
INTRODUCTION
Cervical cancer is the fourth most common malignancy in women worldwide. There are appro-ximately 500,000 new cases of cervical cancer and 233,000 deaths caused by it per year [1]. Thus, cervical cancer is a major health problem worldwide. A deeper understanding of the genetic mechanisms that control the progression of the disease is imperative for enabling early clinical diagnosis and effective treatment of cervical cancer patients.
The study of long non-coding RNAs (lncRNAs) has recently gained prominence in the field of gene regulation research. LncRNAs are RNA molecules that are longer than 200 nucleotides in length and contain no protein-coding capacity [2]. Increasing evidence suggests that lncRNAs are involved in tumorigenic processes such as cell proliferation, apoptosis, differentiation, and invasion in [3, 4] in cervical cancer [5, 6], retinoblastoma [7], and oral squamous cell carcinoma [8].
Colon cancer associated transcript-1 (CCAT-1) is a type of lncRNA that is closely associated with colon cancer [9, 10]. However, recent studies have shown that CCAT-1 is also involved in gastric cancer [11], hepatocellular carcinoma [12], gallbladder cancer [13], and lung cancer [14, 15]. CCAT-1 gene is located on chromosome 8q24.21 in the vicinity of c-Myc, a well-known transcription factor [16–18]. Studies have shown that c-Myc can activate the transcription of CCAT-1, and the depletion of CCAT-1 RNA can reduce the effect of c-Myc [18, 19]. c-MYC is one of the most important oncogenes of cervical cancer [20, 21]; therefore, we propose that CCAT-1 might be involved in the progression of cervical cancer as well. However, the specific role of CCAT-1 in cervical cancer is still unclear. Further understanding of the role of CCAT-1 in cervical cancer development may provide better therapeutic opportunities for cervical cancer patients. Thus, the present study is aimed at investigating the role of CCAT-1 in cervical cancer.
First, the expression of CCAT-1 in cervical cancer tissues and its relationship with clinicopathological parameters were evaluated. Then we assayed the activity of CCAT-1 in cervical cancer cell lines. Finally, we studied the upstream and downstream regulatory factors affecting the expression of CCAT-1 in cervical cancer cell lines.
RESULTS
Expression of CCAT-1 in cervical cancer tissues and adjacent normal tissues
Expression levels of CCAT-1 in cervical cancer tissues (n=94) and matched, adjacent, normal tissues were detected by quantitative real-time PCR (qRT-PCR). Cervical cancer tissues showed a significantly higher expression than matched, adjacent normal tissues (P<0.01, Figure 1A).
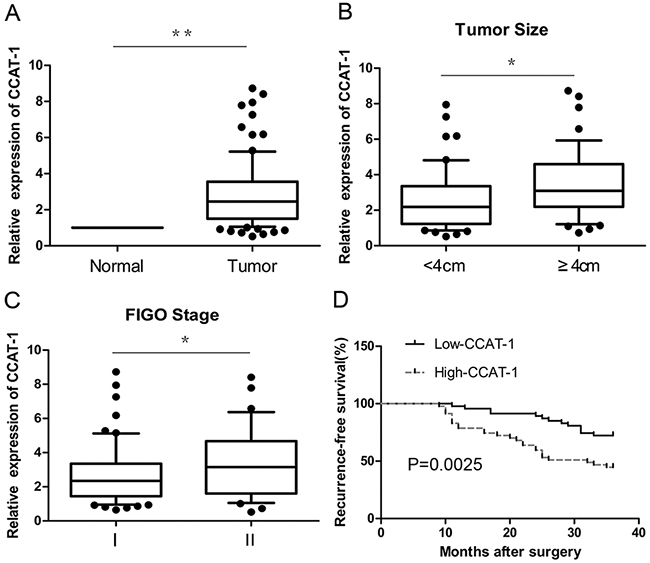
Figure 1: The relative expression of CCAT-1 in cervical cancer tissues. (A) The expression of CCAT-1 in cervical cancer tissues (n=94) was significantly higher than that in adjacent normal tissues. (B) The expression of CCAT-1 in patients with tumor size <4 cm was significantly lower than that in patients with tumor size ≥ 4 cm. (C) The expression of CCAT-1 in patients of FIGO stage I was significantly lower than that in FIGO stage II. (D) The recurrence-free survival of low-CCAT-1 group was significantly higher than that of high-CCAT-1 group. Each assay was performed in triplicate. *P < 0.05, **P < 0.01.
Relationship between CCAT-1 and clinicopathological parameters in cervical cancer
The results of qRT-PCR showed that the expression of CCAT-1 was related to the FIGO (International Federation of Gynecology and Obstetrics) stage and size of the tumor (P<0.05, Figure 1B, 1C). However, no significant relationship was observed between CCAT-1 levels and other parameters such as age and menopausal status of the patients, histological organization of the tissue, degree of differentiation, depth of invasion, lymphatic vascular space invasion (LVSI), and lymph node metastasis of the tumor (Table 1). Then, the median value of all 94 cervical cancer tissue samples was set as the cut-off point to separate tumors with low-level expression of CCAT-1 (low-CCAT-1 group) from those with high-level expression of CCAT-1 (high-CCAT-1 group). Kaplan-Meier survival analysis showed that recurrence-free survival of the low-CCAT-1 group was significantly higher than that of the high-CCAT-1 group (P=0.002, Figure 1D). COX univariate analysis showed that CCAT-1, FIGO stage, lymph node metastasis, and LVSI were prognostic factors for recurrence. Furthermore, COX multivariable analysis showed that only CCAT-1 expression level, FIGO stage, and lymph node metastasis were independent prognostic factors for recurrence (Table 2).
