
INTRODUCTION
Pancreatic cancer (PC) is the fifteenth most common cancer in the world with extremely low 5-year survival rate at only ~5% [1]. Despite decades of efforts, there are still few early detection methods and effective treatments. These grim facts highlight pressing need to identify the risk factors of PC for primary prevention and targeted therapy. Many environmental factors might influence an individual’s PC risk [2, 3], among which cigarette smoking has been confirmed as the surest to date. Both current and former smokers have higher risk than non-smokers [4]. Approximately 25% of PC cases are attributable to smoking [5, 6]. Individuals with family history are prone to PC and risk rises with the increasing number of affected first-degree relatives [7], which indicates inherited genetic background may impact on susceptibility to PC. However, only a small part of sporadic PCs can be explained by the genetic variants identified by recent genome wide association studies [8–14], while some true susceptibility loci with moderate P values might have been overlooked due to stringent P-value threshold [15]. Therefore, the genetic susceptibility to sporadic PC is definitely worth exploring.
STAT3 is a member of the STAT protein family, which are mainly phosphorylated by the receptor or non-receptor associated kinases, in response to cytokines and growth factors, and then form homo- or heterodimers that translocate to the cell nucleus where they act as transcription activators. They regulate cytokine-dependent inflammation and immunity, determining whether immune responses in the tumor microenvironment promote or inhibit cancer [16].
STAT3 plays a central role in many inflammatory pathways including nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and interleukin-6 (IL-6)-GP130-Janus kinase (JAK) pathways [16]. Considering that deregulation of transcriptional control is a central characteristic of cancers [17], and inflammation is a defined cancer-causing factor [18], it is reasonable to presume that alteration of STAT3 may be involved in the process of carcinogenesis. However, the roles it plays may be complicated and two-sided. In some circumstances, STAT3 is regarded as an oncogene [19–24], while in others its tumor suppressing and prognosis ameliorating effects have also been identified [25–28].
In pancreatic ductal adenocarcinomas (PDACs), constitutive activation of STAT3 by phosphorylation of Tyr705 has been reported in 30% to 100% of human tumor specimens, as well as in many PDAC cell lines [29]. Notably, epithelial Stat3 is required particularly during the progression of PanIN to PDAC, but not in the initiation of PanIN in mouse model [30]. Based on these findings, we hypothesized that common variants in STAT3 may contribute to the susceptibility to PC. Since 3′UTR often contains regulatory elements that post-transcriptionally regulate gene expression, we aimed at finding functional variants in this region. Thus we conducted two stage case-control studies to evaluate the correlation between two common variants (rs1053004, rs1053005) in 3′UTR of STAT3, which were predicted most likely to have potential function, and risk of PC. Furthermore, luciferase reporter gene assays were performed for function verification.
RESULTS
Characteristics of population
A total of 774 cases and 777 controls passed quality control of stage one, and 940 cases and 1398 controls were included in stage two. The characteristics of the subjects are summarized in Table 1. The majority of cases were males, with 57.4% in stage one and 79.4% in stage two. The median ages of case/control groups in two stages were 60/59 and 61/60, respectively. Cases and controls were adequately matched in sex and age in both stages.
