
INTRODUCTION
Pancreatic cancer (PaC) is a highly malignant tumor with an overall 5-year survival rate of less than 5% [1, 2]. Therapeutic methods for these patients are scarce, curative resection remains the only potentially therapy for PaC patients until recently. However, patients often present with advanced stage with distant metastases, limiting the chance of surgical resection [3]. Thus, there is a desperate need for the development of new diagnostic biomarkers as well as innovative treatment strategies to improve this situation [4].
PaC is characterized by a prominent desmoplastic reaction and the stroma provides structural support for PaC progression [5, 6]. Therefore, the fibrotic matrix was no longer merely considered as a host barrier against tumor progression, it has now become obvious that it can initiate and promote tumorigenesis [1, 7]. Meanwhile, the stroma is a dynamic cellular microenvironment that is mainly made up of pancreatic stellate cells (PSCs). The typical feature of PaC desmoplastic reaction is quiescent PSCs transform into activated PSCs. Therefore, PSCs are considered to be critical for PaC desmoplastic response. Periostin (POSTN), a 90-kilodalton secretory protein, has been involved in a number of human cancers including PaC. PSCs are the only source of POSTN in PaC. Besides, POSTN expression has been shown to be upregulated 42-fold in PaC compared with normal pancreas at the mRNA level [8]. Once stimulated by tumor cells, PSCs will perpetually be activated and produce excessive extracellular matrix to infiltrate and envelop the normal parenchyma via an autocrine POSTN loop, creating a tumor-supportive microenvironment [1, 5]. In a word, PaC is one of the most malignant cancers and result in a heavy health burden worldwide, new methods are urgently needed to treat this malignancy [9, 10].
Solid tumor growth depends on the vascular vessels to supply nutrients. In general, tumor progression is accompanied by rich angiogenesis to satisfy the metabolic requirements [11]. Tumor metastasis is a major contributor to cancer-related death and the process of capillary formation in tumor is a crucial aspect to enable metastasis, especially when tumors diameter approximates 2 mm [12]. The poor outcome of PaC is closely associate with the tendency to spread to surrounding tissues, approximately 50% of PaC patients present with distant metastatic disease [13], which is significantly correlated with the phenomenon of angiogenesis [14, 15]. Therefore, tumor angiogenesis correlated significantly with metastasis, poor prognosis as well as a poor 5-year overall survival. Many efforts have been made to inhibit the angiogenesis for PaC. However, traditional anti-vascular drug may promote tumor progression by inducing hypoxia which insteadly stimulates angiogenesis, so novel anti-vascular therapy should be applied [12]. To our knowledge, there are various molecular factors involved in vascular growth. Among these, the extracellular signal-regulated kinase (Erk) and its downstream vascular endothelial growth factor (VEGF) signaling pathway have critical roles [16–18]. In particular, VEGF is probably the most important tissue factor and a key angiogenic growth factor that responsible for angioblast differentiation and tube formation [17]. Thus, VEGF overexpression not only considered as a diagnostic marker, but also a poor prognostic factor of PaC.
Technical assessment on tumor angiogenesis over the past decade has focused on contrast-enhanced ultrasound (CEUS) imaging. It is a non-invasive approach to monitor the tumor angiogenesis in real-time with high spatial and temporal resolution. Sonovue is the second-generation ultrasound contrast agent containing stabilized microbubbles of sulfur hexafluoride gas, which exhaled via lung within a few minutes and not impaire the renal function [19]. Thus, CEUS is a feasible and safety way to assess the tumor angiogenesis quantitatively.
In this paper, we elucidate a new finding that sliencing POSTN expression disclosed anti-angiogenesis and anti-metastasis efficacies in PaC. Although POSTN expression has been identified in some other tumors, little is known about its biological functions on PaC, especially on POSTN-related angiogenic mechanism. The aim of this study is to clarify the role of POSTN in angiogenesis in PaC and to explore its potential signaling targets, such as Erk/VEGF signaling pathway. Our new findings will facilitate the understanding of POSTN-mediated angiogenesis mechanisms on PaC.
RESULTS
POSTN promotes tubule formation of HUVECs
Ten thousand Human umbilical vein endothelial cells (HUVECs) were suspended in medium with different concentrations of rPOSTN and plated into 96-well plate coated with matrigel. After 8 h incubation at 37°C, tubule formation of each group was assessed under light microscope. We observed increased tubule formation in the rPOSTN-treated HUVECs group whatever in tubular number, tubular length and tubular intersecting nods, compared with the control group. (Figure 1).
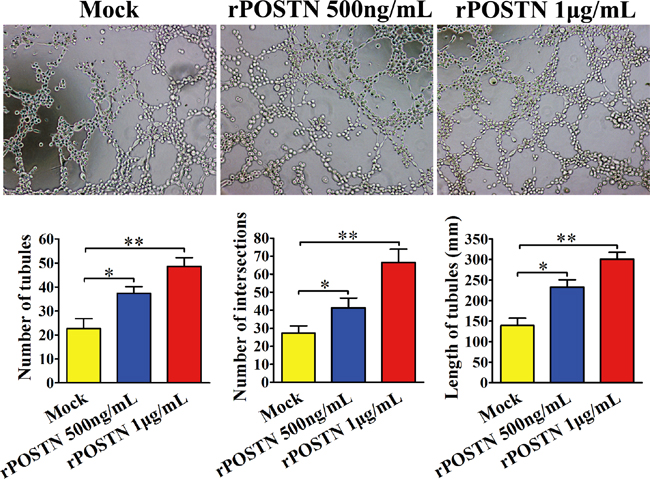
Figure 1: Effect of POSTN on tubule formation in vitro. In the HUVEC assay, abundant tubules were observed in the rPOSTN-treated group compared with the control group (×200 magnification; Zeiss). Bar charts show numbers of tubules, numbers of nodule intersections, and length of tubules between different groups. *P<0.05 and **P<0.01 vs. Mock.
