
INTRODUCTION
Ovarian cancer is one of the most fatal malignancies among women, with high-grade serous ovarian adenocarcinoma being the leading cause of death[1, 2]. The annual incidence of ovarian cancer varies by geographical area and by age worldwide[3]. The majority of ovarian cancer patients succumb to the disease within 5 years of diagnosis[4]. Approximately 80% of ovarian cancer patients are responsive to treatment with platinum-based drugs. However, most patients will relapse and become resistant to subsequent therapy[5, 6].
BRCA1 and BRCA2 are the key genes involved in the DNA damage response and DNA repair processes, mainly via homologous recombination[7]. BRCA1 and BRCA2 work in concert to protect the genome from double-strand DNA damage during DNA replication. BRCA1 and BRCA2 mutations in ovarian cancers, are associated with defects in homologous recombination and genomic instability. Current researches have demonstrated a trend towards favorable outcomes in patients who are deficient in BRCA1 and BRCA2 compared with patients with wild-type BRCA1 and BRCA2 as a result of the DNA damage induced by platinum-based chemotherapy[8, 9]. BRCA-defects include germline mutations, somatic mutations, hypermethylations and copy number deletions. Unfortunately, BRCA-deficiency most likely occurs in only 30% of ovarian cancer patients[8, 10-12].
MiRNAs are small non-coding RNAs (~ 22 nucleotides) that regulate target gene expression by blocking the translation or degradation of their target mRNAs. Deregulation of miRNAs is involved in the initiation and progression of ovarian cancer[13]. MiRNAs participate in a variety of biological processes, such as the immune response, as well as proliferation and metastasis, which are hallmarks of cancer[14-16]. The aberrant expression of miRNAs in cancers indicates their potential to act as oncogenes or tumor suppressor genes[17]. Additionally, miRNA expression patterns have been associated with the prognosis of ovarian cancer[18, 19]. For example, the increased expression of miR-25 has been closely associated with the poor prognosis of epithelial ovarian cancer[20], indicating that miR-25 may serve as a predictive biomarker for the prognosis of epithelial ovarian cancer. Moreover, miRNAs have been shown to participate in the DNA repair pathway by regulating BRCA1/2 during carcinogenesis. Moskwa P et al. reported that the over-expression of miR-182 inhibited the expression of the BRCA1 protein and affected homologous recombination-mediated repair[21]. Sun C et al. reported that miR-9 mediated the down-regulation of BRCA1 and impeded DNA damage repair in ovarian cancer[22].
In this paper, the genes BRCA1 and BRCA2 are abbreviated as BRCA1/2 for clarity. BRCA1/2-deficient patients subjected to platinum-based treatment have significantly improved survival compared with the patients with wild-type BRCA1/2[9]. However, only a small proportion of ovarian cancers exhibit direct genetic or epigenetic alterations in BRCA1/2. By contrast, the deregulation of miRNAs is an alternative mechanism that may affect the expression of BRCA1/2 and further regulate the DNA damage response and DNA repair pathways. Herein, we hypothesized that ovarian cancer patients with wild-type BRCA1/2 but with miRNA deregulation may also have better prognosis than patients with wild-type BRCA1/2 but no deregulation of miRNA. Our work sought to identify miRNAs that could predict the prognosis of ovarian cancer patients who had no alterations in BRCA1/2 and had initially been treated with a platinum-based chemotherapy regimen. Our hypothesis was that these miRNAs could impede DNA damage repair by reducing BRCA1/2 expression, thereby increasing the sensitivity of cancer cells to chemotherapy. We identified that the up-regulation of three miRNAs (hsa-miR-146a, hsa-miR-148a and hsa-miR-545), which target BRCA1/2, in patients with wild-type BRCA1/2 was associated with good overall survival (OS) and progression-free survival (PFS), a finding that has important implications for the clinical management of ovarian cancers.
RESULTS
Survival differences between patients with and without alterations in BRCA1/2
Here, 317 high-grade serous ovarian adenocarcinomas, together with information on mRNA expression, miRNA expression, mutation, promoter methylation, DNA copy number and patient clinical characteristics, were identified in the TCGA data portal[2] (Table 1). Bolton KL et al. reported that BRCA1 alteration carriers and BRCA2 alteration carriers show similar survival patterns[9]. Thus, we combined the data from patients with somatic or germline mutations, copy number alterations and/or hypermethylations in BRCA1 or BRCA2 into the group “BRCA1/2 alteration carriers”. BRCA1 and BRCA2 were altered in 69 and 33 samples, respectively (Supplementary Table 1). Three samples had both BRCA1 and BRCA2 alterations. The 99 BRCA1/2 alteration carriers had a significantly better OS (P=3.00E-03; 49.5 vs 41.9 months; log-rank test; Supplementary Figure 1) than the 218 patients with wild-type BRCA1/2.