Table 1: Correlation between CCAT-1 and clinicopathological characteristics
Clinicopathologic feature |
n(%) |
CCAT-1 (mean±SEM) |
P-value |
|---|---|---|---|
Age |
0.411 |
||
≤50 |
52(55.3%) |
2.80±0.25 |
|
>50 |
42(44.7%) |
3.12±0.28 |
|
Menopause |
0.712 |
||
Yes |
54(57.4%) |
2.88±0.25 |
|
No |
40(42.6%) |
3.02±0.29 |
|
Histology |
0.918 |
||
Squamous cell cancer |
69(73.4%) |
2.95±0.20 |
|
Adenocarcinoma |
17(18.1%) |
2.43±0.36 |
|
Other |
8(8.5%) |
2.90±0.46 |
|
Differentiation |
0.925 |
||
Well to moderately |
58(61.7%) |
2.96±0.32 |
|
Poorly |
36(38.3%) |
2.92±0.32 |
|
Tumor size |
0.022 |
||
<4cm |
50(53.2%) |
2.55±0.23 |
|
≥4cm |
44(46.8%) |
3.41±0.29 |
|
Depth of invasion |
0.353 |
||
≤2/3 |
53(56.4%) |
2.79±0.27 |
|
>2/3 |
41(43.6%) |
3.14±0.26 |
|
Lymphatic vascular space invasion |
0.390 |
||
Negative |
47(50%) |
2.78±0.19 |
|
Positive |
47(50%) |
3.10±0.32 |
|
Lymph node metastasis |
0.122 |
||
Negative |
55(58.5%) |
2.72±0.22 |
|
Positive |
39(41.5%) |
3.33±0.34 |
|
FIGO stage |
0.025 |
||
I |
60(63.8%) |
2.59±0.26 |
|
II |
34(36.2%) |
3.44±0.25 |
Table 2: Univariate and multivariate analyses for recurrence-free survival
Risk factors |
Univariate analysis |
Multivariate analysis |
||||
|---|---|---|---|---|---|---|
HR |
P |
95 % CI |
HR |
P |
95 % CI |
|
CCAT-1 expression |
1.247 |
0.004 |
1.074~1.448 |
1.251 |
0.017 |
1.041~1.504 |
FIGO stage, (I , II) |
4.801 |
<0.001 |
2.482~9.284 |
4.097 |
<0.001 |
2.111~7.950 |
Lymph nodes metastasis (negative, positive) |
2.280 |
0.011 |
1.209~4.299 |
2.106 |
0.044 |
1.987~4.494 |
LVSI (negative, positive) |
2.035 |
0.031 |
1.067~3.883 |
1.422 |
0.307 |
0.724~0.279 |
Depth of invasion (≤2/3 , >2/3) |
1.565 |
0.163 |
0.835~2.934 |
|||
Differentiation (well/moderately, poorly) |
1.572 |
0.158 |
0.839~2.948 |
|||
Tumor size |
1.834 |
0.094 |
1.036~3.396 |
|||
Age |
1.006 |
0.673 |
0.978~1.034 |
|||
Histology (squamous, adenocarcinoma) |
1.246 |
0.580 |
0.572~2.710 |
|||
Menopause (yes, no) |
1.259 |
0.484 |
0.660~2.402 |
|||
HR (hazard ratio)
Effect of pcDNA-CCAT-1, si-CCAT-1, sh-CCAT-1, pcDNA-c-Myc, and si-c-Myc
qRT-PCR results showed that the expression of CCAT-1 was significantly increased in HeLa and CaSki cells transfected with an expression plasmid (pcDNA) carrying the CCAT-1 gene (pcDNA-CCAT-1), compared with cells transfected with an expression plasmid carrying a scrambled negative control (pcDNA-NC; P <0.05, Figure 2A). In contrast, the expression of CCAT-1 was significantly decreased in Hela and CaSki cells transfected with small-interfering CCAT-1 (si-CCAT-1) or short-hairpin CCAT-1 (sh-CCAT-1) constructs compared with their respective negative controls (si-NC and sh-NC; P <0.05, Figure 2B, 2C).
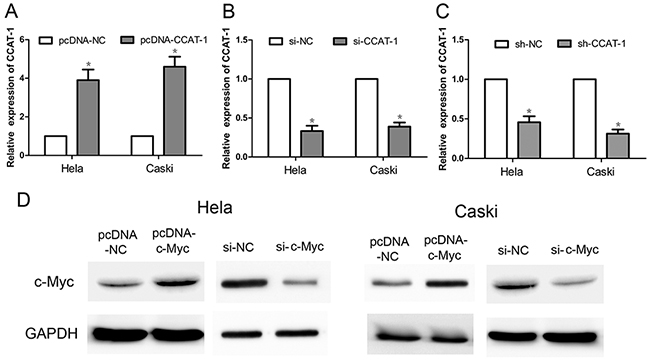
Figure 2: The relative expression of CCAT-1 and c-Myc after transfection of cervical cancer cells. (A) pcDNA-CCAT-1 could significantly increase the expression of CCAT-1 in HeLa and CaSki cells. (B, C) si-CCAT-1 and sh-CCAT-1 could down-regulate the expression of CCAT-1 in HeLa and CaSki cells. (D) pcDNA-c-Myc can significantly increase the expression of c-Myc protein in HeLa and CaSki cells, and si-c-Myc can significantly decrease the c-Myc protein expression in HeLa and CaSki cells. Each assay was performed in triplicate. *P < 0.05.
Western blotting showed that the expression of the c-Myc protein was significantly increased in HeLa and CaSki cells transfected with pcDNA-cMyc, compared with the control group (Figure 2D). Conversely, the expression of the c-Myc protein was significantly reduced in HeLa and CaSki cells transfected with si-cMyc compared with that in the control group (Figure 2D).
Effect of CCAT-1 on tumorigenic ability of cervical cancer cells
Tumorigenesis assays on nude mice showed that the tumorigenic ability of HeLa and CaSki cells transfected with sh-CCAT-1 was significantly lower than cells transfected with sh-NC (P <0.05, Figure 3A). Furthermore, qRT-PCR confirmed that the average CCAT-1 expression in tumors carrying sh-CCAT-1 was significantly lower than that of tumors carrying sh-NC (P <0.05, Figure 3B).
