
Introduction
Aging is a well-known-risk factor for dry eye. Prevalence of dry eye disease increases with age after the 4th decade and it’s more prevalent in women [1-3]. A frequent finding in dry eye is keratinization of the ocular surface epithelium and loss of goblet cells [4-8]. Both IFN-γ and IL-17 have been implicated in the pathogenesis of dry eye associated ocular surface epithelial disease [9-12].
Alterations in cellular immunity are a known aspect of aging [13]. With increasing age, the thymus begins to shrink, leading to a decrease in the number of naïve T-lymphocytes in the circulation. A compensatory auto proliferation of existing, mature T-cells may lead to the expansion of CD4 positive (+) T-cells. These cells are resistant to apoptosis and have been shown to be associated with a variety of autoimmune disorders such as rheumatoid arthritis, multiple sclerosis, and diabetes mellitus [14].
Interferon-gamma (IFN-γ), produced by natural killer, natural killer T cells and T cells, is the signature cytokine of T helper (Th) 1 immune responses and it has been involved in the pathogenesis of colitis and Sjögren Syndrome [15-18]. IFN-γ production by T cells in peripheral blood and conjunctiva progressively increases with age in human studies [19, 20]. There is mounting evidence about a pathogenic role for IFN-γ in dry eye. We have previously shown that desiccating stress decreases goblet cell density mainly to increased IFN-γ, as IFN-γKO mice were resistant to desiccation-induced changes [21]. Subconjunctival injections of IFN-γ or anti-IFN-γ antibody decreased or improved goblet cell density in an experimental dry eye model, respectively [21-24]. IFN-γ produced by NK cells is responsible for upregulation of Th1 cell recruiting chemokines by conjunctival and corneal epithelial cell in experimental dry eye [25, 26]. Cultured conjunctival goblet cells are exquisitely sensitive to IFN-γ, as even minute concentrations of this cytokine promotes goblet cell apoptosis [27]. Increased IFN-γ has been observed in tears and conjunctiva of dry eye patients [9, 12, 28].
IL-17A is the signature cytokine of Th-17 immune responses. It has been implicated in the pathogenesis of several autoimmune diseases, including rheumatoid arthritis, psoriasis and uveitis and inflammatory bowel disease [29-34]. We and others have shown that IL-17A disrupts apical corneal barrier apical function in experimental dry eye [9, 11], by stimulating matrix metalloproteinases that have been implicated in breakdown of epithelial tight junctions [9]. Our published study showed that 24 month-old C57BL/6 mice spontaneously develop a SS-like keratoconjunctivitis sicca, with increased IL-17A and IFN-γ transcript levels in conjunctiva, which was accompanied by loss of conjunctival goblet cells and increased corneal barrier disruption [10]. The role of immune dysfunction in the pathogenesis of age-associated dry eye disease has not been sufficiently investigated.
The purpose of this study was to investigate the roles of IFN-γ and IL-17 in aging and sex-related goblet cell loss and corneal barrier function, respectively. Our studies showed that goblet cell loss in aged IFN-γKO mice was significantly lower than age-matched B6 mice. This was accompanied by increased IFN-γ and IFN-γ inducible-chemokines in the conjunctiva. Increased corneal barrier dysfunction was observed in female mice.
Results
IFN-gamma deficiency reduces aging-related goblet cell loss
One of the hallmarks of aqueous deficient dry eye disease is a decrease in the number of mucin-filled goblet cells [12, 35, 36]. We measured the number of filled goblet cells in 8-week (8W) and 15-month (15M) old B6 and IFN-γKO mice in PAS stained conjunctival sections. There was a significant decrease in the number of PAS+ goblet cells in aged B6 mice, compared to 8-week old mice, irrespective of sex (P < 0.01, Figure 1A-1B). A significant decrease in GC count was also found in male 15M IFN-KO mice P < 0.05, Figure 1A-1B) compared to its baseline, although this was significantly less than the age-related goblet cell loss observed in in B6 mice (~24% vs. 40%, respectively).
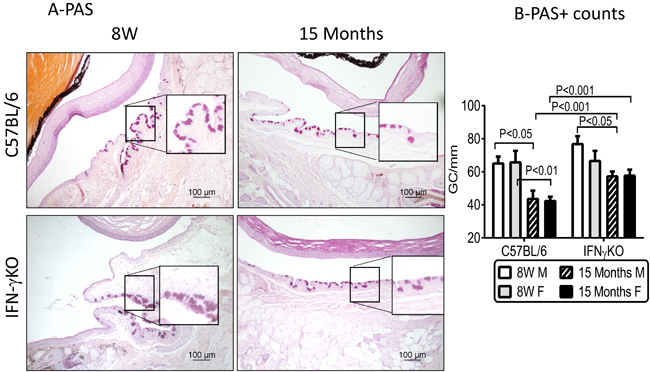
Figure 1: Aging leads to decreased goblet cell density in C57BL/6 mice. A. Representative digital images of PAS stained goblet cells (GC) in conjunctiva (purple-magenta cells) of male C57BL/6 and IFN-γ KO mice. Black squares are higher magnification showing goblet cells in details. B. Number of PAS+ conjunctival GC in C57BL/6 and IFN-γ KO mice of both sexes counted in paraffin-embedded sections. (Mean±SEM, n = 5 right eyes). W = weeks; M = male; F = female.
Age-related increase in T helper (Th) 1 and Th17 associated cytokines in conjunctiva
We have previously shown that T-cell related cytokines are elevated in the conjunctiva of 24 month-old B6 mice [10]. In this study, we evaluated expression of Th1 (IFN-γ, CXCL10), Th17 (IL-17A, CCL20), and Th2 (IL-13) related factors in conjunctiva of young and old mice by real time PCR.