Table 1: Clinical features of ovarian cancer patients.
Characteristic |
TCGA data |
Age (median 59.58, range 27.21-87.47) |
|
<=59.58 |
155 |
>59.58 |
154 |
Stage |
|
II |
15 |
III |
249 |
IV |
52 |
Grade |
|
II |
24 |
III |
286 |
Response Status |
|
CR* |
75 |
Non-CR* |
187 |
Subtype |
|
Proliferative |
89 |
Mesenchymal |
74 |
Differentiated |
92 |
Immunoreactive |
62 |
*CR depicts Complete Response. Non-CR depicts non-complete response, including partial response, stable disease and progressive disease.
Identification of prognostic miRNAs in BRCA1/2 wild-type ovarian cancers
The 218 wild-type BRCA1/2 ovarian cancers were randomly assigned to a training set (n=109) and a testing set (n=109). The training set was used to detect prognostic miRNAs. Fifty-seven miRNAs that are predicted to target the genes BRCA1 or BRCA2 were included in the analysis. According to a univariate Cox proportional hazards regression analysis, 3 of the 57 miRNAs were significantly associated with OS in patients with wild-type BRCA1/2. We calculated a three-miRNA (hsa-miR-146a, hsa-miR-148a and hsa-miR-545) signature risk score for each patient (see Methods). Using the median risk score as the cutoff, patients were classified into a miRNA-related high-risk group and a miRNA-related low-risk group. The patients with low-risk scores were expected to have better survival outcomes. As a result, the miRNA-related low-risk group had longer median OS and PFS than did the miRNA-related high-risk group (P=4.80E-04, median OS= 52.2 vs 26.9 months; Figure 2A).
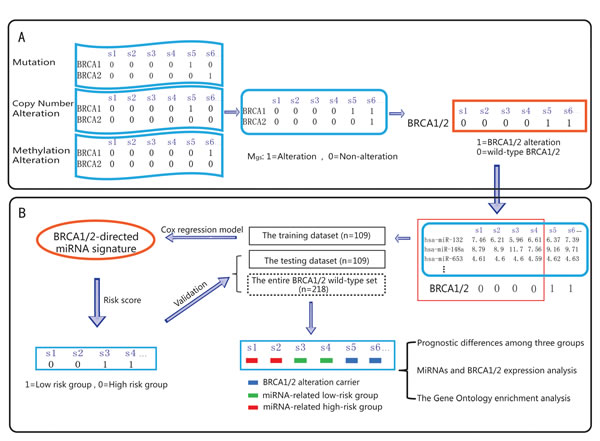
Figure 1: Schematic overview of our analysis procedure. A. By integrating the mutation profile, copy number variation profile and methylation alteration profile, the alteration profile (Mgs) was built: the columns reflect ovarian cancer samples, and the rows reflect genes. If a gene (g) is detected with alterations in a sample (s), Mgs is set to 1; otherwise, Mgs is set to 0. B. The miRNAs that are associated with ovarian cancer prognosis were identified using Cox regression analysis. All of the ovarian cancer samples were divided into three groups: the BRCA 1/2 altered group (BRCA 1/2 alteration carriers), the miRNA-related high-risk group and the miRNA-related low-risk group. Survival difference and differential expression among the groups were then assessed.
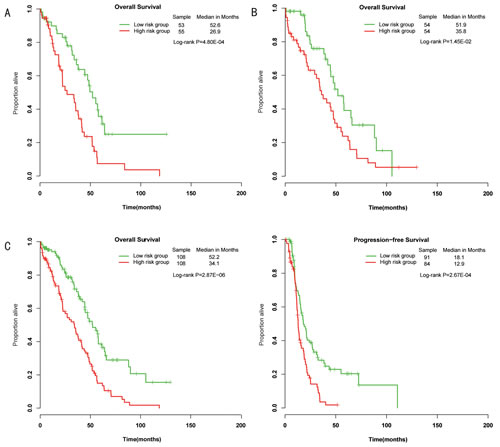
Figure 2: Differences in overall survival and progression-free survival were assessed between the miRNA-related low-risk and high-risk groups. A, The training set. B, The testing set. C, The entire wild-type BRCA1/2 set.
Validation of the BRCA1/2-directed miRNA signature in the testing set and the entire wild-type BRCA1/2 set
To confirm our findings, we validated the BRCA1/2-directed miRNA signature in the testing set. Using the risk score formula, we classified patients in the testing set into a high-risk group (n=54) and a low-risk group (n=54) using the same cutoff as in the training set. Consistent with our findings in the training set, patients in the low-risk group had a significantly longer median overall survival than did those in the high-risk group (P=1.45E-02, median OS= 51.9 vs 35.8 months) (Figure 2B). Combining the training and testing sets, the entire cohort of patients with wild-type BRCA1/2 also yielded similar results (P=2.87E-06, median OS= 52.2 vs 34.1 months; P=2.67E-04, median PFS=18.1 vs 12.9 months; Figure 2C). The distribution of miRNA risk score and miRNA expression is shown in Figure 3. In the entire wild-type BRCA1/2 set, the miRNAs were expressed at significantly higher levels in the low-risk patients than in the high-risk patients (P<0.05).