POSTN promotes proliferation, migration and invasion of HUVECs
The tubule formation of HUVECs was associated with HUVECs proliferation, migration and invasion. We observed HUVECs proliferation was significantly increased in rPOSTN-treated group compared with control groups (Figure 2A). Likewise, a similar tendency was observed in migration and invasion. The migration and invasion ability of HUVECs was enhanced in rPOSTN-treated group compared with control groups (Figure 2B, 2C and 2D). Together, our results suggest that POSTN promoted tumor angiogenesis in PaC by stimulating the proliferation, migration and invasion of HUVECs.
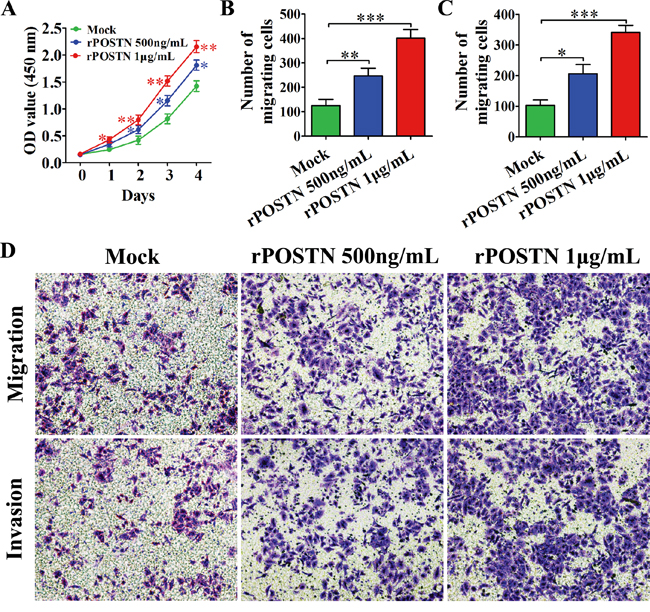
Figure 2: Effects of POSTN on proliferation, migration and invasion of HUVECs. A. The proliferation of HUVECs was increased in rPOSTN-treated groups compared with the control group. B. Histograms showed the numbers of migration cells. C. Histograms showed the numbers of invasion cells. D. POSTN promoted the migration and invasion of HUVECs (×200 magnification; Zeiss). Data are represented as mean ± SD of three independent experiments.
POSTN promotes tumorigenesis in vivo
The effect of POSTN on tumorigenesis was examined by subcutaneous co-injection of SW1990 and POSTN-sh1 transfected PSCs. We observed that knockdown of POSTN in PSCs inhibited tumor growth compared with the control group (Figure 3A). The mean tumor volume was much smaller in POSTN-silenced group than that in control group (Figure 3C). Besides, the tumor weight was decreased in POSTN-silenced group compared with the control group (2.58±0.21 g vs 1.31±0.19 g, P<0.05) (Figure 3D). Conventional ultrasound and contrast-enhanced ultrasound (CEUS) were performed to evaluate the vessels formation before the mice were sacrificed (Figure 3B). The results showed reduced blood flow signals were observed in POSTN-silenced group compared with the control group. Thus, enough nutrition from increasing vessels regulated by POSTN may be the interpretation about the above findings.
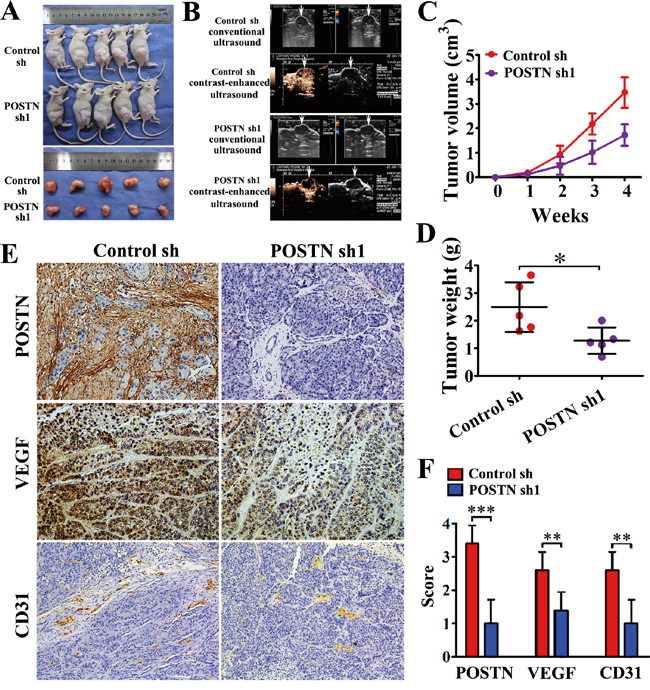
Figure 3: POSTN enhances the tumorigenicity and angiogenesis of pancreatic cancer cells in vivo. A. SW1990 cells were co-injected with control shRNA-transfected PSCs (Control sh) or POSTN shRNA1-transfected PSCs (POSTN sh1) into the right side of nude mice. After 4 weeks the mice were sacrificed. POSTN knockdown group exhibited slower growth than control group. B. Conventional ultrasound and contrast-enhanced ultrasound (CEUS) were performed before the mice were sacrificed to evaluate angiogenesis in the xenografts. Fewer blood vessels were observed in the POSTN knockdown group compared with the control group. C and D. Silencing of POSTN exhibited reduced tumor volume and weight of xenografts. E. Immunohistochemical staining showed POSTN deposits in the stroma of xenografts. Xenograft tumors from the POSTN-shRNA group were decreased in VEGF expression and contained significantly fewer CD31-positive small capillary-like vessels than those from the control group (n=15, five random fields). F. The statistical analysis on immunohistochemical staining by scores.