Table 1: Characteristics of subjects in this study
|
Stage one |
Stage two |
||||||
|---|---|---|---|---|---|---|---|---|
Variables |
Case N(%) |
Control N (%) |
χ2 |
P |
Case N(%) |
Control N (%) |
χ2 |
P |
Total |
774 |
777 |
|
|
940 |
1398 |
|
|
Sex |
|
|
0.045 |
0.833 |
|
|
0.657 |
0.674 |
Male |
444 (57.4) |
450 (57.9) |
|
|
746 (79.4) |
1120 (80.1) |
|
|
Female |
330 (42.6) |
327 (42.1) |
|
|
194 (20.6) |
278 (19.9) |
|
|
Age(Median) |
60 |
59 |
1.240 |
0.743 |
61 |
60 |
0.470 |
0.925 |
< 45 |
94 (12.1) |
94 (12.1) |
|
|
108 (11.5) |
148 (10.6) |
|
|
45~55 |
163 (21.1) |
159 (20.5) |
|
|
184 (19.6) |
277 (19.8) |
|
|
55~65 |
252 (32.6) |
273 (35.1) |
|
|
311 (33.1) |
467 (33.4) |
|
|
> 65 |
265 (34.2) |
251 (32.3) |
|
|
337 (35.9) |
506 (36.2) |
|
|
Association analysis
Both of the two SNPs (rs1053004 and rs1053005) were successfully genotyped. The call rates in both stages were > 95%, and the genotypes in controls conformed to HWE (P > 0.05). In stage one, only rs1053004 T > C polymorphism was found to be significantly associated with a decreased PC risk (CC versus TT: odds ratio (OR) (95% confidence interval (CI)) = 0.71 (0.52–0.96); P = 0.025). While no significant association was found between genotypes of rs1053005 and PC risk (CC versus TT: OR (95% CI) = 0.80 (0.57–1.11); P = 0.176). Rs1053004 was further replicated in stage two (CC versus TT: OR (95% CI) = 0.70 (0.53–0.93); P = 0.013). Consistently, individuals carrying rs1053004 CC genotype had lower risk of PC compared with the TT genotype (OR (95% CI) = 0.72 (0.59–0.88); P = 0.002) in the combined samples. And rs1053004 also showed to be significantly associated with PC risk in additive model in both two stages (OR (95% CI) stage one = 0.85 (0.73–0.98); OR (95% CI) stage two = 0.86 (0.76–0.97)) and combined samples (OR (95% CI) combined = 0.86 (0.78–0.94)). Two other models (dominant, recessive models) were also demonstrated in combined samples. The genotype frequencies and detailed statistical results of rs1053004 and rs1053005 are summarized in Table 2 and Supplementary Table S1, respectively. Also, results of stratified analysis of association between rs1053004 and risk of PC by gender was presented in Supplementary Table S2.
Table 2: Association between rs1053004 and risk of PC
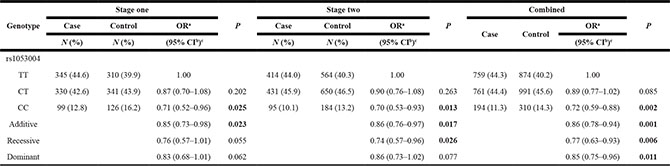
aOR, odds ratio.
bCI, confidence interval.
cOR estimated with logistic regression adjusted for sex and age.
Dual luciferase reporter gene assays
We constructed two luciferase reporter plasmids containing rs1053004 T and C allele, respectively. In all the three PC cell lines, Panc-1, BxPC-3 and SW1990, luciferase activity was significantly higher in the C allelic plasmid compared with the T plasmid (P < 0.05, Figure 1). The results suggested that rs1053004 T > C in 3′UTR could up-regulate gene expression post-transcriptionally.
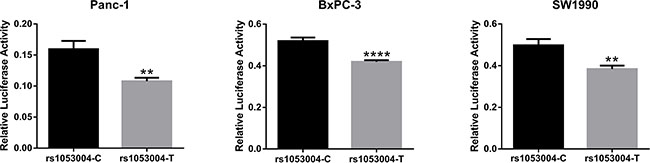
Figure 1: Dual luciferase reporter gene assays: the effects of rs1053004 on gene expression. The figure showed that compared to the construct with rs1053004-T, the construct with rs1053004-C had significantly higher luciferase activity in three PC cell lines. Unpaired Student’s t-test was used to evaluate the differences and p values less than 0.05 was considered significant with ** indicating p < 0.01 and **** indicating p < 0.001.
DISCUSSION
STAT3 plays a pivotal role in a multitude of physiological and pathological processes [31–36]. Its abnormal activity and expression have been found in PC [24, 29, 30, 37]. Nevertheless, whether genetic variations in this gene affect PC risk has been barely investigated. In our current study, using two stage case-control studies integrating luciferase reporter gene assays, we found for the first time that rs1053004 T > C in 3′UTR of STAT3 might decrease risk of PC through up-regulating STAT3 expression.