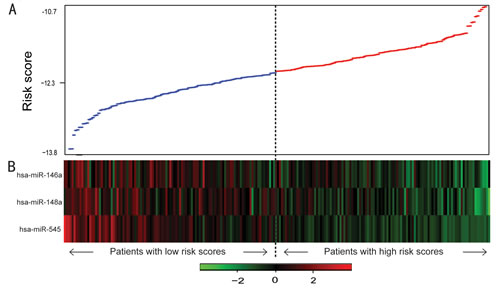
Figure 3: miRNA risk score analysis of ovarian cancers with wild-type BRCA1/2. A, miRNA risk score distribution. B, Heatmap of the miRNA expression profiles. Rows represent miRNAs, and columns represent patients. The black dotted line represents the median risk score cutoff dividing patients into miRNA-related low-risk and high-risk groups.
Independence of prognostic value of the BRCA1/2-directed miRNA signature from other clinical variables
We investigated whether the prognostic value of the BRCA1/2-directed miRNA signature was independent of other clinical variables. The univariate and multivariate Cox analysis consistently revealed that only the BRCA1/2-directed miRNA signature risk score and the treatment response were significantly associated with OS (P<0.05) (Table 2). Next, a data stratification analysis was performed according to treatment response, which stratified the wild-type BRCA1/2 patients into a complete response (CR) group and a non-CR group, which included patients with a partial response, stable disease and progressive disease. The risk score of the three-miRNA signature could further subdivide patients with a CR into groups with significantly different survival times (P=0.01, median OS= 32.0 vs 18.5 months, log-rank test; Supplementary Figure 2). Similarly, even among those patients with a non-CR, the risk score could be used to separate patients into two subgroups with significantly different survival times (P=0.01, median OS= 61.5 vs 48.6 months, log-rank test) (Supplementary Figure 2). These results suggest that the BRCA1/2-directed miRNA signature is an independent prognostic factor for ovarian cancer with wild-type BRCA1/2.
Table 2: Univariate and multivariate Cox regression analysis.
Variables |
Univariate model |
Multivariate model |
||
Hazard Ratio (95% CI) |
P Value |
Hazard Ratio (95% CI) |
P Value |
|
3-miRNA signature risk score |
1.80 (1.34 to 2.42) |
1.04E-04 |
2.04 (1.40 to 2.98) |
2.31E-04 |
Age (<=59.58 vs >59.58) |
1.10 (0.77 to 1.56) |
0.61 |
1.2240 (0.79 to 1.90) |
0.37 |
Stage |
1.29 (0.90 to 1.86) |
0.16 |
1.20 (0.75 to 1.91) |
0.45 |
Grade |
1.89 (0.96 to 3.74) |
0.07 |
2.04 (0.96 to 4.30) |
0.06 |
Treatment response (CR vs non-CR) |
0.27 (0.18 to 0.41) |
8.65E-10 |
0.24 (0.15 to 0.38) |
1.93E-09 |
Subtype |
0.93(0.8 to 1.09) |
0.38 |
0.13 (0.94 to 1.39) |
0.18 |
Prognostic differences among the BRCA1/2 alteration carriers, and miRNA-related high-risk and low-risk patients
The 317 ovarian cancer patients were divided into three groups: the BRCA1/2 alteration carriers group, the miRNA-related high-risk group and the miRNA-related low-risk group. We tested pairwise comparisons of OS and PFS between the groups using the log-rank test. The miRNA-related low-risk group had a significantly better survival than those in the miRNA-related high-risk group (P=2.87E-06, median OS= 52.2 vs 34.1 months; P=2.67E-04, median PFS= 18.1 vs 12.9 months; Figure 4). Additionally, the BRCA1/2 alteration carriers had a significantly longer survival than did patients in the miRNA-related high-risk group (P=8.54E-07, median OS= 49.5 vs 34.1 months; P=6.90E-04, median PFS= 17.8 vs 12.9 months; Figure 4). The BRCA1/2 alteration carriers showed no significant difference in prognosis from patients in the miRNA-related low-risk group (P=0.69, median OS= 49.5 vs 52.2 months; P=0.82, median PFS= 17.8 vs 18.1 months; Figure 4). In other words, patients with wild-type BRCA1/2 and miRNA deregulation also had a better prognosis. These results indicate that patients with either alterations in BRCA1/2 or deregulation in miRNAs targeting BRCA1/2 have a good prognosis and may share a common mechanism, in which miRNAs regulate the DNA repair-related pathway by targeting BRCA1/2.
