
INTRODUCTION
About 72,500 cases of newly diagnosed bladder cancer and 17,960 bladder cancer deaths are estimated to occur in the United States in 2013 [1]. Urothelial carcinomas comprise around 90% of bladder cancers. However, the carcinogenesis of urothelial carcinoma is poorly perceived [2].
The complex phenotypes of bladder cancer cells are controlled by functional response genes [3]. MicroRNAs are small non-coding RNAs that mainly inhibit gene expression [4]. They are increased or decreased in cancers and able to promote or suppress cancer development [5]. Hsa-miR-429 is increased in cancers, such as endometrial carcinoma [6] and colorectal cancer [7]. In contrast, some other findings suggest that hsa-miR-429 functions as a tumor suppressor in osteosarcoma [8], renal cell carcinoma [9] and cervical cancer [10]. We have validated that hsa-miR-429 is indeed increased in bladder cancer [11], while it has been found that hsa-miR-429 reverses epithelial-mesenchymal transition by restoring E-cadherin expression in bladder cancer [12]. Hsa-miR-429 belongs to miR-200 family, and the upregulation of miR200 family has also been reported in bladder cancer [13]. For example, there should be a positive relationship between high microRNA-200C expression and the risk of death from disease in muscle-invasive urothelial carcinoma of the bladder, as revealed by one published work [14].
Partial sequence pairing between miRNA and target mRNA leads to translation inhibition and/or mRNA degradation in animals [15]. We have identified that hsa-miR-125b decreased its target SIRT7 at both mRNA and protein levels through partial sequence pairing with the target sites [16]. Cyclin-dependent kinase inhibitor 2B (CDKN2B) is a tumor suppressor [17]. Deletion, mutation and hypermethylation of CDKN2B gene have been reported to lead to the loss of CDKN2B expression [18]. There is a negative regulatory role of miR-15a-5p in the apoptosis of smooth muscle cells via binding to the CDKN2B mRNA 3’UTR [19]. In silico analysis indicates that CDKN2B 3’UTR has binding site for hsa-miR-429. This finding proposed an important question about the relationship between hsa-miR-429 and CDKN2B in bladder cancer. We hypothesized that hsa-miR-429 may play oncogenic roles via inhibiting CDKN2B in bladder cancer.
To test the proposed hypothesis, in this study, the expression patterns of hsa-miR-429 and CDKN2B in urothelial carcinoma of the bladder were determined. Hsa-miR-429 mimic decreased CDKN2B at both mRNA and protein levels. The binding site between hsa-miR-429 and CDKN2B 3’UTR was confirmed. We found that hsa-miR-429 promoted cell growth and decreased apoptosis via inhibiting CDKN2B.
RESULTS
Hsa-miR-429 and CDKN2B were inversely expressed in bladder cancer
The relative expression levels of Hsa-miR-429 and CDKN2B were evaluated using Real-Time qPCR in 50 patients with bladder cancer. Hsa-miR-429 was increased in bladder cancer compared with matched normal urothelium. As shown in Table 1, upregulation of hsa-miR-429 was positively correlated with bladder cancer clinical pathologic grading (p<0.001) and TNM stage (P=0.005), while gender, age and lymph node metastasis had no associations with hsa-miR-429 expression level. High expression level of miR-429 in the tissue sample appears to be associated with low survival in patients with bladder cancer (Figure 1). For patients with low hsa-miR-429 expression, all survived 5 years after surgery, while for those with high hsa-miR-429 expression, only 58% survived. CDKN2B was decreased in bladder cancer compared with matched normal urothelium. Hsa-miR-429 and CDKN2B were inversely expressed in bladder cancer. The data are provided in Table 2.
