
INTRODUCTION
Vitiligo is an acquired pigmentary disorder of the skin and mucous membranes, and is characterized by a chronic and progressive loss of melanocytes from the epidermis and follicular reservoir. Several hypotheses have been proposed to explain the pathomechanism of vitiligo, including the neural theory, an impaired redux status and autoimmunity [1]. Recently, oxidative stress has been suggested to be the initial pathogenic event in melanocyte degeneration and evidence has been presented for very high levels of H2O2 (1 mM) and peroxynitrite in the epidermis of patients with active vitiligo disease, concomitant with reduced levels and activity of catalase [2–4].
Melanocytes dendrites have crucial roles in the adhesion, migration, and melanosome transfer. In human skin, each melanocyte makes contact with several keratinocytes via their dendrites. A new theory called ‘melanocytorrhagy’ has been proposed that depigmentation in vitiligo results from a chronic detachment and subsequent transepidermal elimination of melanocytes, which is possibly related to increased susceptibility to mechanical and other types of stresses like oxidative stress [5]. Schallreuter et al. [4]. Proved that the addition of H2O2 to established cultures of normal human melanocytes induced a loss of dendricity and in some cases caused melanocyte detachment. Attachment during dendrite outgrowth requires extracellular Ca2+, and H2O2 induces an impaired uptake and efflux of Ca2+ in melanocytes that hinders the dendrite outgrowth and elongation [6].
Mitochondria are the main targets of reactive oxygen species (ROS) which can result in changes of mitochondrial structure and function, including Ca2+ homeostasis, cellular redox state regulation and so on [7]. Picardo’s group has demonstrated an imbalance of antioxidants in the peripheral blood mononuclear cells of active vitiligo patients, which appeared to be due to a mitochondrial impairment[8]. In this regard, it is likely that mitochondria play a pivotal role in the etiopathogenesis of vitiligo.
Centella asiatica (L.) Urb. is a perennial herbaceous creeper of the Apiaceae family, which grows in tropical regions of Asia, Oceania, Africa and America. It contains several triterpenes, namely, asiatic acid, madecassic acids, asiaticoside and madecassoside. The medical applications of C. asiatica involves its anti-inflammatory [9], memory improvement [10], anticancer [11], antihepatoma [12] and antioxidation [13] effects. C. asiatica is also recommended for the treatment of dermatoses and skin lesions such as excoriations, hypertrophic scars, eczema, healing wounds, psoriasis and scleroderma[14]. Madecassoside is the main active ingredient of C. asiatica with a higher content than other triterpenoid constituents, and its antioxidant capacity has been confirmed [15]. Madecassoside is extracted from C. asiatica with a purity of 99%. Some traditional Chinese Medicine doctors have also found its therapeutic effects for external use in the repigmentation of vitiligo and postinflammatory hypopigmentation, but its mechanism of action has not been elucidated. In this study, we evaluated the effects of madecassoside on oxidative stress in human melanocytes, particularly focusing on mitochondria and dendritic morphology, and we discuss its possible mechanism of action.
RESULTS
Effect of H2O2 on normal human melanocyte dendrites
The addition of 0.01, 0.005 or 0.001 mM H2O2 to the culture medium induced the retraction of dendrites and the detachment of melanocytes when replacing the culture medium (Figure 1). Moreover, the dendrites of 0.01 mM H2O2-treated melanocytes retracted faster than the other two groups (Table 1), and dendrite retraction was observed as early as 5 min after H2O2 incubation. Considering that a low concentration of H2O2 would make it rather difficult to observe the obvious dendritic retractions, and would increase factors beyond the control of the experiment, we used 0.01 mM H2O2 throughout the remainder of these experiments.
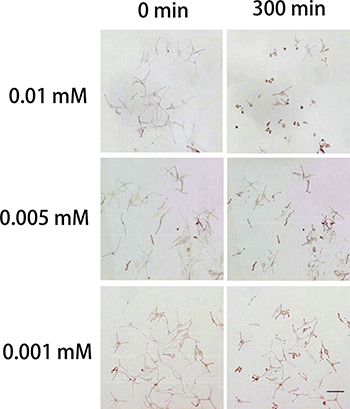
Figure 1: Effects of H2O2 on normal human melanocytes. Images were taken before treatment and at 300 min after treatment with 0.01, 0.005 or 0.001 mM H2O2 in the culture medium. H2O2 induced the retraction of dendrites and the detachment of melanocytes when removing the culture medium and rinsing cells to replace the medium. 0.01 mM H2O2 caused a more obvious dendritic retraction than the other two concentrations of H2O2. Bar = 200 μm (Original magnification 100×)
Table 1: Retraction velocity of melanocytes following H2O2 treatment (n = 60, mean ± SD)
Concentration of H2O2(mM) |
Retraction velocity of melanocytes (μm/min) |
P value |
|---|---|---|
0.01 |
0.310 ± 0.021 |
|
0.005 |
0.199 ± 0.034 |
0.0007* |
0.001 |
0.189 ± 0.017 |
0.0003* |
*indicates a statistically significant difference (P < 0.05) compared with 0.01 mM H2O2-treated melanocytes. 0.005 mM vs 0.001 mM, P = 0.621.
