
INTRODUCTION
The immune system plays an important role in the development and progression of cancer [1]. Immune cells, including T lymphocytes, macrophages, mast cells, and neutrophils present in the tumor microenvironment can either inhibit or enhance tumor growth. Little is known about the impact of B cells on tumor biology. The presence of B cells in human tumors has long been overlooked since the prevailing notion was that antitumor immunity is primarily mediated by T cells and NK cells. Since B cells were solely viewed as antibody producers and antibodies were believed to play a negligible role in tumor immunity their relevance in cancer biology has been ignored. In recent years, it has been demonstrated that B cells do also play an important role in tumor immunology [2]. However, the contribution of B cells to tumor immunology appears to be complex and entails both protumorigenic and antitumor effects. Experimental models have yielded important insights into the mechanism by which B cells affect tumor immunity. Besides antibody-mediated effects, antibody-independent mechanisms such as antigen-presentation [3], cytokine production [4], direct cytotoxicity [5] and indirect effects through modulation of other immune cells have been implicated to be of importance [6]. Whether B cells promote or inhibit tumor growth seems to depend on a number of variables such as temporal and spatial setting as well as on the composition of B-cell subsets. The findings in murine tumor models raised renewed interest in studying the B-cell infiltrate in human tumor samples and its potential impact on the tumor microenvironment. Indeed, B-cell infiltrates can be found in many different human tumor entities, including breast cancer [7], lung cancer [8], ovarian cancer [9], colorectal cancer [10] and germ cell tumors [11].
The multitude of B-cell-directed agents which are on the market or in development, predominantly for the treatment of autoimmune diseases and B-cell malignancies, offer the perspective that insights into the role of B cells in human tumor biology can be rapidly translated into clinical interventions. A more detailed understanding of tumor-associated B-cell subsets and their effects on tumor growth is therefore crucial and will facilitate the therapeutic manipulation of the B-cell compartment with the aim of enhancing tumor immunity.
Since most studies to date used immunohistochemistry on paraffin-embedded tissues they could only assess a limited number of markers and an identification of specific B-cell subsets, which are defined by coexpression of multiple markers, was not possible. We thus set out to perform a comprehensive flow cytometric characterization of tumor-associated B cells in peripheral blood and fresh tumor samples of patients with colorectal cancer.
RESULTS
IgD-CD27+ memory B cells are increased in peripheral blood of CRC patients
We assessed the composition of the B-cell populations in peripheral blood of 46 cancer patients and compared it to 10 age- and sex-matched healthy controls. The clinical characteristics of the patients are summarized in table 1 and the pathologic features are listed in supplementary table 1. The percentage of CD19+ B cells among CD45+ lymphocytes in the peripheral blood of colorectal cancer patients did not differ significantly from healthy controls (8.4% vs. 5.1%, p=0.14, Fig. 1A). Although the percentage of total circulating B cells was similar, the composition of the B cell subsets showed large differences. The percentage of memory B cells (defined as IgD-CD27+) in peripheral blood of CRC patients was more than twice as high (19.3% vs. 8%, p<0.005, Fig. 1C), suggesting an impact of CRC on B-cell homeostasis. Conversely, CD45+ PBMCs of healthy donors contained more virgin naïve (IgD+CD27-CD38-) B cells (p<0.05, Fig. 1D). An analysis of the correlation of B cells and B-cell subsets in peripheral blood of CRC patients with the UICC-tumor-node-metastasis-classification revealed similar compositions throughout the stages (data not shown). As shown in figure 1F and 1G there is a strong colocalisation of CD20 and Ki67 in sequential immunohistochemical stainings, suggesting a local proliferation of tumor associated B cells. This characteristic distribution of CD20 and Ki67 with strong Ki67-positivity especially in germinal centers of peritumorous tertiary lymphoid structures could be confirmed in >90% of the analyzed samples.
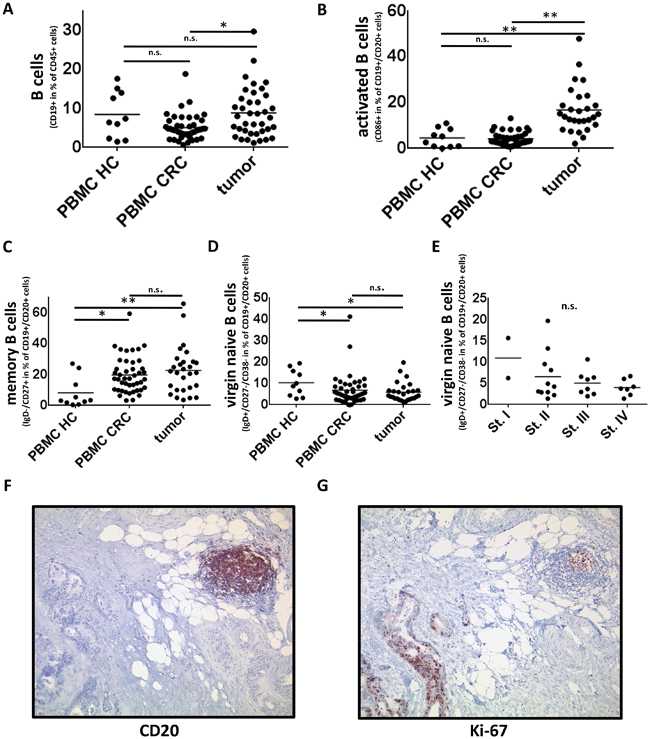
Figure 1: B cells and B-cell subsets in PBMC and tumor samples of CRC patients and PBMC of healthy controls. (A) Flow cytometric analyzes of CD19+ cells in PBMCs of healthy controls (n=10), PBMCs of CRC patients (n=46) and single cell suspensions of tumor samples (n=38). The percentage of CD19+ cells in tumor samples is significantly higher than in PBMC of CRC patients (8.9% vs. 5.1% of CD45+ lymphocytic cells, p<0,05). (B) Tumor associated B cells (n=28) contain a higher percentage of activated B cells (defined by positivity for CD19, CD20 and CD86) than PBMCs of CRC patients (n=44) or healthy controls (n=10) (16.8% vs. 4.1/4.5% respectively, p<0,005). (C) CD27+IgD- (memory) B cells are elevated in PBMCs (n=44) and tumor (n=28) of colorectal cancer patients compared to PBMCs of healthy controls (n=10) (19.3/22.7% vs. 8.2%, p<0,05/<0,005 respectively). (D) According to an increase in memory B cells virgin naïve (IgD+/CD27-/CD38-) B cells are decreased in PBMCs (n=44) and tumor samples (n=28) of CRC patients compared to PBMC of healthy controls (n=10) (p<0,05) (E) The percentage of IgD+/CD27-/CD38- naive B cells decreases with UICC stage (n=28, n.s.). (F) Exemplary immunohistochemical staining of CD20 to visualize the intratumoral distribution of tumor-associated B cells, predominantly in peritumorous lymphoid follicles. (G) Immunohistochemical analyzes of Ki-67 in CD20+ cells revealed a strong local proliferation of B cells within tertiary lympoid structures.