We found that levels of IFN-γ mRNA transcript were significantly higher in female B6 mice at 8W (P < 0.001) and in male B6 at 15M (P < 0.001, Figure 2A), but they were completely absent in the IFN-γKO mice. The mRNA transcripts of CXCL10, a chemokine involved in trafficking of Th-1 cells [37], were significantly elevated in the young female B6 mice and aged male B6 mice (P < 0.0001, Figure 2B) and were higher in B6 than IFN-γKO in young female and aged male.
CCL20 is a chemokine involved in trafficking of Th-17 cells [38, 39]. CCL20 and IL-17A mRNA transcript levels in conjunctiva were elevated in both young and aged female B6 compared to males (P < 0.001). Compared to B6, CCL20 and IL-17A levels were dramatically increased in IFN-γKO mice of both sexes of both ages (P < 0.0001, Figure 2C-2D).
IL-13 is a cytokine that promotes goblet cell homeostasis [40, 41]. IL-13 mRNA transcript levels were higher in female 8W B6 and male 15M B6 (P < 0.0001, Figure 2E), and at the same time were significantly higher in IFN-γKO female mice, in both young and aged groups (P < 0.0001). IL-13/IFN-γ mRNA ratio was decreased in aged male B6, while IFN-γKO mice had unchecked IL-17 and IL-13 (Figure 2F and not shown).
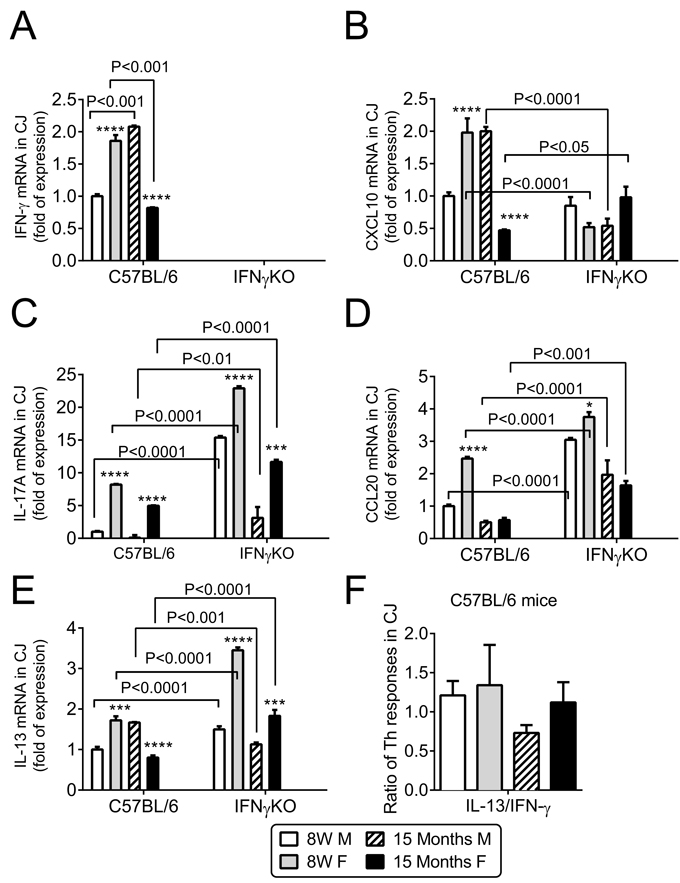
Figure 2: Aging induces sex and age specific findings in conjunctiva. Relative fold of mRNA expression of IFN-γ A. and CXCL10 B., IL-17A C., CCL20 D., IL-13 E., and IL-13/IFN-γ ratio in conjunctiva in B6 mice F. Bar graphs show mean ± SEM (n = 6 animals, conjunctiva from both eyes of the same animal were pooled into one sample, yielding a final sample size of 6). W = weeks; M = male; F = female; * P < 0.05; ** P < 0.01; *** P < 0.001; **** P < 0.0001; sex comparison at same age.
Decreased corneal barrier function in aged female mice
Increased corneal permeability to fluorescent dyes is another hallmark of dry eye. Antibody neutralization of IFN-γ itself or of IFN-γ-producing NK cells has been shown to protect against developing corneal staining in a murine dry eye model [24, 26]. We compared corneal permeability to the 70kDa fluorescent molecule Oregon-Green Dextran (OGD) of young and aged B6 and IFN-γKO mice of both sexes. Sex-specific differences were observed; aged female B6 had increased corneal barrier disruption which was not observed in age-matched male B6 mice (Figure 3A-3B). IFN-γKO mice of both sexes had increased uptake of OGD dye with aging; however, similar to B6 mice, these changes were more pronounced in females. Moreover, corneal permeability in 15M female IFN-γKO mice was also significantly increased compared to 15M B6 mice.
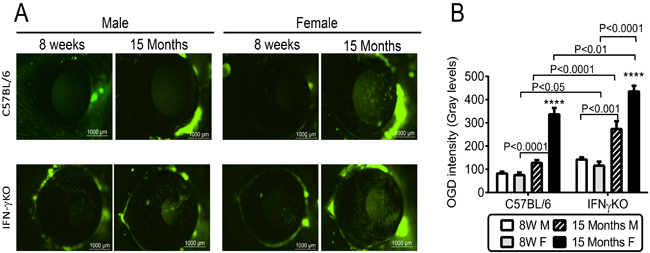
Figure 3: Corneal barrier function in C57BL/6 and IFN-γKO mice of both sexes at 8 weeks and 15 months of age. A. Representative digital images of Oregon Green dextran (OGD) corneal staining that were used to generate the bar graphs in B. B. Corneal Oregon Green dextran (OGD) fluorescence intensity score (mean±SEM, n = 15 animals). W = weeks; M = male; F = female; * P < 0.05; ** P < 0.01; *** P < 0.001; **** P < 0.0001; sex comparison at same age.