Madecassoside reduces the retraction of melanocyte dendrites under oxidative stress
Changes of dendrites retraction were observed over time after the addition of H2O2 with or without madecassoside. Dendrites of only H2O2-treated melanocytes retracted dramatically by 300 min and some dendrites even disappeared. However, the concurrent addition of madecassoside at concentrations of 10, 50 or 100 μg/mL induced only slight dendritic retractions over the same time course (Figure 2), and the retraction velocities of those dendrites were reduced to 0.256 ± 0.021, 0.215 ± 0.017 and 0.167 ± 0.024 μm/min respectively (Table 2). Compared with the retraction velocities in only H2O2-treated melanocytes (0.310 ± 0.021 μm/min), madecassoside inhibited the retraction of melanocyte dendrites in a concentration-dependent manner.
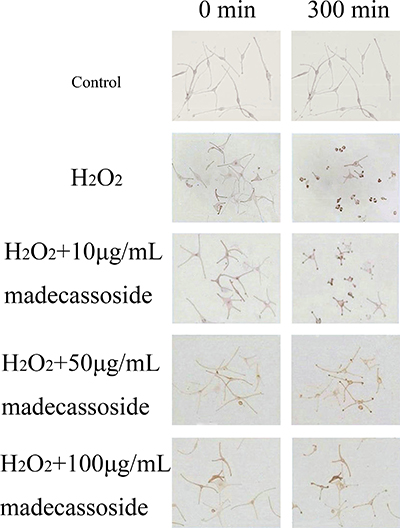
Figure 2: Effect of madecassoside on H2O2-treated human melanocytes. Melanocyte dendrites retracted dramatically after 5 hr when treated only by H2O2. The addition of madecassoside reduced the dendritic retraction velocities under oxidative stress. The higher the concentration of madecassoside, the greater the suppressive effect on dendritic retractions. Bar = 100 μm (Original magnification 200×).
Table 2: Retraction velocity of melanocytes following different treatments (n = 100, mean ± SD)
Treatment |
Retraction velocity of melanocytes (μm/min) |
P value |
|---|---|---|
Control |
0.019 ± 0.008 |
0.001* |
H2O2 |
0.310 ± 0.021 |
|
H2O2+10 μg/mL madecassoside |
0.256 ± 0.021 |
0.004* |
H2O2+50 μg/mL madecassoside |
0.215 ± 0.017 |
0.003* |
H2O2+100 μg/mL madecassoside |
0.167 ± 0.024 |
0.003* |
* indicates a statistically significant difference (P < 0.05) compared with H2O2-treated melanocytes.
Effect of madecassoside on MMP in melanocytes under oxidative stress
As shown in Figure 3, H2O2 treatment reduced MMP which decreased from 98.68 ± 3.28% to 41.57 ± 3.91%, but the concurrent treatment with 10, 50 or 100 μg/mL madecassoside rescued MMP to 58.28 ± 5.07%, 76.42 ± 4.06% and 91.87 ± 4.52%, respectively. In addition, there was a significant difference between each madecassoside-treated group. These results revealed that madecassoside and improves MMP in melanocytes under oxidative stress.
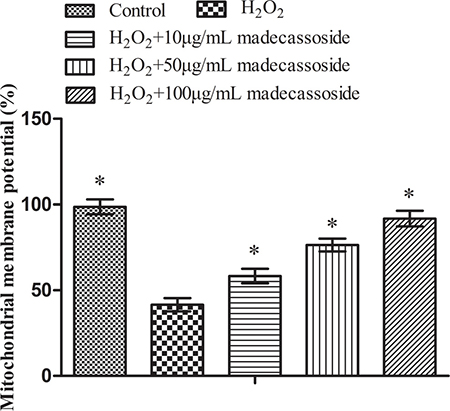
Figure 3: Madecassoside improves the MMP under oxidative stress. *P < 0.05 vs. H2O2 treatment group.