Table 1: Patient characteristics
CRC patients (n=51) |
Age |
70 (±11) |
|
Sex |
32 (61%) male |
19 (39%) female |
|
UICC Stage |
St. I |
2 (4%) |
|
St. II |
17 (34%) |
||
St. III |
14 (27%) |
||
St. IV |
18 (35%) |
||
Grading (n.d. in 4 Metastases) |
G 1 |
0 |
|
G2 |
41 |
||
G3 |
6 |
||
Primary site (RCTx) |
34 Colon (0) |
17 Rectum (1) |
|
Healthy donors (n=10) |
Age |
65 (±11) |
|
sex |
4 (40%) male |
6 (60%) female |
|
Tumor-associated B cells are activated and of a memory phenotype
Single cell suspensions of tumor tissue of 38 CRC patients were produced within 12 hours after surgery. Subsequently we analyzed the tumor-infiltrating lymphocytes by 10-color flow cytometry. CD19+ cells could be detected in all 38 tumor samples and on average made up 8.9% of the CD45+ lymphocytic infiltrate of colorectal cancers compared to 5.1% of the CD45+ PBMCs of CRC patients (p<0,005, Fig. 1A). Of note, about one third (31.7%) of the tumor-infiltrating CD19+ cells were CD19+CD20-CD38high plasma cells (Fig. 2A). CD3+ T cells were also detectable and made up 68% of the CD45+ lymphocytic infiltrate (data not shown). A well-defined population of cells co-expressing CD19+ and CD20+ could be detected in 28 out of 38 tumor samples with a non-significant increase in metastatic disease (8.3% in stage IV vs. 6.4% in stage I-III, p=0.28). In further analyses of these samples tumor-associated B cells have shown several differences to B cells in the peripheral blood of CRC patients and healthy controls. Tumor samples revealed a higher percentage of activated B cells (characterized by expression of CD19, CD20 and CD86) than PBMCs of CRC patients (16.8% vs. 4.1%, p <0,005, Fig. 1B) or healthy controls (16,8% vs. 4,5%, p<0,005, Fig. 1B). To assess the maturity of B cells in the analyzed samples virgin naïve (CD19+CD20+IgD+CD27-CD38-), virgin activated (CD19+CD20+IgD+CD27+ CD38-) and memory B cells (CD19+CD20+IgD-CD27+) were quantified. Whereas memory B cells were relatively rare in peripheral blood of healthy controls (8.2%) they made up 22.7% of CD19+CD20+ cells in tumor samples (p<0,005, Fig.1C). The percentage of memory B cells was similar across all UICC stages (data not shown). According to these results virgin naive B cells were higher in PBMCs of healthy controls compared to PBMCs and tumor tissue of CRC patients (10.2% vs. 6.8%, 6.3% respectively, p<0.05; Fig. 1D). The percentage of virgin naïve B cells in tumor samples showed a tendency to decrease with higher UICC stages (Fig. 1E, n.s.). Virgin activated B cells were hardly detectable in tumor samples and their percentage was similar in PBMCs of tumor patients and healthy controls (data not shown).
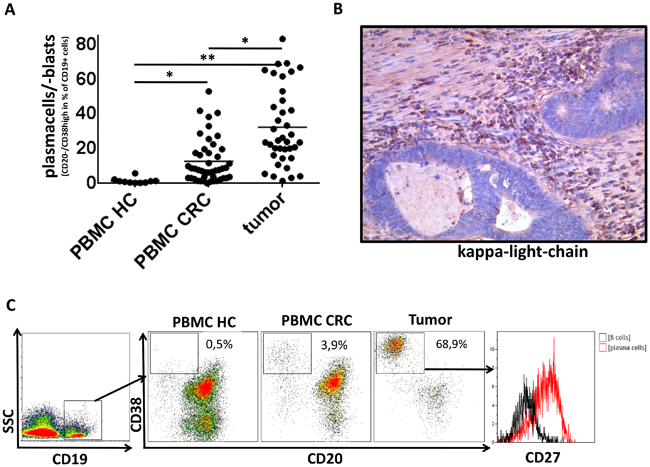
Figure 2: Plasma cells in PBMC and tumor samples of CRC patients and PBMC of healthy controls. (A) Plasma cells (CD19+CD20-CD38high) are strongly elevated in PBMCs (n=46) and tumor tissue (n=38) of CRC patients compared to healthy controls (n=10) (12.6%/31.7% vs. 1.4% respectively; p<0,05/<0,005) (B) Immunohistochemistry showing an infiltration of colorectal cancer by IgG-kappa+ cells into the same tumor sample as shown in C. (C) Exemplary density plots showing CD45+CD19+CD20-CD38high cells in peripheral blood and tumor tissue of CRC patients and peripheral blood of healthy controls. The plasma cell phenotype of these cells was confirmed by expression of CD27.
Plasma cells are elevated in peripheral blood and tumor tissue of colorectal cancer patients
Analysis of plasma cells revealed marked differences between cancer patients and healthy controls. Plasma cells were rarely detectable in PBMCs of healthy donors (n=10) but made up a significant part of CD19+ lymphocytes in peripheral blood (n=46) and tumor tissue (n=38) of colorectal cancer patients. The percentage of CD19+CD20-CD38high cells was 1.4% of CD19+ lymphocytes in PBMCs of healthy controls compared to 12.6% in PBMCs and 31.7% in the tumor tissue of CRC patients (p<0,005, Fig.2A). The plasma cell phenotype was confirmed by a strong expression of CD27 in CD19+CD20-CD38high cells (Fig. 2C). Additionally, the results obtained by flow cytometry were confirmed by immunohistochemical staining for IgG-kappa (Fig. 2B). Nephelometry of IgG subclasses in 11 accessible serum samples of CRC patients revealed a normal distribution of IgG subclasses (IgG1>IgG2>IgG3>IgG4) in 9/11 cases (data not shown).
B-cell subsets with a regulatory phenotype (CD24highCD38high and CD24highCD27+) are detectable in tumor samples and peripheral blood of CRC patients
A regulatory function has been described for transitional B cells (CD19+CD20+CD24highCD38high, Fig. 3A). Our analysis of this subset revealed no difference between PBMCs of CRC patients and healthy donors. Although the frequency of transitional B cells in tumor tissue was lower than that in peripheral blood of CRC patients (1.2% vs. 3.5%, p<0,005) transitional B cells were detectable in tumor samples and their percentage was increased in advanced stage disease (0.7% of B cells in Stage I+II vs. 1,8% in Stage III+IV, p<0,05, Fig.3B). The CD24highCD27+ B cell population has recently been described as another regulatory B cell subset [12]. This population was regularly detected within tumor-associated B cells and PBMCs of CRC patients and was significantly elevated compared to PBMCs of healthy controls (21.1%/18.5% vs. 11.3%, p<0,05, Fig. 3C,D). The percentage of CD24highCD27+ B cells in tumor samples did not correlate with tumor stage (data not shown).