Table 1: Correlation between hsa-miR-429 expression and clinicopathological characteristics of bladder cancer patients
Characteristics |
Expression of has-miR-429 |
P value |
|
|---|---|---|---|
High (n=40) |
Low (n=10) |
||
Gender |
|||
Male |
30(78.9%) |
8(21.1%) |
0.575 |
Female |
10 (83.3%) |
2(16.7%) |
|
Age |
|||
<=60 |
31(81.5%) |
7(18.5%) |
0.587 |
>60 |
9(75.0%) |
3 (25.0%) |
|
Histological grade* |
|||
PUNLMP/Low-grade |
7(46.7%) |
8(53.3%) |
P<0.001* |
High-grade |
33(94.2%) |
2(5.8%) |
|
Lymph node metastasis (N)** |
|||
N0 |
35(79.5%) |
9(20.5%) |
0.663 |
N1 or above |
5(83.3%) |
1 (16.7%) |
|
TNM stage** |
|||
0/I |
10(55.5%) |
8(44.5%) |
0.005* |
II/III/IV |
30(93.7%) |
2(6.3%) |
|
*PUNLMP, papillary urothelial neoplasm of low malignant potential; low-grade, low-grade papillary urothelial carcinoma; high-grade, high-grade papillary urothelial carcinoma (http://www.pathology.jhu.edu/bladder).
**TNM according to the seventh edition of staging TNM of Union Internationale Contre Le Cancer (UICC) in 2009.
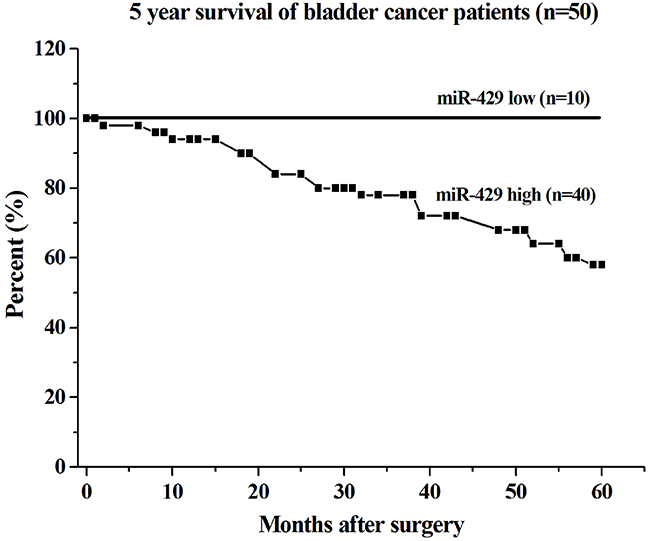
Figure 1: Correlation of hsa-miR-429 expression with 5 year survival of bladder cancer patients. The survival rate of the group with low expression of miR-429 was lower than that of the group with high expression of miR-429 (log-rank test, p < 0.01).
Table 2: The expression of hsa-miR-429 and CDKN2B in bladder cancer
Tumor (n=50) |
Normal (n=50) |
P value |
Pearson’s coefficient correlation* |
||
|---|---|---|---|---|---|
hsa-miR-429 expression |
0.49±0.07 |
0.12±0.03 |
0.019 |
R=−0.602 |
P=0.031 |
CDKN2B expression# |
0.29±0.08 |
0.79±0.15 |
0.014 |
||
#The relative expression levels were evaluated using Real-Time qPCR. Data are shown as mean±SEM.
*log2-Fold-Change (tumor/normal) of hsa-miR-429/CDKN2B in 50 bladder cancer patients.
Hsa-miR-429 decreased the expression of CDKN2B
Forty-eight hours after transfection, CDKN2B expression levels were measured by qRT-PCR and western blot analysis. Both CDKN2B siRNA and hsa-miR-429 mimic decreased CDKN2B at mRNA and protein levels in T24 and 5637 cells (Figure 2A and 2B).
