
INTRODUCTION
Gastric carcinoma (GC) is the fourth most common malignancy and the third leading cause of cancer-related death world-wide [1, 2]. In spite of recent progress in the surgical treatment and chemotherapy, the prognosis of GC patients remains poor [3]. Tumor recurrence including metastasis is the main cause of cancer-related death. Unfortunately, tumor recurrence after radical gastrectomy with curative intent is relatively common, occurring in 20% to 50% of GC patients [4, 5]. At present, the underlying molecular mechanisms responsible for tumor recurrence have not been fully elucidated. The specific tumor markers in detection of tumor recurrence stay elusive. Over the past decades, several cancer stem cells (CSCs) have been identified and characterized in various types of human cancers [6–9]. Because CSCs possess the ability to initiate tumorigenesis, promote progression, and resist conventional chemotherapies [8, 10, 11], the concept that CSCs are responsible for tumor initiation is quite well established. However, the role of CSCs in tumor recurrence remains poorly understood, especially in GC [12, 13].
The cell adhesion molecule, CD44, is a trans-membrane glycoprotein that binds hyaluronic acid, facilitates tumor invasion, and promote tumor recurrence [14–16]. A growing body of evidence suggests that CD44 is also a major cell surface marker of CSCs in several solid tumors, including GC [9, 12, 15–18]. Previous molecular studies show that CD44 is expressed as a standard form (CD44s) and also in numerous variants (CD44v) generated by alternative mRNA splicing [14, 17, 19]. Among CD44 variants, CD44 variant 6 (CD44v6) has been implicated in carcinogenesis, tumor progression, and recurrence in a variety of human cancers [20–25], identified recently as the marker of CSCs in brain tumors, colorectal and bladder carcinomas [21, 26, 27]. Overexpression of CD44v6 found to be an indicator of poor prognosis in hepatocellular carcinoma [28] and GC [29].
Despite the immense clinical importance, molecular mechanisms underlying expression of CD44v6 in GC are unknown. In GC and several other human malignancies in the liver, breast, head and neck, and hematogenetic organs, aberrant activation of signal transducer and activator of transcription 3 (STAT3) has been discovered [30, 31]. STAT3 is critical in inflammation-associated tumorigenesis by regulating numerous oncogenic and inflammatory genes [32], such as interleukin-6 (IL-6), a potent STAT3 activator, and highly expressed in response to hepatitis viral infection and systemic inflammation in the liver [33]. The IL-6/STAT3-signaling pathway has been previously reported to be involved in hepatic inflammation/regeneration, but unknown in GC, in which inflammation is one of the most prominent clinicopathologic characteristics. In this study, we hypothesized that CD44v6 played an important role in GC proliferation, invasion, and migration, and CD44v6 was upregulated via the IL6/STAT3-medicated pathway.
RESULTS
The expression of CD44v6 and pSTAT3 in gastric benign mucosa, premalignant lesion, and carcinoma
By immunohistochemistry, CD44v6 is mainly membranous and p-STAT3 is mainly nuclear. The expression of CD44v6 was significantly progressively increased from minimal immunoreactivity in normal mucosa (0.82±1.006, mean± standard deviation) to moderate in premalignant lesion (2.68±1.887), early GC (2.48±2.129) and marked in advanced GC (4.56±2.912) (p<0.01). A similar immunoreactivity pattern was found with pSTAT3 immunostaining (0.00±0, 0.59±0.805, 0.87±1.254, 0.74±1.209, respectively)(p<0.05). Expression of CD44v6 and pSTAT3 were significantly elevated in different stages of GC, as shown in Figure 1A, B and Supplementary Table 1. The results of Western blotting in 8 paired advanced GC tumors and corresponding adjacent normal tissues confirmed significantly increased immunoreactivity of CD44v6 and pSTAT3 in advanced GC tissues, compared to normal tissue controls (Figure 1C).
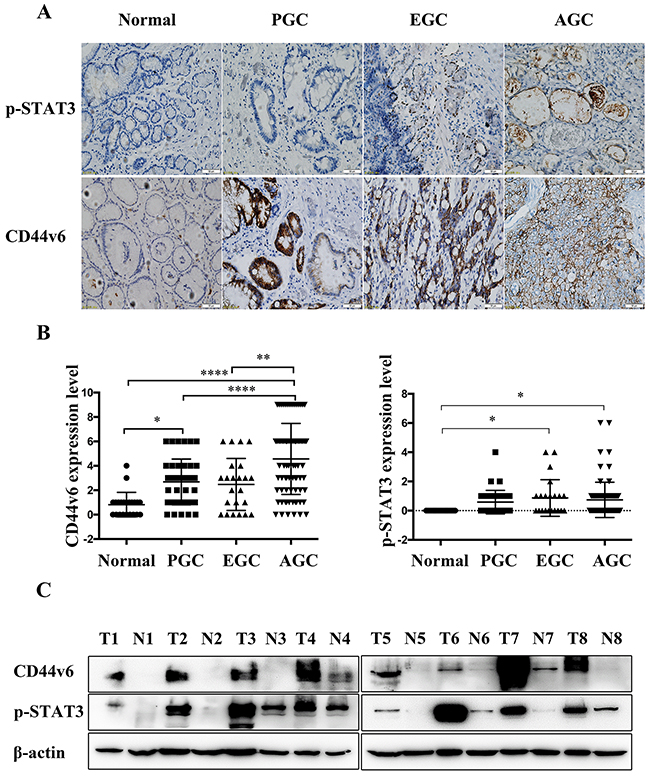
Figure 1: (A) Immunostaining and (B) analysis of the results of CD44v6, and pSTAT3 expression in Normal, precancerous lesions (PGC), early gastric cancer (EGC) and advanced gastric cancer (AGC) tissues (*P<0.05, **P<0.01, ****P<0.005); (C) Western blot analysis of CD44v6 and pSTAT3 expression in 8 paired advanced gastric carcinoma tumors (T1, T2, T3, T4, T5, T6, T7, T8) and corresponding adjacent normal gastric mucosal tissues (N1, N2, N3, N4, N5, N6, N7, N8).