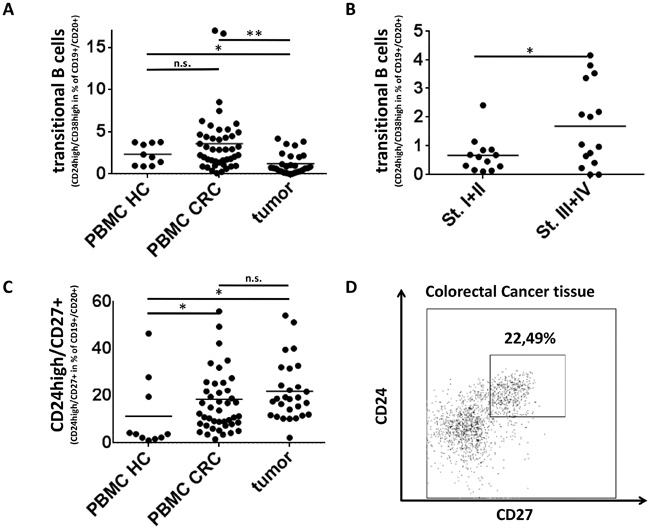
Figure 3: Regulatory B-cell subsets in Colorectal Cancer. (A) The percentage of CD24highCD38high (transitional) B cells is lower in tumor samples (n=28) than in peripheral blood of CRC patients (n=44) or healthy controls (n=10) (1.2% vs. 3.5%/2,3%, p<0,005/p<0,05 respectively). (B) Whereas transitional B cells are very rare in early stage CRC the frequency increases in advanced stage disease (n=28) (0.7% of B cells in Stage I+II vs. 1.8% in Stage III+IV, p<0,05) (C) CD24highCD27+ B cells make up a high percentage of tumor associated B cells in tumor samples (n=28) and PBMC of colorectal cancer (n=44) and are significantly elevated compared to PBMC of healthy controls (n=10) (21.1%/18.5% vs. 11.3%, p<0,05). (D) Exemplary density plot showing CD24highCD27+ B cells in a tumor sample from a patient with colorectal cancer (gated on lymphocytic FSC/SSC,CD45+,CD19+,CD20+).
Characterization of B cell subsets in colorectal liver metastases
To assess differences between infiltrating lymphocytes in metastatic tumor tissue and samples of the primary tumor of colorectal cancer patients, biopsy specimens of six colorectal liver metastases were processed and stained analogous to the primary tumors. Our analyses showed a lower percentage of CD19+CD20+ cells in metastatic samples than in primary tumors (2% vs. 6,9% of CD45+ lymphocytic cells, p<0,05, Fig. 4A). Analyses of the state of activation and maturity revealed a lower percentage of memory B cells (IgD-CD27+) in metastatic tissue than in primary tumors (23.4% vs. 9.9% of CD45+CD19+CD20+ cells, p<0,05, Fig. 4B). Whereas most primary tumor samples contained a distinct plasma cell infiltrate, CD19+CD20-CD38high cells were very rare in metastatic samples (31.7% vs. 3.7% of CD45+CD19+ cells, p<0,005, Fig. 4C). Interestingly CD24highCD38high “transitional” B cells, which represent a regulatory B cell subset, were of low frequency in the primary tumors, but a significant fraction of the metastatic tissue (1,1% vs. 6,3% of CD19+CD20+ cells, p<0,05, Fig. 4D).
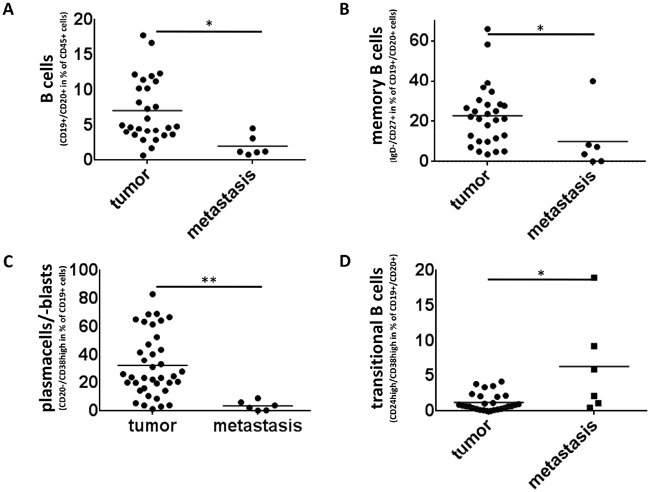
Figure 4: Memory B cells and cells with a plasma cell phenotype are significantly reduced in colorectal liver metastasis. (A) CD19+CD20+ cells make up only 2% of CD45+ lymphocytic cells in metastatic tissue (n=6) compared to 6.9% in samples of primary colorectal cancer (n=28) (p<0,05) (B) The percentage of IgD-/CD27+ memory b cells in colorectal liver metastasis (n=6) is significantly lower than in primary tumors (n=28) (9.9% vs. 23.4%, p<0,05) (C) Whereas cells with a plasma cell phenotype were regularly detectable in tumor samples of CRC patients (n=38) they were heavily reduced in metastatic Tissue (n=6) (3.7% compared to 31.7%, p<0,005). (D) B cells with a regulatory phenotype (CD24highCD38high B cells) were rare in the primary tumors, but well detectable in metastatic tissue (1,1% vs. 6,3% of CD19+CD20+ cells, p<0,05).
Tumor-infiltrating T lymphocytes are mainly CD45-CCR7- effector/memory T cells and CD3+CD4+CD25+CD127low regulatory T cells are significantly elevated in tumor samples
Whereas naive T cells (CD45RA+CCR7+) make up about 25% of T cells in the peripheral blood of CRC patients and healthy controls their percentage in tumor samples is less than 10%. Tumor-infiltrating T cells are predominantly antigen experienced CD45-CCR7- effector/memory T cells (74.7% of CD45+CD3+ lymphocytic cells in tumor tissue compared to 30.2% and 26% in PBMCs of CRC patients and healthy controls respectively, p<0,005, Fig. 5A). Of note, the percentage of effector/memory T cells decreases in metastatic disease (Fig. 5B). In order to assess regulatory T cells we analyzed the CD3+CD4+CD25+CD127low T cell subset, which is known to contain a high percentage of FoxP3+ Treg. CD3+CD4+CD25+CD127low T cells made up 15.5% of CD4+-T cells in tumor samples compared to 7.8% and 5.0% of PBMCs of CRC patients or healthy controls respectively (p<0,005, Fig. 5C). Since regulatory B cells have been proposed to expand regulatory T cells we investigated whether the amount of regulatory B cells and regulatory T cells in the tumor correlated [13,14]. Pearson’s test revealed no significant correlation between CD3+CD4+CD25+CD127low regulatory T cells and either the CD24highCD38high (r=0,283) or the CD24highCD27+ (r=0,183) B-cell subsets. Cytotoxic CD8+ T lymphocytes were present both within and around the tumor while B cells clustered primarily at the invasive margins (Fig. 5D,E). The B cells frequently formed follicle-like structures along the tumor border.