Effect of madecassoside on free intracellular calcium ([Ca2+]i) in H2O2-treated melanocytes
Mitochondria play an important role in maintaining Ca2+ homeostasis. To some extent, the disruption of Ca2+ homeostasis reflects mitochondrial dysfunction. After exposure to H2O2, the level of [Ca2+]i increased significantly. However, a reduction of [Ca2+]i in a concentration-dependent manner was observed in the presence of madecassoside (10, 50 or 100 μg/mL) (Figure 4). These results suggest that madecassoside reduces the accumulation of [Ca2+]i and sequentially recovers Ca2+ homeostasis in melanocytes under oxidative stress.
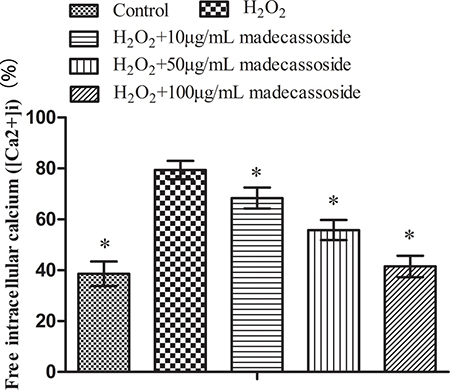
Figure 4: Effect of madecassoside on decreasing the level of free [Ca2+]i in H2O2-treated melanocytes. *P < 0.05 vs. H2O2 treatment group.
Protective effects of madecassoside on oxidative stress-induced ultrastructural changes in melanocytes
Some indications of damage in melanocytes were observed by TEM in the H2O2 treatment group, including the disruption of cytomembrane integrity, a decreased quantity of cristae as well as swelling and vacuolization of mitochondria (Figure 5). However, the degree of mitochondria injury in madecassoside-treated melanocytes was much less than that in H2O2-treated melanocytes, and a large number of normal mitochondria, even some autophagosomes, could be found.
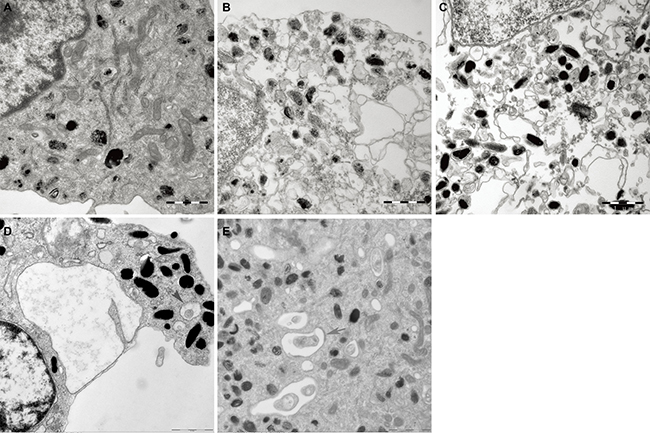
Figure 5: Ultrastructural alterations of mitochondria in melanocytes. (A) The structure and the number of mitochondria in the untreated control group were quite normal. (B) Damaged of mitochondria in H2O2-treated melanocytes were observed, including disruption of the cytomembrane, as well as swelling and vacuolization of mitochondria. (C–E) corresponding 10, 50 and 100 μg/mL madecassoside-treated melanocytes, respectively. Madecassoside attenuated the damage of mitochondria caused by H2O2. Autophagosomes could be also observed (grey arrow).(Original magnification 30,000×).
The stereology results are shown in Table 3. In comparison with the control group, the Vv and Sv of mitochondria in melanocytes treated with H2O2 were significantly higher, whereas the level of Nv was lower, which indicated the swelling of mitochondria. After exposure to madecassoside, the Vv and Sv decreased, accompanied by an increase of Nv. Remarkable differences were also noted between the H2O2 and the madecassoside-treated melanocytes for Vv, Sv and Nv with increasing concentrations of madecassoside.