Correlation with clinicopathological characteristics and prognosis
As shown in Table 1, CD44v6 expression in 103 GC tumors was associated with differentiation grade, depth of invasion, nodal metastasis, and TNM stage (p<0.005). However, there was no significant correlation between CD44v6 expression and other pathological parameters, such as patient age, gender, or tumor size. By the Kaplan–Meier method for post-resection survival analysis. GC patients with low expression of CD44v6 had a significantly better outcome than those with high expression of CD44v6 (P<0.05; Figure 2A). However, expression of pSTAT3 showed neither significant correlation with clinicopathology (Supplementary Table 2) nor prognosis (Figure 2B).
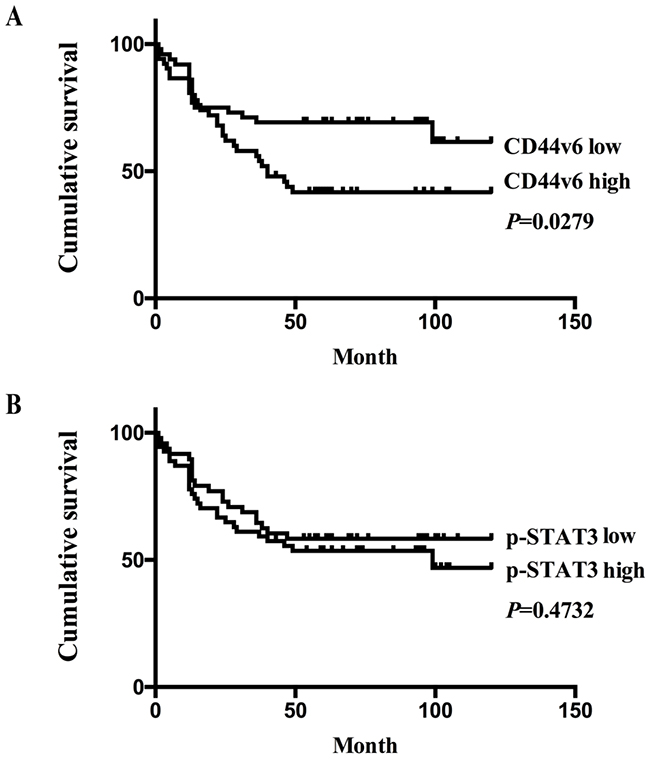
Figure 2: (A) The Kaplan-Meier survival analysis showed significantly poor prognosis in GC patients with high expression of CD44v6,compared to those with low-expression (P<0.05); (B) in contrast, there was no significant survival differences between GC patients with high or low expression of pSTAT3 (P>0.05), on the basis of immunohistochemistry results.
Table 1: The relationship between expression of CD44v6 and clinicopathological features in GC
Parameters |
N |
CD44v6 |
χ2-value |
P value |
|
|---|---|---|---|---|---|
Low(%) |
High(%) |
||||
Gender |
|||||
Male |
69 |
40(58.0) |
29(42.0) |
0.007 |
0.934 |
Female |
34 |
20(58.8) |
14(41.2) |
||
Age |
|||||
<60 |
48 |
25(52.1) |
23(47.9) |
1.407 |
0.236 |
≥60 |
55 |
35(63.6) |
20(36.4) |
||
Size |
|||||
<6.0(cm) |
80 |
50(62.5) |
30(37.5) |
2.658 |
0.103 |
≥6.0(cm) |
23 |
10(43.5) |
13(56.5) |
||
Differentiation |
|||||
Poor |
49 |
21(42.9) |
28(57.1) |
9.109 |
0.003* |
Well/moderate |
54 |
39(72.2) |
15(27.8) |
||
Depth of invasion |
|||||
T1/T2 |
41 |
32(78.0) |
9(22.0) |
10.976 |
0.001* |
T3/T4 |
62 |
28(45.2) |
34(54.8) |
||
Lymph node status |
|||||
No |
42 |
32(76.2) |
10(23.8) |
9.383 |
0.002* |
Yes |
61 |
28(45.9) |
33(54.1) |
||
TNM stage |
|||||
I/II |
51 |
37(72.5) |
14(27.5) |
8.490 |
0.004* |
III/IV |
52 |
23(44.2) |
29(55.8) |
||
Proliferate roles of CD44v6 in gastric cancer cell line
By Western blotting, protein expression of CD44v6 in four GC cell lines was compared with one normal gastric epithelial cell line, GES-1. As exhibited in Supplementary Figure 1, CD44v6 protein expression was significantly higher in 3 GC cell lines, compared to that in control. However, CD44v6 expression was not significantly increased in the N87 GC cell line (Supplementary Figure 1). This inconsistency may be due to the different genetic background of cell lines (Supplementary Table 3).
In order to investigate the functional roles of CD44v6 in GC, we first created two HGC-27 cell lines pre-transfected with CD44v6-siRNA (si#1,si#2) and negative control-siRNA (nc) (Figure 3A). Next, we compared the proliferation of these two cell lines using CCK-8 assay. The growth of cells of decreased CD44v6 is slower than negative-control cells, suggesting CD44v6 is capable of stimulating cell proliferation (Figure 3B). Similar results were observed in colony formation assays as CD44v6-downregulating cells displayed a lower colony-forming ability (Figure 3C).