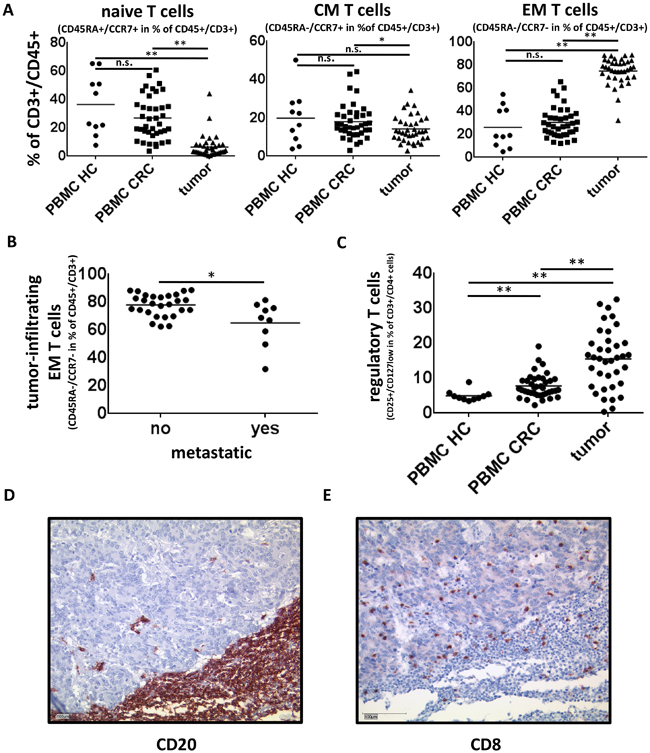
Figure 5: Tumor infiltrating T cells in colorectal cancer. (A) Tumor infiltrating T cells are mainly of a CD45RA-CCR7- effector/memory (EM) phenotype (n=37) whereas PBMC of CRC patients (n=38) and healthy controls (n=10) contain predominantly naive (CD45RA+CCR7+) and central memory (CM) (CD45RA-CCR7+) T cells. (B) The percentage of CD45RA-CCR7- effector memory T cells in primary tumor samples is significantly reduced in Stage IV colorectal cancer (65% vs. 77,9%, p<0,05) (C) CD3+CD4+CD25+CD127low T cells which are known to contain a high percentage of FoxP3+ regulatory T cells are significantly elevated in tumor samples (n=38) compared to PBMC of CRC patients (n=39) and healthy controls (n=10) (15.5% of CD4+-Tcells in tumor samples compared to 7.8% and 5.0% respectively, p<0,005) (D,E) Immunohistochemical staining of CD20 and CD8 were performed to illustrate the intratumoral distribution of B and T cells. B cells were detectable throughout the whole tumor samples, but were predominantly localized in peritumoral tertiary lymphoid structures. Figure E highlights the infiltration of Colorectal Cancer by cytotoxic T cells and also demonstrates a colocalisation of cytotoxic T and B cells especially within the tertiary lymphoid structures. Sequential immunohistochemistries of CD8+-T cells and CD20+-B cells revealed similar colocalisations in 40% of the analyzed samples.
DISCUSSION
The lymphocytic infiltrate of tumors is known to have an important impact on the prognosis of malignant diseases [15–17]. In colorectal cancers a classification based on the immune infiltrate seems to be superior to conventional tumor staging [18]. Although this and other studies focused on T cells, B cells have also been shown to be an important part of the immune cell infiltrate of many tumor entities [2]. However, the role of B cells in tumor development and immunosurveillance is complex [19]. On the one hand, data from animal models suggest that B cells seem to support tumor development and survival. Murine tumor models have provided crucial information about the underlying mechanisms of B-cell mediated effects on tumor biology. Antibodies produced by B lymphocytes participate in carcinogenesis and tumor progression by promoting chronic inflammation [20]. Furthermore, regulatory B cells have been shown to suppress T cell-mediated antitumor responses [21–23]. In line with these results depletion of B cells enhances T cell immunity in response to cancer vaccines [24,25]. In addition to promoting the development and growth of tumors, B lymphocytes can also enhance cancer metastasis [13,26,27].
On the other hand, studies have shown that tumor-associated B cells also contribute to cancer immunosurveillance. B cells are required for optimal CD4 and CD8 T cell tumor immunity [3]. Conversely, depletion of B cells in this setting promotes tumor progression. In addition, B-cell mediated antitumor immunity can also suppress cancer metastasis [28]. Immunohistochemical studies of human cancers found that infiltration of the primary tumor with B cells correlate with a better prognosis and lower rate of metastasis [29,30]. In addition, B-cell based cancer immunotherapy in animal models has yielded promising results and demonstrates that B cells can induce T-cell mediated antitumor immunity [31–34]. How B cells exert their antitumor effects is not precisely known. In some settings the antitumor activity was primarily antibody-mediated [35] whereas other studies suggested a crucial role for antigen-presentation by B cells [36]. Interestingly, a recent retrospective analysis of lymphoma patients receiving high-dose chemotherapy with subsequent autologous transplantation showed that the addition of the B-cell depleting monoclonal anti-CD20 antibody, rituximab, to a high-dose chemotherapy regimen resulted in a significantly higher incidence of solid tumors from 4.90% to 13.26% after ten years of follow-up, suggesting that in humans B cells contribute to tumor immunosurveillance [37].
The heterogeneity of tumor-infiltrating B-cell subsets could provide an explanation as to why studies on B cell-depletion in cancer found both antitumorigenic or protumorigenic effects [38,39]. Understanding the composition of B cell-infiltrates in human tumors is therefore crucial for the effective treatment of cancer patients with B cell-targeted therapies. Several studies have investigated tumor-infiltrating B cells in human malignancies using immunohistochemistry or transcriptomic analysis [9,40,41]. They found that a B-cell infiltrate is present in a wide variety of human tumor entities and that the presence of B cells correlates with survival. Due to the technical limitations of immunohistochemistry and gene expression analysis these studies could not provide a detailed characterization of the different subsets within tumor-associated B cells. We thus used 10-color flow cytometry to perform a more in depth phenotypic analysis of B cells from peripheral blood and tumor samples of patients with colorectal cancer. Using flow cytometry we could identify several B-cell subsets, which are defined by up to five different markers (e.g. transitional B cells (CD45+,CD19+,CD20+,CD24high,CD38high). The use of flow cytometry to study the composition of the intratumoral B-cell compartment enabled us to gain more detailed information on B cell subtypes than would be feasible by immunohistochemical methods.