Table 3: Stereology analysis of mitochondria after different treatments (n = 50, mean ± SD)
Treatment |
Nv (μm-2) |
Sv (‰) |
Vv (%) |
P value |
P value |
P value |
|---|---|---|---|---|---|---|
Control |
8.25 ± 0.98 |
3.66 ± 0.76 |
5.34 ± 0.95 |
0.001* |
0.002Δ |
0.002** |
H2O2 |
3.84 ± 1.03 |
6.93 ± 0.84 |
9.13 ± 1.06 |
|||
H2O2+10μg/mL madecassoside |
5.73 ± 0.94 |
5.81 ± 1.01 |
7.99 ± 0.88 |
0.004* |
0.006Δ |
0.005** |
H2O2+50μg/mL madecassoside |
6.49 ± 0.87 |
4.68 ± 0.96 |
6.85 ± 0.78 |
0.003* |
0.004Δ |
0.003** |
H2O2+100μg/mL madecassoside |
7.22 ± 0.95 |
4.03 ± 0.75 |
6.27 ± 0.92 |
0.001* |
0.004Δ |
0.002** |
* indicates a statistically significant difference in Nv (P < 0.05) compared with H2O2-treated melanocytes.
Δ indicates a statistically significant difference in Sv (P < 0.05) compared with H2O2-treated melanocytes.
**indicates a statistically significant difference in Vv (P < 0.05) compared with H2O2-treated melanocytes.
Effect of madecassoside on autophagy activation in H2O2-treated melanocytes
AO is a fluorescent substance that is often used to detect the occurrence of autophagy. Madecassoside induced a far stronger red punctate fluorescence in the cytoplasm of H2O2-treated melanocytes in comparison with the only H2O2-treated melanocytes (Figure 6). Moreover, the number of AO-positive acidic vesicular organelles (AVOs) in the madecassoside-treated cells was increased in a concentration-dependent manner.
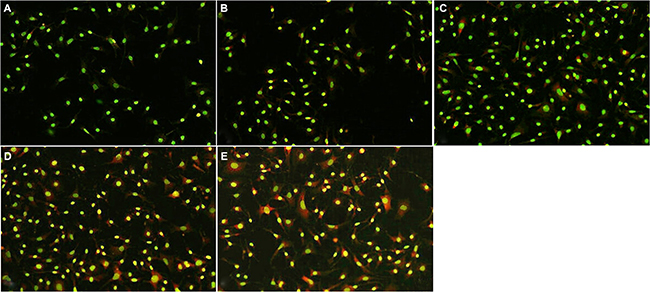
Figure 6: Madecassoside induced autophagy activation in H2O2-treated melanocytes. The number of red fluorescent spots in the cytoplasm of madecassoside-treated melanocytes was increased in a concentration-dependent manner. (A) Untreated control group, (B) H2O2-treated melanocytes, and (C–E) 10, 50 or 100 μg/mL madecassoside-treated melanocytes, respectively. (Original magnification 100×).
Madecassoside increases the LC3-II/LC3-I ratio in melanocytes under oxidative stress
LC3-II is a significant marker of autophagosomes in mammalian cells[17]. Levels of LC3-II and LC3-I in H2O2-treated melanocytes were up-regulated significantly by simultaneous incubation with 10, 50 or 100 μg/mL madecassoside. The relative ratio of LC3-II/LC3-I also increased markedly in a concentration-dependent manner compared with H2O2-treated melanocytes (P < 0.05, Figure 7). Hence, madecassoside activates the LC3-I to LC3-II conversion in melanocytes under oxidative stress.
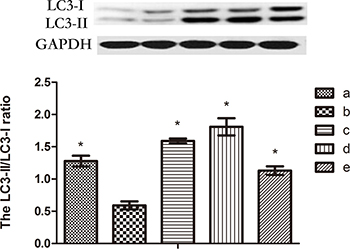
Figure 7: Western blot analysis of LC3. The protein level of the LC3-II/LC3-I ratio increased significantly following madecassoside treatment, where compared with the H2O2 treatment group. (A) Untreated control group, (B) H2O2-treated melanocytes, and (C–E) 10, 50 or 100 μg/mL madecassoside-treated melanocytes, respectively. *P < 0.05 vs. H2O2 treatment group.