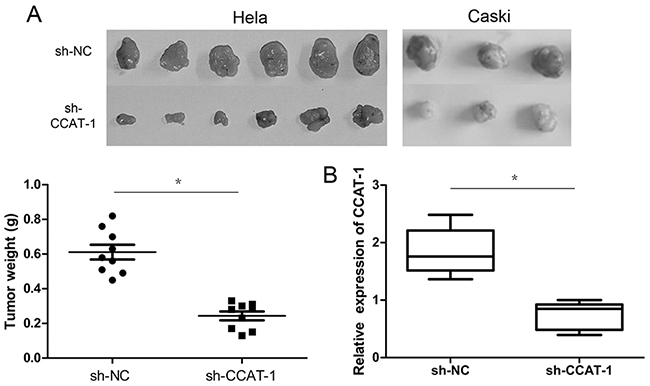
Figure 3: The effect of CCAT-1 on the proliferation of cervical cancer cells in vivo. (A) sh-CCAT-1 significantly inhibited the tumorigenic ability of HeLa and CaSki cells in vivo. (B) Expression levels of CCAT-1 were detected in xenograft tumor tissues by qRT-PCR. Each assay was performed in triplicate. *P < 0.05.
Effect of CCAT-1 on proliferation of cervical cancer cells
Cell Counting Kit-8 (CCK-8) assays showed that the proliferation of HeLa and CaSki cells transfected with pcDNA-CCAT-1 increased significantly compared with that of the negative control (P <0.05, Figure 4A). In contrast, the proliferation of HeLa and CaSki cells transfected with si-CCAT-1 was significantly inhibited compared to that of the control (P <0.05, Figure 4A).
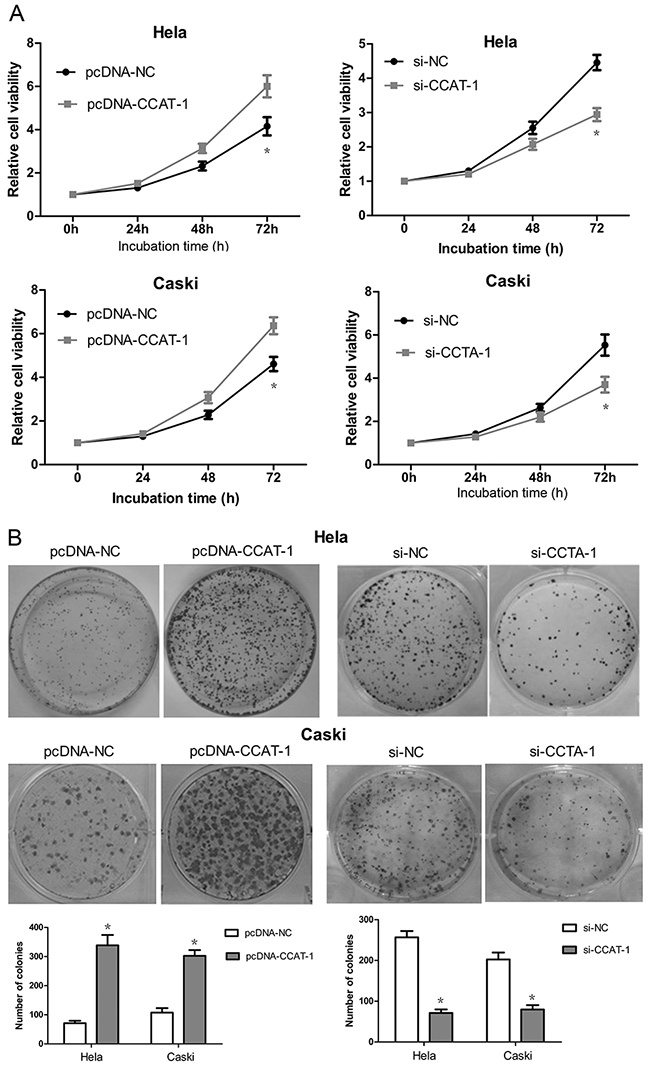
Figure 4: The effect of CCAT-1 on the proliferation of cervical cancer cells in vitro. (A) The growth curve and cell proliferative potential (CCK-8) were determined in HeLa and CaSki cells after pcDNA-CCAT-1 and si-CCAT-1 transfection compared to negative control. (B) Colony-formation assay was used to determine the cell proliferation ability at 48 h post-transfection. Each assay was performed in triplicate. *P < 0.05.
Effect of CCAT-1 on colony formation by cervical cancer cells
As depicted in Figure 4B, pcDNA-CCAT-1 transfection led to a significant increase in the number of colonies formed by HeLa and CaSki cells compared with that by the negative controls. We also observed a significant decrease in the number of colonies formed by si-CCAT-1-transfected cells compared with the negative controls.
Effect of CCAT-1 on apoptosis of cervical cancer cells
Flow cytometry showed that the percentage of early apoptotic cells among HeLa and CaSki cells transfected with pcDNA-CCAT-1 was significantly lower than that among control cells (P <0.05, Fig. 5). And compared with the control group, the percentage of early apoptotic cells in HeLa and CaSki cells transfected with si-CCAT-1 was significantly higher (P <0.05, Figure 5).
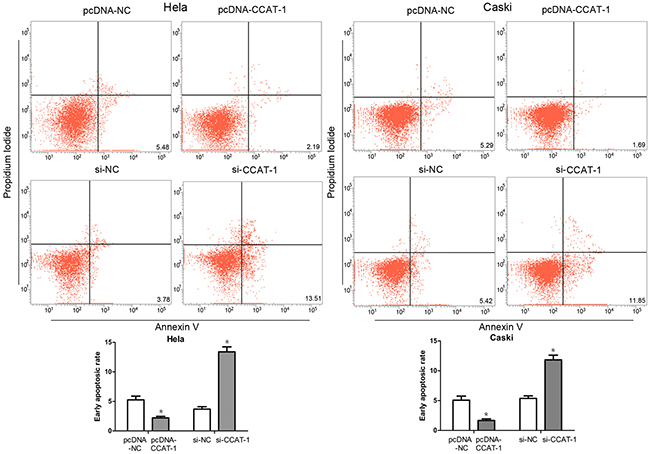
Figure 5: Flow cytometry was used to assess the early apoptotic rate of cervical cancer cells. The proportion of early apoptotic cells in pcDNA-CCAT-1 group was significantly lower than that in pcDNA-NC group. And the percentage of early apoptotic cells in HeLa and CaSki cells transfected with si-CCAT-1 was significantly higher than that in control cells. Each assay was performed in triplicate. *P < 0.05.