Since the importance of the immune response has been well established in colorectal cancer this entity appears to be an attractive model to study the impact of B cells on the clinical course of solid cancers in humans. [18]. A small case series reported a beneficial effect of B-cell depletion in patients with colorectal cancer suggesting that therapeutic targeting of B cells could be a promising treatment approach in CRC patients [38]. Many solid tumors induce a disturbance of the B cell compartment. In a recent study which investigated B cells in the peripheral blood of cancer patients, cancer progression lead to a disturbance of B-cell homeostasis and resulted in a loss of memory B cells as well as an increase in plasmablasts in the peripheral blood of cancer patients [42]. Regarding peripheral blood, our results were similar with a remarkable increase of cells with a CD19+CD20-CD38highCD27+ plasma cell phenotype. Of note, we could also detect the same population within colorectal cancer tissues, confirming immunohistochemical studies showing a pronounced plasma cell infiltration in colorectal cancer [43] and other tumor entities [9,44,45]. In colorectal cancer a plasma cell infiltrate is associated with favorable outcome [43,46]. Furthermore, several studies yielded indirect evidence that tumor-infiltrating B cells in some solid tumors contain a high proportion of tumor antigen-specific B cells. Analysis of the immunoglobulin genes from tumor-infiltrating B cells revealed a restricted repertoire with a high frequency of B cells specific for tumor-associated antigens [47,48]. Our analyses of IgG-subclasses in 11 accessible serum samples showed a normal distribution of IgG subclasses suggesting that the peripheral antibody repertoire is not significantly disturbed. However, we could not assess whether the intratumoral antibody repertoire was affected.
In comparison to the published literature on B cells in solid tumors our study in CRC patients reveals some commonalities but also important differences. Similar to the results described by Carpenter et al we found an increase of plasma cells in peripheral blood of cancer patients with advanced cancer [42]. Concerning the activation status of B cells, they found no significant difference in the percentage of CD86+ cells (6.7% vs. 4.2%) between tumor patients and controls. This is in line with our results for CRC patients. With regard to other B-cell populations our results differ from those of Carpenter et al [42]. In their report the percentage of circulating CD19+ cells was increased in malignant melanoma patients, whereas our study shows a significant decrease compared to healthy controls. They also describe a decrease in memory B cells with tumor progression. On the contrary, we observed an increased percentage of memory B cells in the peripheral blood of CRC patients. The differences between the results of Carpenter and our study can be due to a number of methodological differences between the two studies. The patients in the study of Carpenter et al consisted primarily of patients with malignant melanoma. Additionally, since the peripheral blood samples in our study were exclusively obtained prior to surgery, our study only included the subgroup with active disease as described by Carpenter et al. The different results concerning memory B cells of our study compared to the results of Carpenter et al in peripheral blood of malignant melanoma patients are most likely a result of differences in the gating strategy. We gated on CD19+CD20+ lymphocytic cells whereas Carpenter et al described the percentage of CD19+ cells. In addition, we included IgD-negativity to define CD27+ memory B cells. In line with our findings, the percentage of IgD+ cells was decreased in peripheral blood of patients with active malignant melanoma [42].
In addition to peripheral blood, we also analyzed B cells in freshly isolated and processed solid tumor samples of colorectal cancer patients. To ensure that samples comprised of representative tumor tissue the tumors were dissected by an experienced pathologist. To our knowledge, there are only two other studies addressing this issue: Nielsen et al analyzed tumor-associated B cells in ovarian cancer, including flow cytometric analyses of 9 tumor samples and Zirakzadeh et al. studied 7 urinary bladder cancer and 1 prostate cancer sample [9,40]. Our analysis revealed a highly activated B cell infiltrate. CD86+ activated B cells were significantly elevated in tumor tissue compared with PBMCs of healthy controls and CRC patients. This is in line with the results obtained in ovarian and urinary bladder cancer specimens using Ig-class-switch or CD69-positivity to characterize activated B cells [40]. In the study published by Nielsen et al. the majority of CD20+ B cells were IgG+ and classified as memory B cells with an unusual down-regulation of CD27. We observed a similar population (IgD-CD27-) but also a significant increase of classical IgD-CD27+ memory B cell subset suggesting that a down regulation of CD27 is less frequent in colorectal cancer than in ovarian cancer (Fig. 2B). Zirakzadeh et al also described an increased proportion of memory B cells in urinary bladder cancer. We visualized the distribution of tumor associated T and B cells by immunohistochemistry. Sequential stainings revealed a colocalization of Ki67 and CD20, suggesting a local proliferation, especially in peritumorous tertiary lymphoid structures. For T cells it is well established that tumor-infiltrating cells are mainly of an effector memory phenotype and our analyses confirmed this finding (Fig. 5A).
For both T and B cells an immunosuppressive/regulatory subset has been described with FoxP3+ regulatory T cells being the most well known subset. We observed a significant increase of regulatory T cells in colorectal cancer specimens thereby confirming prior studies [49]. Whereas regulatory T cell subsets have been extensively studied there is little knowledge about the role of regulatory B cells in human cancer [50]. To address this issue we included two recently described regulatory B-cell subsets, i.e. CD24highCD38high and CD24high CD27+B cells, in our analyses. CD24highCD38high “transitional” B cells could be detected in most colorectal cancer samples, but their frequency was generally low in tumors and even lower in peripheral blood. Of interest, the percentage of CD24highCD38high B cells was significantly elevated in advanced stage disease (Fig. 3A,B). Iwata et al. recently described CD24highCD27+ B cells as the major B cell population producing the immunosuppressive cytokine IL-10 in humans [12]. This population was present in the majority of tumor samples and in some cases represented more than 50% of CD19+CD20+ B cells (Fig.3 C,D). As we did not include IL-10 in our analyses, it needs to be further studied whether this population reflects the same IL-10 producing B cell subset. However, since we did not find a correlation between any of the two regulatory B cell subsets and the amount of regulatory T cells this finding would argue against a role of these regulatory B cell subsets in the expansion of regulatory T cells.
Since we observed changes in the proportion of the different B-cell subsets with disease progression we also analyzed biopsy specimens from liver metastases of six patients with metastatic colorectal cancer. Studies investigating the role of B cells in cancer metastasis have yielded conflicting results. Some studies found that a strong B cell infiltrate of the primary tumor was associated with a lower risk of distant metastasis [28,51] while others found the exact opposite [13,26,27,30,52]. We detected B cells in a significantly lower frequency than in primary tumor samples comprising less terminally differentiated B cells. IgD+CD27- memory B cells were decreased from 23.4% in tumor tissue to 9.9% among CD19+CD20+ B cells in colorectal liver metastases. The absence of B cells in colorectal cancer metastases is in line with results in metastatic melanoma [53]. Contrary to primary tumors, where we saw a strong infiltration by cells with a plasma cell phenotype this subset made up only 3.7% of CD19+ cells in metastatic tissue. However, regulatory B cells were significantly increased in metastatic tissue indicating a shift in the balance of B-cell subsets towards a more immunosuppressive state. These findings may reflect an immune escape mechanism of metastatic tumor cells, which results in a decreased detection by and activation of B cells.
T follicular helper cells (Tfh) are a novel T cell subset. It has recently been shown that Tfh cells also contribute to tumor immunity and that there is a correlation between B cells and Tfh within the tumor microenvironment [41,54]. Due to the importance of the interaction between Tfh and B cells during the germinal center reaction it would be interesting to investigate the cooperation of both cell types within the tumor microenvironment. The first reports that implied a role of Tfh in tumor immunology were published after the samples of our study had been analyzed. Therefore Tfh were not included in our FACS panel. In the future we are planning to study both populations using flow cytometry to clearly identify Tfh by multi-parameter flow cytometric analysis.