DISCUSSION
Vitiligo is a multifactorial polygenic disorder with a complex pathogenesis. Recently, melanocytorrhagy arising from defective cell-cell adhesion has been proposed as a pathomechanism for vitiligo, which considers nonsegmental vitiligo as a disease caused by the chronic detachment and transepidermal loss of melanocytes[18]. Kumar et al. demonstrrated that the dendrites of melanocytes from patients with unstable nonsegmental vitiligo are small with clubbed ends, and show a significantly lower adhesion to collagen type IV compared with control and stable vitiligo melanocytes [19]. These observations support the melanocytorrhagic theory. Dendrites are important not only for melanosome transfer, but also dramatically increase the adhesion and anchoring of melanocytes within the basal layer of the epidermis. Routine observation of melanocytes in culture indicates that dendrite retraction comes first, before cell detachment and eventual death. Increasing evidence shows that oxidative stress caused by H2O2 is a key factor in the onset and progression of vitiligo [1]. More importantly, it has been suggested that the loss of melanocytes in vitiligo may be the result of oxidative stress and melanocytorrhagy acting in unison. ROS could induce the loss of dendricity of melanocytes, which might not only affect melanosome transfer but could also exaggerate traumatic transepidermal loss of genetically adhesion-deficient melanocytes [5]. Additionally, extracellular Ca2+ is essential for dendrite outgrowth and attachment. It is well known that mitochondria play an essential role in maintaining calcium homeostasis, and they are the main source of ROS, as well as an unifying target of ROS, so the status of mitochondrial structure and function may be strongly related with the onset of vitiligo.
In this study, 0.01 mM H2O2 was employed to induce of oxidative stress in human melanocytes, and we found an obvious retraction of dendrites, structural damage and functional disruption of mitochondria, including a decreased quantity of their cristae, swelling and vacuolization a decrease of MMP and the accumulation of [Ca2+]i in melanocytes under oxidative stress. It has been reported that the major mechanism for mitochondrial Ca2+ transport is primarily via the mitochondrial Ca2+ uniporter (MCU) and Ca2+ uptake is driven by the MMP [20]. Accordingly, we speculate that the stimulation of ROS might lead to the opening the mitochondrial permeability transition pore (MPTP) and MMP depolarization, followed by the disruption of Ca2+ homeostasis, suggesting that dendrite retraction might be related to the overload of [Ca2+]i.
Our results show that the detrimental effects of H2O2 on melanocytes are alleviated by madecassoside in a concentration-dependent manner. Our observations showed that madecassoside remarkably reverses the decrease of MMP and inhibits the accumulation of [Ca2+]i induced by oxidative stress. In addition, madecassoside protects the structure of mitochondria against oxidative stress. Mitochondrial swelling and/or vacuolization was only rarely observed and a large number of normal mitochondria could be found by TEM. The stereology data revealed that Vv and Sv of mitochondria in melanocytes treated with H2O2 were much higher than those in the untreated control melanocytes and decreased following treatment with increasing concentrations of madecassoside, and the changes of Nv were contrary to the Vv and Sv. Taken together, the underlying mechanism involved in the protective effect of madecassoside on melanocyte dendrites might be its antioxidative impact including the protection of MMP and the recovery of Ca2+ homeostasis.
Autophagy is a cellular degradation pathway essential for survival. As a protective mechanism, autophagy safeguards organisms against normal and pathological aging by regulating the turnover of dysfunctional organelles and proteins [21]. It is thought that damaged or excess mitochondria are eliminated by autophagy, and TEM revealed the formation of autophagic vacuoles in melanocytes treated with madecassoside. Accordingly, we speculate that madecassoside might play a role in the antioxidant defense against melanocyte loss through autophagy. To further investigate that possibility, we also examined the effect of madecassoside treatment on the formation of AVOs in melanocytes using fluorescence microscopy. As shown in Figure 6, madecassoside-treated melanocytes displayed primarily red fluorescence in their cytoplasm with minimal green fluorescence, indicating the abundance of AVOs. Futher, the LC3-II/LC3-I ratio was analyzed using western blots. During autophagy, the cytosolic form of LC3 (LC3-I) is cleaved and lipidated to form the phosphatidylethanolamine-conjugated form (LC3-II) that is recruited to the autophagosomal membrane [22]. The degree of autophagosome formation can be measured using the LC3-II/LC3-I ratio and/or by the expression level of LC3-II. We observed that madecassoside-treated melanocytes had a higher LC3-II expression and also an increased LC3-I conversion into LC3-II compared to H2O2-treated melanocytes, suggesting an increased activation of autophagy. This proves that autophagy occurs in melanocytes following treatment with madecassoside against oxidative stress.