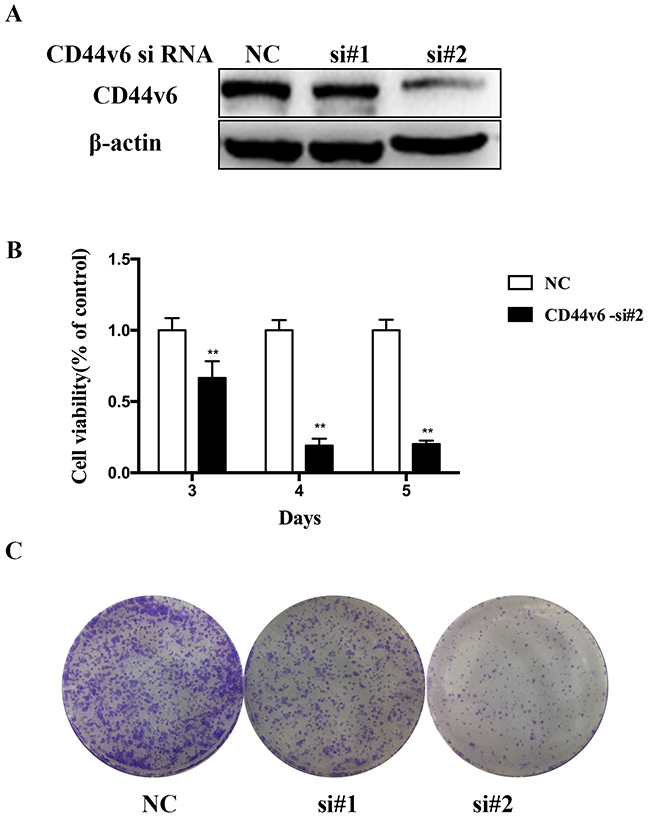
Figure 3: Investigation of the oncogenic functions of CD44v6. HGC-27 cells were transfected with negative-contronl siRNA and CD44v6-siRNA for 3 days. (A) The expression of CD44v6 and β-actin was detected by Western blot. (B) Proliferation of HGC-27 was examined using CCK8 assay. Cell viability was calculated by the following formula: relative cell viability = (absorbance450nm of treated group − absorbance450nm of blank)/(absorbance450nm of control group − absorbance450nm of blank). (C) Colony-forming ability of HGC-27 cell was investigated by colony formation assay (*P <0.05).
Effect of CD44v6 on expression of epithelial-mesenchymal transition molecules, gastric carcinoma cell migration and invasion in vitro
In the GC cell line (HCG-27) after knockdown of CD44v6 with siRNA, decreased expression of CD44v6 was accompanied by downregulation of key epithelial-mesenchumal transition inducers, snail and zeb1, and by upregulation of E-cadherin (Figure 4A). Further investigation in HGC-27 (Figure 4B) and AGS (Figure 4C) cell lines demonstrated significant inhibition of the GC cell migration and invasion capability.
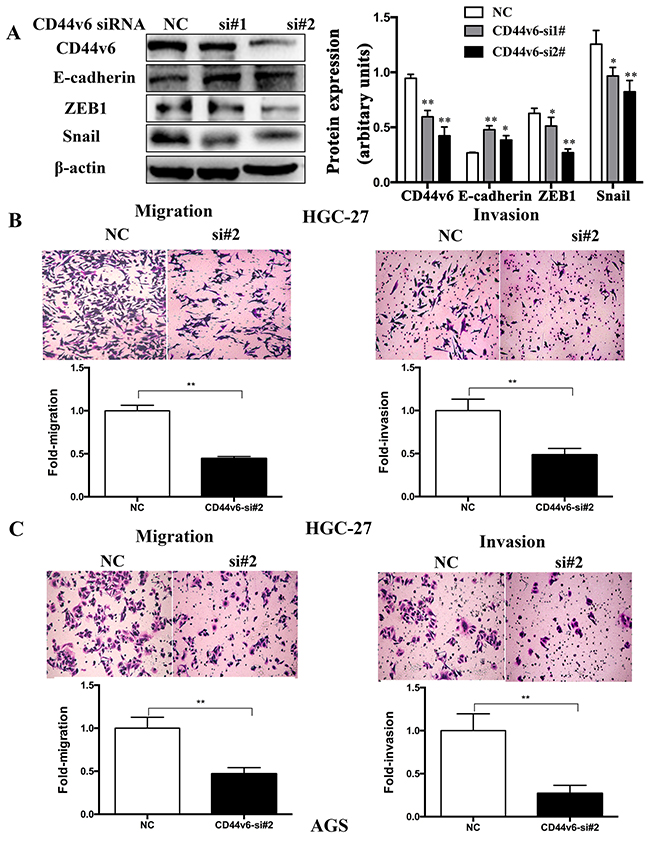
Figure 4: After knockdown of CD44v6 expression with siRNA (si#1,si#2) in the gastric cancer cell line, HGC-27, AGS. (A) In HGC-27 cells, expression of epithelial-mesenchymal transition molecules, ZEB1 and Snail was markedly reduced, but E-cadherin expression increased. The right lane Further cell migration and invasion assays demonstrated conspicuous inhibition of (B) HGC-27 and (C) AGS cells after knockdown of CD44v6, compared to cells transfected with negative-si RNA (NC). The upper panel was the representative result images of cells. The lower panel was the quantitative comparison. NC was used as 1 for fold change calculation (*P <0.05, ** P <0.01).
CD44v6 expression by activation of the IL-6/STAT3 signaling pathway
GC is known to be associated with chronic inflammation, in which IL-6-activated STAT3 expression is an important signaling pathway. To investigate whether pSTAT3 involves in activation of CD44v6 expression, we used the GC cell lines, HGC-27 and AGS with endogenously overexpressed CD44v6, to test if siRNA-mediated knockdown of STAT3 affected CD44v6 expression. As shown in Figure 4, a significantly decreased expression of CD44v6 after STAT3 knockdown was indeed observed (Figure 5A, 5B). Furthermore, functional inactivation of STAT3 by pan-JAK inhibitor AG-490 decreased the expression of CD44v6 (Figure 5C, 5D).