One limitation of our study is the low number of samples. This resulted in a low statistical power and limited our ability to identify statistically significant differences between the immune cell subsets. Compared to immunohistochemistry or gene expression analysis the flow cytometric assessment of the tumor immune infiltrate is more expensive and more time-consuming. Nonetheless, our study represents the largest flow cytometric characterization of tumor-infiltrating B lymphocytes published so far. To our knowledge this is the first report providing a detailed phenotypic analysis of the B-cell infiltrate in colorectal cancer.
In summary, our results show that patients with colorectal cancer have substantial alterations in their B cell compartment. Tumor-associated B cells are predominantly activated and of a mature phenotype suggesting a specific response to the tumor. On the other hand, metastatic lesions are characterized by a low number of B cells and an increased proportion of regulatory B cells, which might reflect immune escape. Importantly, this study also demonstrates that the composition of immune cells subsets in peripheral blood reflects changes in the tumor microenvironment. Our results provide important information for B cell-targeted therapeutic interventions and suggest that B cell-depletion at early tumor stages would be detrimental whilst in metastatic disease B cell-depletion could potentially enhance antitumor immunity through the depletion of regulatory B cells.
MATERIALS AND METHODS
Patient characteristics
Tumor samples (n=38), peripheral blood mononuclear cells (PBMCs) (n=46) and metastatic tissue (n=6) of 51 patients with a diagnosis of colorectal cancer were obtained between August 2010 and January 2013. Tumor stage was assessed according to the UICC tumor-node-metastasis criteria. Peripheral blood samples of 10 healthy donors were included as controls for PBMCs. The patient characteristics are summarized in table 1. Written informed consent was obtained from all patients prior to surgery and our institutional review board approved the study.
Cell isolation from human peripheral blood, tumor and metastatic tissue
Peripheral blood was obtained after written, informed consent from patients and age- matched healthy donors immediately prior to surgery. Peripheral blood mononuclear cells were purified using density-based separation with Ficoll-Paque PLUS (GE Healthcare Life Sciences).
Fresh unfixed tissue from primary tumor or metastatic lesions, which were not required for pathological analyses, were transferred to our laboratory immediately after surgical resection and processed within 12 hours. The samples were provided by an experienced pathologist (S. Eidt) who made sure that the samples we obtained were indeed tumor tissue. Furthermore histological analysis of the resection margins of our samples was performed. The fresh tumor tissue was manually minced using a scalpel and then transferred into single cells suspensions using a gentle MACS Dissociator (Miltenyi, Bergisch Gladbach, Germany). Single cell suspensions were obtained by sequential dissociation and incubation according to the company’s instructions. Afterwards, the cells were filtered through 70 μm nylon cell strainers (BD).
Flow cytometry
If possible at least 1x106 events per sample were acquired on a Gallios 10-color flow cytometer (Beckman Coulter, Krefeld, Germany). B-cell subsets were identified by multicolor staining using IgD, CD10, CD19, CD20, CD21, CD24, CD27, CD38, CD45, CD80, CD86 and HLA-DR (BD,NJ USA). Phenotypic characterization of T-cell subsets was performed using CD3, CD4, CD8, CD25, CD45, CD45RA, CCR7, CD127 and CD161 (BD, NJ USA). The data was analyzed using the Kaluza Software (Version 1.1, Beckman Coulter, Krefeld, Germany).
Immunohistochemistry
Immunohistochemistry on paraffin-embedded tumor samples was performed with an automated stainer (Medac) at the institute of pathology, Hildegardis-Hospital-Cologne. CD79a, CD20, Ki67, CD8 and IgG-kappa (Dako, Hamburg, Germany) antibodies were used for immunohistochemistry.
Nephelometry of IgG subclasses
Immunoglobulin-G-subclasses (IgG1, IgG2, IgG3 and IgG4) were analyzed on a Siemens BN ProSpec Analyzer using a Human IgG-Subclass liquid reagent kit (Binding Site, Birmingham, UK) according to the companies’ instructions.
Statistical analysis
Statistical analyses were performed with SPSS Version 20 (IBM corp.) and GraphPad Prism 5 (Graphpad Software, Inc.). Normal distribution was tested using the Shapiro-Wilk test of normality. In case of normal distribution student’s T-test was performed otherwise significance was analyzed by the Mann-Whitney-U-test.
FINANCIAL SUPPORT
The Else-Kröner-Fresenius-Stiftung and the Deutsche Forschungsgesellschaft (SFB832) supported research on tumor associated B cells.
DISCLOSURE
The authors declare no conflict of interests
References
1. Schreiber RD, Old LJ and Smyth MJ. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011; 331:6024. 1565–70.
2. Nelson BH. CD20+ B cells: the other tumor-infiltrating lymphocytes. J Immunol. 2010; 185:9. 4977–82.
3. DiLillo DJ, Yanaba K and Tedder TF. B cells are required for optimal CD4+ and CD8+ T cell tumor immunity: therapeutic B cell depletion enhances B16 melanoma growth in mice. J Immunol. 2010; 184:7. 4006–16.
4. Ammirante M, Luo J-L, Grivennikov S, Nedospasov S and Karin M. B-cell-derived lymphotoxin promotes castration-resistant prostate cancer. Nature. 2010; 464:7286. 302–5.
5. Jahrsdörfer B, Blackwell SE, Wooldridge JE, Huang J, Andreski MW, Jacobus LS, Taylor CM and Weiner GJ. B-chronic lymphocytic leukemia cells and other B cells can produce granzyme B and gain cytotoxic potential after interleukin-21-based activation. Blood. 2006; 108:8. 2712–9.
6. Zhang Y, Eliav Y, Shin S-U, Schreiber TH, Podack ER, Tadmor T and Rosenblatt JD. B lymphocyte inhibition of anti-tumor response depends on expansion of Treg but is independent of B-cell IL-10 secretion. Cancer immunology, immunotherapy: CII. 2012;
7. Menegaz RA, Michelin MA, Etchebehere RM, Fernandes PCJ and Murta EFC. Peri- and intratumoral T and B lymphocytic infiltration in breast cancer. European Journal of Gynaecological Oncology. 2008; 29:4. 321–6.
8. Yasuda M, Takenoyama M, Obata Y, Sugaya M, So T, Hanagiri T, Sugio K and Yasumoto K. Tumor-infiltrating B lymphocytes as a potential source of identifying tumor antigen in human lung cancer. Cancer research. 2002; 62:6. 1751–6.
9. Nielsen JS, Sahota RA, Milne K, Kost SE, Nesslinger NJ, Watson PH and Nelson BH. CD20+ tumor-infiltrating lymphocytes have an atypical CD27- memory phenotype and together with CD8+ T cells promote favorable prognosis in ovarian cancer. Clin Cancer Res. 2012; 18:12. 3281–92.
10. Maletzki C, Jahnke A, Ostwald C, Klar E, Prall F and Linnebacher M. Ex-vivo clonally expanded B lymphocytes infiltrating colorectal carcinoma are of mature immunophenotype and produce functional IgG. PLoS ONE. 2012; 7:2. e32639
11. Willis SN, Mallozzi SS, Rodig SJ, Cronk KM, McArdel SL, Caron T, Pinkus GS, Lovato L, Shampain KL, Anderson DE, Anderson RCE, Bruce JN and O’Connor KC. The microenvironment of germ cell tumors harbors a prominent antigen-driven humoral response. Journal of Immunology (Baltimore, Md: 1950). 2009; 182:5. 3310–7.