STAT3, a member of a transcription factor family, was first identified in 1994 as an IL-6-activated acute-phase response factor (APRF) [38]. Other from its transient activation in normal cells, STAT3 was often found aberrantly expressed and constitutively activated in a variety of malignancies [39], thereby was regarded as an oncogene in multiple cancers [20–23]. Nonetheless, the tumor suppressing role of STAT3 was also reported [25–28, 40, 41]. For example, STAT3 was found as a negative regulator of thyroid cancer since it could activate transcription of the tumor suppressor insulin-like growth factor binding protein 7 (IGFBP7), and negatively regulate aerobic glycolysis [26]. In the context of lung cancer, STAT3 prevented disease initiation by maintaining pulmonary homeostasis under oncogenic stress [28]. Besides, loss of IL-6/Stat3 signaling in prostate cancer might bypass senescence and accelerates cancer progression via disrupting the ARF-Mdm2-p53 tumor suppressor axis [27]. In addition to these malignancies, other cancers were reported to benefit from STAT3 in phenotype and prognosis as well, such as breast cancer [40, 42], head and neck neoplasms [25], and brain tumor [43, 44]. In respect of PC, STAT3 was suggested to be frequently over-expressed in vivo and vitro [24, 29, 37] and play a pivotal role in the carcinogenesis of PC, while its anti-tumor effect in PC is scarcely reported so far.
rs1053004 lies in the 3′UTR of STAT3, the mutant C allele is predicted to cause binding site loss for some miRNAs (such as hsa-miR-31-5p and hsa-miR-99b-3p, predicted by MicroSNiper web tool [45]). Consistently, dual luciferase reporter gene assays demonstrated in three different PC cell lines that C allele was significantly associated with higher luciferase activity. We therefore inferred that, in Chinese population, the C allele of rs1053004 astricted the binding of miRNA to STAT3, thus increasing the expression of this gene with potential benefit, and consequently conferred individuals with CC genotype lower risk to PC compared with TT genotypes, notwithstanding the exactly anti-PC mechanism of STAT3 still needs further investigation.
To the best of our knowledge, this is the first study to investigate the association between genetic variants in STAT3 and risk of PC. Our work has three major strengths. First, two stage case-control studies with large sample size offer us enough statistical power to identify the association. Second, functional validation experiments not only indicated that the association might be reliable, but also provided the plausible underlying mechanisms of how this variant influenced risk of PC. Last but not least, our work suggests that STAT3 might exert positive effects in preventing PC, particularly in Chinese population, despite the stereotype that STAT3 was an oncogene. However, functional experiments such as RNA interference were still needed for deeper mechanism exploration. Besides, environmental factors such as smoking and obesity were not included in our current study. Further researches concerning gene-environment interaction will be required.
In conclusion, we identified a regulatory SNP rs1053004 mapping to 3′UTR of STAT3 associated with a decreased risk of PC in Chinese population. Our findings could lead to new insights to the etiology of PC and provide a potential biomarker. Future studies conducted in different population and with deeper biological experiments are warranted to validate our results and more attention should be paid to the variants in STAT3 when investigating genetic susceptibility to PC.
MATERIALS AND METHODS
Study subjects
We carried out a two-stage case-control study to investigate the association between variants of STAT3 and risk of PC. Stage one included 774 cases and 777 cancer-free controls, part of which were also involved in our previous studies [46–49]. Patients were consecutively recruited from January 2009 to September 2014 at Tongji Hospital of Huazhong University of Science and Technology, Wuhan, China. While controls were volunteers randomly selected from healthy screenings over the same period. In stage two, 940 cases were enrolled at the Peking Union Medical College Hospital, Beijing from January 2008 to December 2012, and 1398 controls were cancer-free individuals from a community cancer screening program for early detection from the same region over the same period. All cases were individuals with newly diagnosed, pathologically confirmed and previously untreated primary PC. The diagnosis of PC was confirmed by histopathological or cytological analysis according to the World Health Organization classification. Controls were frequency matched with cases by sex and age (interval of 5 years). Demographic characteristics (sex, age) were obtained from medical records. At recruitment, a 2-mL peripheral venous blood sample was collected from each subject with informed consent. This study was approved by the ethics committee of Tongji Medical College of Huazhong University of Science and Technology and Peking Union Medical College Hospital.