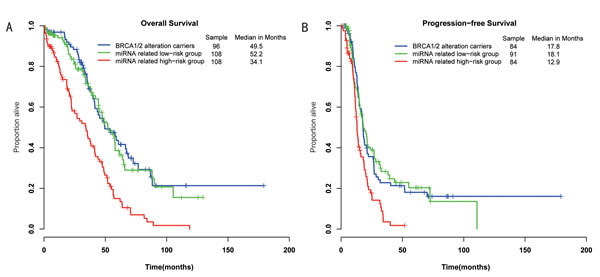
Figure 4: Differences in overall survival and progression-free survival were assessed among the three groups. A, The log-rank P value of overall survival for the miRNA-related low-risk group vs the miRNA-related high-risk group (P=2.87E-06); the BRCA-altered group vs the miRNA-related low-risk group (P=0.69); the BRCA-altered group vs the miRNA-related high-risk group (P=8.54E-07). B, The log-rank P value of progression-free survival for the miRNA-related low-risk group vs the miRNA-related high-risk group (P=2.70E-04); the BRCA-altered group vs the miRNA-related low-risk group (P=0.82); the BRCA-altered group vs the miRNA-related high-risk group (P=6.90E-04).
MiRNAs and BRCA1/2 expression in ovarian cancers
According to the miRNA-target regulation data from the databases (see Methods), hsa-miR-146a is predicted to target BRCA1 and BRCA2; hsa-miR-545 is predicted to target BRCA1; and hsa-miR-148a is predicted to target BRCA2 (Supplementary Figure 3). We found that the three miRNAs were significantly up-regulated in the miRNA-related low-risk group compared with the miRNA-related high-risk group, the BRCA1/2 alteration carriers, and the normal samples (P<0.01) (Figure 5). There were no significant differences in the expression values of the three miRNAs between the miRNA-related high-risk group and the normal samples (P=0.39 for hsa-miR-545, P=0.17 for hsa-miR-148a and P=0.25 for hsa-miR-146a). Additionally, there were no significant differences in the expression of these miRNAs between the BRCA1/2 alteration carriers and the normal samples (P=0.076 for hsa-miR-545, P=0.065 for hsa-miR-148a and P=0.022 for hsa-miR-146a). Detailed results are shown in Supplementary Table 2. Those results suggested that the three miRNAs were deregulated in a portion of the wild-type BRCA1/2 cases and that this deregulation might have facilitated the good prognosis of these ovarian cancers.
Based on the above analysis, we combined the BRCA1/2 alteration carriers and miRNA-related low-risk patients into a good prognosis group, and the miRNA-related high-risk patients were defined as the poor prognosis group. We found that although the genes BRCA1 and BRCA2 trended toward being down-regulated in the miRNA-related low-risk group compared with the poor prognosis group, this fell short of significance. Nevertheless, the expression values of BRCA1 and BRCA2 were significantly lower in the good prognosis group than in the poor prognosis group (P=1.90E-05 for BRCA1 and P=1.60E-02 for BRCA2). These results indicate that the three miRNAs may facilitate a good prognosis for ovarian cancer by down-regulating BRCA1/2 and deregulating the DNA repair-related pathways.

Figure 5: The expression of three BRCA1/2-directed miRNAs in ovarian cancer cases and normal cases. BRCA-altered refers to the group of BRCA1/2 alteration carriers. miRNA low-risk and miRNA high-risk refer to the patients with wild-type BRCA1/2 who are predicted to fall into the high-risk and low-risk groups, respectively, according to the BRCA1/2-directed miRNA signature. ** means P<0.01.
Enrichment analysis of DNA damage response pathways
Deregulation of BRCA1/2-directed miRNA and alteration of BRCA1/2 may share a common mechanism that affects the prognosis of ovarian cancer patients treated with platinum-based chemotherapy. Based on this hypothesis, we first detected differentially expressed genes (DEGs) between the miRNA-related low-risk group and the miRNA-related high-risk group (miRNA-DEGs) and between the BRCA1/2 alteration carrier group and the miRNA-related high-risk group (BRCA-DEGs), respectively. We found that 98% of overlapping genes between the two DEG lists (miRNA-DEGs and BRCA-DEGs) were highly consistent in their deregulation directions (up-regulation or down-regulation). Next, using functional enrichment analysis, we further investigated whether the DEGs participated in the DNA repair-related biological processes derived from the GO database. The results revealed that the genes in the two DEG lists were significantly enriched in the DNA repair-related pathways (Supplementary Table 3) (P<0.05). The pathways most enriched in BRCA-DEGs were “DNA damage response, signal transduction by p53 class mediator resulting in cell cycle arrest” (GO: 0006977, P=8.74E-05) and “signal transduction involved in mitotic G1 DNA damage checkpoint” (GO: 0072431, P=8.74E-05). The pathway most enriched in the miRNA-DEGs was “DNA damage response, signal transduction by p53 class mediator” (GO: 0030330, P=2.30E-03). Some pathways, such as “signal transduction in response to DNA damage” (GO: 0042770) and “DNA damage response, signal transduction by p53 class mediator” (GO: 0030330), were enriched in both the BRCA-DEGs and miRNA-DEGs. These results suggested that similar to BRCA1/2 alterations, the deregulation of miRNAs affected the prognosis of ovarian cancers by regulating DNA damage response-related pathways.