Age-related increase in T-cell associated cytokines in cornea
IL-17A and matrix-metalloproteinase -9 (MMP-9) have been found to participate in desiccation-induced corneal barrier disruption [9, 11, 42]. We hypothesized that aged-related corneal barrier disruption would be accompanied by an increase in the same factors as those previously observed in inducible dry eye models. Therefore, we compared the expression of IL-17A, CCL20 and MMP-9, in corneas of aged mice to young mice of both groups by real-time PCR.
In B6 mice, IL-17A mRNA transcript levels were elevated only in aged females (P < 0.001), and they were dramatically increased in IFN-γKO mice of both sexes (P < 0.001) and particularly in young and old (P < 0.0001, Figure 4A). CCL20 mRNA transcript level in cornea was elevated in the both young and aged female B6 mice (P < 0.0001), and significantly increased in young IFN-γKO mice of both sexes (P < 0.0001) and in aged IFN-γKO female mice (P < 0.0001; Figure 4B). MMP-9 has been shown to mediate corneal barrier disruption in dry eye [42]. MMP-9 mRNA transcript levels were significantly elevated in aged female B6 cornea compared to young mice (P < 0.0001), and significantly higher in female IFN-γKO mice, in both young (P < 0.0001) and aged (P < 0.05) groups (Figure 4C).
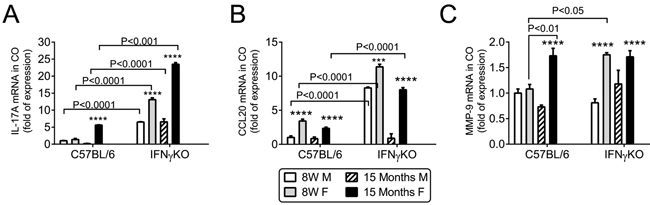
Figure 4: Aging increases IL-17, CCL20 and MMP-9 mRNA expression in cornea. Relative fold of mRNA expression of IL-17A A., CCL20 B. and MMP-9 C. in corneal epithelium. Bar graphs show mean ± SEM (n = 6, corneal epithelium pooled from both eyes of the same animal). W = weeks; M = male; F = female; * P < 0.05; ** P < 0.01; *** P < 0.001; **** P < 0.0001; sex comparison at same age.
Discussion
Dry eye is a very prevalent disease in the aging population [1, 2]. It is estimated that dry eye is one of the top reasons for seeking eye care, mostly because corneal barrier disruption is responsible for irritation and decreased vision [43, 44]. Among dry eye patients, 11% have been estimated to have the systemic autoimmune condition Sjögren Syndrome [45, 46], a severe and potentially blinding condition. Strong correlation between age and female sex and prevalence of DED has been well established [2, 47]. However, the exact mechanisms why dry eye is more prevalent in aged women have not been elucidated yet.
Goblet cells are highly secretory epithelial cells that are abundant in conjunctiva, and digestive tract. Cornification and goblet cell loss in the conjunctival epithelium has been recognized in dry eye patients for over than three decades [8, 12, 48, 49]. Our previous studies showed that 24-month old B6 mice of both sexes had a decreased number of mucin-filled goblet cells [10]. This was accompanied by increased IFN-γ expression, leading us to hypothesize that IFN-γ may contribute to the goblet cell loss in aged mice. Our current study demonstrated that IFN-γKO mice had a significantly less goblet cells loss than age-matched B6 mice. The decline in goblet cells was accompanied by increased IFN-γ and CXL10 expression in the conjunctiva of B6 mice. Interestingly, aged female B6 and IFN-γKO mice displayed greater corneal barrier disruption and increased IL-17A and CCL20 mRNA levels in cornea than aged male mice.
Our findings indicated that IFN-γ is partially responsible for aging-related goblet cell loss. Williams and colleagues showed an age-related increase in IFN-γ-producing CD4+ T cells in conjunctiva of human subjects [20]. Recent studies have shown a close relationship between the mucosal immune system and conjunctival goblet cells, a relationship that has been identified in other mucosal tissues, including the intestine and airways [50, 51]. Studies in the lung have demonstrated that IL-13 induces goblet cell hyperplasia [52, 53]. In the conjunctiva, goblet cell differentiation and MUC5AC production is reciprocally regulated by the T helper cytokines IFN-γ and IL-13 [21, 27, 40, 41]. We have shown that IL-13 has a homeostatic role in maintaining conjunctival goblet cells, whereas IFN-γ promotes conjunctival cornification and goblet cell loss [21, 24]. IL-13KO mice have decreased goblet cell density at baseline and subconjunctival injections of IL-13 during experimental dry eye rescued goblet cell density compared to placebo-injected mice [40]. We have also shown that IL-13 -stimulated cultured conjunctival goblet cells have increased proliferation and production of Muc5ac and Muc2 [41]. Prior studies from our laboratory showed that IFN-γKO mice were resistant to desiccation-induced goblet cell loss and subconjunctival administration of IFN-γ in IFN-γKO mice decreased goblet cell to similar levels as wild-type mice [21], as well as upregulated cornified envelope proteins that also increase in dry eye [4]. We have also demonstrated that subconjunctival injections of anti-IFN-γ during desiccating stress prevented goblet cell loss [24]. We also discovered that IFN-γ causes endoplasmic reticulum stress and induces an unfolded protein response in cultured conjunctival goblet cells, as a molecular mechanism responsible for the reduced mucin secretion induced by IFN-γ [27]. Our primary conjunctival cultures are exquisitely sensitive to IFN-γ as even minute concentrations of this cytokine decreased proliferation, inhibited synthesis of mature mucins and induced profound morphological changes, despite increased levels of mucin gene transcripts [27]. Taken together, these results indicate a critical role for IFN- in conjunctival goblet cell loss/dysfunction, in vitro, and in two different models of dry eye, including aging and desiccating stress in young mice.