Effect of c-Myc on the expression of CCAT-1 in cervical cancer cells
First, our results showed that the expression of CCAT-1 was positively correlated with the expression of c-Myc in 94 samples of cervical cancer tissue (P<0.0001, Figure 6A). Then, in HeLa and CaSki cells the expression of CCAT-1 was significantly increased upon pcDNA-cMyc transfection and significantly decreased upon si-cMyc transfection, compared with that in their controls (P <0.05, Figure 6B, 6C).
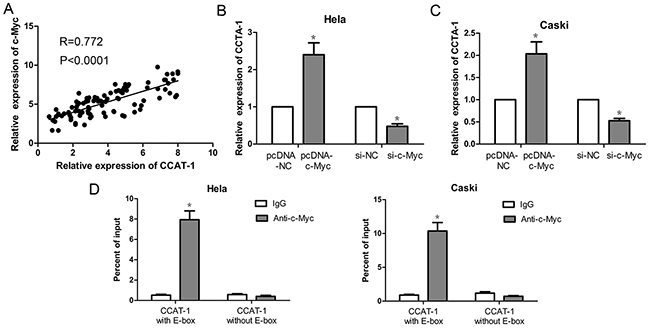
Figure 6: Expression of CCAT-1 was up-regulated by c-Myc protein. (A) There was a positive correlation between CCAT-1 and c-Myc mRNA expression in cervical cancer tissues (n=94). (B, C) Up-regulation of the expression of c-Myc protein in HeLa and CaSki cells could promote the transcription of CCAT-1. And down-regulated the expression of c-Myc protein in HeLa and CaSki cells could inhibit the transcription of CCAT-1. (D) ChIP assay was used to detect the binding of c-Myc protein to E-box of CCAT-1 promoter. Each assay was performed in triplicate. *P < 0.05.
Furthermore, we examined the binding of c-Myc to the E-box of the CCAT-1 promoter region using chromatin immunoprecipitation assay. For CCAT-1 promoter containing a wild-type E-box, we found that the amount of c-Myc immunoprecipitates was significantly higher than that of lgG immunoprecipitates (P <0.05, Figure 6D). However, for the CCAT-1 promoter with the E-box absent, there was no difference in binding between c-Myc and lgG as revealed by the lack of statistically significant difference in the amount of c-Myc and IgG immunoprecipitates (Figure 6D).
Effect of CCAT-1 on the activity of Wnt pathway
The activity of luciferase was found to be significantly higher in HeLa and CaSki cells transfected with pcDNA-CCAT-1 construct compared with cells transfected with the control pcDNA-NC construct by TOP Flash assays. Conversely, the activity of luciferase was significantly lower in HeLa and CaSki cells transfected with si-CCAT-1 compared with that in control cells (P <0.05, Figure 7A). In addition, western blotting showed that β-catenin protein levels were significantly increased in the pcDNA-CCAT-1 transfected group compared with the pcDNA-NC transfected group. Conversely, the level of β-catenin in HeLa and CaSki cells transfected with si-CCAT-1 was significantly lower than that in the controls (P <0.05, Figure 7B).
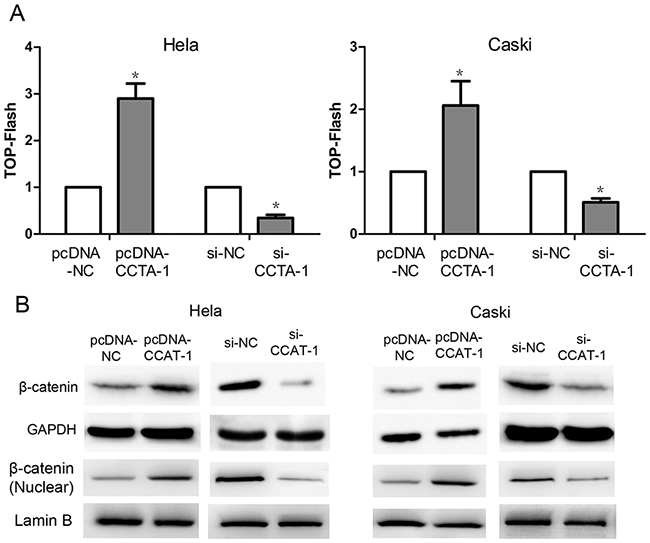
Figure 7: The regulation of Wnt pathway by CCAT-1. (A) TOP-Flash assays showed that CCAT-1 might up-regulate the Wnt pathway activity and down-regulation of CCAT-1 might significantly decrease the Wnt pathway activity of HeLa and CaSki cells. (B) Western blot showed that up-regulation of CCAT-1 expression in HeLa and CaSki cells resulted in a significant increase of β-catenin protein level, while down-regulation of CCAT-1 in HeLa and CaSki cells resulted in a significant decrease of β-catenin protein level. Each assay was performed in triplicate. *P < 0.05.
Finally, we used the Wnt/β-catenin pathway anta-gonist Dkk-1 and the Wnt/β-catenin pathway acti-vator LiCl to reverse the effects of pcDNA-CCAT-1 and si-CCAT-1 in HeLa and CaSki cells (Figure 8A). CCK-8 assays showed that the proliferation of HeLa and CaSki transfected with pcDNA-CCAT-1 and Dkk-1 was significantly lower than cells transfected only with pcDNA-CCAT-1. Conversely, the proliferation of HeLa and CaSki si-CCAT-1 transfected cells was significantly enhanced when they were treated with LiCl compared with untreated cells (P <0.05, Figure 8B).