Immunohistochemical staining showed VEGF expression decreased in POSTN-silenced group and significantly fewer CD31 positive capillary-like vessels in POSTN-sh1 xenograft tumors compared with the control group (Figure 3E and 3F). This tendency was congruity with CEUS and consistent with the results in vitro. These data collectively indicate that POSTN promoted PaC growth via stimulating vessels formation in vivo.
POSTN upregulates phosphorylated Erk in PaC
Erk signaling is not only related with tumor angiogenesis, but also tumor metastasis. In this study, POSTN increased Thr202 and Tyr204 phosphorylation of Erk and VEGF expression. Opposing results were observed in POSTN-silenced group compared with the control groups (Figure 4A). Meanwhile, we found Erk phosphorylation and VEGF expression were decreased in POSTN-sh1 xenograft tumor (Figure 4B). As shown in Figure 4C, the effect of POSTN on Erk phosphorylation was completely abrogated when cells were treated with Erk inhibitor SCH772984. Moreover, the phosphorylation of downstream molecule VEGF was correspondingly inhibited. Next, we examined the effects of SCH772984 on cell function and it is worth mentioning that the proliferation, migration and tubule formation of HUVECs were decreased after treated with Erk inhibitor SCH772984 (Figure 5A, 5B, 5C and 5D). Together, these findings suggested that POSTN promoted tumor angiogenesis mainly via Erk/VEGF signaling.
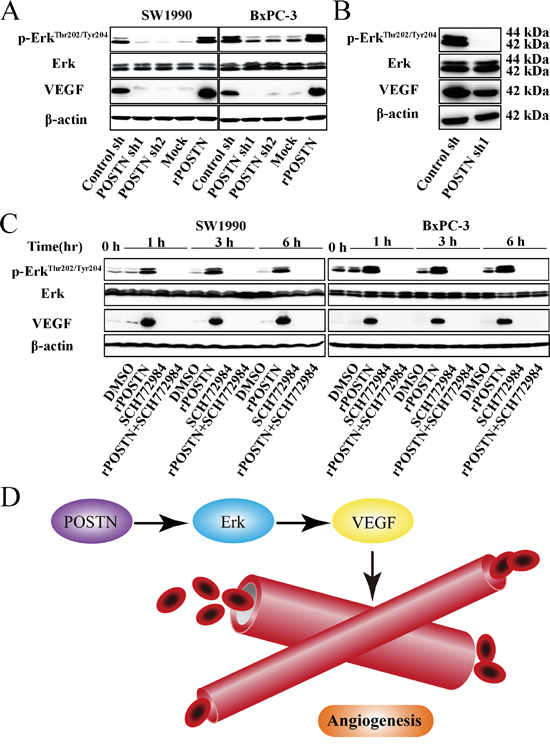
Figure 4: POSTN activates Erk/VEGF signaling to regulate the angiogenesis of pancreatic cancer cells. A. SW1990 and BxPC-3 cells were treated with the supernatant of control shRNA-transfected PSCs (Control sh), POSTN shRNA1-transfected PSCs (POSTN sh1), POSTN shRNA2-transfected PSCs (POSTN sh2), or human recombinant protein (rPOSTN, 1 μg/mL). After 12 h, cells were harvested and the basal expression of Erk and its downstream molecule VEGF was determined by western blotting. B. Xenograft tumors of nude mice from the control-shRNA group and POSTN-shRNA1 group were also subjected to western blotting using the indicated antibodies. C. SW1990 and BxPC-3 cells were treated with DMSO, rPOSTN, Erk inhibitor (SCH772984, 20 μM), or rPOSTN plus SCH772984. Cells were harvested after 0, 1, 3, and 6 h, and the basal expression of Erk and its downstream molecule was determined by western blotting. D. POSTN activates Erk-VEGF signaling to regulate the angiogenesis of pancreatic cancer cells.
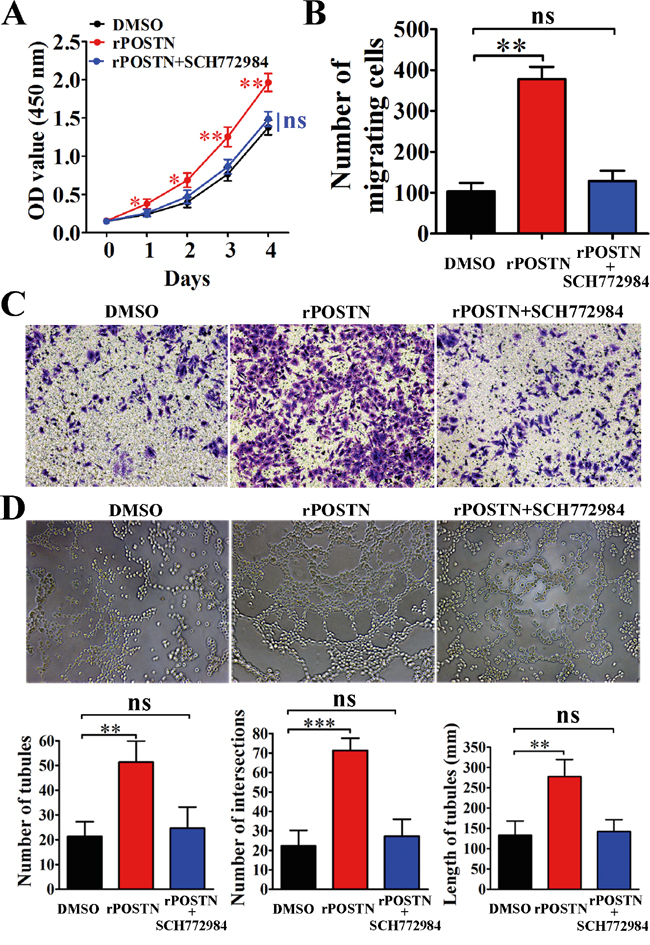
Figure 5: The effects of Erk inhibitor on pancreatic cancer cells. A. Proliferation of HUVECs was increased by treated with rPOSTN (1 μg/mL) and was completely inhibited by SCH772984. B. Histograms showed the numbers of migration cells. C. Migration of HUVECs was significantly reduced by treatment with SCH772984. D. Tubule formation ability of HUVECs was completely inhibited by SCH772984. Bar charts show numbers of tubules, numbers of nodule intersections, and length of tubules between different groups. *P<0.05, **P<0.01 and ***P<0.001vs. DMSO.