IL-17A has also been noted to increase in mice and humans with aging and to participate in autoimmunity [54-57]. IL-17A has been shown to cause epithelial barrier disruption in mice [9, 11] and to increase expression of MMPs [58, 59]. MMP-induced lysis of tight junctions and desquamation of the corneal epithelium is responsible for corneal barrier disruption in dry eye [42]. The discovery of Th-17 cells has challenged the Th1/Th2 paradigm of cross regulation of autoimmunity. While IFN-γ has been shown to inhibit the development of IL-17-producing effector CD4+ T cells in vitro [60, 61], an in vivo suppressive effect of IFN-γ on IL-17 production has not been clearly demonstrated. Both Th1 and Th17 cells are pathologic in some experimental models, such as experimental autoimmune uveitis, dry eye and rheumatoid arthritis [62-67]. In the collagen-induced arthritis model, young IFN-γKO B6 mice produced higher levels of IL-17 in response to immunization with type II collagen compared to wild-type B6 mice [68]. Anti-IL-17 antibody prevented collagen-induced arthritis in the IFN-γ-receptor KO, which has elevated levels of IL-17 compared to wild-type mice [64]. Antibody depletion of IL-17 prior to and during desiccating stress did not increase IFN-γ mRNA in conjunctiva [9]. Here we report that suppression of IL-17A by IFN-γ occurs naturally even in young mice, but a significant clinical phenotype (corneal barrier disruption) is only found with advanced aging, demonstrating that a certain accumulation of still unknown events is necessary for a full dry eye phenotype. Interestingly, greater age-related corneal barrier disruption was observed in female mice. This is in agreement with a recent study demonstrating that young female B6 mice are more susceptible to corneal barrier disruption in a desiccating environment and have greater levels of IL-17 in draining lymph nodes and lacrimal gland than young male mice subjected to the same protocol [69]. IL-17 mRNA levels in corneal epithelia peaked at the same time as maximum corneal barrier disruption in the 8-week old CD25 (IL-2 receptor alpha) knock-out mouse model of Sjögren Syndrome [33]. Our group was the first to describe that aged female B6 mice spontaneously develop corneal barrier disruption [10]. Increased levels of IL-17A have been reported in both tears and conjunctiva of dry eye patients. [9, 70-72]. In humans, dry eye disease affects both sexes, but it has a great incidence in female patients [1, 2, 73-75]. We can speculate that increased corneal disease and increased IL-17 in females may be one of the reasons why dry eye is more prevalent in female sex, but further studies are necessary to investigate this correlation.
In conclusion, this study provides evidence that IFN-γ actively participates in aging-related conjunctival goblet cell loss and also provides insights about the complex cytokine interactions in dry eye pathogenesis. Therapies targeting both IFN-γ and IL-17A cytokines may be needed to fully address the spectrum of dry eye manifestations and to improve the quality of visual function and life of an aging population.
Materials and Methods
Animals
This research protocol was approved by the Baylor College of Medicine Center for Comparative Medicine, and it conformed to the standards of the Association for Research in Vision and Ophthalmology Statement for the Use of Animals in Ophthalmic and Vision Research. C57BL/6 (B6), IFN-γKO (B6.129S7-ifngtm1Ts/J, on a B6 background) mice of both sexes were purchased from Jackson Laboratories (Bar Harbor, ME) for establishing breeding colonies in our facility and were used at 8 weeks and 15 months of age. Offspring mice housed in our vivarium until they reach appropriate age. Twenty-six mice per age/strain/sex were used: five for histologic sections, 15 for measuring corneal permeability, six for gene analysis.
Corneal permeability to oregon-green dextran
Corneal epithelial permeability to Oregon-Green dextran [OGD; 70 000 molecular weight (MW); Invitrogen, Eugene, OR, USA] was assessed at 8 weeks and 15 months in both sexes and strains (C57BL/6 and IFN-γKO) as previously described [9]. Briefly, 0.5 ml of 50µg/ml OGD was instilled onto the ocular surface 1 min before euthanasia. Corneas were rinsed with 2 μl of phosphate buffered saline (PBS) and photographed with a stereoscopic zoom microscope (model SMZ 1500; Nikon, Melville, NY, USA), under fluorescence excitation at 470 nm. The severity of corneal OGD staining was digitally graded as previously reported [9]. The mean fluorescence intensity measured inside the 2-mm central zone by the image analysis software (NIS Elements, version 3.0, Nikon, Melville, NY) was transferred to a database and the results averaged within each group.
Histology, periodic acid schiff staining
Five right eyes from each sex, at 8W and 15M, were surgically excised, fixed in 10% formalin, paraffin embedded, and 8-µm sections were cut. Ocular sections were cut at the center of the eye, where the lens has its maximum diameter. Sections were stained with H&E for evaluating morphology and with periodic-Schiff (PAS) reagent for measuring goblet cell density. The superior and inferior conjunctiva were examined and photographed with a microscope equipped with a digital camera (Eclipse E400 with a DS-Fi1; Nikon, Melville, NY, USA). The number of goblet cells in the superior and inferior conjunctiva was measured in 3 sections from each eye that were at least 300 µm apart, using image-analysis software (NIS Elements Software, version 3.0, BR, Nikon) and expressed as number of goblet cells per mm as previously reported [21].