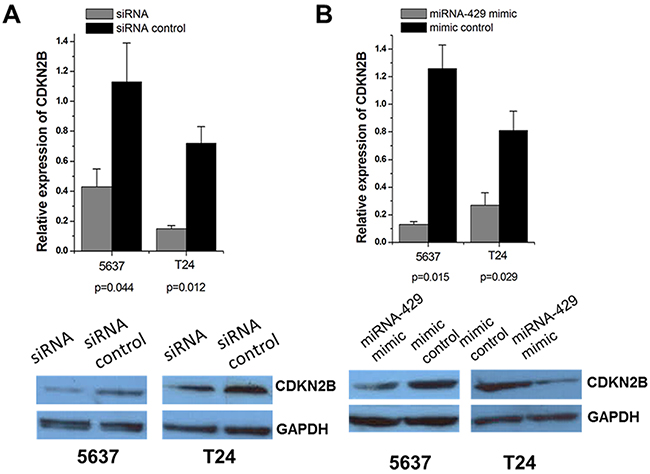
Figure 2: Expression changes of CDKN2B after transfection of CDKN2B siRNA and hsa-miR-429 mimic in T24 and 5637 cells. The relative mRNA expression levels were evaluated using Real-Time qPCR. Data are indicated as mean±SD. CDKN2B protein levels were determined using Western blot assay. Each experiment was performed in triplicate for three independent times. (A) CDKN2B specific siRNA significantly decreased the expression of CDKN2B at mRNA levels. Representative images of Western blot results indicated CDKN2B specific siRNA significantly decreased the expression of CDKN2B at protein levels. (B) Hsa-miR-429 mimic significantly decreased the expression of CDKN2B at mRNA levels. Representative images of Western blot results indicated that hsa-miR-429 mimic significantly decreased the expression of CDKN2B at protein levels.
Hsa-miR-429 reduced the luciferase activities
Forty-eight hours after transfection, the dual luciferase reporter assay was performed. Hsa-miR-429 inhibited the luciferase activities in T24 and 5637 cells transfected with the reporter vector CDKN2B 3’UTR-WT, but not in cells transfected with the reporter vector CDKN2B 3’UTR-MUT (Figure 3A and 3B), which indicated the hsa-miR-429 binding site within CDKN2B 3’UTR was functional.
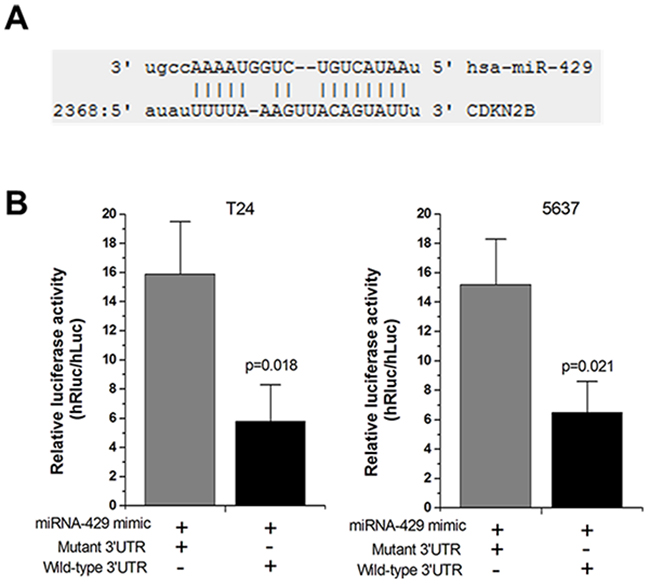
Figure 3: Hsa-miR-429 mimic reduced the luciferase activities. (A) The predicted hsa-miR-429 binding site located in CDKN2B 3’UTR. (B) The relative luciferase activities were inhibited in the cells transfected with the reporter vector CDKN2B 3’UTR-WT, not in the cells transfected with the reporter vector CDKN2B 3’UTR-MUT. Data are shown as mean ± SD. Each experiment was performed in triplicate for three independent times.
Cell proliferation and apoptosis changes caused by transfection of CDKN2B siRNA/hsa-miR-429 mimic
Forty-eight hours after transfection, cell proliferation was determined using MTT assay. Cell proliferation promotion was observed in T24 and 5637 cells by transfection of hsa-miR-429 mimic (Figure 4A and 4B) or CDKN2B siRNA (Figure 4C and 4D). EdU incorporation assay was also used as a further study to determine the effects of CDKN2B siRNA/hsa-miR-429 mimic on proliferation of bladder cancer cell lines. The results showed that CDKN2B siRNA/hsa-miR-429 could promote the cell growth of T24 (Figure 5A and 5B) and 5637 (Figure 5C and 5D) remarkably. As revealed by ELISA assay, decreased cell apoptosis was observed in both bladder cancer cells by transfection of hsa-miR-429 mimic (Figure 6A) or CDKN2B siRNA (Figure 6B and 6C).