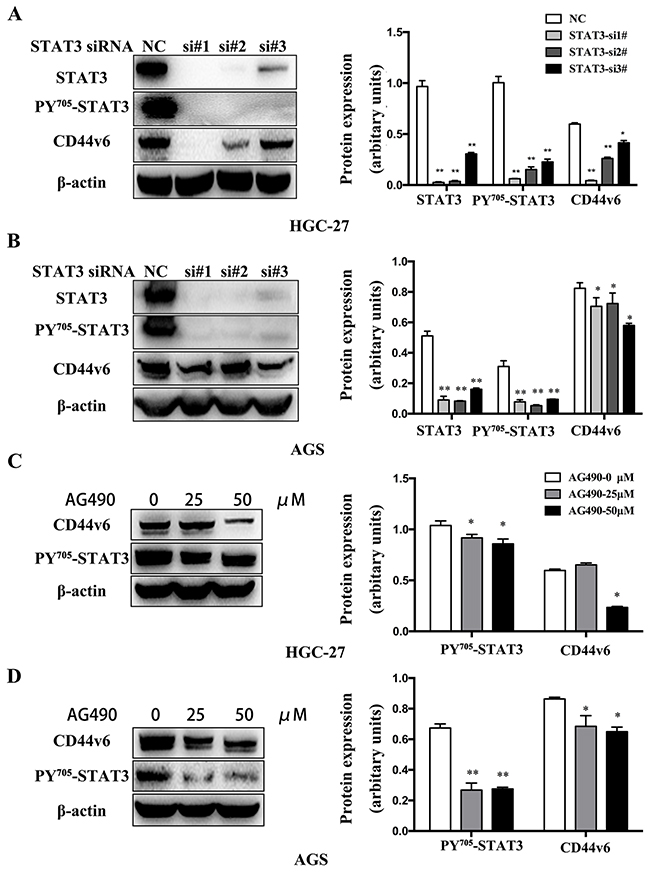
Figure 5: Western blot analysis of CD44v6 expression that was regulated by activation of STAT3 signaling. (A) HGC-27 and (B) AGS cells were treated with STAT3 siRNA (si#1, si#2, si#3) for 3 days to knockdown STAT3 expression. As a result, CD44v6 expression was markedly reduced, compared to normal control cells (nc). In (C) HGC-27 and (D) AGS cells pretreated with AG490 to inactivate pan-JAK signaling to disable STAT3 for 12 hours, expression of CD44v6 was decreased (*P<0.05, ** P <0.01).
Next, we tested whether IL-6 treatment could modulate CD44v6 expression. As shown in Figure 6, IL-6 treatment resulted in not only high expression of CD44v6 but also pSTAT3 in both HGC-27 (Figure 6A) and AGS (Figure 6B) cells. The activation of STAT3 expression by IL-6 stimulation was also confirmed by Western blot in both HGC-27 (Figure 6A) and AGS cells (Figure 6B). Furthermore, a combination treatment with AG490 and IL-6 significantly reduced CD44v6 expression in HGC-27 cells (Figure 6C) but not in AGS cells (Figure 6D), suggesting that pSTAT3 is necessary for IL-6-induced CD44v6 expression in some gastric cancer cells. The difference of AGS and HGC-27 cells may be due to the different sensitivity to AG490 and IL-6. These results strongly imply that proinflammatory signaling can facilitate GC tumorigenesis through the IL-6/STAT3-mediated CD44v6 up-regulation.
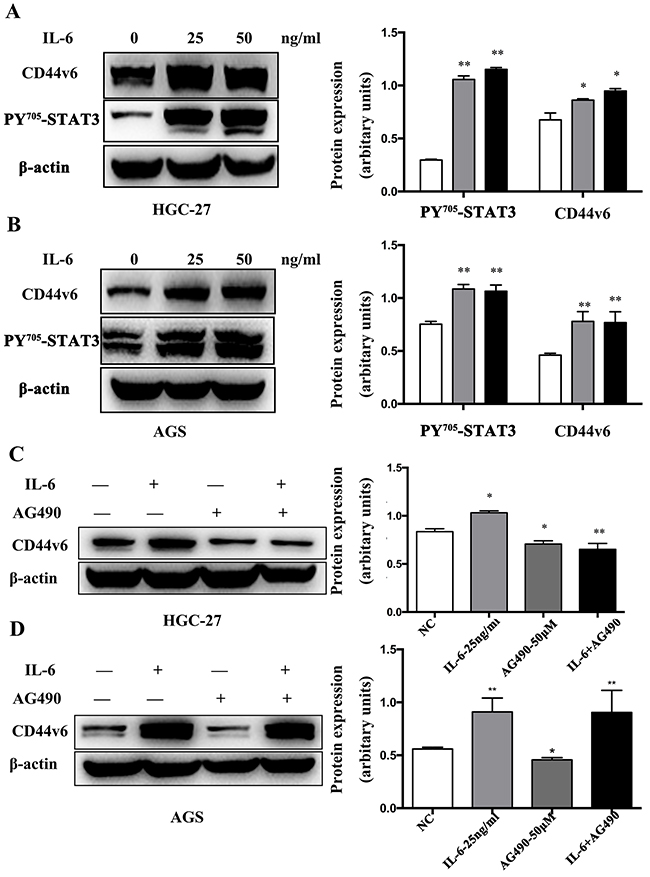
Figure 6: Western blot analysis of IL-6 induced CD44v6 expression through activation of STAT3 signaling. (A) HGC-27 and (B) AGS cells were cultured in serum-free media for 6 hours. Then, cells were incubated with IL-6 for 6 hours, and expression of CD44v6 and p-STAT3 was dose-dependently increased. Pretreatment with AG490 (50μM) for 12 hours in (C) HGC-27 and (D) AGS cells to inactivate STAT3, next cultured in serum-free media for 6 hours and then incubation with IL-6 (50ng/ml) in a serum-free condition for 6 hours markedly reduced expression of CD44v6 (*P <0.05, ** P <0.01).
DISCUSSION
It is well established that the tumor CSC surface marker, CD44v6, is causally involved in cancer metastasis [29], correlated with tumorigenesis in some cancer types. For example, Mikami et al. reported that CD44v6 was overexpressed in extrahepatic bile duct carcinomas and linked to carcinoma differentiation [34]. In GC, Xin et al. first discovered that CD44v6-high expression was associated with poor survival in patients with advanced GC [35]. In 2014, two meta-analysis studies confirmed the association between high CD44v6 expression and worse overall post-resection survival in GC patients [36]. However, most studies have concentrated on advanced carcinomas, including GC. Studies focused on different stages of GC and functions of CSC marker, including CD44v6 and the molecular signaling pathways in tumor microenvironment are lacking.