SNP selection genotyping and statistical analysis
It was primarily acknowledged that the 3′UTR often contains regulatory elements that post-transcriptionally regulate gene expression. For example, miRNAs usually interact with 3′UTR of target mRNAs leading to mRNA degradation and/or translational repression [50]. Accordingly, our hypothesis was that the variants in 3′UTR of STAT3 might affect its expression and further concern PC risk. We utilized the University of California Santa Cruz (UCSC) Table Browser (http://genome.ucsc.edu/cgi-bin/hgTables) to retrieve variants in 3′UTR of STAT3, finding four SNPs (rs1053004, rs1053005, rs1053023 and rs3744483) with minor allele frequency greater than 0.05. Among which, rs1053005, rs1053023 and rs3744483 are in perfect linkage disequilibrium (LD) (r2 = 1, using 1000 genome phase 3 data). While the LD relations between rs1053004 and others are slightly weaker (r2 = 0.8). Then we utilized SNPinfo Web Server (http://snpinfo.niehs.nih.gov/snpinfo/snpfunc.htm) to predict the function of these polymorphisms. All of the four SNPs lie in putative microRNA targets, and rs1053005 is predicted to regulate more microRNAs′ binding than the other two SNPs in perfect LD (rs1053023 and rs3744483). Taken together, the results of LD analysis and functional prediction suggested both rs1053004 and rs1053005 as most promising SNPs, and to be subsequently genotyped.
Genotyping and statistical analysis
Genomic DNA was extracted from peripheral blood using RelaxGene Blood DNA System (Tiangen, DP319) according to the manufacturer’s instructions. Quantitative and qualitative DNA analysis was performed with Nanodrop.
The two candidate SNPs were genotyped using a TaqMan assay on the ABI PRISM 7900 HT platform (Applied Biosystems, Inc.) in two stages. Genotyping analysis was conducted with Sequence Detection System (SDS) Software v2.4.1. For quality control, 5% duplicate samples were independently reanalyzed in a blinded fashion. The call rate of candidate SNPs were over 95%.
Deviation from Hardy-Weinberg equilibrium (HWE) in controls was examined by a goodness-of-fit χ2 test. Demographic characters and genotype frequencies differences were assessed by Pearson’sχ2 test. Odds ratios (ORs) and 95% confidence intervals (CIs) were calculated applying unconditional logistic regression analysis adjusting for age and sex. For all analyses, statistical significance was set at P < 0.05, and all tests were two sided. All statistical analyses were conducted by SPSS Statistics 20.0.
Dual luciferase reporter gene assays
We chose psiCHECK™-2 Vector (Promega) to construct luciferase reporter gene plasmid since it is an ideal biosensor for miRNA-target interaction. This vector contains two kinds of luciferase gene, Renilla and Firefly. The former’s activity reflected STAT3 expression, as the 3′ UTR of which was appended to the Renilla luciferase gene. Inclusion of the firefly luciferase reporter served as the internal control. The whole 3′UTRs of STAT3 containing different alleles of SNP were cloned into the vectors (Genewiz). The reporter constructs were verified for sequence. Human pancreatic cancer cell lines Panc-1, BxPC-3 and SW1990 were used for luciferase reporter gene assays. The constructed reporter plasmid was transfected to the cells cultured in Dulbecco’s Modified Eagle Medium (DMEM) with high glucose (Gibco) supplemented with 10% fetal bovine serum (Gibco) and 1% Penicillin-Streptomycin (Gibco) under 37°C in humidified atmosphere containing 5% CO2 using Attractene Transfection Reagent (QIAGEN). After 24 hours’ incubation, the cells were assayed for two luciferase activity with the Dual-Luciferase Reporter Assay System (Promega). Relative activities were evaluated by calculating the ratio of Renilla to Firefly. For each plasmid, three independent transfection experiments were performed with each in triplicate. The differences in reporter gene expression were examined by Student’s t-test.