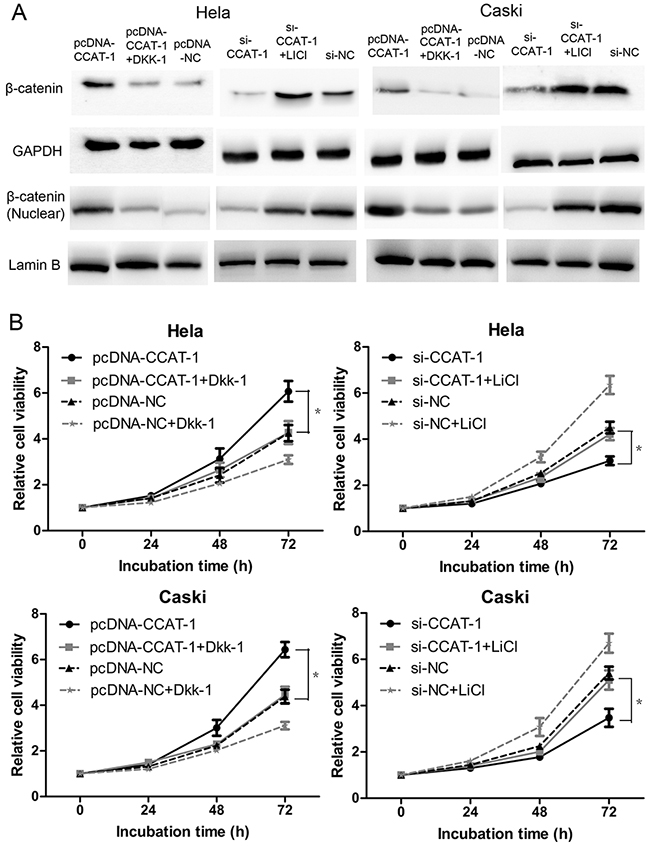
Figure 8: CCAT-1 achieved its biological functions via regulating Wnt pathway. (A) Wnt pathway antagonist Dkk-1 and activator LiCl just reversed the regulatory effect of pcDNA-CCAT-1 and si-CCAT-1 on Wnt pathway. (B) The CCAT-1 biological function was impaired after reversing the regulatory effect of CCAT-1 on Wnt pathway. Each assay was performed in triplicate. *P < 0.05.
DISCUSSION
The study of lncRNAs has gained prominence in the field of gene regulation in recent years. Accumulated evidence from recently published studies suggests that aberrant lncRNA expression may be a crucial component of tumorigenesis in cervical cancer [22–24]. CCAT-1, a newly discovered lncRNA, is involved in the development of tumors in many different tissues [9–16]. However, its role in cervical cancer is unknown.
In this study, we first discovered that CCAT-1 was overexpressed in cervical cancer tissue compared with that in normal, adjacent tissue. Then we analyzed the relationship between CCAT-1 expression and clinicopathological characteristics of the patients. We found that CCAT-1 expression was associated with the FIGO stage and size of the tumor. CCAT-1 shows a similar overexpression in other cancers [9–16]. Since FIGO stage and tumor size are key factors in the prognosis of cervical cancer, we hypothesized that CCAT-1 expression levels too might show a direct correlation with prognosis. Our data confirmed that CCAT-1 expression served as an independent prognostic factor for cervical cancer. Our results indicate that CCAT-1 expression can be employed as a potential tool for preoperative identification and postoperative follow-up of patients at high risk for recurrent cervical cancer.
Since we observed that CCAT-1 expression was associated with tumor size, we speculated that it has a role in promoting the proliferation of cervical cancer cells. Therefore, we conducted CCK-8 and flow cytometry assays on cells that were made to overexpress CCAT-1. We found that CCAT-1 overexpression promoted proliferation and inhibited apoptosis in cervical cancer cell lines, HeLa and CaSki. Conversely, silencing the expression of CCAT-1 by RNA interference inhibited proliferation and promoted apoptosis in cervical cancer cell lines. These results confirmed that CCAT-1 functions by promoting cervical cancer cell growth. We simultaneously conducted tumor formation assays on nude mice injected with HeLa and CaSki cells carrying recombinant CCAT-1 constructs and confirmed that silencing of CCAT-1 expression significantly inhibited the tumorigenicity of cervical cancer cells. Although these results may be corroborated by manipulating CCAT1 expression in tumors of other tissues, they did demonstrate that CCAT-1 promoted tumor growth and functioned as an oncogene in cervical cancer in our study.
Previous studies have shown that c-Myc can activate CCAT-1 expression in colon and gastric cancers, and human bronchial epithelial cells [10, 11, 14, 15]. Additionally, the c-Myc gene has also been confirmed to promote the tumorigenicity of cervical cancer cells [20, 21]. However, whether c-Myc can activate CCAT-1 expression in cervical cancer cells remains unknown. We first analyzed the relationship between the expression of CCAT-1 and c-Myc in samples of cancerous tissue and found that the two are positively correlated, suggesting that there may be a regulatory relationship between CCAT-1 and c-Myc. Subsequently, we found that CCAT-1 expression was significantly upregulated upon overexpressing c-Myc in cervical cancer cells. In contrast, silencing c-Myc expression significantly inhibited CCAT-1 expression in cervical cancer cells. These results suggested that c-Myc activity upregulates the expression of CCAT-1 in cervical cancer cells. Furthermore, to determine whether c-Myc could directly bind to sites within the CCAT-1 promoter, chromatin immunoprecipitation assays were performed. We found that c-Myc could bind to the wild-type E-box within the CCAT-1 promoter, but could not bind to the CCAT-1 promoter when the E-box was absent. Together, these results systematically revealed a mechanism for expression of CCAT-1 in cervical cancer cells: c-Myc promotes the expression of CCAT-1 by binding to the E-box of its promoter, leading to the proliferation of cervical cancer cells.
The Wnt/β-catenin pathway regulates several cellular processes such as proliferation, invasion, and differentiation [25]. Previous studies have shown that Wnt/β-catenin pathway activation can promote cell proliferation and inhibit cell apoptosis in cervical cancer [26, 27]. Therefore, we also aimed to explore the regulatory relationship between CCAT-1 and the Wnt/β-catenin pathway. We used TOP-Flash reporter assays to first show that downregulation of CCAT-1 can significantly inhibit Wnt/β-catenin pathway activity in cervical cancer cells. Furthermore, we found that upregulating the expression of CCAT-1 could significantly activate the Wnt/β-catenin pathway in cervical cancer cells. Western blotting confirmed that upregulation of CCAT-1 significantly increased the expression of the β-catenin protein, while downregulation of CCAT-1 expression resulted in a significant reduction in the expression of the β-catenin protein. It has been demonstrated in previous studies that LiCl activates the Wnt/β-catenin pathway by inhibiting GSK3β and stabilizing β-catenin, while DKK-1 can inhibit the Wnt/β-catenin pathway by binding to LRP6 [28, 29]. Therefore, we used DKK-1 as an antagonist and LiCl as an agonist to reverse the effects of CCAT-1 manipulation on the Wnt/β-catenin pathway in cervical cancer cells. The functional assays revealed that the biological function of CCAT-1 was significantly weakened in cervical cancer cells after the reversal. These results suggested that the Wnt/β-catenin pathway is an important downstream target of CCAT-1 for promoting proliferation and inhibiting apoptosis in cervical cancer cells.
