
INTRODUCTION
About 30% of endometrial cancers, predominantly of endometrioid histology, can be molecularly characterized by microsatellite instability (MSI). MSI is a hypermutable phenotype caused by the loss of DNA mismatch repair (MMR) activity mostly due to sporadic MLH1 promoter hypermethylation [1]. Tumors that exhibit this phenotype have numerous insertions and deletions also in coding microsatellites causing frameshift mutations and loss of protein function. The coding microsatellite-containing genes frequently affected by MSI are believed to be involved in progression of MSI tumors [2]. Some target genes, such as BAX, are altered in diverse MSI tumor types (e.g. colorectal- and ovarian cancer), whereas others, such as JAK1, have a very restricted occurrence in MSI endometrial cancers [3-6]. MSI endometrial cancers show a remarkably high number of JAK1 frameshift mutations that may have clinical implications [4, 5].
JAK1 plays a role in the JAK/STAT pathway, which is activated by cytokines such as IFNγ, that influence several cellular processes such as cell growth and immune response [7-9]. Ren et al. have shown that JAK1 mutant gynecological cancer cell lines were defective in interferon gamma (IFNγ) induced STAT1 tyrosine phosphorylation and thereby impede upregulation of antigen processing machinery components such as LMP2 and TAP1 [10]. Impaired antigen processing and presentation due to hindered expression of LMP and/or TAP proteins are associated with lack of HLA class I upregulation and resistance to cytotoxic T-cell mediated lysis [11, 12]. HLA class I expression has been reported as a prognostic marker in endometrial cancer patients [13-15] and upregulation of HLA class I was frequently impaired in MSI endometrial cancers [14, 16]. The high rate of JAK1 mutations in MSI endometrial cancer is suggestive of an adaptation favoring tumor survival by blocking the JAK/STAT pathway activity, and impeding an adequate immune response.
MSI tumors exhibit a high number of somatic mutations that could facilitate an immune response by presentation of neo-antigen-epitopes in the context of HLA class I molecules. Programmed death 1 expressed on cytotoxic T-cells is a checkpoint involved in immune suppression. Checkpoint inhibitors, as potential mechanism for T-cell activation, recently showed promising results in treatment of mismatch repair deficient tumors independent of tumor origin [17]. However, JAK1 mutations and other mechanisms involved in impeding antigen presentation and expression of antigen processing machinery components in MSI endometrial cancers may interfere with new treatment regiments for MSI tumors such as the programmed death 1 inhibitor pembroluzimab. [17, 18].
In this study, MSI and JAK1 mutation status were analyzed in a study cohort of 181 tissue samples of endometrial cancer patients with the aim to evaluate that the JAK1 locus is frequently affected by MSI, and to determine its functional implication in immune evasion by analyzing expression of antigen presenting machinery components and the presence of cytotoxic T-cells specifically in MSI endometrial cancers. Finally, the effect of JAK1 mutation status on survival was evaluated in a large independent cohort of 198 MSI endometrial cancer patients with mature long-term follow-up from the PORTEC-1 and -2 clinical trials [19, 20].
RESULTS
Of the 181 endometrial cancers from the study cohort, MSI was detected in 62 (34%) cases, in nine cases MSI status remained unclear due to technical failure. Twenty-two (35%) MSI endometrial cancers had a JAK1 frameshift mutation, mainly at position K860, whereas only 3 of 110 (3%) MSS endometrial cancers had a JAK1 mutation (P<0.001, Supplementary Table S1). Two of these three JAK1 mutant MSS cases showed focal loss of MLH1 protein expression in part of the tumor as a result of MLH1 promoter hypermethylation. There were no significant differences in age, FIGO stage, differentiation grade or tumor type between JAK1 wildtype and mutant MSI endometrial cancers (Table 1). However, JAK1 mutations were associated with deeper myometrial invasion (P=0.030; odds ratio 3.500, 95% confidence interval 1.102-11.116).