In the present study, we studied the expression of CD44v6 and pSTAT3 in normal tissues, precancerous lesions, the early and advanced GC and discovered the progressive involvement of CD44v6 in GC tumorigenesis. The differences in CD44v6 expression between normal tissues, precancerous lesions, and advanced GC are statistically significant. In contrast, expression of pSTAT3 is significantly increased only in early and advanced GC tissue, compared to normal controls, as confirmed with both immunohistochemistry and Western blotting. The CD44v6 expression in normal mucosa was very low, but in premalignant lesions (including low and high grade intraepithelial neoplasia), the expression of CD44v6 was increased. Probably due to the premalignant lesions had many similar phenotype with the early GC, the CD44v6 expression was not significantly different between the two groups. However, the CD44v6 expression in advanced GC sharply elevated. This also showed the CD44v6 played a great role on the tumor progression. Importantly, CD44v6 expression is significantly predictive for poor prognosis in GC patients and correlated with poor tumor differentiation, deeper invasion, nodal metastasis, and advanced TNM stage. The findings of this study are compatible with the property of CD44v6 being an important prognosis marker in GC [29].
Despite the significant relevance of enhanced expression of CD44v6 to GC progression, little is known about the mechanisms of how it exerts its oncogenic function in GC. Herein, we investigated the functional roles of CD44v6 in GC cell proliferation, colony-forming ability, migration and invasion. The results are consistent with other two studies [29, 37], except for that CD44v6 did not activate cell migration in the scratch wound-healing assays [29]. The reason for this discrepancy maybe differences in background of GC cell lines and further studies will be needed to elucidate this question. That how CD44v6 involve in the cell proliferation, invasion and migration in GC also need further research.
Despite the studies on the CD44v6 expression, fundamental mechanisms underlying deregulating of CD44v6 are not known. The improved understanding of molecular pathway by which CD44v6 is deregulated in GC will provide useful information for elucidation of stomach CSC origin and development of novel treatment strategies against the deadly disease. IL-6 and its downstream molecules, such as STAT3, play an essential role in inflammation, aberrant immunity, and also carcinogenesis in some carcinomas [38–40]. IL-6 has been shown to enhance invasion of GC cells through sustained activation of STAT3 [31, 41, 42]. IL-6/STAT3-mediated CSC marker, CD133 up-regulation contributes to promotion of liver carcinoma [43]. Herein, we provide additional critical evidence on how IL-6/STAT3 signaling promotes GC invasion by inducing CD44v6 expression. In this study, a significantly decreased expression of CD44v6 after STAT3 knockdown was observed and functional inactivation of STAT3 by the pan-JAK inhibitor, AG-490, decreased the expression level of CD44v6 in both HGC-27 and AGS GC cells in vitro. The results suggest that IL-6 is able to activate expression CD44v6 and STAT3. On the other hand, obviously decreased CD44v6 expression in AG490-treated GC cells in vitro even with the concurrent IL-6 treatment suggests that activation of STAT3 is necessary for IL-6-induced CD44v6 expression. Therefore, our study results demonstrate that IL-6 is critical for induction of CD44v6 expression by STAT3 activation. IL-6 is also elevated in lots of cancers and is a potential regulator of stem cell renewal and proliferation [44–46]. However, the mechanism by which IL-6 regulates CD44v6 expression through STAT3 requires more detailed studies.
Taken together, our study results showed that CD44v6 is an important regulator of GC tumorigenesis, angiogenesis, and survival in an IL-6 mediated, pSTAT3-dependent manner; pSTAT3-mediated CD44v6 up-regulation may represent a promising target molecular signaling pathway for systemic therapy of human GC.
MATERIALS AND METHODS
Patients and tissue samples
One hundrend sixty six patients treated at the Nanjing Drum Tower Hospital in the Jiangsu Province, China, were enrolled over the period from Jan 2006 to Dec 2013, including 80 with GC, 23 with early GC staged at pT1, 41 with premalignant lesions (low and high grade intraepithelial neoplasia) in the gastric mucosa, and 22 normal controls. Patients without enough tissue sample or necessary clinicopathological information, or loss to follow-up were excluded from the study. The paired formalin-fixed paraffin-embedded tissue blocks were retrieved and recut for immunohistochemistry. Proteins were extracted with the conventional methods in fresh frozen matched tumor and non-tumor tissues stored in the Biobank at this hospital. The study protocol was approved by the Medical Ethics Committee of the Nanjing Drum Tower Hospital.
Immunohistochemistry
Immunohistochemical (IHC) analysis for CD44v6, and p-STAT3 expression was performed on formalin-fixed, paraffin-embedded sections of surgical specimens. Briefly, sections were deparaffinized in xylene and rehydrated in gradient ethanol solutions up to distilled water. Endogenous peroxidase activity was blocked by 0.3% H2O2 in methanol for 20 min. The slides were immersed in 10mM citric buffer (pH 6.0) with heating for 15 min for antigen retrieval. Nonspecific binding sites were blocked with 10% normal goat serum for 10 min. Then, sections were incubated in a humidified chamber overnight with CD44v6 and p-STAT3 antibody. Immunostaining was visualized with Diaminobenzidine (DAB) and hematoxylin counterstain. The scoring for CD44v6 and p-STAT3 (expressed at a high level) was based on the area intensity score method (AIS) as previously descibed [49]. The protein expression was scored independently according to the intensity of cellular staining and the proportion of stained tumor cells. The staining intensity was scored as 0 (no staining), 1 (weak staining, light brown), 2 (moderate staining, yellow brown) and 3 (strong staining, brown). The proportions of stained tumor cells were graded as 0 (≤5 % positive cells), 1 (6–25 % positive cells), 2 (26–50 % positive cells) and 3 (≥51 % positive cells). The total scores for intensity and proportion were used to represent the level of protein expression. Positive controls consisted of each staining run and consisted of GCs known to express each of the antigens. Negative controls were normal mouse serum instead of the primary antibody.
Reagents, siRNAs, and antibodies
Anti-CD44v6 (clone: ab78960) and anti-Snail (clone: ab82846) antibodies were purchased from Abcam (Cambridge, UK). Anti-phospho-(Tyr705)-STAT3 (p-STAT3) (clone: 4113; clone: 9131), E-cadherin(clone: 3195s) and ZEB1(clone:3396) antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA). Anti-β-actin (clone: A5441) antibody was from Sigma-Aldrich (St Luis, MO, USA). AG490 were purchased from Selleck Chemicals (Houston, TX, USA). SiRNAs target STAT3 was purchased from Invitrogen (Carlsbad, CA, USA), and CD44v6 as well as a negative control siRNA (sequences are detailed in Supplementary Table 4) were purchased from (RiboBio, GuangZhou, China).