DISCUSSION
BRCA1 and BRCA2 function at different stages of the DNA damage response and DNA repair but act in concert to protect the genome from double-stranded DNA damage during DNA replication[7]. Our results confirmed the findings that 30% of ovarian cancer patients carrying alterations in BRCA1/2 had a better prognosis than BRCA1/2 alteration non-carriers. However, a proportion of patients with wild-type BRCA1/2 may also have favorable prognosis as a result of other mechanisms, such as the deregulation of miRNAs. In this study, an analysis of 218 high-grade serous ovarian cancer cases with wild-type BRCA1/2 revealed a BRCA1/2-directed miRNA signature model. The deregulation of the three miRNAs (hsa-miR-545, hsa-miR-146a and hsa-miR-148a) was significantly associated with favorable OS and PFS in wild-type BRCA1/2 ovarian cancer patients. By analyzing the expression profiles and GO functional enrichment, we unraveled that carriers of BRCA1/2 alterations and patients with miRNA deregulation shared a common mechanism that affected the prognosis of ovarian cancer treated with platinum-based chemotherapy. In other words, these miRNAs also participated in the DNA damage response and repair-related pathways by regulating BRCA1/2. In summary, our work identified that some BRCA1/2 alteration non-carriers benefit from platinum-based chemotherapy, a finding that has potentially important implications for the clinical management of patients with ovarian cancer.
Kang et al. showed that some genes that are differentially expressed between ovarian cancer patients with poor and favorable outcomes are involved in the repair of platinum-induced DNA damage and extracted 151 DNA repair-related genes from the literature[23] . The three miRNAs (hsa-miR-545, hsa-miR-146a and hsa-miR-148a) that we identified targeted 29 of these 151 DNA repair genes, and their regulatory relationships were constructed as a network (Supplementary Figure 3). Six of the network genes were significantly down-regulated in the miRNA-related low-risk group compared with the miRNA-related high-risk group. These results indicate that the three miRNAs target not only BRCA1 or BRCA2 but also multiple DNA repair pathway genes to response platinum-based chemotherapy.
Some studies have reported that miRNAs can impact sensitivity to cancer therapy by targeting BRCA1/2. Moskwa et al. found that the miR-182-mediated down-regulation of BRCA1 affected DNA repair and sensitivity to inhibitors of poly (ADP-ribose) polymerase 1 in breast cancer cell lines[21]. miR-9 down-regulated BRCA1 and impeded DNA damage repair in ovarian cancer cells, which could improve chemotherapeutic efficacy[22]. Some previous studies have confirmed the relationships between hsa-miR-146a and BRCA1 and between hsa-miR-545 and BRCA1[24, 25]. Our future work will focus on validating the miRNA-target relationships among hsa-miR-146a, hsa-miR-148a and BRCA2 and exploring the effect of deregulation of miRNAs on sensitivity to platinum-based chemotherapy using wet experiments on cell lines and clinical samples. Despite the functional evidence that we have presented, one limitation of our study is the lack of an independent multidimensional dataset to validate our conclusion. Nevertheless, the discovery that miRNAs represent another mechanism that affects the prognosis of ovarian cancer patients with wild-type BRCA1/2 may have important implications for clinical prediction and trial design. Moreover, long non-coding RNAs (lncRNAs), which are non-protein-coding transcripts ranging from 200 nucleotides (nt) to ~100 kilobases (kb) in length[26], are frequently aberrantly expressed in cancers and can be used for cancer diagnosis and prognosis[27, 28]. However, the roles of lncRNAs in regulating BRCA1/2 and their impact on the prognosis of ovarian cancer have yet to be elucidated, and these warrant detailed research in the future.