Table 1: Clinicopathological characteristics of 58 MSI endometrial cancers of the study cohort and 187 MSI endometrial cancers of the PORTEC cohort according to JAK1 mutation status
Study cohort |
PORTEC cohort |
|||||
|---|---|---|---|---|---|---|
JAK1 wildtype |
JAK1 mutation |
P-value |
JAK1 wildtype |
JAK1 mutation |
P-value |
|
Age |
1.000 |
0.914 |
||||
<60 years |
18 (50.0) |
11 (50.0) |
19 (14.1) |
7 (13.5) |
||
>60 years |
18 (50.0) |
11 (50.0) |
116 (85.9) |
45 (86.5) |
||
Tumor type |
0.430 |
0.534 |
||||
Endometrioid |
35 (97.2) |
22 (100) |
134 (99.3) |
52 (100) |
||
Serous |
1 (2.8) |
0 (0.0) |
1 (0.7) |
0 |
||
FIGO (2009)* |
0.650 |
0.270 |
||||
I |
22 (61.1) |
11 (52.4) |
135 (100) |
52 (100) |
||
II |
3 (8.3) |
4 (19.0) |
0 |
0 |
||
III |
10 (27.8) |
5 (23.8) |
0 |
0 |
||
IV |
1 (2.8) |
1 (4.8) |
0 |
0 |
||
Grade |
0.885 |
0.725 |
||||
1 |
13 (36.1) |
7 (31.8) |
77 (57.0) |
33 (63.5) |
||
2 |
14 (38.9) |
10 (45.5) |
31 (23.0) |
10 (19.2) |
||
3 |
9 (25.0) |
5 (22.7) |
27 (20.0) |
9 (17.3) |
||
Myometrial invasion* |
0.030 |
0.172 |
||||
<50% |
21 (55.3) |
6 (28.6) |
45 (33.3) |
12 (23.1) |
||
>50% |
15 (41.7) |
15 (71.4) |
90 (66.7) |
40 (76.9) |
||
* 1 missing value in the study cohort
The functional implication of JAK1 in tumor immune evasion was analyzed by expression analysis of TAP1, LMP7, HLA class I and presence of CD8-positive T-cells in the MSI endometrial cancers (Table 2). Distribution of TAP1 expression was similar for both JAK1 wildtype and mutant (P=0.151). Upregulation of LMP7 was impaired in JAK1 mutant tumors, although not statistically significant (P=0.074). Upregulation of HLA class I was significantly impaired in JAK1 mutant tumors (P<0.001). The expression of HLA class I was related to LMP7 expression in contrast to TAP1 (P=0.001 vs. P=0.381). Presence of CD8-positive T-cells was not related to JAK1 mutation (Figure 1).
Table 2: Expression of antigen processing machinery components TAP1, LMP7 and HLA class I in JAK1 wildtype and mutant MSI endometrial cancers
JAK1 wildtype n=36 (62.1%) |
JAK1 mutation n=22 (37.9%) |
P-value |
|
|---|---|---|---|
TAP1* |
0.151 |
||
Impaired/Normal |
24 (68.6) |
18 (85.7) |
|
Upregulated |
11 (31.4) |
3 (14.3) |
|
LMP7* |
0.074 |
||
Impaired/Normal |
8 (22.9) |
10 (45.5) |
|
Upregulated |
27 (77.1) |
12 (54.5) |
|
HLA class I |
<0.001 |
||
Impaired |
7 (19.4) |
12 (54.5) |
|
Normal |
3 (8.3) |
6 (27.3) |
|
Upregulated |
26 (72.2) |
4 (18.2) |
* 1 missing value
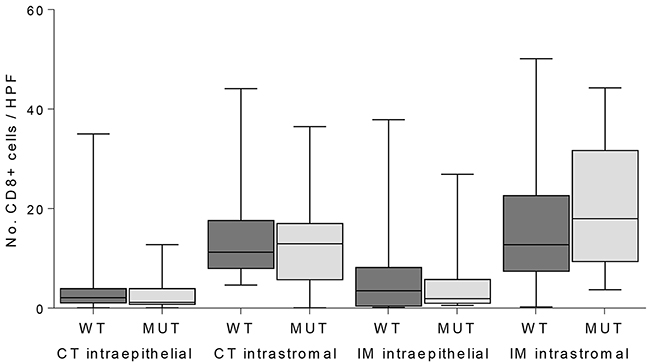
Figure 1: Quantification of CD8-positive T-cells in intraepithelial and intrastromal components in the center of the tumor (CT) and invasive margin (IM) in JAK1 wildtype (WT) and mutant (MUT) MSI endometrial cancers.
In order to validate these findings in an independent cohort, RNAseq data was used from 25 TCGA MSI endometrial cancers with analyzed DNA slippage events. The 13 JAK1 mutant endometrial cancers showed significantly lower expression of TAP1 (2.1-fold, P<0.001), LMP7 (3.0-fold, P<0.001), and HLA class I (2.5-fold, P<0.001) in comparison to JAK1 wildtype endometrial cancers (Figure 2). Consistent with the results in our study cohort, JAK1 mutation status did not correlate with CD8 expression (P=0.112).