MATERIALS AND METHODS
Cell culture
Human melanocytes were obtained from normal foreskins of 10 young healthy male subjects aged 7–10 years old (skin phototype III or IV, Fitzpatrick classification) after circumcision, according to the method of Swope et al. [16]. This study was conducted after signed consent of each participant in accordance with the guidelines in the Declaration of Helsinki Principles and was approved by the local Ethics Committees of the First Affiliated Hospital of Nanjing Medical University. Briefly, each foreskins was sterilized in 70% ethanol, cut into small pieces (4 × 4 mm), and then incubated in 5 mg/mL Dispase II (Sigma, St Louis, MO, USA) overnight at 4°C. The epidermis was separated using forceps and was further digested with 0.25% trypsin/EDTA (Gibco, Carlsbad, CA, USA) at 37°C for 30 min. After centrifugation, cells were resuspended in growth medium Medium 254 with human melanocyte growth supplement (Cascade Biologics, Portland, OR, USA) and were cultured in a humidified incubator with 5% CO2/95% air at 37°C. During the primary culture of human melanocytes, 200 mg/mL G418 (geneticin sulfate, Duchefa, Haarlem, Netherlands) was added to the growth medium to suppress the proliferation of fibroblasts. The culture medium was changed every other day thereafter until the cells became confluent. All experiments were performed using human melanocytes at passages 2–5. Melanocytes were identified by staining with the antibody HMB45 and Masson-Fontana (both from Life Technologies, Carlsbad, CA, USA) as markers of PMEL17 and melanin, respectively.
Observation of melanocytes dendrites
To determinate the optimal concentration of H2O2 to observe melanocytes dendrites under oxidative stress conditions in vitro, the cells were photographed using a light microscope (Olympus BX51, Aizu, Japan) before treatment and every 5 min after treatment with various concentrations of H2O2 (0.01, 0.005, 0.001 mM, Nanjing Chemical Reagent Factory, Nanjing, China). The lengths of dendrites were measured using AutoCAD Software (Autodesk, Inc. San Rafael, CA, USA). Twenty melanocytes in each group were randomly selected to calculate the retraction velocity according to Eq. (1).
Retraction velocity = (Lengthinitial− Lengthfinal )/time (μm/min)1
Subsequently, cells were treated with H2O2 (0.01 mM) and final concentrations of madecassoside (10, 50, 100 μg/mL) concurrently, and the assessment of retraction velocity of melanocyte dendrites was performed as noted above. Cells cultured without madecassoside or H2O2 served as a control group. Madecassoside powder was a gift from Shanghai Shyndec Pharmaceutical Co. Ltd. (Shanghai, China)
Determination of mitochondrial membrane potential (MMP)
JC-1 dye exhibits potential dependent accumulation in mitochondria, indicated by a change in fluorescence emission from green to red. Consequently, mitochondrial depolarization is indicated by a decrease in the red/green fluorescence intensity ratio. In brief, human melanocytes were grown in 6-well plates to approximately 90% confluence, and then were treated with H2O2 (0.01 mM) and/or a variety of concentrations of madecassoside (10, 50, 100 μg/mL) for 5 hr. A total of 0.5 mL of each cell suspension collected from the different groups was added to 0.5 mL JC-1 solution (Sigma, St Louis, MO, USA) followed by incubation at 37°C for 15 min. Each cell suspension was then centrifuged at 60 g for 5 min to remove the supernatant. The cell pellets were resuspended in JC-1 solution and analyzed by flow cytometry using a FACScan and Cell Quest software (Becton Dickinson, Mountain View, CA, USA). Mean fluorescence intensity values for FL1 (green fluorescence) and FL2 (red fluorescence) were obtained, and the ratio of Red/Green Fluorescence intensity values provided an estimate of the MMP (ΔΨm).
Intracellular calcium measurements by flow cytometry
Levels of free intracellular calcium ([Ca2+]i) were measured using the cell-permeable calcium-sensitive fluorescent dye Fluo-3/AM (Sigma, St Louis, MO, USA). After treatment with H2O2 (0.01 mM) and/or final concentrations of madecassoside (0, 10, 50, 100 μg/mL) for 5 hr, cells were collected and incubated with 5 μM Fluo-3/AM for 35 min at 37°C. The cells were washed twice with PBS, and the fluorescence intensity of Fluo-3/AM probes was analyzed by flow cytometry using Cell Quest software (Becton Dickinson, Mountain View, CA) with excitation at 488 nm and emission at 530 nm.
Transmission electron microscopy (TEM)
Human melanocytes were treated with H2O2 (0.01 mM) and/or a variety of final concentrations of madecassoside (10, 50, 100 μg/mL) for 5 hr and were harvested. Cell pellets were washed twice with PBS and fixed with 3% glutaraldehyde at 4 °C overnight. Thereafter they were postfixed with PBS containing 1% (v/v) OsO4, dehydrated in a series of graded ethanol (70–100%), embedded in Durcopan (Durcopan, Fluka Chemie, Buchs, Switzerland) and sectioned using a LEICA Ultracut UCT ultramicrotome (Leica Microsystems, Wetzlar, Germany). Ultrathin sections were stained with uranyl acetate and lead citrate, and were examined using TEM (JEOL Ltd., Tokyo, Japan). Images were captured and input into the electron microscope image analysis system for morphological quantitative analysis of melanocytes and mitochondria. The mean long axis, minor axis, circumference and area of 10 melanocytes in each specimen were measured. The numeral density (Nv), surface density (Sv) and volume density (Vv) of mitochondria were calculated by Eq. (2), Eq. (3) and Eq. (4), respectively.