MATERIALS AND METHODS
Datasets and processing
317 high-grade serous ovarian adenocarcinomas and 8 normal fallopian tube samples, including all information on mRNA expression, miRNA expression, mutation, promoter methylation, DNA copy number and clinical features, were downloaded from the TCGA data portal[2]. The level 3 mutation profile (somatic mutation and germline mutation information) was used. The level 1 copy number data were processed using the dChip software, the GLAD algorithm and the GISTIC algorithm, as described by Gu et al[29]. Level 3 IlluminaInfinium DNA methylation data were processed as described by Yang et al[30]. By integrating the discrete mutation profile, copy number alteration profile and methylation profile, we obtained the gene alteration profile (Mgs), in which the columns reflect ovarian cancer samples and the rows reflect genes. If a gene (g) was detected with alterations in a sample (s), we set Mgs to 1; otherwise, Mgs was set to 0 (Figure 1A). Here, alterations in the Mgs included somatic mutations, germline mutations, copy number amplifications/deletions and hypo/hyper-methylations. All patients received platinum-based chemotherapy after surgery. The detailed clinical features of the patients are listed in Table 1.
Considering that miRNA targets predicted by multiple algorithms might be more reliable, we used miRNA-target interactions appearing in at least two of nine databases: TargetScan[31], miRanda[32], PicTar[33], miRBase[34], DIANA-microT[35], PITA[36], miRNAMap[37], miRTarBase[38] and miRecords[39].
Identification of predictive BRCA1/2-directed miRNA signature
In total, germline and somatic mutations, hypermethylations or deletions in the genes BRCA1 or BRCA2 (Supplementary Table 1) were detected in 99 patients. The 218 samples with wild-type BRCA1/2 were randomly divided into a training set (n=109 samples) and a testing set (n=109 samples). First, in the training set, the association between the miRNAs targeting BRCA1/2 and survival was assessed by univariate Cox regression analysis (Figure 1B). Second, a miRNA prediction classifier was constructed by linear combination of the expression values of the BRCA1/2-directed miRNAs and the Cox regression coefficient as the weight. The patients were classified into high-risk and low-risk groups by utilizing the median risk score as the cutoff point (low-risk group was 1, high-risk group was 0). Then, the log-rank test was used to assess the overall survival between the low-risk and high-risk groups. In the Cox analysis, miRNAs with P<0.05 were selected as signature that were significantly associated with the OS of ovarian cancer. Finally, we validated the miRNA risk-prediction model in the testing set. The 317 ovarian cancer samples from the TCGA data portal were divided into three groups: the BRCA1/2 alteration carrier group, the miRNA related high-risk group and the miRNA related low-risk group. Kaplan-Meier survival plots and the log-rank test were used to assess the differences in OS and PFS among the three groups. Multivariate Cox analysis was applied to test whether the BRCA1/2-directed miRNA signature was independent of other clinical characteristics such as age, tumor grade and so on.
Statistical analysis
A two-sample t-test was performed to identify the differentially expressed miRNAs or mRNAs in the level 3 miRNA expression profile and level 3 mRNA expression profile, respectively. The hypergeometric distribution model was used to test whether the DNA repair-related terms from the Gene Ontology (GO) database[40] were significantly enriched in the differentially expressed genes[41].
Conflict of Interests
The authors declare that they have no conflict of interests.
Abbreviations
CR, Complete Response; DEG, Differential Expression Gene; HR, Hazard Ratio; OS, Overall Survival; PFS, Progression-Free Survival; GO: Gene Ontology.
ACKNOWLEDGEMENTS
This work was supported by the National Natural Science Foundation of China (Grant No. 81201822, 31300943 and 81372213), Research Fund for the Doctoral Program of Higher Education of China (Grant No. 20112307110011), Postdoctoral Science Foundation of China (Grant No. 2012M511509) and Postdoctoral Foundation of Heilongjiang Province (Grant No. LBH-Z11083).
REFERENCES
1. Kim J, Coffey DM, Creighton CJ, Yu Z, Hawkins SM and Matzuk MM. High-grade serous ovarian cancer arises from fallopian tube in a mouse model. Proc Natl Acad Sci USA. 2012; 109(10):3921-3926.
2. Integrated genomic analyses of ovarian carcinoma. Nature. 2011; 474(7353):609-615.
3. Jemal A, Siegel R, Xu J and Ward E. Cancer statistics, 2010. CA Cancer J Clin. 2010; 60(5):277-300.
4. Ow GS, Ivshina AV, Fuentes G and Kuznetsov VA. Identification of two poorly prognosed ovarian carcinoma subtypes associated with CHEK2 germ-line mutation and non-CHEK2 somatic mutation gene signatures. Cell Cycle. 2014; 13(14):2262-2280.
5. Cooke SL and Brenton JD. Evolution of platinum resistance in high-grade serous ovarian cancer. Lancet Oncol. 2011; 12(12):1169-1174.
6. Ledermann J, Harter P, Gourley C, Friedlander M, Vergote I, Rustin G, Scott C, Meier W, Shapira-Frommer R, Safra T, Matei D, Macpherson E, Watkins C, et al. Olaparib maintenance therapy in platinum-sensitive relapsed ovarian cancer. N Engl J Med. 2012; 366(15):1382-1392.