RNA isolation and real-time PCR
Total RNA from the cornea and conjunctiva was collected as previously described [9] and extracted using a Qiagen RNeasy Plus® Micro RNA isolation Kit (Qiagen, Valencia, CA) according to the manufacturer’s instructions, quantified by a NanoDrop® ND-1000 Spectrophotometer (Thermo Scientific, Wilmington, DE) and stored at -80°C. Six samples per group/age were used, and 1 sample consisted of pooled conjunctiva or cornea of right and left eyes of the same animal. The RNA concentration was measured by its absorption at 260 nm and samples were stored at -80°C until use. First-strand cDNA was synthesized with random hexamers by M-MuLV reverse transcription (Ready-To-Go You-Prime First-Strand Beads; GE Healthcare, Inc., Arlington Heights, NJ), as previously described [9]. Real-time PCR was performed with specific MGB probes (Taqman; Applied Biosystems, Inc., Foster City, CA) and PCR master mix (Taqman Gene Expression Master Mix), in a commercial thermocycling system (StepOnePlus Real-Time PCR System, Applied Biosystems), according to the manufacturer’s recommendations. Murine MGB probes were HPRT1 (Mm00446968), CCL20 (Mm00444228), CXCL10 (Mm445235), IFN-γ (Mm00801778), IL-13 (Mm99999190), IL-17A (Mm00439618) and MMP-9 (Mm00442991). The HPRT1 gene was used as an endogenous reference for each reaction. The results of quantitative PCR were analyzed by the comparative CT method where target change = 2ΔΔCT. The results were normalized by the CT value of HPRT1. The mean CT of relative mRNA level in the eight week male group was used as the calibrator.
Statistical analysis
Two-way analysis of variance (ANOVA) or the Kruskall-Wallis tests were used for overall statistical comparisons, followed by Sidak’s multiple comparison test. P < 0.05 was considered statistically significant. These tests were performed using GraphPad Prism 6.0 software (GraphPad Software, Inc., San Diego, CA).
Abbreviations
ANOVA: analysis of variance; APC: antigen-presenting cells; CCL20: CC-chemokine attractant ligand 20; CXCL10: C-X-C motif chemokine ligand 10 (or IP-10: IFN-γ inducible protein 10); DED: dry eye disease; GC: goblet cells; HPRT1: hypoxanthine phosphoribosyltransferase 1 gene; IFN-γ: interferon-gamma; IL: interleukin; MGB: 3’-minor groove binder-DNA probes; MGD: meibomian gland disease; MHC: major histocompatibility complex; MMPs: matrix metallo-proteinases; M-MuLV: Moloney murine leukemia virus; MUC5AC: mucin 5AC gene with protein product; OCT: optimal cutting temperature compound; OGD: Oregon Green dextran; PBS: phosphate-buffered saline;; RT-PCR: real-time polymerase chain reaction.
Acknowledgments
We want to thank Kevin Tesareski for his expert technical assistance.
Conflicts of Interest
The sponsor agencies had no involvement in the study design; data collection, analysis, and interpretation of data; writing of the report; or the decision to submit the paper for publication. The authors declare no conflicts of interest.
References
1. Schaumberg DA, Dana R, Buring JE and Sullivan DA. Prevalence of dry eye disease among US men: estimates from the Physicians’ Health Studies. Arch Ophthalmol. 2009; 127:763-768.
2. Schaumberg DA, Sullivan DA, Buring JE and Dana MR. Prevalence of dry eye syndrome among US women. Am J Ophthalmol. 2003; 136:318-326.
3. Uchino M, Dogru M, Uchino Y, Fukagawa K, Shimmura S, Takebayashi T, Schaumberg DA and Tsubota K. Japan Ministry of Health study on prevalence of dry eye disease among Japanese high school students. Am J Ophthalmol. 2008; 146:925-929 e922.
4. Corrales RM, de Paiva CS, Li DQ, Farley WJ, Henriksson JT, Bergmanson JP and Pflugfelder SC. Entrapment of conjunctival goblet cells by desiccation-induced cornification. Invest Ophthalmol Vis Sci. 2011; 52:3492-3499.
5. Lemp MA. The mucin-deficient dry eye. Int Ophthalmol Clin. 1973; 13:185-189.
6. Murube J and Rivas L. Biopsy of the conjunctiva in dry eye patients establishes a correlation between squamous metaplasia and dry eye clinical severity. Eur J Ophthalmol. 2003; 13:246-256.
7. Murube J and Rivas L. Impression cytology on conjunctiva and cornea in dry eye patients establishes a correlation between squamous metaplasia and dry eye clinical severity. Eur J Ophthalmol. 2003; 13:115-127.
8. Pflugfelder SC, Huang AJW, Schuchovski PT, Pereira IC and Tseng SCG. Conjunctival cytological features of primary Sjogren syndrome. Ophthalmology. 1990; 97:985-991.
9. de Paiva CS, Chotikavanich S, Pangelinan SB, Pitcher JD, III, Fang B, Zheng X, Ma P, Farley WJ, Siemasko KS, Niederkorn JY, Stern ME, Li DQ and Pflugfelder SC. IL-17 disrupts corneal barrier following desiccating stress. Mucosal Immunol. 2009; 2:243-253
10. McClellan AJ, Volpe EA, Zhang X, Darlington GJ, Li DQ, Pflugfelder SC and de Paiva CS. Ocular Surface Disease and Dacryoadenitis in Aging C57BL/6 Mice. Am J Pathol. 2014; 184:631-643. .
11. Chauhan SK, El AJ, Ecoiffier T, Goyal S, Zhang Q, Saban DR and Dana R. Autoimmunity in dry eye is due to resistance of Th17 to Treg suppression. JImmunol. 2009; 182:1247-1252.
12. Pflugfelder SC, De Paiva CS, Moore QL, Volpe EA, Li DQ, Gumus K, Zaheer ML and Corrales RM. Aqueous Tear Deficiency Increases Conjunctival Interferon-gamma (IFN-gamma) Expression and Goblet Cell Loss. Invest Ophthalmol Vis Sci. 2015; 56:7545-7550.