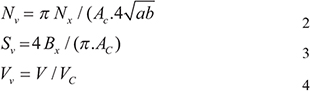
Nx: number of mitochondria in the cytoplasm, Ac: area of the cytoplasm, a: long axis of mitochondria, b: minor axis of mitochondria, Bx: circumference of mitochondria, V : mitochondrial volume, Vc: cytoplasmic volume
Acridine orange (AO) staining
Melanocytes for AO fluorescent staining were cultured in 6-well plates at a density of 1 × 105 cells/well. After treatment with various concentrations of madecassoside (0, 10, 50, 100 μg/mL) as well as 0.01 mM H2O2 for 5 hr, cells were incubated in PBS with 1 μg/mL AO (Sigma, St. Louis, MO, USA) for 15 min at room temperature in the dark, and then were washed twice by PBS. Acidic vacuoles were detected using a fluorescence microscope (Nikon TE2000-U, Shizvoka, Japan). The protonated form of AO accumulates in acidic compartments and forms aggregates, which are characterized by red fluorescence.
Western blot analysis
Melanocytes were inoculated in 6-well plates at 2 × 105 cells/well, and were concurrently treated with H2O2 (0.01 mM) and madecassoside at final concentrations of 0, 10, 50, 100 μg/mL for 5 hr. Each well was then harvested by rinsing twice with cold PBS, and was lysed in RIPA buffer containing protease inhibitors. Lysates were cleared by centrifugation at 290g for 15 min and protein concentrations were determined using a Bradford protein assay kit (Bio-Rad Laboratories, Hercules, CA, USA). Equal amounts of protein were resolved by SDS-PAGE and were transferred to nitrocellulose membranes (Schleicher & Schuell, Keene, NH, USA) by electroblotting. Membranes were then blocked and incubated at 4°C overnight with primary antibodies [rabbit anti-LC3 (1:1000, L7543, Sigma, St. Louis, MO, USA) and mouse anti-GAPDH (1:1000, G8795, Sigma, St. Louis, MO, USA)]. Subsequently, the membranes were washed and incubated for 1–2 hr at room temperature with horseradish peroxidase-conjugated anti-rabbit or anti-mouse as the secondary antibody (1:1000, Jackson ImmunoResearch Lab., West Grove, PA, USA). Specific signals were detected with the ECL detection reagent (Pierce, Rockford, IL USA). The quantification of LC3 was analyzed by Quantity One software (Bio-Rad, Hercules, CA, USA).
Statistics
Data are expressed as means ± SD. All statistical analyses were performed with SPSS 16.0 software (International Business Machines). Comparisons among 3 or more groups were assessed via one-way analysis of variance, and differences among means were analyzed using Least significant difference (LSD). A P < 0.05 is considered significant.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards. This article does not contain any studies with animals performed by any of the authors.
ACKNOWLEDGMENTS
This study was supported by the Science and Technology Social Development Plan of Jiangsu Province (No. BS2007072) and by the National Natural Science Foundation of China (No. 81171517).
CONFLICTS OF INTEREST
The authors have no conflicts of interest to declare.
REFERENCES
1. Namazi MR. Neurogenic dysregulation, oxidative stress, autoimmunity, and melanocytorrhagy in vitiligo: can they be interconnected? Pigment Cell Res. 2007; 20:360–3.
2. Schallreuter KU, Gibbons NC, Zothner C, Abou Elloof MM, Wood JM. Hydrogen peroxide-mediated oxidative stress disrupts calcium binding on calmodulin: more evidence for oxidative stress in vitiligo. Biochem Biophys Res Commun. 2007; 360:70–5.
3. Schallreuter KU, Moore J, Wood JM, Beazley WD, Gaze DC, Tobin DJ, Marshall HS, Panske A, Panzig E, Hibberts NA. In vivo and in vitro evidence for hydrogen peroxide (H2O2) accumulation in the epidermis of patients with vitiligo and its successful removal by a UVB-activated pseudocatalase. J Investig Dermatol Symp Proc. 1999; 4:91–6.