ACKNOWLEDGMENTS
The authors wish to appreciate all the study participants, research staff, and students who participated in this work, especially blood sample donors.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
GRANT SUPPORT
This work is supported by National Program for Support of Top-notch Young Professionals, National High-Tech Research and Development Program of China 2014AA020609 and Specialized Research Fund for the Doctoral Program of Higher Education 20130142110017 for Xiaoping Miao.
REFERENCES
1. Wolfgang CL, Herman JM, Laheru DA, Klein AP, Erdek MA, Fishman EK, Hruban RH. Recent progress in pancreatic cancer. CA Cancer J Clin. 2013; 63:318–348.
2. Zhu B, Zou L, Han J, Chen W, Shen N, Zhong R, Li J, Chen X, Liu C, Shi Y, Miao X. Parity and pancreatic cancer risk: evidence from a meta-analysis of twenty epidemiologic studies. Sci Rep. 2014; 4:5313.
3. Lowenfels AB, Maisonneuve P. Epidemiology and risk factors for pancreatic cancer. Best pract Res Cl G. 2006; 20:197–209.
4. Iodice S, Gandini S, Maisonneuve P, Lowenfels AB. Tobacco and the risk of pancreatic cancer: a review and meta-analysis. Langenbeck Arch Su. 2008; 393:535–545.
5. Maisonneuve P, Lowenfels AB. Epidemiology of pancreatic cancer: an update. Digest Dis. 2010; 28:645–656.
6. Zou L, Zhong R, Shen N, Chen W, Zhu B, Ke J, Lu X, Zhang T, Lou J, Wang Z, Liu L, Qi L, Miao X. Non-linear dose-response relationship between cigarette smoking and pancreatic cancer risk: evidence from a meta-analysis of 42 observational studies. Eur J Cancer. 2014; 50:193–203.
7. Tersmette AC, Petersen GM, Offerhaus GJ, Falatko FC, Brune KA, Goggins M, Rozenblum E, Wilentz RE, Yeo CJ, Cameron JL, Kern SE, Hruban RH. Increased risk of incident pancreatic cancer among first-degree relatives of patients with familial pancreatic cancer. Clin cancer Res. 2001; 7:738–744.
8. Wu C, Miao X, Huang L, Che X, Jiang G, Yu D, Yang X, Cao G, Hu Z, Zhou Y, Zuo C, Wang C, Zhang X, et al. Genome-wide association study identifies five loci associated with susceptibility to pancreatic cancer in Chinese populations. Nat Genet. 2012; 44:62–66.
9. Petersen GM, Amundadottir L, Fuchs CS, Kraft P, Stolzenberg-Solomon RZ, Jacobs KB, Arslan AA, Bueno-de-Mesquita HB, Gallinger S, Gross M, Helzlsouer K, Holly EA, Jacobs EJ, et al. A genome-wide association study identifies pancreatic cancer susceptibility loci on chromosomes 13q22.1, 1q32.1 and 5p15.33. Nat Genet. 2010; 42:224–228.
10. Amundadottir L, Kraft P, Stolzenberg-Solomon RZ, Fuchs CS, Petersen GM, Arslan AA, Bueno-de-Mesquita HB, Gross M, Helzlsouer K, Jacobs EJ, LaCroix A, Zheng W, Albanes D, et al. Genome-wide association study identifies variants in the ABO locus associated with susceptibility to pancreatic cancer. Nat Genet. 2009; 41:986–990.
11. Wolpin BM, Rizzato C, Kraft P, Kooperberg C, Petersen GM, Wang Z, Arslan AA, Beane-Freeman L, Bracci PM, Buring J, Canzian F, Duell EJ, Gallinger S, et al. Genome-wide association study identifies multiple susceptibility loci for pancreatic cancer. Nat Genet. 2014; 46:994–1000.