Altogether, we propose the following model for the proliferation of cervical cancer cells: overexpression of c-Myc → enhancement of CCAT-1 gene transcription → activation of Wnt/β-catenin pathway → enhanced cell proliferation. This study helps us to better understand the specific role of CCAT-1 in malignant growth of cervical cancer cells.
In summary, our study indicated that CCAT-1 overexpression was related to tumor size and prognosis in cervical cancer. The expression of CCAT-1 might be caused by the binding of the c-Myc protein to the promoter of CCAT-1. CCAT-1 might promote proliferation and inhibit apoptosis of cervical cancer cells by activating the Wnt/β-catenin pathway. In conclusion, the study and manipulation of CCAT-1 may prove to be useful for prognosis and treatment of cervical cancer.
MATERIALS AND METHODS
Clinical samples
A total of 94 paired cervical cancer tissues and adjacent normal tissues were obtained from the cervical cancer patients with surgical treatment between January 2011 and May 2012. Freshly specimens were immediately frozen in liquid nitrogen and stored at -80 ° C until used. All patients did not receive any preoperative tumor-related treatment, and pathology was confirmed as cervical cancer by two pathologists. Written informed consent was obtained from each patient, and the present study was approved by the ethics committee of University. The clinical and pathological data of patients was shown in Table 1.
Cell culture conditions
Human cervical cancer cell lines (HeLa, CaSki) were obtained from the Cell Bank of the Chinese Academy of Sciences. Cells were cultured in DMEM (Thermo Scientific, Belmont, Massachusetts, US) containing 10% fetal bovine serum (Gibco, Gaithersburg, USA) at 37°C in a humidified chamber supplemented with 5% CO2.
Establish the stable shRNA-CCAT-1 cervical cancer cell lines by lentivirus
The lentiviral vectors with shRNA-CCAT-1 were constructed by the GenePharma Company (Shanghai, China). Briefly, the oligonucleotide encoding shRNA-CCAT-1 and negative control were annealed and inserted into the pCMV vector (GenePharma). The recombinational vectors and the packaging vectors were co-transfected into 293T cells (Invitrogen, Carlsbad, USA) with Lipofectamine2000 (Invitrogen, USA). The culture supernatants were collected at 48 hours after transfection, concentrated. The cells were divided into two groups according our objectives: infection of control virus (sh-NC); infection of shRNA-CCAT-1 virus (sh-CCAT-1). The shRNA-CCAT1 sequences used showed in Supplementary Table 1.
Transient transfection
pcDNA-CCAT-1, si-CCAT-1, pcDNA-c-Myc, si-c-Myc and their negative controls were purchased from GenePharma Company (Shanghai, China). Transient transfection was performed using the Lipofectamine 2000 (Invitrogen, USA) following the manufacturer’s instructions. Forty-eight hours after transfection, the cells were collected for further experiments. siRNA sequences were listed in Supplementary Table 1.
RNA extraction and quantitative real-time PCR (qRT-PCR)
Total RNAs from tissues and cells were extracted using TRIzol (TaKaRa, Otsu, Japan). After reverse transcription reaction with Prime-ScriptTM one step RT-PCR kit (TaKaRa, Otsu, Japan), qPCR was conducted with SYBR Premix Ex Taq (TaKaRa, Otsu, Japan) on CFX96™ Real-Time PCR Detection System supplied with analytical software (Bio-Rad, USA). GAPDH were used as endogenous control and the relative expression of RNAs was calculated using the 2−ΔΔCt method. The primers used in this study were synthesized by GenePharma Company (Shanghai, China) and the sequences were shown in Supplementary Table 2.
Cell proliferation assay
Cell viability was determined using the CCK8 assay according to the manufacturer’s protocol. Briefly, Cells were plated in 96-well plates in triplicate at 5×103 per well in a final volume of 90 μl. Then 10 μl CCK8 solutions was added to each well and incubated for 2h at 37 °C and 5% CO2 at each time point (0, 24, 48, 72h). After incubating, the absorbance at 450 nm was measured by an SpectraMax M5 (Molecular Devise, California, USA).
Apoptosis assay
Apoptotic fractions cells were analysed by Annexin-V-FLUOS Kit (BD Pharminogen, USA) according to the manufacturer’s instructions. Annexin V and PI was added 48h after transfection and apoptotic cells were identified with a flow cytometer (FACSCalibur, BD Biosciences). The relative ratio of early apoptotic cells were counted for further comparisons.
Colony-formation assay
Cells were plated at 500 per well into 6-well plates in DMEM supplemented with 10% FBS. Dishes were incubated at 37°C in a humidified atmosphere containing 5% CO2. After 7 days, cells were washed twice with PBS, fixed with formaldehyde for 30 min, stained with crystal violet (Beyotime; Shanghai, China) for 10 min, washed with ddH2O three times and then photographed with a digital camera.
Xenograft tumor model
HeLa and CaSki cells (2×107cells) that stably transfected with pCMV/shRNA-CCAT-1 or pCMV/shRNA-NC were re-suspended in 150 μl PBS and subcutaneously injected into the flank tissues of 5-week-old BALB/c-nu female mice, respectively. 21 days after inoculation, the animals were sacrificed, the xenografts were isolated and the weight (gram) of the xenografts was determined. The protocols for animal experiment study were approved by the Institutional Committee for Animal Research in University.
Western blot analysis
Cells were lysed at 4°C for 30 min using RIPA buffer (Thermo Scientific, Massachusetts, USA) that contained the protease inhibitor cocktail (Roche, Basel, Switzerland). Nuclear protein extracts were prepared using the Nuclear Extraction Kit (Sangon Biotechology, Shanghai, China) according to the manufacturer’s instructions. Protein lysates were separated by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto PVDF membrane (Roche, Basel, Switzerland). The membrane was blocked in 5% nonfat milk for 1 h at room temperature and subsequently incubated with specific antibody against β-catenin (1:1000, Cell Signaling Technology, Danvers, USA) and c-Mcy (1:1000, Cell Signaling Technology, Danvers, USA) overnight at 4°C. Afterwards, the membrane was washed and incubated with HRP Goat-anti-Rabbit (1:2000; Santa Cruz Biotechnology; Dallas, USA) for 1 h. Protein detection was performed using an ECL chemiluminescence reagent (Thermo Fisher Scientific, MA, USA). Lamin B (1:200, Santa Cruz Biotechnology) served as a nuclear protein internal control and GAPDH (1:1000, Cell Signaling Technology, Danvers, USA) served as a total protein internal control.