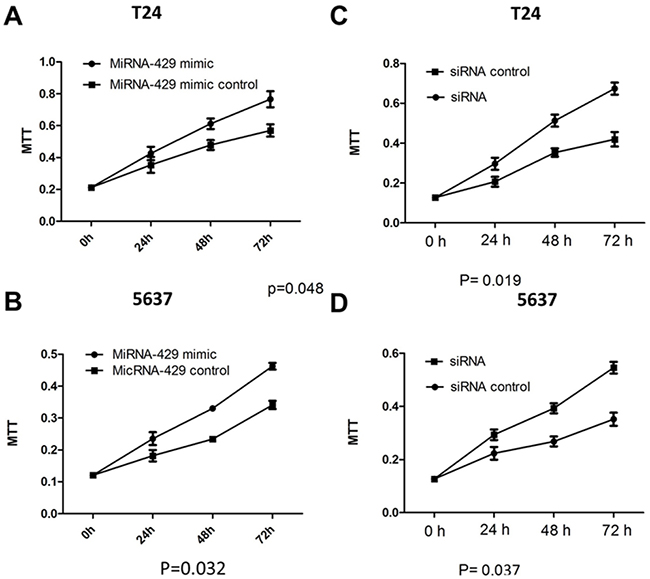
Figure 4: Cell proliferation changes caused by transfection of CDKN2B siRNA and hsa-miR-429 mimic Cell proliferation was measured by MTT assay. (A) and (B) Hsa-miR-429 mimic promoted T24 and 5637 proliferation. (C) and (D) CDKN2B siRNA promoted T24 and 5637 proliferation. Data are indicated as mean ± SD. Each experiment in both cell lines was performed in triplicate for three independent times.
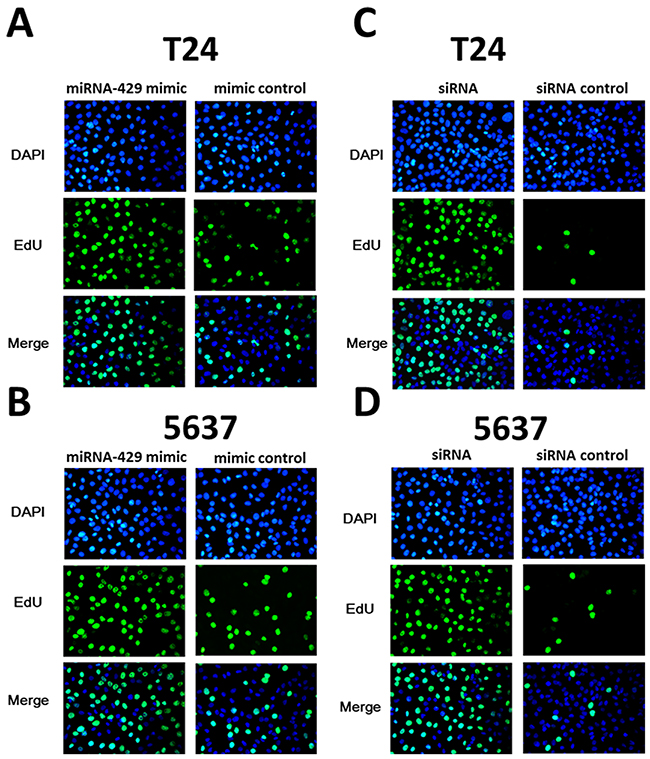
Figure 5: The effects of CDKN2B siRNA /hsa-miR-429 mimic on cell proliferation determined by EdU assay. (A) and (B) Hsa-miR-429 mimic promoted T24 and 5637 proliferation. (C) and (D) CDKN2B siRNA promoted T24 and 5637 proliferation.
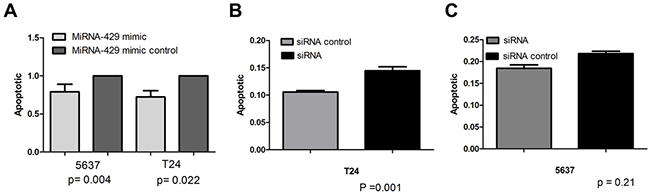
Figure 6: Cell apoptosis changes caused by transfection of CDKN2B siRNA and hsa-miR-429 mimic forty-eight hours after transfection, cell apoptosis changes were determined by Caspase 3 ELISA. (A) Hsa-miR-429 mimic inhibited T24 and 5637 apoptosis. (B) and (C) CDKN2B siRNA inhibited T24 and 5637 apoptosis. Data are indicated as mean ± SD. Each experiment in both cell lines was performed in triplicate for three independent times.