VEGF in PaC tissues was positively associated with POSTN expression
To further investigate the correlation between POSTN expression and VEGF expression, we used TMAs to study POSTN and VEGF expression levels in PaC and corresponding paired normal tissues. We performed immunohistochemical staining for POSTN and VEGF on a large cohort of primary PaC patients simultaneously (n=30). Semiquantitative analysis showed an increased intensity of POSTN and VEGF staining in PaC compared with normal tissues. Both VEGF and POSTN had similar expression pattern in PaC tissues. Besides, POSTN expression was positively associated with VEGF expression in the same patient tissue (Figure 6, Table 1). Positive correlation was found between the expression of these two proteins (r=0.472, P<0.01).
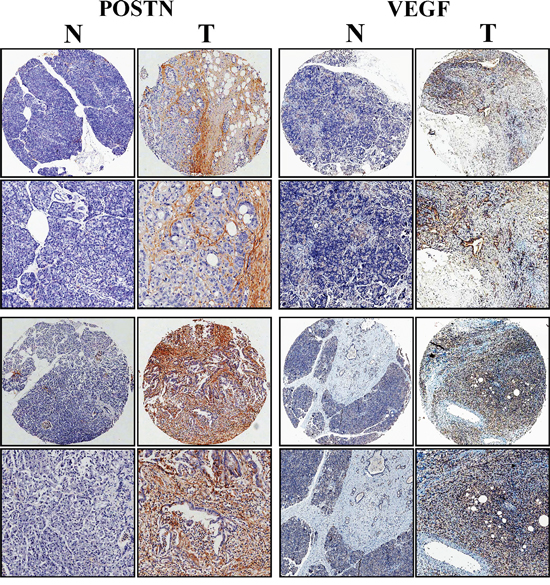
Figure 6: VEGF in pancreatic cancer tissues was significantly correlated with POSTN expression. Immunohistochemical staining of the same pancreatic cancer and matched normal tissue with anti-POSTN and anti-VEGF antibody simultaneously. Representative photomicrographs of POSTN and VEGF in pancreatic specimens showed VEGF immunostaining was significantly associated with POSTN expression (magnification, ×50, ×200).
Table 1: Expression of POSTN and VEGF in pancreatic cancer tissues
VEGF |
P |
|||
|---|---|---|---|---|
- |
+ |
|||
POSTN |
- |
8 |
2 |
<0.05 |
+ |
6 |
14 |
||
DISCUSSION
The microenvironment of cancer is complicated and can maintain stable survival advantages of PaC cells despite various perturbations [20, 21]. When the environment is not optimal, PaC cells can mediate adaptive changes such as stimulation of tumor angiogenesis. Angiogenesis plays a critical role in tumor initiation, progression and metastasis [22], and now it has been considered to be one of the major hallmarks of cancer [23–25]. Tumors require blood supply for nutrition, growth as well as distant metastasis. More and more attention has been focused on tumor angiogenesis, which is significantly correlated with cancer metastasis. The importance of angiogenesis in PaC progression cannot be ignored [26–28]. Surgical resection is a reasonable therapy to PaC patients, patients who have no distant metastasis should undergo surgical resection [29]. However, over 80 ~ 90% patients are diagnosed at the advanced stage, which attributed to the early metastasis and dissemination of the PaC cells [3]. POSTN, originally designated osteoblast-specific factor 2, was found to be overexpressed in various types of human tumors [30, 31], including PaC [1, 32]. POSTN mRNA elevated 42-fold in PaC, and patients with increased expression had a shorter survival tendence [1]. Thus, cancer metastasis is significantly correlated with tumor angiogenesis. However, the detailed tumor-associated angiogenesis function of POSTN in PaC remain to be explored. Based on these observations, we aim to determine the role of POSTN in mediating tumor angiogenesis of PaC and its potential clinical significance. The endothelial tube formation assay is based on the ability of endothelial cells to form 3D tubular structures when coated with a gel of basement membrane extract.