12. Low SK, Kuchiba A, Zembutsu H, Saito A, Takahashi A, Kubo M, Daigo Y, Kamatani N, Chiku S, Totsuka H, Ohnami S, Hirose H, Shimada K, et al. Genome-wide association study of pancreatic cancer in Japanese population. PloS One. 2010; 5:e11824.
13. Diergaarde B, Brand R, Lamb J, Cheong SY, Stello K, Barmada MM, Feingold E, Whitcomb DC. Pooling-based genome-wide association study implicates gamma-glutamyltransferase 1 (GGT1) gene in pancreatic carcinogenesis. Pancreatology. 2010; 10:194–200.
14. Childs EJ, Mocci E, Campa D, Bracci PM, Gallinger S, Goggins M, Li D, Neale RE, Olson SH, Scelo G, Amundadottir LT, Bamlet WR, Bijlsma MF, et al. Common variation at 2p13.3, 3q29, 7p13 and 17q25.1 associated with susceptibility to pancreatic cancer. Nat Genet. 2015; 47:911–916.
15. Huang L, Wu C, Yu D, Wang C, Che X, Miao X, Zhai K, Chang J, Jiang G, Yang X, Cao G, Hu Z, Zhou Y, et al. Identification of common variants in BRCA2 and MAP2K4 for susceptibility to sporadic pancreatic cancer. Carcinogenesis. 2013; 34:1001–1005.
16. Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer. 2009; 9:798–809.
17. Gallinari P, Di Marco S, Jones P, Pallaoro M, Steinkuhler C. HDACs, histone deacetylation and gene transcription: from molecular biology to cancer therapeutics. Cell Res. 2007; 17:195–211.
18. Okada F. Inflammation-Related Carcinogenesis: Current Findings in Epidemiological Trends, Causes and Mechanisms. Yonago Acta Med. 2014; 57:65–72.
19. Corcoran RB, Contino G, Deshpande V, Tzatsos A, Conrad C, Benes CH, Levy DE, Settleman J, Engelman JA, Bardeesy N. STAT3 plays a critical role in KRAS-induced pancreatic tumorigenesis. Cancer Res. 2011; 71:5020–5029.
20. Bollrath J, Phesse TJ, von Burstin VA, Putoczki T, Bennecke M, Bateman T, Nebelsiek T, Lundgren-May T, Canli O, Schwitalla S, Matthews V, Schmid RM, Kirchner T, et al. gp130-mediated Stat3 activation in enterocytes regulates cell survival and cell-cycle progression during colitis-associated tumorigenesis. Cancer Cell. 2009; 15:91–102.
21. Grivennikov S, Karin E, Terzic J, Mucida D, Yu GY, Vallabhapurapu S, Scheller J, Rose-John S, Cheroutre H, Eckmann L, Karin M. IL-6 and Stat3 are required for survival of intestinal epithelial cells and development of colitis-associated cancer. Cancer Cell. 2009; 15:103–113.
22. Bronte-Tinkew DM, Terebiznik M, Franco A, Ang M, Ahn D, Mimuro H, Sasakawa C, Ropeleski MJ, Peek RM, Jr. and Jones NL. Helicobacter pylori cytotoxin-associated gene A activates the signal transducer and activator of transcription 3 pathway in vitro and in vivo. Cancer Res. 2009; 69:632–639.
23. Gao SP, Mark KG, Leslie K, Pao W, Motoi N, Gerald WL, Travis WD, Bornmann W, Veach D, Clarkson B, Bromberg JF. Mutations in the EGFR kinase domain mediate STAT3 activation via IL-6 production in human lung adenocarcinomas. J Clin Invest. 2007; 117:3846–3856.