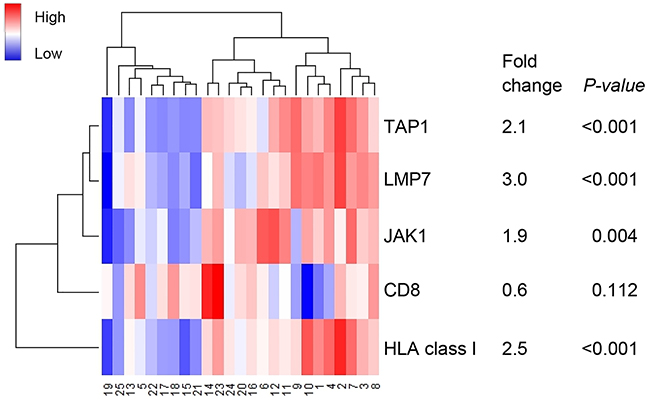
Figure 2: Heatmap of RNA expression of genes encoding for antigen machinery components. Case numbers 1-12 represent JAK1 wildtype and 13-25 JAK1 mutant MSI endometrial cancers. A broader analysis independent of MSI status was previously shown by Kim et al.
Two patient cohorts derived from the PORTEC-1 and -2 randomized trials with MSI early-stage endometrial cancers (n=198) with long-term mature follow-up data were used to investigate a possible prognostic effect of JAK1 frameshift mutations. In this independent cohort, fifty-two (28%) of MSI endometrial cancers had a JAK1 mutation (Supplementary Table S1). No significant differences were found between JAK1 mutation status and clinicopathological characteristics, and the association of JAK1 mutation and deep myometrial invasion could not be confirmed (Table 1). This discrepancy could be explained by the fact that this cohort consisted of significant more tumors with deep myometrial invasion compared to the study cohort (Supplementary Table S2). For JAK1 wildtype and mutant endometrial cancers, 10-year recurrence free rates were 84% versus 77%, respectively (P=0.301) and 10-year overall survival was 64.4% and 63.5% (P=0.716) (Figure 3). Neither did subanalysis (e.g. in grade 3 cancers, or when analyzing only pelvic and distant recurrences) show a significant difference in outcome between JAK1 wildtype and mutant endometrial cancer patients. Survival analysis within the TCGA microsatellite unstable endometrial cancers with known JAK1 mutation status (n=30) showed neither a survival benefit for JAK1 wildtype tumors (data not shown).
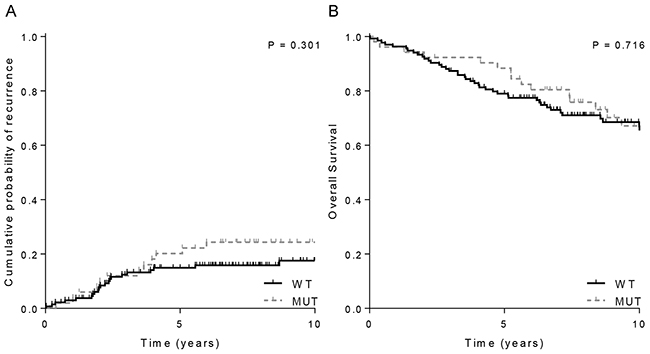
Figure 3: Clinical outcome of 198 MSI endometrial cancer stratified by JAK1 mutations status. A. Overall recurrence rate and B. Overall survival. WT= wildtype; MUT=mutant
.DISCUSSION
This research shows that frameshift mutations in JAK1 frequently and almost exclusively occur in MSI endometrial cancers. JAK1 mutations were associated with impaired upregulation of antigen presenting machinery proteins LMP7 and HLA class I. The association of JAK1 mutations with limited expression of the antigen presentation pathway was validated using RNAseq data of the TCGA MSI endometrial cancers. Impaired upregulation of HLA class I expression interferes with tumor lysis by cytotoxic T-cells, and therefore JAK1 mutations may facilitate an immune escape. However, no effect was observed of JAK1 mutation status on recurrence rate and overall survival in a large independent cohort of 198 MSI endometrial cancers. These findings suggest a functional role for JAK1, although with no prognostic value which suggests that JAK1 mutations are pivotal to cancer initiation and/or maintenance, in an intriguing tissue-specific manner.