Overexpression of hsa-miR-429 promoted in vivo tumor growth
To investigate the possible impacts of hsa-miR-429 on in vivo growth of bladder cancer cells, the cell growth changes of T24 (Figure 7A) and 5637 (Figure 7B) cells were determined by tumourigenicity assay. Significant differences were demonstrated between the miR-429 expression vector and empty vector-transfected cells. Has-miR-429 was able to promote tumor development, compared to empty control vector treatment. These results further confirmed the positive role of has-miR-429 in the growth of bladder cancer.
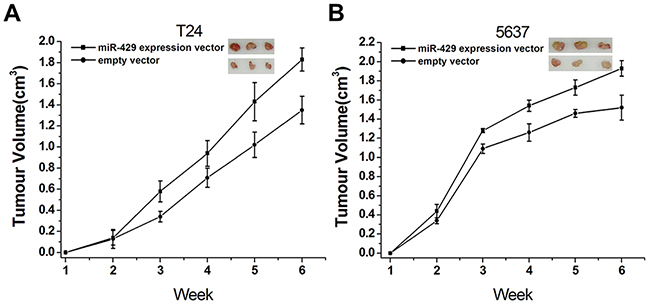
Figure 7: Involvement of hsa-miR-429 in vivo cell growth. (A) Overexpression of miR-429 inhibited T24 cell growth in vivo (p < 0. 01). (B) Overexpression of miR-429 inhibited 5637 cell growth in vivo (p < 0.01). Data are shown as mean ± SD.
DISCUSSION
In this study, we found that the expression levels of hsa-miR-429 were increased in bladder cancer. Hsa-miR-429 mimic promoted bladder cancer cell proliferation both in vitro and in vivo and decreased apoptosis. These findings suggested that hsa-miR-429 is an oncogene in bladder cancer.
The data also reveal that the expression levels of CDKN2B were decreased in bladder cancer. Down-regulation of CDKN2B promoted bladder cancer cell proliferation and decreased apoptosis. These effects were similar to that of transfection of hsa-miR-429 mimic in bladder cancer cells.
CDKN2B was an in silico predictable target of hsa-miR-429. The mature hsa-miR-429 sequence was partially complementary with CDKN2B 3’UTR target sequence. In consistent with the prediction, we showed that hsa-miR-429 and CDKN2B were inversely expressed in bladder cancer. Hsa-miR-429 mimic decreased the expression of CDKN2B at both mRNA and protein levels. Binding site was confirmed between hsa-miR-429 and CDKN2B. These findings suggested that CDKN2B is a real target of hsa-miR-429. In this study, we identified that overexpression of hsa-miR-429 is a novel contributor to the down-regulation of CDKN2B in bladder cancer, suggesting that post-transcriptional inhibition plays roles in repressing CDKN2B expression.
The exact role of hsa-miR-429 in bladder cancer is still controversial. Hsa-miR-429 is considered to have a tumor suppressive role based on the previous report in which hsa-miR-429 was found to reverse epithelial-mesenchymal transition in bladder cancer. However, in our works, hsa-miR-429 seems to be a promoter of proliferation in bladder cancer cells. Interestingly, it has also been concluded that TGFβ mediates downregulation of hsa-miR-429 [20] and that TGFβ decreases cell viability and induces epithelial-mesenchymal transition in bladder cancer [21]. We speculated that these seemingly contradictory results from hsa-miR-429 may reflect the diversity of TGFβ functions. TGFβ may exert these opposite effects partially through down-regulating hsa-miR-429.
In conclusion, overexpression of hsa-miR-429 contributes to the underexpression of CDKN2B in bladder cancer. Hsa-miR-429 promotes bladder cancer development via down-regulating CDKN2B.