7. Roy R, Chun J and Powell SN. BRCA1 and BRCA2: different roles in a common pathway of genome protection. Nat Rev Cancer. 2012; 12(1):68-78.
8. Ruscito I, Dimitrova D, Vasconcelos I, Gellhaus K, Schwachula T, Bellati F, Zeillinger R, Benedetti-Panici P, Vergote I, Mahner S, Cacsire-Tong D, Concin N, Darb-Esfahani S, et al. BRCA1 gene promoter methylation status in high-grade serous ovarian cancer patients--a study of the tumour Bank ovarian cancer (TOC) and ovarian cancer diagnosis consortium (OVCAD). Eur J Cancer. 2014; 50(12):2090-2098.
9. Bolton KL, Chenevix-Trench G, Goh C, Sadetzki S, Ramus SJ, Karlan BY, Lambrechts D, Despierre E, Barrowdale D, McGuffog L, Healey S, Easton DF, Sinilnikova O, et al. Association between BRCA1 and BRCA2 mutations and survival in women with invasive epithelial ovarian cancer. JAMA. 2012; 307(4):382-390.
10. Soegaard M, Kjaer SK, Cox M, Wozniak E, Hogdall E, Hogdall C, Blaakaer J, Jacobs IJ, Gayther SA and Ramus SJ. BRCA1 and BRCA2 mutation prevalence and clinical characteristics of a population-based series of ovarian cancer cases from Denmark. Clin Cancer Res. 2008; 14(12):3761-3767.
11. McLaughlin JR, Rosen B, Moody J, Pal T, Fan I, Shaw PA, Risch HA, Sellers TA, Sun P and Narod SA. Long-term ovarian cancer survival associated with mutation in BRCA1 or BRCA2. J Natl Cancer Inst. 2013; 105(2):141-148.
12. Swisher EM, Gonzalez RM, Taniguchi T, Garcia RL, Walsh T, Goff BA and Welcsh P. Methylation and protein expression of DNA repair genes: association with chemotherapy exposure and survival in sporadic ovarian and peritoneal carcinomas. Mol Cancer. 2009; 8:48.
13. Vaksman O, Stavnes HT, Kaern J, Trope CG, Davidson B and Reich R. miRNA profiling along tumour progression in ovarian carcinoma. J Cell Mol Med. 2011; 15(7):1593-1602.
14. Montagner S, Orlandi EM, Merante S and Monticelli S. The role of miRNAs in mast cells and other innate immune cells. Immunol Rev. 2013; 253(1):12-24.
15. Guan Y, Yao H, Zheng Z, Qiu G and Sun K. MiR-125b targets BCL3 and suppresses ovarian cancer proliferation. Int J Cancer. 2011; 128(10):2274-2283.
16. Liu C, Kelnar K, Liu B, Chen X, Calhoun-Davis T, Li H, Patrawala L, Yan H, Jeter C, Honorio S, Wiggins JF, Bader AG, Fagin R, et al. The microRNA miR-34a inhibits prostate cancer stem cells and metastasis by directly repressing CD44. Nat Med. 2011; 17(2):211-215.
17. Garofalo M and Croce CM. microRNAs: Master regulators as potential therapeutics in cancer. Annu Rev Pharmacol Toxicol. 2011; 51:25-43.
18. Marchini S, Cavalieri D, Fruscio R, Calura E, Garavaglia D, Fuso Nerini I, Mangioni C, Cattoretti G, Clivio L, Beltrame L, Katsaros D, Scarampi L, Menato G, et al. Association between miR-200c and the survival of patients with stage I epithelial ovarian cancer: a retrospective study of two independent tumour tissue collections. Lancet Oncol. 2011; 12(3):273-285.
19. Leskela S, Leandro-Garcia LJ, Mendiola M, Barriuso J, Inglada-Perez L, Munoz I, Martinez-Delgado B, Redondo A, de Santiago J, Robledo M, Hardisson D and Rodriguez-Antona C. The miR-200 family controls beta-tubulin III expression and is associated with paclitaxel-based treatment response and progression-free survival in ovarian cancer patients. Endocr Relat Cancer. 2011; 18(1):85-95.
20. Wang X, Meng X, Li H, Liu W, Shen S and Gao Z. MicroRNA-25 expression level is an independent prognostic factor in epithelial ovarian cancer. Clin Transl Oncol. 2014; 16(11):954-958.
21. Moskwa P, Buffa FM, Pan Y, Panchakshari R, Gottipati P, Muschel RJ, Beech J, Kulshrestha R, Abdelmohsen K, Weinstock DM, Gorospe M, Harris AL, Helleday T, et al. miR-182-mediated downregulation of BRCA1 impacts DNA repair and sensitivity to PARP inhibitors. Mol Cell. 2011; 41(2):210-220.