The overall JAK1 mutation frequency of 28% in our large series of MSI endometrial cancers is in line with findings from two recent smaller studies [4, 5]. Both studies also showed that JAK1 frameshift mutations in MSI tumors are tissue specific and significantly less important in colorectal cancers as compared with endometrial cancers. Similarly, Ren et al. identified JAK1 frameshift mutations mainly in gynecological cancers, primarily in endometrial and cervical cancer, when sequencing more than 3,000 tumors from various human tissues [10]. Altogether, the occurrence of JAK1 mutation specifically in endometrial cancer is suggestive of a positive selection for this mutation in endometrial cancer.
To date, there is very little evidence for the functionality of JAK1 frameshift mutations in endometrial cancer. An in vitro study with one endometrial cancer and two ovarian cancer cell lines harboring JAK1 frameshift mutations demonstrated that JAK1 mutations impede STAT1 posphorylation and upregulation of antigen presenting machinery components LMP2 and TAP1 [10]. Kim et al. have shown that JAK1 mutations were associated with hampered JAK/STAT signaling and lymphocyte activation [4]. These findings suggest that JAK1 mutations have a negative effect on tumor immune surveillance. Our study now confirmed impaired upregulation of LMP7 and HLA class I with no effect on TAP1 expression and the number of CD8-positive T-cells in JAK1 mutant endometrial cancer tissue samples. The findings on protein expression were validated using RNAseq data of TCGA MSI endometrial cancers, except for TAP1. TAP1 gene expression may not be equal to its protein expression because of the small number of cases with RNAseq data (n=25) or due to the (post-)translational process into proteins. The lack of correlation between JAK1 mutations and CD8-positive T-cells might be explained by the fact that recruitment and migration of T-cells do not rely on recognition of peptides presented by HLA class I molecules. In addition, previous studies have also shown a high correlation between LMP7 expression and HLA class I expression, but not with LMP2 or TAP1/2 [21, 22]. This further strengthens the argument that JAK1 mutations favor immune escape via the JAK/STAT signaling, although, this study did not evaluate the activation status of the JAK/STAT signaling by phospho-STAT1 expression.
The association of JAK1 mutations and lack of HLA class I upregulation in MSI endometrial cancers with no effect on clinical course was an unanticipated finding of our study. Bijen et al. and Yakabe et al. reported impaired upregulation of HLA class I as a prognostic marker for survival in endometrial cancer patients [13, 15]. Of note, a large proportion of HLA class I negative endometrial cancers are MSI [14], therefore, separate analysis of MSI and MSS tumors would be of interest to determine the prognostic impact of HLA class I. This study indirectly showed no effect of HLA class I expression on survival via the JAK1 mutation status in a large cohort of MSI, early-stage endometrial cancers. HLA class I expression was not analyzed on the precious PORTEC tissue samples as this will only validate previous findings of Ren et al., the study and TCGA cohort and the fact that JAK1 mutations have no effect on survival will remain. However, the relatively good prognosis of this cohort of early-stage cancers needs to be taken into account. The previous studies were performed in heterogeneous groups of endometrial cancers consisting of different histologic subtypes and different FIGO stages [13-15]. Therefore, the prognostic role remains to be determined in higher risk endometrial cancers. These results, in contrast to the finding that JAK1 mutations favor immune evasion, may suggest a lack of negative selection of JAK1 mutations specifically in endometrial cancer.
Considering the process of immune surveillance in cancer [11, 12], it is likely that JAK1 mutant tumors are still recognizable to cells of the innate immune system (natural killer cells) or that JAK1 wildtype tumors have encountered other mechanisms to evade immune-mediated killing. However, low numbers of natural killer cells that lack an association with HLA class I expression were observed in endometrial cancer (unpublished data by Versluis et al.). On the contrary, IFNγ production might be responsible for a CD4-positive T-cell-mediated antitumor immunity [23]. ]. Furthermore, JAK1 wildtype tumors may not demonstrate a survival benefit because of other strategies to dampen immune response such as upregulation of anti-apoptotic molecules, expression of immune-inhibitory ligands or secretion of immunosuppressive cytokines. Nevertheless, our findings imply that JAK1 mutations may exert in part its oncogenic effects by immune escape, but we cannot exclude other contributions of JAK1 in the JAK/STAT signaling [7].