MATERIALS AND METHODS
Patient samples
The primary tumor samples and paired adjacent normal urothelia were obtained from 50 patients with bladder urothelial carcinoma newly diagnosed at the Department of Urology, Shenzhen Second People’s Hospital. The clinicopathological features of the patients are provided in Table 1. The informed consent from the patients was obtained. All the patients have been followed up for at least 5 years. All procedures performed in studies involving human participants were in accordance with the ethical standards of the Institutional Review Board of the Second People’s Hospital of Shenzhen and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Cell culture and transfection
Bladder cancer T24 and 5737 cells and human embryo kidney 293T (HEK 293T, 293T) cells were purchased from the Institute of Cell Research, Chinese Academy of Sciences, Shanghai, China. The cells were routinely cultured in DMEM (Invitrogen, Carlsbad, CA, USA) plus 10% fetal bovine serum.
The cells were transfected with CDKN2B siRNA (target sequence: GAGAGCAATTGTAACGGTTAA)/Allstars Negative Control siRNA (final concentration: 20nM) (Qiagen, Hilden, Germany) or hsa-miR-429 mimic/negative control mimic (final concentration: 100nM) (Ribo, Guangzhou, China) using NanofectinTM Transfection Reagent (Excell Bio, Shanghai, China).
Total RNA extraction and reverse transcription
Total RNA was routinely extracted from the tissues or cells using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Ten micrograms of total RNA was converted to cDNA using the All-in-OneTM miRNA qRT-PCR Detection Kit (GeneCopoiea Inc, Rockville, MD, USA).
Real-time quantitative polymerase chain reaction (qPCR)
Hsa-miR-429 and homo sapiens snRNA U6 qPCR primers were purchased from GeneCopoiea Inc, Rockville, MD, USA. CDKN2B primers were forward:5’-GGAATGCGCGAGGAGAACAA-3’,reverse:5’-CATCA TCATGACCTGGATCGC-3’; TBP primers: 5’-CCCGAA ACGCCGAATATAATCC-3’ (forward), 5’-AATCAGTG CCGTGGTTCGTG-3’(reverse). The PCR procedures were as previously described [22, 23]. Expression fold changes were calculated using 2-ΔΔCt methods [24].
Western blot assay
Western blotting was performed as described elsewhere [25]. Antibodies specific to CDKN2B and GAPDH were obtained from Santa Cruz, Dallas, Texas, USA.
miRNA target prediction
miRanda (http://www.microrna.org), DIANA - microT v3.0 (http://www.http://diana.cslab.ece.ntua.gr/microT) and TargetScan (http://www.Targetscan.org) were used to predict binding sites between hsa-miR-429 and CDKN2B 3’UTR.
Dual luciferase reporter assay
The reporter vector CDKN2B 3’UTR-wild-type (WT) was created by chemically synthesizing and cloning the CDKN2B 3’UTR fragment 5’-ATATTTTTAAAGTTA CAGTATTT-3’(position2368, NM_004936) containing the predicted hsa-miR-429 binding site into the Xhol and Notl sites of the psiCHECK-2 vector (Promega, Madison, WI, USA). The reporter vector CDKN2B 3’UTR-mutated-type (MUT) was created by mutating the hsa-miR-429 seed region binding site (seed sequence binding fragment 5’-AGTATT-3’ changed to 5’-CCAGAA-3’).
Cell proliferation and cell apoptosis assays
Cell proliferation was determined using MTT assay. Apoptosis caused by CDKN2B siRNA and hsa-miR-429 mimic was determined using the Caspase 3 ELISA assay kit (R&D, Minneapolis, MN, USA). The procedures for cell proliferation and cell apoptosis assays were performed as previously described [22].
EdU incorporation
Cell proliferation was also detected using the EdU incorporation assay (Beyotime Inst Biotech, China). Briefly, 1×105 cells were seeded in each well of a 96-well flat-bottom plate. The cells within a single well were incubated with 100 μl of 50 μM EdU for 2 h, and then fixed with 50μl of fixing buffer for 30 min at room temperature. After removing the buffer, the cells were incubated with 50μl of 2 mg/ml glycine for 5 min and then washed with 100μl of PBS. The cells were also added with 100μl of permeabilization buffer followed by washing with 100μl of PBS. Subsequently, cells were added with 100μl of 1X Apollo solution for 30 min at room temperature in the dark. After that, cells were incubated with 100μl of 1X DAPI solution for 30 min at room temperature in the dark followed by washing with 100μl of PBS. The cells were finally observed by fluorescence microscopy.