13. De Benedictis G, Carrieri G, Varcasia O, BonaFe M and Franceschi C. Inherited variability of the mitochondrial genome and successful aging in humans. AnnNYAcadSci. 2000; 908:208-218.
14. Goronzy J, Weyand, CM. Aging, autoimmunity and arthritis: T-cell senescence and contraction of T-cell repertoire diversity - catalysts of autoimmunity and chronic inflammation. Arthritis Res Ther. 2003; 5:225-234.
15. Brookes SM, Price EJ, Venables PJ and Maini RN. Interferon-gamma and epithelial cell activation in Sjogren’s syndrome. Br J Rheumatol. 1995; 34:226-231.
16. Kullberg MC, Jankovic D, Feng CG, Hue S, Gorelick PL, McKenzie BS, Cua DJ, Powrie F, Cheever AW, Maloy KJ and Sher A. IL-23 plays a key role in Helicobacter hepaticus-induced T cell-dependent colitis. J Exp Med. 2006; 203:2485-2494.
17. Kang SS, Bloom SM, Norian LA, Geske MJ, Flavell RA, Stappenbeck TS and Allen PM. An antibiotic-responsive mouse model of fulminant ulcerative colitis. PLoS Med. 2008; 5:e41.
18. Bouma G and Strober W. The immunological and genetic basis of inflammatory bowel disease. Nat Rev Immunol. 2003; 3:521-533.
19. Bandres E, Merino, J, Vazquez, B, Inoges, S, Moreno, C, Subira, ML, Sanchez-Ibarrola, A. The increase of IFN-g production through aging correlates with the expanded CD8(+high)CD28(-)CD57(+) subpopulation. Clin Immunol. 2000; 96:230-235.
20. Williams GP, Denniston AK, Oswal KS, Tomlins PJ, Barry RJ, Rauz S and Curnow SJ. The dominant human conjunctival epithelial CD8alphabeta+ T cell population is maintained with age but the number of CD4+ T cells increases. Age (Dordr). 2012; 34:1517-1528. Epub 2011 Sep 1527.
21. de Paiva CS, Villarreal AL, Corrales RM, Rahman HT, Chang VY, Farley WJ, Stern ME, Niederkorn JY, Li DQ and Pflugfelder SC. Dry Eye-Induced Conjunctival Epithelial Squamous Metaplasia Is Modulated by Interferon-{gamma}. Invest Ophthalmol Vis Sci. 2007; 48:2553-2560.
22. Zhang X, Chen W, de Paiva CS, Volpe EA, Gandhi NB, Farley WJ, Li DQ, Niederkorn JY, Stern ME and Pflugfelder SC. Desiccating Stress Induces CD4(+) T-Cell-Mediated Sjogren’s Syndrome-Like Corneal Epithelial Apoptosis via Activation of the Extrinsic Apoptotic Pathway by Interferon-gamma. Am J Pathol. 2011; 179:1807-1814.
23. Zhang X, Chen W, de Paiva CS, Corrales RM, Volpe EA, McClellan AJ, Farley WJ, Li DQ and Pflugfelder SC. Interferon-gamma exacerbates dry eye-induced apoptosis in conjunctiva through dual apoptotic pathways. Invest Ophthalmol Vis Sci. 2011; 52:6279-6285.
24. Zhang X, de Paiva CS, Su Z, Volpe EA, Li DQ and Pflugfelder SC. Topical interferon-gamma neutralization prevents conjunctival goblet cell loss in experimental murine dry eye. Exp Eye Res 2014; 118:117-124.
25. Coursey TG, Bohat R, Barbosa FL, Pflugfelder SC and de Paiva CS. Desiccating stress-induced chemokine expression in the epithelium is dependent on upregulation of NKG2D/RAE-1 and release of IFN-gamma in experimental dry eye. J Immunol. 2014; 193:5264-5272.
26. Chen Y, Chauhan SK, Saban DR, Sadrai Z, Okanobo A and Dana R. Interferon-{gamma}-secreting NK cells promote induction of dry eye disease. J Leukoc Biol. 2011; 89:965-972.
27. Coursey TG, Henriksson JT, Barbosa FL, de Paiva CS and Pflugfelder SC. Interferon-gamma-Induced Unfolded Protein Response in Conjunctival Goblet Cells as a Cause of Mucin Deficiency in Sjogren Syndrome. Am J Pathol. 2016; S0002-9440:30005-30000.
28. Lam H, Blieden L, de Paiva CS, Farley WJ, Stern ME and Pflugfelder SC. Tear Cytokine Profiles in Dysfunctional Tear Syndrome Am J Ophthalmol. 2009; 147:198-205.
29. Lowes MA, Kikuchi T, Fuentes-Duculan J, Cardinale I, Zaba LC, Haider AS, Bowman EP and Krueger JG. Psoriasis vulgaris lesions contain discrete populations of Th1 and Th17 T cells. J Invest Dermatol. 2008; 128:1207-1211.
30. Ogawa A, Andoh A, Araki Y, Bamba T and Fujiyama Y. Neutralization of interleukin-17 aggravates dextran sulfate sodium-induced colitis in mice. Clin Immunol. 2004; 110:55-62.
31. Yang W, Yao Y, Yang YQ, Lu FT, Li L, Wang YH, Nakajima T, Tsuneyama K, Ridgway WM, Gershwin ME and Lian ZX. Differential modulation by IL-17A of Cholangitis versus Colitis in IL-2Ralpha deleted mice. PLoS One. 2014; 9:e105351.
32. Nguyen CQ, Hu MH, Li Y, Stewart C and Peck AB. Salivary gland tissue expression of interleukin-23 and interleukin-17 in Sjogren’s syndrome: findings in humans and mice. Arthritis Rheum. 2008; 58:734-743.