Cell culture and transfection
The human gastric cancer cell lines, AGS and HGC-27, were purchased from the Cell Bank of Chinese Academy of Sciences, and were authenticated by China Center for Type Culture Collection (CCTCC) (Shanghai, China). All cell lines were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium supplemented with 10% fetal calf serum (FBS) in a humidified incubator at 37 °C with 5% CO2. Transfection was carried out using Lipofectamine RNAiMax Reagent (Invitrogen, California, USA) as described elsewhere (reverse transfection method) [47]. In brief, 50 pmol siRNA and 0.5 ml Opti-MEM I Medium (Invitrogen) without serum was mixed in each well of the six-well plate. Then 7.5 μl of Lipofectamine RNAiMAX reagent was added and gently mixed. After incubation for 20min at room temperature, 2ml of cell suspension including 3×105 cells in complete growth medium without antibiotics was added into each well. This generated a final siRNA concentration of 20 nM.
Western blot analysis
The total protein were extracted from target tissues and cells prepared with ice-cold lysis buffer (Biosharp). The supernatant was used for Western blot analysis. Protein concentrations were determined using a BCA protein kit (Beyotime Institute of Biotechnology, China). Samples containing equal amounts of protein were mixed with loading buffer containing 5% 2-mercaptoethanol and then heated for 10 min at 95 °C. Twenty to forty micrograms of protein lysates were separated on 8–12 % sodium dodecyl sulfate-polyacrylamide gels and then transferred to the NC (Nitrocellulose) membranes (Millipore, Bedford, MA, USA). Tris Buffered Saline, with Tween-20 (TBST) containing with 5% nonfat milk or bovine serum albumin was used to block nonspecific binding for 2h at room temperature. Then, the membranes were incubated with the primary antibodies. Subsequently, the membranes were rinsed three times with Tris Buffered Saline (TBS) and 0.1% Tween-20 (TBS-T) for 10min and re-incubated for 1h at room temperature in blocking buffer with each Horseradish Peroxidase (HRP)-conjugated secondary antibody (1:5000), then washed three times for 10min each. Signals generated by enhanced chemiluminescence (Millipore) were recorded with a CCD camera (CLINX, Shanghai, China). Data are representative of at least three independent experiments.
Cell proliferation
Cell viability was detected by Cell Counting Kit (CCK-8) assay. Cells pretreated with siRNA for 2 days were seeded into 96-well plates at 5 × 103 cells/well and then cultured for 24h, 48h, and 72h at 37 °C. After treatment for indicated times, 10 μl CCK-8 solutions were added to each well of the plate. Plates were incubated at 37 °C for 1 h, and then the absorbance at 450 nm was measured. All experiments were carried out in triplicate and repeated three times independently.
Colony formation assays
Colony-formation assay was performed as previously described [48]. In brief, the cell lines were transfected with CD44v6 siRNA (si#1,si#2), negative control-siRNA, for 3 days. Then 1 × 103 cells/well were seeded into six-well plates. After 14 days, the colonies were fixed with methanol and stained with crystal violet. Crystal violet stained colonies were photographed. All experiments were done in triplicate and repeated three times independently.
Cell migration and invasion analysis
Transfilter migration and invasion assays were performed on the HGC-27 and AGS cell lines in serum-free RPMI with 8.0-μm pore inserts on a 24-well Transwell (Corning Costar, Lowell, MA). The HGC-27 cell lines were transfected with CD44v6 siRNA, scRNA, for 3 days and then transferred to the upper chamber of the Transwell coated with 0.5mg/ml collagen type I and Matrigel (BD Bioscience, San Jose, CA) at 1:8 dilution. Migrating and invading cells were quantified after hematoxylin and eosin staining. Migration and invasion assays were performed after transfection, as previously described [50, 51].
Statistical analysis
All statistical analyses were performed by the software “Statistical Package for Social Science” (SPSS) version 22.0 for Windows (SPSS, Chicago, IL, USA) and GraphPad Prism software v.6.01. Differences in expression of CD44v6, and pSTAT3 among advanced and early GC, gastric premalignant lesions, and normal gastric mucosal tissues were compared by the one-way analysis of variance (ANOVA) test. The associations among expression of CD44v6, pSTAT3, and clinicopathological characteristics were analyzed using the Students’ t test or the Chi-square test, where appropriate. The probability of survival was estimated by the Kaplan-Meier method with a log-rank test. All P values were two-sided and considered statistically significant if less than 0.05.
Author contributions
Conception and design: Gui-Fang Xu, Wei-Jie Zhang, Lei Wang
Development of methodology: Gui-Fang Xu, Wei-Jie Zhang, Qin Huang
Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc.): Xiao-Ping Zou, Lei Wang
Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational analysis): Yuan-Yuan Xu, Ming Guo, Liu-Qing Yang, Fan Zhou
Writing, review, and/or revision of the manuscript: Yuan-Yuan Xu, Qin Huang,
Administrative, technical, or material support (i.e., reporting or organizing data, constructing databases): Cao Yu, Aixiu Wang, Tao-Hong Pang, Hong-Yan Wu
Study supervision: Gui-Fang Xu
ACKNOWLEDGMENT
We gratefully thank the staff members in the Department of Gastroenterology and Pathology at the Nanjing Drum Tower Hospital for their suggestions and assistance.
CONFLICTS OF INTEREST
All authors declare no conflicts of interest.
FUNDING
This work was supported by grants from the National Natural Science Foundation of China (Grant Nos. 81201909, 81572338 and 81672380), the Nanjing City Medical Science and Technology Development Program (Nos. YKK12072, YKK15061 and YKK16078). This work was also part of a C-class sponsored research project of the Jiangsu Provincial Six Talent Peaks (WSN-078).