24. Akihisa Fukuda SCW, John P. Morris, Alexandra E. Folias, Angela Liou, Graece E. Kim, Shizuo AKira, Kenneth M. Boucher, Matthew A. Firpo, Sean J. Mulvihill, Matthias Hebrok. Stat3 and MMP7 Contribute to Pancreatic Adenocarcinama Initiation and Progression. Cancer Cell. 2011; 19:441–455.
25. Pectasides E, Egloff AM, Sasaki C, Kountourakis P, Burtness B, Fountzilas G, Dafni U, Zaramboukas T, Rampias T, Rimm D, Grandis J, Psyrri A. Nuclear localization of signal transducer and activator of transcription 3 in head and neck squamous cell carcinoma is associated with a better prognosis. Clin Cancer Res. 2010; 16:2427–2434.
26. Couto JP, Daly L, Almeida A, Knauf JA, Fagin JA, Sobrinho-Simoes M, Lima J, Maximo V, Soares P, Lyden D, Bromberg JF. STAT3 negatively regulates thyroid tumorigenesis. P Natil Acad Sci USA. 2012; 109:E2361–2370.
27. Pencik J, Schlederer M, Gruber W, Unger C, Walker SM, Chalaris A, Marie IJ, Hassler MR, Javaheri T, Aksoy O, Blayney JK, Prutsch N, Skucha A, et al. STAT3 regulated ARF expression suppresses prostate cancer metastasis. Nat Commun. 2015; 6:7736.
28. Zhou J, Qu Z, Yan S, Sun F, Whitsett JA, Shapiro SD, Xiao G. Differential roles of STAT3 in the initiation and growth of lung cancer. Oncogene. 2015; 34:3804–3814.
29. Scholz A, Heinze S, Detjen KM, Peters M, Welzel M, Hauff P, Schirner M, Wiedenmann B, Rosewicz S. Activated signal transducer and activator of transcription 3 (STAT3) supports the malignant phenotype of human pancreatic cancer. Gastroenterology. 2003; 125:891–905.
30. Lesina M, Kurkowski MU, Ludes K, Rose-John S, Treiber M, Kloppel G, Yoshimura A, Reindl W, Sipos B, Akira S, Schmid RM, Algul H. Stat3/Socs3 activation by IL-6 transsignaling promotes progression of pancreatic intraepithelial neoplasia and development of pancreatic cancer. Cancer Cell. 2011; 19:456–469.
31. Takeda K, Kaisho T, Yoshida N, Takeda J, Kishimoto T, Akira S. Stat3 activation is responsible for IL-6-dependent T cell proliferation through preventing apoptosis: generation and characterization of T cell-specific Stat3-deficient mice. J Immunol. 1998; 161:4652–4660.
32. Chapman RS, Lourenco PC, Tonner E, Flint DJ, Selbert S, Takeda K, Akira S, Clarke AR, Watson CJ. Suppression of epithelial apoptosis and delayed mammary gland involution in mice with a conditional knockout of Stat3. Gene Dev. 1999; 13:2604–2616.
33. Dauer DJ, Ferraro B, Song L, Yu B, Mora L, Buettner R, Enkemann S, Jove R, Haura EB. Stat3 regulates genes common to both wound healing and cancer. Oncogene. 2005; 24:3397–3408.
34. Barry SP, Townsend PA, McCormick J, Knight RA, Scarabelli TM, Latchman DS, Stephanou A. STAT3 deletion sensitizes cells to oxidative stress. Biochem Bioph Res C. 2009; 385:324–329.
35. Guo X, Qiu J, Tu T, Yang X, Deng L, Anders RA, Zhou L, Fu YX. Induction of innate lymphoid cell-derived interleukin-22 by the transcription factor STAT3 mediates protection against intestinal infection. Immunity. 2014; 40:25–39.
36. Resemann HK, Watson CJ, Lloyd-Lewis B. The Stat3 paradox: a killer and an oncogene. Mol Cell Endocrino. 2014; 382:603–611.
37. Wei D, Le X, Zheng L, Wang L, Frey JA, Gao AC, Peng Z, Huang S, Xiong HQ, Abbruzzese JL, Xie K. Stat3 activation regulates the expression of vascular endothelial growth factor and human pancreatic cancer angiogenesis and metastasis. Oncogene. 2003; 22:319–329.