33. de Paiva CS, Hwang CS, Pitcher JD, III, Pangelinan SB, Rahimy E, Chen W, Yoon KC, Farley WJ, Niederkorn JY, Stern ME, Li DQ and Pflugfelder SC. Age-related T-cell cytokine profile parallels corneal disease severity in Sjogren’s syndrome-like keratoconjunctivitis sicca in CD25KO mice. Rheumatology. 2010; 49:246-258.
34. Lin X, Rui K, Deng J, Tian J, Wang X, Wang S, Ko KH, Jiao Z, Chan VS, Lau CS, Cao X and Lu L. Th17 cells play a critical role in the development of experimental Sjogren’s syndrome. Ann Rheum Dis. 2014.
35. Raphael M, Bellefqih S, Piette JC, Le HP, Debre P and Chomette G. Conjunctival biopsy in Sjogren’s syndrome: correlations between histological and immunohistochemical features. Histopathology. 1988; 13:191-202.
36. Pflugfelder SC, Tseng SCG, Yoshino K, Monroy D, Felix C and Reis BL. Correlation of goblet cell density and mucosal epithelial membrane mucin expression with rose bengal staining in patients with ocular irritation. Ophthalmology. 1997; 104:223-235.
37. Mo F, Proia, AD, Johnson, WH, Cyr, D, Lashkari, K. Interferon gamma-inducible protein-10 (IP-10) and eotaxin as biomarkers in age-related macular degeneration. Invest Ophthalmol Vis Sci. 2010; 51:4226-4236.
38. Harper E, Guo, C, Rizzo, H, Lillis, JV, Kurtz, SE, Skorcheva, I, Purdy, D, Fitch, E, Iordanov, M, Blauvelt, A. Th17 cytokines stimulate CCL20 expression in keratinocytes in vitro and in vivo: implications for psoriasis pathogenesis. J Invest Dermatol. 2009; 129:2175-2183.
39. Reboldi A, Coisne, C, Baumjohann, D, Benvenuto, F, Bottinelli, D, Lira, S, Uccelli, A, Lanzavecchia, A, Engelhardt, B, Sallusto, F. C-C chemokine receptor 6-regulated entry of TH-17 cells into the CNS through the choroid plexus is required for the initiation of EAE. Nature Immunol. 2009; 10:514-523.
40. de Paiva CS, Raince JK, McClellan AJ, Shanmugam KP, Pangelinan SB, Volpe EA, Corrales RM, Farley WJ, Corry DB, Li DQ and Pflugfelder SC. Homeostatic control of conjunctival mucosal goblet cells by NKT-derived IL-13. Mucosal Immunol. 2011; 4:397-408. .
41. Tukler Henriksson J, Coursey TG, Corry DB, De Paiva CS and Pflugfelder SC. IL-13 Stimulates Proliferation and Expression of Mucin and Immunomodulatory Genes in Cultured Conjunctival Goblet Cells. Invest Ophthalmol Vis Sci. 2015; 56:4186-4197.
42. Pflugfelder SC, Farley W, Luo L, Chen LZ, de Paiva CS, Olmos LC, Li DQ and Fini ME. Matrix metalloproteinase-9 knockout confers resistance to corneal epithelial barrier disruption in experimental dry eye. Am J Pathol. 2005; 166:61-71.
43. Bourcier T, Acosta MC, Borderie V, Borras F, Gallar J, Bury T, Laroche L and Belmonte C. Decreased corneal sensitivity in patients with dry eye. Invest Ophthalmol Vis Sci. 2005; 46:2341-2345.
44. Ding J, Sullivan, DA. Aging and dry eye disease. Exp Gerontol. 2012; 47:483-490.
45. Haugen AJ, Peen E, Hulten B, Johannessen AC, Brun JG, Halse AK and Haga HJ. Estimation of the prevalence of primary Sjogren’s syndrome in two age-different community-based populations using two sets of classification criteria: the Hordaland Health Study. Scand J Rheumatol. 2008; 37:30-34.
46. Kosrirukvongs P, Ngowyutagon P, Pusuwan P, Koolvisoot A and Nilganuwong S. Prevalence of dry eye syndrome and Sjogren’s syndrome in patients with rheumatoid arthritis. J Med Assoc Thai. 2012; 95 Suppl 4:S61-69.
47. Moss SE, Klein R and Klein BE. Incidence of dry eye in an older population. Arch Ophthalmol. 2004; 122:369-373.
48. Adams AD. The morphology of human conjunctival mucus. ArchOphthalmol. 1979; 97:730-734.
49. Ralph RA. Conjunctival goblet cell density in normal subjects and in dry eye syndromes. Invest Ophthalmol. 1975; 14:299-302.
50. Kondo M, Tamaoki J, Takeyama K, Isono K, Kawatani K, Izumo T and Nagai A. Elimination of IL-13 reverses established goblet cell metaplasia into ciliated epithelia in airway epithelial cell culture. Allergol Int. 2006; 55:329-336.
51. Marillier R, Michels, C, Smith, EM, Fick, LC, Leeto, M, Dewals, B, Horsnell, WG, Brombacher, F. IL-4/IL-13 independent goblet cell hyperplasia in experimental helminth infections. BMC Immunol. 2008; 9:11.
52. Kondo M, Tamaoki J, Takeyama K, Nakata J and Nagai A. Interleukin-13 induces goblet cell differentiation in primary cell culture from Guinea pig tracheal epithelium. Am J Respir Cell Mol Biol. 2002; 27:536-541.
53. Laoukili J, Perret E, Willems T, Minty A, Parthoens E, Houcine O, Coste A, Jorissen M, Marano F, Caput D and Tournier F. IL-13 alters mucociliary differentiation and ciliary beating of human respiratory epithelial cells. J Clin Invest. 2001; 108:1817-1824.