12. Iwata Y, Matsushita T, Horikawa M, Dilillo DJ, Yanaba K, Venturi GM, Szabolcs PM, Bernstein SH, Magro CM, Williams AD, Hall RP, St Clair EW and Tedder TF. Characterization of a rare IL-10-competent B-cell subset in man that parallels mouse regulatory B10 cells. Blood [Internet]. 2010; [cited 2010 Nov 4]; Available from: http://www.ncbi.nlm.nih.gov/pubmed/20962324
13. Olkhanud PB, Damdinsuren B, Bodogai M, Gress RE, Sen R, Wejksza K, Malchinkhuu E, Wersto RP and Biragyn A. Tumor-evoked regulatory B cells promote breast cancer metastasis by converting resting CD4+ T cells to T-regulatory cells. Cancer Res. 2011; 71:10. 3505–15.
14. Ray A, Basu S, Williams CB, Salzman NH and Dittel BN. A novel IL-10-independent regulatory role for B cells in suppressing autoimmunity by maintenance of regulatory T cells via GITR ligand. J Immunol. 2012; 188:7. 3188–98.
15. Gooden MJM, de Bock GH, Leffers N, Daemen T and Nijman HW. The prognostic influence of tumour-infiltrating lymphocytes in cancer: a systematic review with meta-analysis. Br J Cancer. 2011; 105:1. 93–103.
16. Ogino S, Nosho K, Irahara N, Meyerhardt JA, Baba Y, Shima K, Glickman JN, Ferrone CR, Mino-Kenudson M, Tanaka N, Dranoff G, Giovannucci EL and Fuchs CS. Lymphocytic reaction to colorectal cancer is associated with longer survival, independent of lymph node count, microsatellite instability, and CpG island methylator phenotype. Clin Cancer Res. 2009; 15:20. 6412–20.
17. De Miranda NFCC, Goudkade D, Jordanova ES, Tops CMJ, Hes FJ, Vasen HFA, van Wezel T and Morreau H. Infiltration of Lynch colorectal cancers by activated immune cells associates with early staging of the primary tumor and absence of lymph node metastases. Clin Cancer Res. 2012; 18:5. 1237–45.
18. Galon J, Costes A, Sanchez-Cabo F, Kirilovsky A, Mlecnik B, Lagorce-Pages C, Tosolini M, Camus M, Berger A, Wind P, Zinzindohoue F, Bruneval P, Cugnenc PH, Trajanoski Z, Fridman WH and Pages F. Type, density, and location of immune cells within human colorectal tumors predict clinical outcome. Science (New York, NY.. 2006; 313:5795. 1960–4.
19. Fremd C, Schuetz F, Sohn C, Beckhove P and Domschke C. B cell-regulated immune responses in tumor models and cancer patients. Oncoimmunology. 2013; 2:7. e25443
20. De Visser KE, Korets LV and Coussens LM. De novo carcinogenesis promoted by chronic inflammation is B lymphocyte dependent. Cancer Cell. 2005; 7:5. 411–23.
21. Shah S, Divekar AA, Hilchey SP, Cho HM, Newman CL, Shin SU, Nechustan H, Challita-Eid PM, Segal BM, Yi KH and Rosenblatt JD. Increased rejection of primary tumors in mice lacking B cells: inhibition of anti-tumor CTL and TH1 cytokine responses by B cells. International journal of cancer. 2005; 117:4. 574–86.
22. Schioppa T, Moore R, Thompson RG, Rosser EC, Kulbe H, Nedospasov S, Mauri C, Coussens LM and Balkwill FR. B regulatory cells and the tumor-promoting actions of TNF-α during squamous carcinogenesis. Proc Natl Acad Sci USA. 2011; 108:26. 10662–7.
23. Balkwill F, Montfort A and Capasso M. B regulatory cells in cancer. Trends Immunol. 2013; 34:4. 169–73.
24. Qin Z, Richter G, Schuler T, Ibe S, Cao X and Blankenstein T. B cells inhibit induction of T cell-dependent tumor immunity. Nature medicine. 1998; 4:5. 627–30.
25. Inoue S, Leitner WW, Golding B and Scott D. Inhibitory effects of B cells on antitumor immunity. Cancer research. 2006; 66:15. 7741–7.
26. Staquicini FI, Tandle A, Libutti SK, Sun J, Zigler M, Bar-Eli M, Aliperti F, Pérez EC, Gershenwald JE, Mariano M, Pasqualini R, Arap W and Lopes JD. A subset of host B lymphocytes controls melanoma metastasis through a melanoma cell adhesion molecule/MUC18-dependent interaction: evidence from mice and humans. Cancer Res. 2008; 68:20. 8419–28.
27. Ruddell A, Harrell MI, Furuya M, Kirschbaum SB and Iritani BM. B lymphocytes promote lymphogenous metastasis of lymphoma and melanoma. Neoplasia. 2011; 13:8. 748–57.
28. Quan N, Zhang Z, Demetrikopoulos MK, Kitson RP, Chambers WH, Goldfarb RH and Weiss JM. Evidence for involvement of B lymphocytes in the surveillance of lung metastasis in the rat. Cancer Res. 1999; 59:5. 1080–9.
29. Martinet L, Garrido I, Filleron T, Le Guellec S, Bellard E, Fournie J-J, Rochaix P and Girard J-P. Human solid tumors contain high endothelial venules: association with T- and B-lymphocyte infiltration and favorable prognosis in breast cancer. Cancer Res. 2011; 71:17. 5678–87.
30. Eiró N, Pidal I, Fernandez-Garcia B, Junquera S, Lamelas ML, Del Casar JM, González LO, López-Muñiz A and Vizoso FJ. Impact of CD68/(CD3+CD20) Ratio at the Invasive Front of Primary Tumors on Distant Metastasis Development in Breast Cancer. PLoS ONE. 2012; 7:12. e52796
31. Li Q, Lao X, Pan Q, Ning N, Yet J, Xu Y, Li S and Chang AE. Adoptive transfer of tumor reactive B cells confers host T-cell immunity and tumor regression. Clin Cancer Res. 2011; 17:15. 4987–95.
32. Jackaman C, Cornwall S, Graham PT and Nelson DJ. CD40-activated B cells contribute to mesothelioma tumor regression. Immunol Cell Biol. 2011; 89:2. 255–67.
33. Sorenmo KU, Krick E, Coughlin CM, Overley B, Gregor TP, Vonderheide RH and Mason NJ. CD40-activated B cell cancer vaccine improves second clinical remission and survival in privately owned dogs with non-Hodgkin’s lymphoma. PLoS ONE. 2011; 6:8. e24167
34. Ding C, Wang L, Marroquin J and Yan J. Targeting of antigens to B cells augments antigen-specific T-cell responses and breaks immune tolerance to tumor-associated antigen MUC1. Blood. 2008; 112:7. 2817–25.