4. Schallreuter KU, Wood JM, Lemke KR, Levenig C. Treatment of vitiligo with a topical application of pseudocatalase and calcium in combination with short-term UVB exposure: a case study on 33 patients. Dermatology. 1995; 190:223–9.
5. Gauthier Y, Cario AM, Taieb A. A critical appraisal of vitiligo etiologic theories. Is melanocyte loss a melanocytorrhagy? Pigment Cell Res. 2003; 16:322–32.
6. Hara M, Yaar M, Tang A, Eller MS, Reenstra W, Gilchrest BA. Role of integrins in melanocyte attachment and dendricity. J Cell Sci. 1994; 107:2739–48.
7. Prakash A, Kumar A. Mitoprotective effect of Centella asiatica against aluminum-induced neurotoxicity in rats: possible relevance to its anti-oxidant and anti-apoptosis mechanism. Neurol Sci. 2013; 34:1403–9.
8. Dell'Anna ML, Maresca V, Briganti S, Camera E, Falchi M, Picardo M. Mitochondrial impairment in peripheral blood mononuclear cells during the active phase of vitiligo. J Invest Dermatol. 2001; 117:908–13.
9. Guo JS, Cheng CL, Koo MW. Inhibitory effects of Centella asiatica water extract and asiaticoside on inducible nitric oxide synthase during gastric ulcer healing in rats. Planta Med. 2004; 70:1150–4.
10. Veerendra KM, Gupta YK. Effect of different extracts of Centella asiatica on cognition and markers of oxidative stress in rats. J Ethnopharmacol. 2002; 79:253–60.
11. Babu TD, Kuttan G, Padikkala J. Cytotoxic and anti-tumour properties of certain taxa of Umbelliferae with special reference to Centella asiatica (L.) Urban. J Ethnopharmacol. 1995; 48:53–7.
12. Lin LT, Liu LT, Chiang LC, Lin CC. In vitro anti-hepatoma activity of fifteen natural medicines from Canada. Phytother Res. 2002; 16:440–4.
13. Gupta R, Flora SJ. Effect of Centella asiatica on arsenic induced oxidative stress and metal distribution in rats. J Appl Toxicol. 2006; 26:213–22.
14. Bylka W, Znajdek-Awiżeń P, Studzińska-Sroka E, Dańczak-Pazdrowska A, Brzezińska M. Centella asiatica in dermatology: an overview. Phytother Res. 2014; 28:1117–24.
15. Leng DD, Han WJ, Rui Y, Dai Y, Xia YF. In vivo disposition and metabolism of madecassoside, a major bioactive constituent in Centella asiatica (L.) Urb. J Ethnopharmacol. 2013; 150:601–8.
16. Swope VB, Medrano EE, Smalara D, Abdel-Malek ZA. Long-term proliferation of human melanocytes is supported by the physiologic mitogens alpha-melanotropin, endothelin-1, and basic fibroblast growth factor. Exp Cell Res. 1995; 217:453–9.
17. Kabeya Y, Mizushima N, Ueno T, Yamamoto A, Kirisako T, Noda T, Kominami E, Ohsumi Y, Yoshimori T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000; 19:5720–8.
18. Gauthier Y, Cario-Andre M, Lepreux S, Pain C, Taïeb A. Melanocyte detachment after skin friction in non lesional skin of patients with generalized vitiligo. Br J Dermatol. 2003; 148:95–101.
19. Kumar R, Parsad D, Kanwar AJ. Role of apoptosis and melanocytorrhagy: a comparative study of melanocyte adhesion in stable and unstable vitiligo. Br J Dermatol. 2011; 164:187–91.
20. Brookes PS, Yoon Y, Robotham JL, Anders MW, Sheu SS. Calcium, ATP, and ROS: a mitochondrial love-hate triangle. Am J Physiol Cell Physiol. 2004; 287:C817–33.
21. López de Figueroa P, Lotz MK, Blanco FJ, Caramés B. Autophagy activation and protection from mitochondrial dysfunction in human chondrocytes. Arthritis Rheumatol. 2015; 67:966–76.
22. Miracco C, Cevenini G, Franchi A, Luzi P, Cosci E, Mourmouras V, Monciatti I, Mannucci S, Biagioli M, Toscano M, Moretti D, Lio R, Massi D. Beclin 1 and LC3 autophagic gene expression in cutaneous melanocytic lesions. Hum Pathol. 2010; 41:503–12.