Tumourigenicity assay
All experiments involving animals were approved by Institutional Review Board of Shenzhen 2nd People’s Hospital (Shenzhen, China). Stable transfection cell lines were constructed in our lab by transfecting the miR-429 expression vector or empty vector into T24 and 5637 cells and selecting with G-418. Four-week-old BALB/c athymic nude mice were bred in laminar flow cabinets and cultured at a constant humidity and temperature (25–28°C). In detail, 1×107 T24 or 5637cells were suspended in PBS and injected subcutaneously into the flank of nude mice (3 in each group) on day 0. Tumor growth was monitored by the tumor volume, which was calculated by the formula: Volume (mm3) = width2 (mm2) × length (mm) /2. The mice were sacrificed on day 42, and the tumors were excised. The in vivo experiment was repeated three times.
Statistical analysis
Paired samples t-test was used to analyze the expression differences between bladder cancer and matched urothelium. Pearson’s coefficient correlation was used for expression correlation assay. Survival curves were plotted using the Kaplan–Meier method and compared using the log-rank test. Independent samples t-test was used to analyze expression changes after transfection, luciferase activity and cell apoptosis. ANOVA was used to analyze cell proliferation. SPSS (Version 19.0) was used for statistical analyses. All P values were two-sided. A P value <0.05 was considered to be statistically significant.
ACKNOWLEDGMENTS
The authors thank all the donors whose names were not included in the author list, but who participated in this program.
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest with this work.
FUNDING
This work was supported by grants from the Shenzhen Key Medical Discipline Fund, Special Support Funds of Shenzhen for Introduced High-Level Medical Team, the National Science Foundation for Young Scientists of China [81602421],Shenzhen Foundation of Science and Technology (JCYJ20150330102720182). Shenzhen Health and Family Planning Commission Scientific Research Project [201506026, 201601025].
REFERENCES
1. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013; 63:11-30.
2. Lacy S, Lopez-Beltran A, MacLennan GT, Foster SR, Montironi R, Cheng L. Molecular pathogenesis of urothelial carcinoma: the clinical utility of emerging new biomarkers and future molecular classification of bladder cancer. Anal Quant Cytol Histol. 2009; 31:5-16.
3. Liu Y, Zhan Y, Chen Z, He A, Li J, Wu H, Liu L, Zhuang C, Lin J, Guo X, Zhang Q, Huang W, Cai Z. Directing cellular information flow via CRISPR signal conductors. Nat Methods. 2016; 13:938-44.
4. Guo H, Ingolia NT, Weissman JS, Bartel DP. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature. 2010; 466:835-40.
5. Croce CM. Causes and consequences of microRNA dysregulation in cancer. Nat Rev Genet. 2009; 10:704-14.
6. Yoneyama K, Ishibashi O, Kawase R, Kurose K, Takeshita T. miR-200a, miR-200b and miR-429 are onco-miRs that target the PTEN gene in endometrioid endometrial carcinoma. Anticancer Res. 2015; 35:1401-10.
7. Dong SJ, Cai XJ, Li SJ. The clinical significance of MiR-429 as a predictive biomarker in colorectal cancer patients receiving 5-fluorouracil treatment. Med Sci Monit. 2016; 22:3352-61.
8. Li X, Jiang H, Xiao L, Wang S, Zheng J. miR-200bc/429 inhibits osteosarcoma cell proliferation and invasion by targeting PMP22. Med Sci Monit. 2017; 23:1001-8.
9. Machackova T, Mlcochova H, Stanik M, Dolezel J, Fedorko M, Pacik D, Poprach A, Svoboda M, Slaby O. MiR-429 is linked to metastasis and poor prognosis in renal cell carcinoma by affecting epithelial-mesenchymal transition. Tumour Biol. 2016; 37:14653-8.
10. Wang Y, Dong X, Hu B, Wang XJ, Wang Q, Wang WL. The effects of Micro-429 on inhibition of cervical cancer cells through targeting ZEB1 and CRKL. Biomed Pharmacother. 2016; 80:311-21.
11. Han Y, Chen J, Zhao X, Liang C, Wang Y, Sun L, Jiang Z, Zhang Z, Yang R, Chen J, Li Z, Tang A, Li X, et al. MicroRNA expression signatures of bladder cancer revealed by deep sequencing. PLoS One. 2011; 6:e18286.