Chromatin immunoprecipitation assay
Chromatin immunoprecipitation (ChIP) assays were performed according to the EZ ChIP Chromatin Immunoprecipitation Kit (Millipore, Bedford, MA, USA). Briefly, HeLa and CaSki cells were treated with formaldehyde and incubated for 10 min to generate DNA-protein cross-links. Cell lysates were then sonicated to generate chromatin fragments of 200–300 bp and immunoprecipitated with antibodies against c-Myc (Santa Cruz Biotechnology, Santa Cruz, CA, USA) or IgG as control. The antibody-bound complex was precipitated by Protein A-Sepharose beads. The DNA fragments in the immunoprecipitated complex were released by reversing the cross-linking at 65 °C for 5 hour and purified DNA was analyzed by qRT-PCR with SYBR-Green incorporation (Applied Biosystems, Foster City, CA, USA). The ΔCt was calculated as ΔCt (normalized ChIP) = [Ct (ChIP) – Ct (Input)]. The % input was shown as 2[-ΔCt(normalized ChIP)]. The primers specific for the CCAT1 promoter containing E-box and non-E-box were shown in Supplementary Table 2.
TOP Flash/FOP-Flash reporter assay
The β-catenin reporter plasmid (TOP-flash) and its mutant control (FOP-flash) were constructed by Millipore Corporation (Massachusetts, USA). Cells were serum-starved overnight and co-transfected with 0.2 μg TOP flash or FOP flash expression plasmids and 0.1 μg pRL-TK (Renilla TK-luciferase vector; Promega, Madison, USA) using Lipofectamine 2000. The activities of both firefly and Renilla luciferase reporters were determined at 48 hours after transfection using a Dual Luciferase Assay Kit (Promega, Madison, WI, USA) according to the manufacturer’s instructions. The TOP-Flash reporter activity is presented as the relative ratio of firefly luciferase activity to Renilla luciferase activity, and the TOP/FOP ratio was used as a measure of β-catenin-driven transcription.
Treatment with wnt activators and wnt antagonists
The following regulators of the Wnt signalling pathway were used: Wnt activators LiCl (Sigma-Aldrich, MO, USA) and Wnt antagonist Dickkopf-1 (Dkk1, PeproTech, New Jersey, USA). At 24 hours after transfection, the transfected cells were treated for 48 hours with LiCl (25mmol/L) or Dkk1 (120ng/ml). Following that, cells were collected for further experiments.
Statistical analysis
SPSS 11.0 and GraphPad Prism software was used for statistical analyses. Values were expressed as mean ± standard error. Student’s t-test was used to analyze the differences between two groups and P < 0.05 was considered statistically significant. All experiments were performed at least three times and all samples analyzed in triplicate.
Abbreviations
lncRNA: long noncoding RNA; CCAT-1: colon cancer associated transcript 1; ChIP: chromatin immunoprecipitation; CCK-8: Cell Counting Kit-8; LVSI: lymphatic vascular space invasion; qRT-PCR: quantitative real-time polymerase chain reaction.
Author contributions
ZJ and GY conceived and designed the project. ZJ performed patient collection and clinical data interpretation. ZJ and GY participated performed the statistical analysis. ZJ and GY performed the experiments and wrote the manuscript. ZJ and GY contributed to the writing and to the critical reading of the paper. All authors read and approved the final manuscript. All authors read and gave their approval for the final version of the manuscript.
ACKNOWLEDGMENTS
Not applicable.
CONFLICTS OF INTEREST
The authors have no conflicts of interest to declare.
FUNDING
The Project Supported by Health and Family Commission of Shenzhen Municipality Foundation (Grant No. 201507010), Medical Scientific and Technology Research Foundation of Guangdong Province, China (Grant No. A2017569) and Shenzhen Science and Technology Plan Project (Grant No.JCYJ20170307095222274).
REFERENCES
1. Tewari KS, Sill MW, Long HJ, Penson RT, Huang H, Ramondetta LM, Landrum LM, Oaknin A, Reid TJ, Leitao MM, Michael HE, Monk BJ. Improved survival with bevacizumab in advanced cervical cancer. N Engl J Med. 2014; 370:734-743.
2. Spizzo R, Almeida MI, Colombatti A, Calin GA. Long noncoding RNAs and cancer: a new frontier of translational research? Oncogene. 2012; 31:4577-4587.
3. Wapinski O, Chang HY. Long noncoding RNAs and human disease. Trends Cell Biol. 2011; 21:354-361.
4. Gutschner T, Diederichs S. The hallmarks of cancer: a long non-coding RNA point of view. RNA Biol. 2012; 9:703-719.
5. Zhang J, Yao T, Wang Y, Yu J, Liu Y, Lin Z. Long noncoding RNA MEG3 is downregulated in cervical cancer and affects cell proliferation and apoptosis by regulating miR-21. Cancer Biol Ther. 2016; 17:104-113.
6. Zhang J, Lin Z, Gao Y, Yao T. Downregulation of long noncoding RNA MEG3 is associated with poor prognosis and promoter hypermethylation in cervical cancer. J Exp Clin Cancer Res. 2017; 36:5.
7. Gao Y, Lu X. Decreased expression of MEG3 contributes to retinoblastoma progression and affects retinoblastoma cell growth by regulating the activity of Wnt/β-catenin pathway. Tumour Biol. 2016; 37:1461-1469.
8. Zhou X, Liu S, Cai G, Kong L, Zhang T, Ren Y, Wu Y, Mei M, Zhang L, Wang X. Long non coding RNA MALAT1 promotes tumor growth and metastasis by inducing epithelial-mesenchymal transition in oral squamous cell carcinoma. Sci Rep. 2015; 5:15972.