35. Julien S, Picco G, Sewell R, Vercoutter-Edouart A-S, Tarp M, Miles D, Clausen H, Taylor-Papadimitriou J and Burchell JM. Sialyl-Tn vaccine induces antibody-mediated tumour protection in a relevant murine model. Br J Cancer. 2009; 100:11. 1746–54.
36. Candolfi M, Curtin JF, Yagiz K, Assi H, Wibowo MK, Alzadeh GE, Foulad D, Muhammad AKMG, Salehi S, Keech N, Puntel M, Liu C, Sanderson NR, Kroeger KM, Dunn R, Martins G and et al. B cells are critical to T-cell-mediated antitumor immunity induced by a combined immune-stimulatory/conditionally cytotoxic therapy for glioblastoma. Neoplasia. 2011; 13:10. 947–60.
37. Tarella C, Passera R, Magni M, Benedetti F, Rossi A, Gueli A, Patti C, Parvis G, Ciceri F, Gallamini A, Cortelazzo S, Zoli V, Corradini P, Carobbio A, Mulé A and Bosa M, et al. Risk factors for the development of secondary malignancy after high-dose chemotherapy and autograft, with or without rituximab: a 20-year retrospective follow-up study in patients with lymphoma. J Clin Oncol. 2011; 29:7. 814–24.
38. Barbera-Guillem E, Nelson MB, Barr B, Nyhus JK, May KF, Feng L and Sampsel JW. B lymphocyte pathology in human colorectal cancer. Experimental and clinical therapeutic effects of partial B cell depletion. Cancer Immunol Immunother. 2000; 48:10. 541–9.
39. Wejksza K, Lee-Chang C, Bodogai M, Bonzo J, Gonzalez FJ, Lehrmann E, Becker K and Biragyn A. Cancer-produced metabolites of 5-lipoxygenase induce tumor-evoked regulatory B cells via peroxisome proliferator-activated receptor α. J Immunol. 2013; 190:6. 2575–84.
40. Zirakzadeh AA, Marits P, Sherif A and Winqvist O. Multiplex B cell characterization in blood, lymph nodes, and tumors from patients with malignancies. J Immunol. 2013; 190:11. 5847–55.
41. Bindea G, Mlecnik B, Tosolini M, Kirilovsky A, Waldner M, Obenauf AC, Angell H, Fredriksen T, Lafontaine L, Berger A, Bruneval P, Fridman WH, Becker C, Pagès F, Speicher MR and Trajanoski Z, et al. Spatiotemporal dynamics of intratumoral immune cells reveal the immune landscape in human cancer. Immunity. 2013; 39:4. 782–95.
42. Carpenter EL, Mick R, Rech AJ, Beatty GL, Colligon TA, Rosenfeld MR, Kaplan DE, Chang K-M, Domchek SM, Kanetsky PA, Fecher LA, Flaherty KT, Schuchter LM and Vonderheide RH. Collapse of the CD27+ B-cell compartment associated with systemic plasmacytosis in patients with advanced melanoma and other cancers. Clin Cancer Res. 2009; 15:13. 4277–87.
43. Richards CH, Flegg KM, Roxburgh CSD, Going JJ, Mohammed Z, Horgan PG and McMillan DC. The relationships between cellular components of the peritumoural inflammatory response, clinicopathological characteristics and survival in patients with primary operable colorectal cancer. Br J Cancer. 2012; 106:12. 2010–5.
44. Mohammed ZMA, Going JJ, Edwards J, Elsberger B, Doughty JC and McMillan DC. The relationship between components of tumour inflammatory cell infiltrate and clinicopathological factors and survival in patients with primary operable invasive ductal breast cancer. Br J Cancer. 2012; 107:5. 864–73.
45. Lohr M, Edlund K, Botling J, Hammad S, Hellwig B, Othman A, Berglund A, Lambe M, Holmberg L, Ekman S, Bergqvist M, Pontén F, Cadenas C, Marchan R, Hengstler JG and Rahnenführer J, et al. The prognostic relevance of tumour-infiltrating plasma cells and immunoglobulin kappa C indicates an important role of the humoral immune response in non-small cell lung cancer. Cancer Lett. 2013;
46. Schmidt M, Hellwig B, Hammad S, Othman A, Lohr M, Chen Z, Boehm D, Gebhard S, Petry I, Lebrecht A, Cadenas C, Marchan R, Stewart JD, Solbach C, Holmberg L, Edlund K and et al. A comprehensive analysis of human gene expression profiles identifies stromal immunoglobulin κ C as a compatible prognostic marker in human solid tumors. Clin Cancer Res. 2012; 18:9. 2695–703.
47. Pavoni E, Monteriu G, Santapaola D, Petronzelli F, Anastasi AM, Pelliccia A, D’Alessio V, De Santis R and Minenkova O. Tumor-infiltrating B lymphocytes as an efficient source of highly specific immunoglobulins recognizing tumor cells. BMC Biotechnology. 2007; 7:70
48. Nzula S, Going JJ and Stott DI. Antigen-driven clonal proliferation, somatic hypermutation, and selection of B lymphocytes infiltrating human ductal breast carcinomas. Cancer Res. 2003; 63:12. 3275–80.
49. Tosolini M, Kirilovsky A, Mlecnik B, Fredriksen T, Mauger S, Bindea G, Berger A, Bruneval P, Fridman W-H, Pagès F and Galon J. Clinical impact of different classes of infiltrating T cytotoxic and helper cells (Th1, th2, treg, th17) in patients with colorectal cancer. Cancer Res. 2011; 71:4. 1263–71.
50. Bodogai M, Lee-Chang C, Wejksza K, Lai J-P, Merino M, Wersto RP, Gress RE, Chan AC, Hesdorffer C and Biragyn A. Anti-CD20 antibody promotes cancer escape via enrichment of tumor-evoked regulatory B cells expressing low levels of CD20 and CD137L. Cancer Res. 2013;
51. Nagalla S, Chou JW, Willingham MC, Ruiz J, Vaughn JP, Dubey P, Lash TL, Hamilton-Dutoit SJ, Bergh J, Sotiriou C, Black MA and Miller LD. Interactions between immunity, proliferation and molecular subtype in breast cancer prognosis. Genome Biol. 2013; 14:4. R34
52. Camus M, Tosolini M, Mlecnik B, Pagès F, Kirilovsky A, Berger A, Costes A, Bindea G, Charoentong P, Bruneval P, Trajanoski Z, Fridman W-H and Galon J. Coordination of intratumoral immune reaction and human colorectal cancer recurrence. Cancer Res. 2009; 69:6. 2685–93.
53. Hernberg M, Turunen JP, Muhonen T and Pyrhönen S. Tumor-infiltrating lymphocytes in patients with metastatic melanoma receiving chemoimmunotherapy. J Immunother. 1997; 20:6. 488–95.
54. Gu-Trantien C and Willard-Gallo K. Tumor-infiltrating follicular helper T cells: The new kids on the block. Oncoimmunology. 2013; 2:10. e26066