REFERENCES
1. Correia M, Machado JC, Ristimaki A. Basic aspects of gastric cancer. Helicobacter. 2009; 14 Suppl 1:36-40.
2. Lozano R, Naghavi M, Foreman K, Lim S, Shibuya K, Aboyans V, Abraham J, Adair T, Aggarwal R, Ahn SY, Alvarado M, Anderson HR, Anderson LM, et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: a systematic analysis for the Global Burden of Disease Study 2010. Lancet. 2012; 2095-2128.
3. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA: A Cancer Journal for Clinicians. 2015; 65:5-29.
4. Huang KH, Chen JH, Wu CW, Lo SS, Hsieh MC, Li AF, Lui WY. Factors affecting recurrence in node-negative advanced gastric cancer. Journal of gastroenterology and hepatology. 2009; 24:1522-1526.
5. Sakar B, Karagol H, Gumus M, Basaran M, Kaytan E, Argon A, Ustuner Z, Bavbek SE, Bugra D, Aykan FN. Timing of death from tumor recurrence after curative gastrectomy for gastric cancer. American Journal of Clinical Oncology. 2004; 27:205-209.
6. Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001; 414:105-111.
7. Visvader JE, Lindeman GJ. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nature Reviews Cancer. 2008; 8:755-768.
8. Visvader JE, Lindeman GJ. Cancer stem cells: current status and evolving complexities. Cell Stem Cell. 2012; 10:717-728.
9. Zhang S, Balch C, Chan MW, Lai HC, Matei D, Schilder JM, Yan PS, Huang TH, Nephew KP. Identification and characterization of ovarian cancer-initiating cells from primary human tumors. Cancer Research. 2008; 68:4311-4320.
10. Alvero AB, Chen R, Fu HH, Montagna M, Schwartz PE, Rutherford T, Silasi DA, Steffensen KD, Waldstrom M, Visintin I, Mor G. Molecular phenotyping of human ovarian cancer stem cells unravels the mechanisms for repair and chemoresistance. Cell Cycle. 2009; 8:158-166.
11. Kulkarni-Datar K, Orsulic S, Foster R, Rueda BR. Ovarian tumor initiating cell populations persist following paclitaxel and carboplatin chemotherapy treatment in vivo. Cancer Letters. 2013; 339:237-246.
12. Yae T, Tsuchihashi K, Ishimoto T, Motohara T, Yoshikawa M, Yoshida GJ, Wada T, Masuko T, Mogushi K, Tanaka H, Osawa T, Kanki Y, Minami T, et al. Alternative splicing of CD44 mRNA by ESRP1 enhances lung colonization of metastatic cancer cell. Nature Communications. 2012; 3:883.
13. Baccelli I, Trumpp A. The evolving concept of cancer and metastasis stem cells. The Journal of Cell Biology. 2012; 198:281-293.
14. Nagano O, Saya H. Mechanism and biological significance of CD44 cleavage. Cancer Science. 2004; 95:930-935.
15. Orian-Rousseau V. CD44, a therapeutic target for metastasising tumours. European Journal of Cancer. 2010; 46:1271-1277.
16. Zoller M. CD44: can a cancer-initiating cell profit from an abundantly expressed molecule? Nature Reviews Cancer. 2011; 11:254-267.
17. Ishimoto T, Nagano O, Yae T, Tamada M, Motohara T, Oshima H, Oshima M, Ikeda T, Asaba R, Yagi H, Masuko T, Shimizu T, Ishikawa T, et al. CD44 variant regulates redox status in cancer cells by stabilizing the xCT subunit of system xc(-) and thereby promotes tumor growth. Cancer Cell. 2011; 19:387-400.
18. Nagano O, Okazaki S, Saya H. Redox regulation in stem-like cancer cells by CD44 variant isoforms. Oncogene. 2013; 32:5191-5198.
19. Yoshikawa M, Tsuchihashi K, Ishimoto T, Yae T, Motohara T, Sugihara E, Onishi N, Masuko T, Yoshizawa K, Kawashiri S, Mukai M, Asoda S, Kawana H, et al. xCT inhibition depletes CD44v-expressing tumor cells that are resistant to EGFR-targeted therapy in head and neck squamous cell carcinoma. Cancer Research. 2013; 73:1855-1866.
20. Afify AM, Tate S, Durbin-Johnson B, Rocke DM, Konia T. Expression of CD44s and CD44v6 in lung cancer and their correlation with prognostic factors. The International Journal of Biological Markers. 2011; 26:50-57.
21. Jijiwa M, Demir H, Gupta S, Leung C, Joshi K, Orozco N, Huang T, Yildiz VO, Shibahara I, de Jesus JA, Yong WH, Mischel PS, Fernandez S, et al. CD44v6 regulates growth of brain tumor stem cells partially through the AKT-mediated pathway. PLoS One. 2011; 6:e24217.
22. Kamura T, Sakai K, Kaku T, Kobayashi H, Mitsumoto M, Tsuneyoshi M, Nakano H. Comparison of p53 and CD44 variant 6 expression between paired primary and recurrent ovarian cancer: an immunohistochemical analysis. Oncology Reports. 1999; 6:97-101.
23. Seiter S, Arch R, Reber S, Komitowski D, Hofmann M, Ponta H, Herrlich P, Matzku S, Zoller M. Prevention of tumor metastasis formation by anti-variant CD44. The Journal of Experimental Medicine. 1993; 177:443-455.
24. Shi J, Zhou Z, Di W, Li N. Correlation of CD44v6 expression with ovarian cancer progression and recurrence. BMC Cancer. 2013; 13:182.
25. Sliutz G, Tempfer C, Winkler S, Kohlberger P, Reinthaller A, Kainz C. Immunohistochemical and serological evaluation of CD44 splice variants in human ovarian cancer. British Journal of Cancer. 1995; 72:1494-1497.
26. Todaro M, Gaggianesi M, Catalano V, Benfante A, Iovino F, Biffoni M, Apuzzo T, Sperduti I, Volpe S, Cocorullo G, Gulotta G, Dieli F, De Maria R, Stassi G. CD44v6 is a marker of constitutive and reprogrammed cancer stem cells driving colon cancer metastasis. Cell Stem Cell. 2014; 14:342-356.