12. Wu CL, Ho JY, Chou SC, Yu DS. MiR-429 reverses epithelial-mesenchymal transition by restoring E-cadherin expression in bladder cancer. Oncotarget. 2016; 7:26593-603. doi: 10.18632/oncotarget.8557.
13. Navai N, Williams MB, Wen S, Siefker-Radtke AO, McConkey DJ, Dinney CP, Adam L. Relationship between high microRNA-200C expression and the risk of death from disease in muscle-invasive urothelial carcinoma of the bladder. J Clin Oncol. 2012; 30:279.
14. Martínez-Fernández M, Dueñas M, Feber A, Segovia C, García-Escudero R, Rubio C, López-Calderón FF, Díaz-García C, Villacampa F, Duarte J, Gómez-Rodriguez MJ, Castellano D, Rodriguez-Peralto JL, et al. A Polycomb-mir200 loop regulates clinical outcome in bladder cancer. Oncotarget. 2015; 6:42258-75. doi: 10.18632/oncotarget.5546.
15. Wu L, Fan J, Belasco JG. MicroRNAs direct rapid deadenylation of mRNA. Proc Natl Acad Sci U S A. 2006; 103:4034-9.
16. Han Y, Liu Y, Zhang H, Wang T, Diao R, Jiang Z, Gui Y, Cai Z. Hsa-miR-125b suppresses bladder cancer development by down-regulating oncogene SIRT7 and oncogenic long non-coding RNA MALAT1. FEBS Lett. 2013; 587:3875-82.
17. Jafri M, Wake NC, Ascher DB, Pires DE, Gentle D, Morris MR, Rattenberry E, Simpson MA, Trembath RC, Weber A, Woodward ER, Donaldson A, Blundell TL, et al. Germline mutations in the CDKN2B tumor suppressor gene predispose to renal cell carcinoma. Cancer Discov. 2015; 5:723-9.
18. Kim M, Yim SH, Cho NS, Kang SH, Ko DH, Oh B, Kim TY, Min HJ, She CJ, Kang HJ, Shin HY, Ahn HS, Yoon SS, et al. Homozygous deletion of CDKN2A (p16, p14) and CDKN2B (p15) genes is a poor prognostic factor in adult but not in childhood B-lineage acute lymphoblastic leukemia: a comparative deletion and hypermethylation study. Cancer Genet Cytogenet. 2009; 195:59-65.
19. Gao P, Si J, Yang B, Yu J. Upregulation of microRNA-15a contributes to pathogenesis of abdominal aortic aneurysm (AAA) by modulating the expression of cyclin-dependent kinase inhibitor 2B (CDKN2B). Med Sci Monit. 2017; 23:881-8.
20. Katoh Y, Katoh M. Hedgehog signaling, epithelial-to-mesenchymal transition and miRNA (review). Int J Mol Med. 2008; 22:271-5.
21. Al-Azayzih A, Gao F, Goc A, Somanath PR. TGFβ1 induces apoptosis in invasive prostate cancer and bladder cancer cells via Akt-independent, p38 MAPK and JNK/SAPK-mediated activation of caspases. Biochem Biophys Res Commun. 2012; 427:165-70.
22. Han Y, Liu Y, Nie L, Gui Y, Cai Z. Inducing cell proliferation inhibition, apoptosis, and motility reduction by silencing long noncoding ribonucleicacid metastasis-associated lung adenocarcinoma transcript 1 in urothelial carcinoma of the bladder. Urology. 2013; 81:209 e1-7.
23. VanGuilder HD, Vrana KE, Freeman WM. Twenty-five years of quantitative PCR for gene expression analysis. Biotechniques. 2008; 44:619-26.
24. Zhao J, Lei T, Xu C, Li H, Ma W, Yang Y, Fan S, Liu Y. MicroRNA-187, down-regulated in clear cell renal cell carcinoma and associated with lower survival, inhibits cell growth and migration though targeting B7-H3. Biochem Biophys Res Commun. 2013; 438:439-44.
25. Hunt S, Jones AV, Hinsley EE, Whawell SA, Lambert DW. MicroRNA-124 suppresses oral squamous cell carcinoma motility by targeting ITGB1. FEBS Lett. 2011; 585:187-92.