There is plenty of evidence to support the significance of angiogenesis in tumor development. For instance, Zhang et al. found that upregulate of POSTN expression may promote angiogenesis in keloids and concluded that POSTN may be a key factor in keloid development [33]. Wang et al. reported that high expression of POSTN is closely associated with tumor angiogenesis, progression and poor survival of esophageal squamous cell carcinoma (ESCC) [25]. Hu et al. examined POSTN expression in osteosarcoma and explored the relationship of POSTN expression with tumor angiogenesis. They showed that high expression of POSTN significantly correlated with VEGF expression and microvessal density compared to the normal bone tissues [34]. Our experiments also confirmed that rPOSTN significantly promoted angiogenesis in PaC. We verified that POSTN promoted tubule formation dependents on HUVECs and POSTN was also able to accelerate HUVECs proliferation, migration and invasion, which contributed to tubule formation in PaC. Beside, the nude mice tumor immunohistochemical staining showed fewer capillary-like vessels in POSTN-silenced group. Diagnostic imaging has significant clinical value and contrast-enhanced ultrasound (CEUS) has been increasingly used to evaluate tumor angiogenesis [19, 35]. The contrast agent SonoVue is a blood pool contrast agent [36] that can distribute entirely within the blood vessels and dynamically reveal the blood supply in the tumor mass and the microvessel structure in real time [37, 38]. We performed contrast-enhanced ultrasound on the subcutaneous tumors of mice before they were sacrificed. Consistent with the in vitro experiments, tumors in the control group showed high enhancement reflecting a rich blood flow signal, whereas reduced blood flow signals were observed in the POSTN knockdown group. Not surprisingly, the volume and weight of subcutaneous xenografts were decreased in nude mice derived from the POSTN knockdown group as insufficient nutrients were supplied for tumor growth. Along with these important clues, we further researched the potential association between POSTN expression and angiogenesis to better understand the mechanisms for how these processes could be regulated by POSTN.
As to blood supply of tumor, we turned our attention to the “classic” gene, such as VEGF. It is the most potent angiogenic factor for its high specificity to endothelial cells [33, 39, 40]. Our study demonstrated that increased POSTN expression significantly promoted Erk Phosphorylation and VEGF expression. Erk, affecting cellular angiogenesis, was detected in this study. Interestingly, we observed that Erk inhibitor SCH772984 inhibited Erk phosphorylation and significantly decreased VEGF expression as well as tubule formation of HUVECs in rPOSTN-treated PaC cells. In accordance with these observations, we found that knockdown of POSTN in PaC cells and tissues decreased Erk phosphorylation and its downstream VEGF expression. Additionally, the proliferation and migration of HUVECs were also decreased in SCH772984-treated group. Besides, in our study, we observed VEGF expression was decreased in POSTN-silenced group by nude mice tumor immunochemistry. Taken together, these findings suggested that POSTN promoted PaC angiogenesis, at least in part, via Erk signaling.
In conclusion, our findings elucidated the critical role of POSTN in PaC angiogenesis, POSTN could promote PaC metastasis and tumor angiogenesis via Erk/VEGFsignaling pathways, which may be serve as a marker in highly aggressive phenotype of PaC. In addition, inhibition of POSTN could suppress PaC progression in vitro and in vivo, suggesting that POSTN inhibition might represent new and potential strategies against human PaC. Collectively, we may identify a promising targeted therapy for PaC patients based on POSTN.
MATERIALS AND METHODS
Ethical statement
Informed consent was obtained from all participants and this research was approved by the ethics committee of Shanghai General Hospital affiliated of Shanghai Jiaotong University and performed in accordance with ethical principles. All mouse experiments were manipulated and housed according to the protocols approved by Shanghai Medical Experimental Animal Care Commission.
Cell lines and reagent
The human PaC cell lines BxPC3 and SW1990 were purchased from American Type Culture Collection (Manassas, VA), Human umbilical vein endothelial cells (HUVECs) were purchased from Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences. These cells were maintained in RPMI 1640 with 10% FBS. Human pancreatic stellate cells (PSCs) were purchased from ScienCell research laboratory (Carlsbad, CA) and maintained in stellate cell medium (ScienCell). All cells were cultured in a humidified atmosphere of 5% CO2 at 37°C. Human recombinant POSTN protein (rPOSTN) was purchased from Biovendor (Heidelberg, Germany) and dissolved in 0.1 M acetate buffer (pH 4) at a concentration of 1 μg/mL.
Tissue microarray construction
PaC samples and paired adjacent non-tumor tissues with informed consent were collected from 30 patients who underwent pancreatic surgery and were stored at Biobank Center of National Engineering Center for Biochip at Shanghai. Tissue microarray was stained for expression analysis of POSTN (ab14041, Abcam, 1:50 dilution), VEGF (sc-152, Santa Cruz Biotechnology, 1:50 dilution). All immunohistochemically stained sections were dependently scored by two in-house pathologists who were blinded to clinical outcome.
Lentivirus transduction for gene silencing
The lentivirus suspension used for shRNA silencing of the POSTN gene in PSCs was purchased from Ebioeasy Ltd (Shanghai, China). The target sequences for POSTN were 5’-CGGTGACAGTATAACAGTAAA-3’ named POSTN sh1, 5’-CACTTGTAAGAACTGGTATAA-3’ named POSTN sh2, respectively. The sequence for scrambled negative control shRNA was 5’-CCTAAGGTTAAGTCGCCCTCG-3’ named Control sh (Supplementary Figures 1). The PSCs lentivirus infection was performed according to the manufacturer’s instructions.
Endothelial tube formation assay
Briefly, each well of prechilled 96-well plates was covered with a thin layer of Matrigel, which was allowed to polymerize at 37°C for 1 h. HUVECs were resuspended in medium with different concentrations of rPOSTN. HUVECs (50 μl, 1 × 104 cells/well) were added to the polymerized Matrigel. After incubation for 8 h at 37°C with 5% CO2, the tube formation ability was evaluated by measuring the tubular length and counting the number of tubes and tubular intersecting nodes in five random fields using Image Pro Plus software. Each experiment was performed at least three times.