54. Huang MC, Liao JJ, Bonasera S, Longo DL and Goetzl EJ. Nuclear factor-kappaB-dependent reversal of aging-induced alterations in T cell cytokines. FASEB J. 2008; 22:2142-2150.
55. Schmitt V, Rink L and Uciechowski P. The Th17/Treg balance is disturbed during aging. Exp Gerontol. 2013; 48:1379-1386.
56. Ouyang X, Yang Z, Zhang R, Arnaboldi P, Lu G, Li Q, Wang W, Zhang B, Cui M, Zhang H, Liang-Chen J, Qin L, Zheng F, Huang B and Xiong H. Potentiation of Th17 cytokines in aging process contributes to the development of colitis. Cell Immunol. 2011; 266:208-217.
57. Kang JY, Lee SY, Rhee CK, Kim SJ, Kwon SS and Kim YK. Effect of aging on airway remodeling and muscarinic receptors in a murine acute asthma model. Clin Interv Aging. 2013; 8:1393-1403.
58. Sylvester J, Liacini A, Li WQ and Zafarullah M. Interleukin-17 signal transduction pathways implicated in inducing matrix metalloproteinase-3, -13 and aggrecanase-1 genes in articular chondrocytes. Cell Signal. 2004; 16:469-476.
59. Cortez DM, Feldman MD, Mummidi S, Valente AJ, Steffensen B, Vincenti M, Barnes JL and Chandrasekar B. IL-17 stimulates MMP-1 expression in primary human cardiac fibroblasts via p38. Am J Physiol Heart Circ Physiol. 2007; 293:H3356-H3365.
60. Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM and Weaver CT. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol. 2005; 6:1123-1132.
61. Park H, Li Z, Yang XO, Chang SH, Nurieva R, Wang YH, Wang Y, Hood L, Zhu Z, Tian Q and Dong C. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17. Nat Immunol. 2005; 6:1133-1141.
62. Pelegrino FS, Volpe EA, Gandhi NB, Li DQ, Pflugfelder SC and de Paiva CS. Deletion of interferon-gamma delays onset and severity of dacryoadenitis in CD25KO mice. Arthritis Res Ther. 2012; 14:R234.
63. Cha S, Brayer J, Gao J, Brown V, Killedar S, Yasunari U and Peck AB. A dual role for interferon-gamma in the pathogenesis of Sjogren’s syndrome-like autoimmune exocrinopathy in the nonobese diabetic mouse. Scand J Immunol. 2004; 60:552-565.
64. Kelchtermans H, Schurgers, E, Geboes, L, Mitera, T, Van Damme, J, Van Snick, J, Uyttenhove, C, Matthys, P. Effector mechanisms of interleukin-17 in collagen-induced arthritis in the absence of interferon- and counteraction by interferon-. Arthritis Res Ther. 2009; 11:R122.
65. Luger D, Silver PB, Tang J, Cua D, Chen Z, Iwakura Y, Bowman EP, Sgambellone NM, Chan CC and Caspi RR. Either a Th17 or a Th1 effector response can drive autoimmunity: conditions of disease induction affect dominant effector category. J Exp Med. 2008; 205:799-810.
66. Billiau A, Heremans H, Vandekerckhove F, Dijkmans R, Sobis H, Meulepas E and Carton H. Enhancement of experimental allergic encephalomyelitis in mice by antibodies against IFN-gamma. JImmunol. 1988; 140:1506-1510.
67. Caspi RR, Chan CC, Grubbs BG, Silver PB, Wiggert B, Parsa CF, Bahmanyar S, Billiau A and Heremans H. Endogenous systemic IFN-gamma has a protective role against ocular autoimmunity in mice. JImmunol. 1994; 152:890-899.
68. Chu C-Q, Swart, D, Alcorn, D, Tocker, J, Elkon, KB. Interferon-g regulates susceptibility to collagen-induced arthritis through suppression of interleukin-17. Arthritis Rheum. 2007; 56:1145-1151.
69. Gao Y, Min K, Zhang Y, Su J, Greenwood M and Gronert K. Female-Specific Downregulation of Tissue Polymorphonuclear Neutrophils Drives Impaired Regulatory T Cell and Amplified Effector T Cell Responses in Autoimmune Dry Eye Disease. J Immunol. 2015; 195:3086-3099.
70. Meadows JF, Dionne K and Nichols KK. Differential Profiling of T-Cell Cytokines as Measured by Protein Microarray Across Dry Eye Subgroups. Cornea. 2016; 35:329-335.
71. Enriquez-de-Salamanca A, Castellanos E, Stern ME, Fernandez I, Carreno E, Garcia-Vazquez C, Herreras JM and Calonge M. Tear cytokine and chemokine analysis and clinical correlations in evaporative-type dry eye disease. MolVis. 2010; 16:862-873.
72. Nguyen CQ, Yin H, Lee BH, Carcamo WC, Chiorini JA and Peck AB. Pathogenic effect of interleukin-17A in induction of Sjogren’s syndrome-like disease using adenovirus-mediated gene transfer. Arthritis ResTher. 2010; 12:R220.
73. McCarty CA, Bansal AK, Livingston PM, Stanislavsky YL and Taylor HR. The epidemiology of dry eye in Melbourne, Australia. Ophthalmology. 1998; 105:1114-1119.
74. Pflugfelder SC. Prevalence, burden, and pharmacoeconomics of dry eye disease. AmJManagCare. 2008; 14:S102-S106.
75. Schein OD, Munoz B, Tielsch JM, Bandeen-Roche K and West S. Prevalence of dry eye among the elderly. AmJOphthalmol. 1997; 124:723-728.