9. Nissan A, Stojadinovic A, Mitrani-Rosenbaum S, Halle D, Grinbaum R, Roistacher M, Bochem A, Dayanc BE, Ritter G, Gomceli I, Bostanci EB, Akoglu M, Chen YT, et al. Colon cancer associated transcript-1: a novel RNA expressedin malignant and pre-malignant human tissues. Int J Cancer. 2012; 130:1598-1606.
10. He X, Tan X, Wang X, Jin H, Liu L, Ma L, Yu H, Fan Z. c-Myc-activated long noncoding RNA CCAT1 promotes colon cancer cell proliferation and invasion. Tumour Biol. 2014; 35:12181-8.
11. Yang F, Xue X, Bi J, Zheng L, Zhi K, Gu Y, Fang G. Long noncoding RNA CCAT-1, which could be activated by c-Myc, promotes the progression of gastric carcinoma. J Cancer Res Clin Oncol. 2013; 139:437-445.
12. Deng L, Yang SB, Xu FF, Zhang JH. Long noncoding RNA CCAT1 promotes hepatocellular carcinoma progression by functioning as let-7 sponge. J Exp Clin Cancer Res. 2015; 34:18.
13. Ma MZ, Chu BF, Zhang Y, Weng MZ, Qin YY, Gong W, Quan ZW. Long non-coding RNA CCAT1 promotes gallbladder cancer development via negative modulation of miRNA-218-5p. Cell Death Dis. 2015; 6:e1583.
14. Lu L, Qi H, Luo F, Xu H, Ling M, Qin Y, Yang P, Liu X, Yang Q, Xue J, Chen C, Lu J, Xiang Q, et al. Feedback circuitry via let-7c between lncRNA CCAT1 and c-Myc is involved in cigarette smoke extract-induced malignant transformation of HBE cells. Oncotarget. 2017; 8:19285-19297. doi: 10.18632/oncotarget.15195.
15. Chen J, Zhang K, Song H, Wang R, Chu X, Chen L. Long noncoding RNA CCAT1 acts as an oncogene and promotes chemoresistance in docetaxel-resistant lung adenocarcinoma cells. Oncotarget. 2016; 7:62474-62489. doi: 10.18632/oncotarget.11518.
16. Xin Y, Li Z, Shen J, Chan MT, Wu WK. CCAT1: a pivotal oncogenic long non-coding RNA in human cancers. Cell Prolif. 2016; 49:255-260.
17. Kam Y, Rubinstein A, Naik S, Djavsarov I, Halle D, Ariel I, Gure AO, Stojadinovic A, Pan H, Tsivin V, Nissan A, Yavin E. Detection of a long non-coding RNA (CCAT1) in living cells and human adenocarcinoma of colon tissues using FIT-PNA molecular beacons. Cancer Lett. 2014; 352:90-96.
18. Xiang JF, Yin QF, Chen T, Zhang Y, Zhang XO, Wu Z, Zhang S, Wang HB, Ge J, Lu X, Yang L, Chen LL. Human colorectal cancer-specific CCAT1-L lncRNA regulates long-range chromatin interactions at the MYC locus. Cell Res. 2014; 24:513-531.
19. Younger ST, Rinn JL. ‘Lnc’-ing enhancers to MYC regulation. Cell Res. 2014; 24:643-644.
20. Liao LM, Sun XY, Liu AW, Wu JB, Cheng XL, Lin JX, Zheng M, Huang L. Low expression of long noncoding XLOC_010588 indicates a poor prognosis and promotes proliferation through upregulation of c-Myc in cervical cancer. Gynecol Oncol. 2014; 133:616-623.
21. Kübler K, Heinenberg S, Rudlowski C, Keyver-Paik MD, Abramian A, Merkelbach-Bruse S, Büttner R, Kuhn W, Schildhaus HU. c-myc copy number gain is a powerful prognosticator of disease outcome in cervical dysplasia. Oncotarget. 2015; 6:825-835. doi: 10.18632/oncotarget.2706.
22. Kim HJ, Eoh KJ, Kim LK, Nam EJ, Yoon SO, Kim KH, Lee JK, Kim SW, Kim YT. The long noncoding RNA HOXA11 antisense induces tumor progression and stemness maintenance in cervical cancer. Oncotarget. 2016; 7:83001-83016. doi: 10.18632/oncotarget.12863.
23. Jiang B, Sun R, Fang S, Qin C, Pan X, Peng L, Li Y, Li G. Lnc-CC3 increases metastasis in cervical cancer by increasing Slug expression. Oncotarget. 2016; 7:41650-41661. doi: 10.18632/oncotarget.9519.
24. Yang L, Yi K, Wang H, Zhao Y, Xi M. Comprehensive analysis of lncRNAs microarray profile and mRNA-lncRNA co-expression in oncogenic HPV-positive cervical cancer cell lines. Oncotarget. 2016; 7:49917-49929. doi: 10.18632/oncotarget.10232.
25. MacDonald BT, Tamai K, He X. Wnt/beta-catenin signaling: components, mechanisms, and diseases. Dev Cell. 2009; 17:9-26.
26. Ramachandran I, Thavathiru E, Ramalingam S, Natarajan G, Mills WK, Benbrook DM, Zuna R, Lightfoot S, Reis A, Anant S, Queimado L. Wnt inhibitory factor 1 induces apoptosis and inhibits cervical cancer growth, invasion and angiogenesis in vivo. Oncogene. 2012; 31:2725-2737.
27. Chung MT, Lai HC, Sytwu HK, Yan MD, Shih YL, Chang CC, Yu MH, Liu HS, Chu DW, Lin YW. SFRP1 and SFRP2 suppress the transformation and invasion abilities of cervical cancer cells through Wnt signal pathway. Gynecol Oncol. 2009; 112:646-653.
28. Rowe MK, Chuang DM. Lithium neuroprotection: molecular mechanisms and clinical implications. Expert Rev Mol Med. 2004; 6:1-18.
29. Binnerts ME, Kim KA, Bright JM, Patel SM, Tran K, Zhou M, Leung JM, Liu Y, Lomas WE 3rd, Dixon M, Hazell SA, Wagle M, Nie WS, et al. R-Spondin1 regulates Wnt signaling by inhibiting internalization of LRP6. Proc Natl Acad Sci U S A. 2007; 104:14700-14705.