27. Yang YM, Chang JW. Bladder cancer initiating cells (BCICs) are among EMA-CD44v6+ subset: novel methods for isolating undetermined cancer stem (initiating) cells. Cancer Investigation. 2008; 26:725-733.
28. Endo K, Terada T. Protein expression of CD44 (standard and variant isoforms) in hepatocellular carcinoma: relationships with tumor grade, clinicopathologic parameters, p53 expression, and patient survival. Journal of Hepatology. 2000; 32:78-84.
29. Xie JW, Chen PC, Zheng CH, Li P, Wang JB, Lin JX, Lu J, Chen QY, Cao LL, Lin M, Lin Y, Huang CM. Evaluation of the prognostic value and functional roles of CD44v6 in gastric cancer. Journal of Cancer Research And Clinical Oncology. 2015; 141:1809-1817.
30. Yu H, Jove R. The STATs of cancer--new molecular targets come of age. Nature Reviews Cancer. 2004; 4:97-105.
31. Wang Z, Si X, Xu A, Meng X, Gao S, Qi Y, Zhu L, Li T, Li W, Dong L. Activation of STAT3 in human gastric cancer cells via interleukin (IL)-6-type cytokine signaling correlates with clinical implications. PLoS One. 2013; 8:e75788.
32. Turkson J, Jove R. STAT proteins: novel molecular targets for cancer drug discovery. Oncogene. 2000; 19:6613-6626.
33. Wang C, Yang W, Yan HX, Luo T, Zhang J, Tang L, Wu FQ, Zhang HL, Yu LX, Zheng LY, Li YQ, Dong W, He YQ, et al. Hepatitis B virus X (HBx) induces tumorigenicity of hepatic progenitor cells in 3,5-diethoxycarbonyl-1,4-dihydrocollidine-treated HBx transgenic mice. Hepatology. 2012; 55:108-120.
34. Mikami T, Saegusa M, Mitomi H, Yanagisawa N, Ichinoe M, Okayasu I. Significant correlations of E-cadherin, catenin, and CD44 variant form expression with carcinoma cell differentiation and prognosis of extrahepatic bile duct carcinomas. American Journal of Clinical Pathology. 2001; 116:369-376.
35. Xin Y, Grace A, Gallagher MM, Curran BT, Leader MB, Kay EW. CD44V6 in gastric carcinoma: a marker of tumor progression. Appl Immunohistochem Mol Morphol. 2001; 9:138-142.
36. Chen J, Li T, Liu Q, Jiao H, Yang W, Liu X, Huo Z. Clinical and prognostic significance of HIF-1alpha, PTEN, CD44v6, and survivin for gastric cancer: a meta-analysis.PLoS One. 2014; 9:e91842.
37. Eom BW, Joo J, Park B, Jo MJ, Choi SH, Cho SJ, Ryu KW, Kim YW, Kook MC. Nomogram incorporating CD44v6 and clinicopathological factors to predict lymph node metastasis for early gastric cancer. PLoS One. 2016; 11:e0159424.
38. Hodge DR, Hurt EM, Farrar WL. The role of IL-6 and STAT3 in inflammation and cancer. European Journal of Cancer. 2005; 41:2502-2512.
39. Bowman T, Garcia R, Turkson J, Jove R. STATs in oncogenesis. Oncogene. 2000; 19:2474-2488.
40. Huang S. Regulation of metastases by signal transducer and activator of transcription 3 signaling pathway: clinical implications. Clin Cancer Res. 2007; 13:1362-1366.
41. Lin MT, Lin BR, Chang CC, Chu CY, Su HJ, Chen ST, Jeng YM, Kuo ML. IL-6 induces AGS gastric cancer cell invasion via activation of the c-Src/RhoA/ROCK signaling pathway. International Journal of Cancer. 2007; 120:2600-2608.
42. Kinoshita H, Hirata Y, Nakagawa H, Sakamoto K, Hayakawa Y, Takahashi R, Nakata W, Sakitani K, Serizawa T, Hikiba Y, Akanuma M, Shibata W, Maeda S, Koike K. Interleukin-6 mediates epithelial-stromal interactions and promotes gastric tumorigenesis. PLoS One. 2013; 8:e60914.
43. Won C, Kim BH, Yi EH, Choi KJ, Kim EK, Jeong JM, Lee JH, Jang JJ, Yoon JH, Jeong WI, Park IC, Kim TW, Bae SS, et al. Signal transducer and activator of transcription 3-mediated CD133 up-regulation contributes to promotion of hepatocellular carcinoma. Hepatology. 2015; 62:1160-1173.
44. Grivennikov S, Karin E, Terzic J, Mucida D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H, Eckmann L, Karin M. IL-6 and Stat3 are required for survival of intestinal epithelial cells and development of colitis-associated cancer. Cancer Cell. 2009; 15:103-113.
45. Schafer ZT, Brugge JS. IL-6 involvement in epithelial cancers. The Journal of Clinical Investigation. 2007; 117:3660-3663.
46. Lin L, Amin R, Gallicano GI, Glasgow E, Jogunoori W, Jessup JM, Zasloff M, Marshall JL, Shetty K, Johnson L, Mishra L, He AR. The STAT3 inhibitor NSC 74859 is effective in hepatocellular cancers with disrupted TGF-beta signaling. Oncogene. 2009; 28:961-972.
47. Sousa JF, Nam KT, Petersen CP, Lee HJ, Yang HK, Kim WH, Goldenring JR. miR-30-HNF4gamma and miR-194-NR2F2 regulatory networks contribute to the upregulation of metaplasia markers in the stomach. Gut. 2016; 65:914-924.
48. Zhang B, Yang Y, Shi X, Liao W, Chen M, Cheng AS, Yan H, Fang C, Zhang S, Xu G, Shen S, Huang S, Chen G, et al. Proton pump inhibitor pantoprazole abrogates adriamycin-resistant gastric cancer cell invasiveness via suppression of Akt/GSK-beta/beta-catenin signaling and epithelial-mesenchymal transition. Cancer Letters. 2015; 356:704-712.
49. Kurzen H, Münzing I, Hartschuh W. Expression of desmosomal proteins in squamous cell carcinomas of the skin. J Cutan Pathol. 2003;30:621-30.