Cell proliferation assay
Cell proliferation was measured by Cell Counting Kit-8 (Dojindo, Kumamoto, Japan) according to the manufacturer’s instructions. HUVECs (2×103 cells/well) were cultured with different concentrations of rPOSTN in 96-well plates. The rPOSTN concentration was selected based on data shown in Supplementary Figures 2. Cell proliferation was examined every 24 h for 5 days after incubation for 2 h at the absorbance 450 nm.
Cell migration and invasion assays
For cell migration and invasion assays, 6.5-mm Transwells® chamber with 8.0-μm Pore Polycarbonate Membrane Insert (Corning, NY) was used according to the manufacturer’s instructions. In invasion assay, diluted Matrigel (BD Biosciences) was plated in transwell chambers. HUVECs (4 × 104) in 200 μL serum-free medium were added to the upper chamber and cultured for 48 h. In migration assay, the membranes were not coated with Matrigel and HUVECs (2 × 104) were cultured in the same condition. For both assays, 500 μl of medium with different concentrations of rPOSTN was added to the lower chamber as a chemoattractant. Finally, the migrated cells and invasive cells on the bottom of the membrane were fixed with 20% methanol and stained with 0.04% crystal violet and cells were photographed and counted under a microscope.
Western blotting
Whole cell protein lysates were electrophoresed on 10% sodium dodecyl sulfate-polyacrylamide gels and transferred onto polyvinylidene difluoride membranes (Millipore). The membranes were blocked with 5% non-fat milk in Tris-buffered saline and then incubated with primary antibodies at 4°C overnight. The primary antibodies were as follows: anti-POSTN (1:1,000; ab14041, Abcam, Cambridge, UK), anti-Erk (1:1,000; #4695; Cell Signaling Technology), anti-P-ErkThr202/Tyr204 (1:1,000; #9101; Cell Signaling Technology), anti-VEGF (1:1,000; sc-152; Santa Cruz Biotechnology) and anti-β-actin (1:5,000; Abcam). Membranes were then incubated with peroxidase-linked secondary antibody for 2 h at room temperature. Target protein signals were visualized by an enhanced chemiluminescence detection system (Amersham Bioscience, Piscataway, NJ, USA) according to the manufacturer’s protocol. All western blotting analyses were performed three times.
Nude mice tumorigenesis
SW1990 (3×106 cells/mouse) and POSTN sh1 or control sh stably transfected PSCs (3×106 cells/mouse) were subcutaneously co-injected into 4-week-old male nude mice (Institute of Zoology, Chinese Academy of Sciences, Shanghai, China). Tumor nodules were examined weekly, and were evaluated using the following formula: tumor volume = (Width2×Length)/2 [41]. Mice were killed 4 weeks after injection. Before sacrifice, contrast-enhanced ultrasound (CEUS) was performed using SonoVue as a contrast agent to assess angiogenesis in vivo. Lyophilized SonoVue powder was dissolved in 5 mL saline and 100 μL of suspension was injected via the post-glomus venous plexus within 2–3 s, followed by a 100 μL saline flush. Real-time observation of the xenograft was performed for at least 120 s. Dynamic images were preserved for later analysis. Finally, tumors were excised, weighed and fixed for immunohistochemistry staining.
Immunohistochemistry
The paraffin-embedded nude mice tumor tissue was sliced into 4-μm thick sections and then deparaffinized and rehydrated. Immunohistochemical staining of sections was performed using primary antibody mouse anti-POSTN (1:100; Abcam, Cambridge, UK), primary antibody rabbit anti-VEGF (1:100; Santa Cruz Biotechnology), mouse anti-CD31 (1:100; Santa Cruz Biotechnology) at 37 °C for 2 h. CD31 positive vessels indicated blood vessels in tissues. Finally, the analysis was assessed and scored by two independent investigators simultaneously by combining both the scores and staining intensity. Scores representing the percentage of positive staining tumor cells were categorized into: 0 (0–5%), 1 (6–25%), 2 (26–50%), 3 (51%–75%) and 4 (>75%) [42]. The intensity of staining was categorized into 0 (negative), 1 (weak, light yellow), 2 (moderate, yellow brown), or 3 (strong, brown) [43]. The final score was determined by the product of staining intensity and percentage of positive tumor cells (final score = intensity score × percentage score). The final score of ≤ 3 was considered as low expression, and > 3 as high expression [42].
Statistical analysis
All statistical analyses were carried out using the SPSS 16.0 statistical software package. Statistical comparisons were conducted using Student’s t-test and presented as mean ± SD. Correlation between POSTN and VEGF expression was analyzed by Spearman test. In all cases, P-value <0.05 was considered as statistical significant.
ACKNOWLEDGMENTS
We gratefully acknowledge the entire staff of the Department of Pathophysiology in the Key Laboratory of Cell Differentiation and Apoptosis of the National Ministry of Education.
CONFLICTS OF INTEREST
We do not have any potential conflicts of interest for all the authors.
GRANT SUPPORT
This study was supported by grants from the National Natural Science Foundation of China (Nos. 81271596, Nos. 81571679 and Nos. 81471666), and the Medical and Engineering Cross-Foundation of Shanghai Jiaotong University (Nos. YG2015ZD09).
REFERENCES
1. Erkan M, Kleeff J, Gorbachevski A, Reiser C, Mitkus T, Esposito I, Giese T, Buchler MW, Giese NA, Friess H. Periostin creates a tumor-supportive microenvironment in the pancreas by sustaining fibrogenic stellate cell activity. Gastroenterology. 2007; 132:1447-1464.
2. Razidlo GL, Magnine C, Sletten AC, Hurley RM, Almada LL, Fernandez-Zapico ME, Ji B, McNiven MA. Targeting Pancreatic Cancer Metastasis by Inhibition of Vav1, a Driver of Tumor Cell Invasion. Cancer research. 2015; 75:2907-2915.