22. Sun C, Li N, Yang Z, Zhou B, He Y, Weng D, Fang Y, Wu P, Chen P, Yang X, Ma D, Zhou J and Chen G. miR-9 regulation of BRCA1 and ovarian cancer sensitivity to cisplatin and PARP inhibition. J Natl Cancer Inst. 2013; 105(22):1750-1758.
23. Kang J, D’Andrea AD and Kozono D. A DNA repair pathway-focused score for prediction of outcomes in ovarian cancer treated with platinum-based chemotherapy. J Natl Cancer Inst. 2012; 104(9):670-681.
24. Garcia AI, Buisson M, Bertrand P, Rimokh R, Rouleau E, Lopez BS, Lidereau R, Mikaelian I and Mazoyer S. Down-regulation of BRCA1 expression by miR-146a and miR-146b-5p in triple negative sporadic breast cancers. EMBO Mol Med. 2011; 3(5):279-290.
25. Dimitrov SD, Lu D, Naetar N, Hu Y, Pathania S, Kanellopoulou C and Livingston DM. Physiological modulation of endogenous BRCA1 p220 abundance suppresses DNA damage during the cell cycle. Genes Dev. 2013; 27(20):2274-2291.
26. Mercer TR, Dinger ME and Mattick JS. Long non-coding RNAs: insights into functions. Nat Rev Genet. 2009; 10(3):155-159.
27. Qi P and Du X. The long non-coding RNAs, a new cancer diagnostic and therapeutic gold mine. Mod Pathol. 2013; 26(2):155-165.
28. Zhang XQ, Sun S, Lam KF, Kiang KM, Pu JK, Ho AS, Lui WM, Fung CF, Wong TS and Leung GK. A long non-coding RNA signature in glioblastoma multiforme predicts survival. Neurobiol Dis. 2013; 58:123-131.
29. Gu Y, Wang H, Qin Y, Zhang Y, Zhao W, Qi L, Wang C and Guo Z. Network analysis of genomic alteration profiles reveals co-altered functional modules and driver genes for glioblastoma. Mol Biosyst. 2013; 9(3):467-477.
30. Yang D, Khan S, Sun Y, Hess K, Shmulevich I, Sood AK and Zhang W. Association of BRCA1 and BRCA2 mutations with survival, chemotherapy sensitivity, and gene mutator phenotype in patients with ovarian cancer. JAMA. 2011; 306(14):1557-1565.
31. Lewis BP, Burge CB and Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005; 120(1):15-20.
32. Betel D, Koppal A, Agius P, Sander C and Leslie C. Comprehensive modeling of microRNA targets predicts functional non-conserved and non-canonical sites. Genome Biol. 2010; 11(8):R90.
33. Krek A, Grun D, Poy MN, Wolf R, Rosenberg L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M and Rajewsky N. Combinatorial microRNA target predictions. Nat Genet. 2005; 37(5):495-500.
34. Kozomara A and Griffiths-Jones S. miRBase: integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011; 39(Database issue):D152-157.
35. Paraskevopoulou MD, Georgakilas G, Kostoulas N, Vlachos IS, Vergoulis T, Reczko M, Filippidis C, Dalamagas T and Hatzigeorgiou AG. DIANA-microT web server v5.0: service integration into miRNA functional analysis workflows. Nucleic Acids Res. 2013; 41(Web Server issue):W169-173.
36. Kertesz M, Iovino N, Unnerstall U, Gaul U and Segal E. The role of site accessibility in microRNA target recognition. Nat Genet. 2007; 39(10):1278-1284.
37. Hsu SD, Chu CH, Tsou AP, Chen SJ, Chen HC, Hsu PW, Wong YH, Chen YH, Chen GH and Huang HD. miRNAMap 2.0: genomic maps of microRNAs in metazoan genomes. Nucleic Acids Res. 2008; 36(Database issue):D165-169.
38. Hsu SD, Lin FM, Wu WY, Liang C, Huang WC, Chan WL, Tsai WT, Chen GZ, Lee CJ, Chiu CM, Chien CH, Wu MC, Huang CY, et al. miRTarBase: a database curates experimentally validated microRNA-target interactions. Nucleic Acids Res. 2011; 39(Database issue):D163-169.
39. Xiao F, Zuo Z, Cai G, Kang S, Gao X and Li T. miRecords: an integrated resource for microRNA-target interactions. Nucleic Acids Res. 2009; 37(Database issue):D105-110.
40. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, et al. Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet. 2000; 25(1):25-29.
41. Hong G, Zhang W, Li H, Shen X and Guo Z. Separate enrichment analysis of pathways for up- and downregulated genes. J R Soc Interface. 2014; 11(92):20130950.