38. Lutticken C, Wegenka UM, Yuan J, Buschmann J, Schindler C, Ziemiecki A, Harpur AG, Wilks AF, Yasukawa K, Taga T, et al. Association of transcription factor APRF and protein kinase Jak1 with the interleukin-6 signal transducer gp130. Science. 1994; 263:89–92.
39. Yu H, Jove R. The STATs of cancer—new molecular targets come of age. Nat Rev Cancer. 2004; 4:97–105.
40. Dolled-Filhart M, Camp RL, Kowalski DP, Smith BL, Rimm DL. Tissue microarray analysis of signal transducers and activators of transcription 3 (Stat3) and phospho-Stat3 (Tyr705) in node-negative breast cancer shows nuclear localization is associated with a better prognosis. Clin Cancer Res. 2003; 9:594–600.
41. de la Iglesia N, Puram SV, Bonni A. STAT3 regulation of glioblastoma pathogenesis. Curr Mol Med. 2009; 9:580–590.
42. Sonnenblick A, Uziely B, Nechushtan H, Kadouri L, Galun E, Axelrod JH, Katz D, Daum H, Hamburger T, Maly B, Allweis TM, Peretz T. Tumor STAT3 tyrosine phosphorylation status, as a predictor of benefit from adjuvant chemotherapy for breast cancer. Breast Cancer Res T. 2013; 138:407–413.
43. de la Iglesia N, Konopka G, Puram SV, Chan JA, Bachoo RM, You MJ, Levy DE, Depinho RA, Bonni A. Identification of a PTEN-regulated STAT3 brain tumor suppressor pathway. Gene Dev. 2008; 22:449–462.
44. de la Iglesia N, Konopka G, Lim KL, Nutt CL, Bromberg JF, Frank DA, Mischel PS, Louis DN, Bonni A. Deregulation of a STAT3-interleukin 8 signaling pathway promotes human glioblastoma cell proliferation and invasiveness. J NeuroSci. 2008; 28:5870–5878.
45. Barenboim M, Zoltick BJ, Guo Y, Weinberger DR. MicroSNiPer: a web tool for prediction of SNP effects on putative microRNA targets. Hum Mutat. 2010; 31:1223–1232.
46. Zhong R, Liu L, Zou L, Sheng W, Zhu B, Xiang H, Chen W, Chen J, Rui R, Zheng X, Yin J, Duan S, Yang B, et al. Genetic variations in the TGFbeta signaling pathway, smoking and risk of colorectal cancer in a Chinese population. Carcinogenesis. 2013; 34:936–942.
47. Zhu B, Tian J, Zhong R, Tian Y, Chen W, Qian J, Zou L, Xiao M, Shen N, Yang H, Lou J, Qiu Q, Ke J, et al. Genetic Variants in the SWI/SNF Complex and Smoking Collaborate to Modify the Risk of Pancreatic Cancer in a Chinese Population. Mol Carcinogen. 2014.
48. Gong J, Tian J, Lou J, Ke J, Li L, Li J, Yang Y, Gong Y, Zhu Y, Zhang Y, Zhong R, Chang J, Miao X. A functional polymorphism in lnc-LAMC2–1:1 confers risk of colorectal cancer by affecting miRNA binding. Carcinogenesis. 2016.
49. Tian J, Liu Y, Zhu B, Tian Y, Zhong R, Chen W, Lu X, Zou L, Shen N, Qian J, Li H, Miao X, Wang L. SF3A1 and pancreatic cancer: new evidence for the association of the spliceosome and cancer. Oncotarget. 2015; 6:37750–37757. doi: 10.18632/oncotarget.5647.
50. Valinezhad Orang A, Safaralizadeh R, Kazemzadeh-Bavili M. Mechanisms of miRNA-Mediated Gene Regulation from Common Downregulation to mRNA-Specific Upregulation. Int J Genomics. 2014; 2014:970607.