3. Xia X, Zhang K, Cen G, Jiang T, Cao J, Huang K, Huang C, Zhao Q, Qiu Z. MicroRNA-301a-3p promotes pancreatic cancer progression via negative regulation of SMAD4. Oncotarget. 2015; 6:21046-21063. doi: 10.18632/oncotarget.4124.
4. Wolfgang CL, Herman JM, Laheru DA, Klein AP, Erdek MA, Fishman EK, Hruban RH. Recent progress in pancreatic cancer. CA Cancer J Clin. 2013; 63:318-348.
5. Liu Y, Du L. Role of pancreatic stellate cells and periostin in pancreatic cancer progression. Tumour biology. 2015; 36:3171-3177.
6. Lunardi S, Jamieson NB, Lim SY, Griffiths KL, Carvalho-Gaspar M, Al-Assar O, Yameen S, Carter RC, McKay CJ, Spoletini G, D’Ugo S, Silva MA, Sansom OJ, et al. IP-10/CXCL10 induction in human pancreatic cancer stroma influences lymphocytes recruitment and correlates with poor survival. Oncotarget. 2014; 5:11064-11080. doi: 10.18632/oncotarget.2519.
7. Wang LM, Silva MA, D’Costa Z, Bockelmann R, Soonawalla Z, Liu S, O’Neill E, Mukherjee S, McKenna WG, Muschel R, Fokas E. The prognostic role of desmoplastic stroma in pancreatic ductal adenocarcinoma. Oncotarget. 2015. doi: 10.18632/oncotarget.6770.
8. Erkan M, Hausmann S, Michalski CW, Fingerle AA, Dobritz M, Kleeff J, Friess H. The role of stroma in pancreatic cancer: diagnostic and therapeutic implications. Nature reviews Gastroenterology & hepatology. 2012; 9:454-467.
9. D’Angelo FA, Antolino L, La Rocca M, Petrucciani N, Magistri P, Aurello P, Ramacciato G. Adjuvant and neoadjuvant therapies in resectable pancreatic cancer: a systematic review of randomized controlled trials. Med Oncol. 2016; 33:28.
10. Heestand GM, Kurzrock R. Molecular landscape of pancreatic cancer: implications for current clinical trials. Oncotarget. 2015; 6:4553-4561. doi: 10.18632/oncotarget.2972.
11. Jiang J, Liu W, Guo X, Zhang R, Zhi Q, Ji J, Zhang J, Chen X, Li J, Gu Q, Liu B, Zhu Z, Yu Y. IRX1 influences peritoneal spreading and metastasis via inhibiting BDKRB2-dependent neovascularization on gastric cancer. Oncogene. 2011; 30:4498-4508.
12. Zang M, Zhang Y, Zhang B, Hu L, Li J, Fan Z, Wang H, Su L, Zhu Z, Li C, Yan C, Gu Q, Liu B, et al. CEACAM6 promotes tumor angiogenesis and vasculogenic mimicry in gastric cancer via FAK signaling. Biochimica et biophysica acta. 2015; 1852:1020-1028.
13. Yang YF, Cao XH, Bao CE, Wan X. Concurrent radiotherapy with oral fluoropyrimidine versus gemcitabine in locally advanced pancreatic cancer: a systematic review and meta-analysis. OncoTargets and therapy. 2015; 8:3315-3322.
14. Hong SM, Park JY, Hruban RH, Goggins M. Molecular signatures of pancreatic cancer. Archives of pathology & laboratory medicine. 2011; 135:716-727.
15. Wang SH, Sun YF, Liu Y, Zhou Y. CT contrast enhancement correlates with pathological grade and microvessel density of pancreatic cancer tissues. International journal of clinical and experimental pathology. 2015; 8:5443-5449.
16. Carmeliet P, Jain RK. Angiogenesis in cancer and other diseases. Nature. 2000; 407:249-257.
17. Costache MI, Ioana M, Iordache S, Ene D, Costache CA, Saftoiu A. VEGF Expression in Pancreatic Cancer and Other Malignancies: A Review of the Literature. Romanian journal of internal medicine. 2015; 53:199-208.
18. Hayes TK, Neel NF, Hu C, Gautam P, Chenard M, Long B, Aziz M, Kassner M, Bryant KL, Pierobon M, Marayati R, Kher S, George SD, et al. Long-Term ERK Inhibition in KRAS-Mutant Pancreatic Cancer Is Associated with MYC Degradation and Senescence-like Growth Suppression. Cancer cell. 2016; 29:75-89.
19. Paprottka PM, Rosspunt S, Ingrisch M, Cyran CC, Nikolaou K, Reiser MF, Mack B, Gires O, Clevert DA, Zengel P. Reducing tumor growth and angiogenesis using a triple therapy measured with Contrast-enhanced ultrasound (CEUS). BMC cancer. 2015; 15:373.
20. Wilson JS, Pirola RC, Apte MV. Stars and stripes in pancreatic cancer: role of stellate cells and stroma in cancer progression. Frontiers in physiology. 2014; 5:52.
21. Coffman LG, Choi YJ, McLean K, Allen BL, di Magliano MP, Buckanovich RJ. Human carcinoma-associated mesenchymal stem cells promote ovarian cancer chemotherapy resistance via a BMP4/HH signaling loop. Oncotarget. 2016; 7:6916-32. doi: 10.18632/oncotarget.6870.
22. Blagosklonny MV. Antiangiogenic therapy and tumor progression. Cancer cell. 2004; 5:13-17.
23. Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011; 144:646-674.
24. Yang D, Wang LL, Dong TT, Shen YH, Guo XS, Liu CY, Liu J, Zhang P, Li J, Sun YP. Progranulin promotes colorectal cancer proliferation and angiogenesis through TNFR2/Akt and ERK signaling pathways. American journal of cancer research. 2015; 5:3085-3097.
25. Wang W, Sun QK, He YF, Ma DC, Xie MR, Ji CS, Hu B. Overexpression of periostin is significantly correlated to the tumor angiogenesis and poor prognosis in patients with esophageal squamous cell carcinoma. International journal of clinical and experimental pathology. 2014; 7:593-601.
26. Craven KE, Gore J, Korc M. Overview of pre-clinical and clinical studies targeting angiogenesis in pancreatic ductal adenocarcinoma. Cancer letters. 2015.
27. Khan MA, Srivastava SK, Bhardwaj A, Singh S, Arora S, Zubair H, Carter JE, Singh AP. Gemcitabine triggers angiogenesis-promoting molecular signals in pancreatic cancer cells: Therapeutic implications. Oncotarget. 2015; 6:39140-39150. doi: 10.18632/oncotarget.3784.
28. Gore J, Craven KE, Wilson JL, Cote GA, Cheng M, Nguyen HV, Cramer HM, Sherman S, Korc M. TCGA data and patient-derived orthotopic xenografts highlight pancreatic cancer-associated angiogenesis. Oncotarget. 2015; 6:7504-7521. doi: 10.18632/oncotarget.3233.
29. Wagner M, Redaelli C, Lietz M, Seiler CA, Friess H, Buchler MW. Curative resection is the single most important factor determining outcome in patients with pancreatic adenocarcinoma. The British journal of surgery. 2004; 91:586-594.
30. Sung PL, Jan YH, Lin SC, Huang CC, Lin H, Wen KC, Chao KC, Lai CR, Wang PH, Chuang CM, Wu HH, Twu NF, Yen MS, et al. Periostin in tumor microenvironment is associated with poor prognosis and platinum resistance in epithelial ovarian carcinoma. Oncotarget. 2016; 7:4036-47. doi: 10.18632/oncotarget.6700.
31. Errico A. CNS cancer: periostin-a new potential target for the treatment of glioblastoma. Nature reviews Clinical oncology. 2015; 12:128.
32. Kanno A, Satoh K, Masamune A, Hirota M, Kimura K, Umino J, Hamada S, Satoh A, Egawa S, Motoi F, Unno M, Shimosegawa T. Periostin, secreted from stromal cells, has biphasic effect on cell migration and correlates with the epithelial to mesenchymal transition of human pancreatic cancer cells. International journal of cancer. 2008; 122:2707-2718.
33. Zhang Z, Nie F, Chen X, Qin Z, Kang C, Chen B, Ma J, Pan B, Ma Y. Upregulated periostin promotes angiogenesis in keloids through activation of the ERK 1/2 and focal adhesion kinase pathways, as well as the upregulated expression of VEGF and angiopoietin1. Molecular medicine reports. 2015; 11:857-864.
34. Hu F, Shang XF, Wang W, Jiang W, Fang C, Tan D, Zhou HC. High-level expression of periostin is significantly correlated with tumour angiogenesis and poor prognosis in osteosarcoma. International journal of experimental pathology. 2016.
35. Klotz LV, Clevert DA, Scheckinger S, Strieth S, Eichhorn ME. Contrast-enhanced Ultrasound Imaging of Antiangiogenic Tumor Therapy. Anticancer research. 2015; 35:2571-2576.
36. Pysz MA, Guracar I, Foygel K, Tian L, Willmann JK. Quantitative assessment of tumor angiogenesis using real-time motion-compensated contrast-enhanced ultrasound imaging. Angiogenesis. 2012; 15:433-442.
37. Fan Z, Yan K, Wang Y, Qiu J, Wu W, Yang L, Chen M. Application of Contrast-Enhanced Ultrasound in Cystic Pancreatic Lesions Using a Simplified Classification Diagnostic Criterion. BioMed research international. 2015; 2015:974621.
38. Li ZJ, Du LF, Luo XH. Evaluation of ventricular-vascular coupling in patients with type 2 diabetes mellitus using 2-dimensional speckle tracking imaging. Journal of Huazhong University of Science and Technology Medical sciences. 2014; 34:929-934.
39. Eichmann A, Simons M. VEGF signaling inside vascular endothelial cells and beyond. Current opinion in cell biology. 2012; 24:188-193.
40. Blagosklonny MV. How Avastin potentiates chemotherapeutic drugs: action and reaction in antiangiogenic therapy. Cancer biology & therapy. 2005; 4:1307-1310.
41. Zhou C, Ji J, Cai Q, Shi M, Chen X, Yu Y, Liu B, Zhu Z, Zhang J. MTA2 promotes gastric cancer cells invasion and is transcriptionally regulated by Sp1. Molecular cancer. 2013; 12:102.
42. Luo Q, Wang C, Jin G, Gu D, Wang N, Song J, Jin H, Hu F, Zhang Y, Ge T, Huo X, Chu W, Shu H, et al. LIFR functions as a metastasis suppressor in hepatocellular carcinoma by negatively regulating phosphoinositide 3-kinase/AKT pathway. Carcinogenesis. 2015; 36:1201-1212.
43. Xu Y, Dong B, Huang J, Kong W, Xue W, Zhu Y, Zhang J, Huang Y. Sphingosine kinase 1 is overexpressed and promotes adrenocortical carcinoma progression. Oncotarget. 2016; 7:3233-3244. doi: 10.18632/oncotarget.6564.