Better understanding of the antigen-specific immune responses and tumor microenvironment may guide immunotherapy. Recently, immune checkpoint blockade were reported as promising therapies for tumors with a high mutational load, including mismatch repair deficient endometrial cancers, as a result of an increased neo-antigen specific T-cell response [17]. JAK1 mutations in MSI endometrial cancers may interfere with the T-cell response due to impaired HLA class I or PD-L1 expression [17, 18]. In melanoma, one patient without a clinical response to PD-1 blockade and increased T-cell response showed a loss of function JAK1 mutation that unables PD-L1 upregulation. All other fifteen melanoma patients did not show genetic alterations in the interferon receptor signaling pathway [18]. These limited data suggest that JAK1 mutations may be used as negative selective predictive biomarker for immune blockade therapy. However, additional studies are required on the immune microenvironment of JAK1 mutant endometrial cancers (e.g. PD-L1 expression).
We have identified a high frequency of JAK1 mutations in MSI endometrial cancers in two relatively large series of MSI endometrial cancers. In addition, JAK1 mutations may have a negative effect on tumor immune surveillance due to lack of HLA class I upregulation on the cell surface. It must be noted that it remains unclear why JAK1 mutations are limited to gynecological cancers and mainly to MSI endometrial cancer. However, B2M frameshift mutations, also leading to immune escape via loss of HLA class 1 expression, frequently occur in MSI colorectal cancer and are a rare phenomenon in MSI endometrial cancer [14, 24]. In addition, Xiong et al. showed that JAK1 inhibition is associated with cell cycle arrest and apoptosis in colorectal cancer [25]. No effect on cell viability upon IFNγ was found in JAK1 mutant gynecological cell lines [10]. In conclusion, we confirmed the remarkably high frequency of JAK1 mutations and associations with impaired upregulation of antigen presenting machinery components in MSI endometrial cancers, which suggest a functional role for JAK1 in an intriguing tissue-specific manner.
MATERIALS AND METHODS
Patients
DNA analysis and immunohistochemical staining was performed on a study cohort of 181 endometrial cancers with endometrioid histology, treated at the University Medical Center Groningen between 1985-2004 or at the University Medical Center Leiden between 2000-2013. Classification and grading was done according to the World Health Organization criteria and staging was according to FIGO guidelines (2009). No follow-up data was available for this study cohort.
To validate our findings regarding the JAK1 mutation frequency in MSI endometrial cancer, an independent cohort of 198 MSI early-stage endometrial cancers derived from the randomized PORTEC-1 and -2 clinical trials was used [19, 20]. To estimate the impact of JAK1 mutation on survival the same cohort of 198 MSI endometrial cancers was used [26].
MSI and JAK1 mutation status
DNA was isolated as previously described [27]. The MSI status of each tumor was determined using the Promega MSI analysis system (version 1.2, Promega). Tumors with instability in at least two markers were defined as being high-frequency MSI whereas those showing no instability or instability in one marker were classified as being stable (MSS). JAK1 frameshift mutations (k142fs, p430fs, k860fs) were detected by Sanger sequencing. The following primers were used: exon 5-F: 5’-GTCACATCTGGGTCCCCTTTGCCAC-3’, exon 5-R: 5’-CACAAACTCCAGCTTCTCCTGGGC-3’, exon 9-F: 5’-GTCGAGGAGGCCTTG TCCTTTGTGTC-3’, exon 9-R: 5’-ACACGGGCTCTCTGCACACC-3’, exon 19-F: 5’-GTATCGACTGCCTTTCACTCTG-3’, exon 19-R: 5’-CTTACCTCTCCCAAGTCACGG-3’.
Immunohistochemistry
Formalin-fixed paraffin embedded 4-μm tissue sections of MSI endometrial cancers (n=58) with sufficient tumor tissue were immunohistochemically stained for expression of LMP7, TAP1, HLA class I (HCA2 and HC10) and CD8 (marker of cytotoxic T-cells). Sections were deparaffinized in xylene, rehydrated in graded concentration of ethanol and microwave antigen retrieval was performed in 10 mM citrate pH 6.0 (LMP7, TAP1), 10 mM Tris/1 mM EDTA pH 9.0 (LMP7, TAP1, HCA2, HC10 and CD8) before staining. Endogenous peroxidase was blocked by incubation in a 0.3% hydrogen peroxide solution. LMP7 and TAP1 were stained using anti-LMP7 mouse monoclonal 1B3 (Novus Biologicals) and anti-TAP1 rabbit polyclonal H300 (Santa Cruz) as primary antibodies by incubation overnight at 4°C (dilution 1:100 and 1:50 respectively). HLA class I was stained using HCA2 and HC10 as previously described [16]. Antigen-antibody reactions were visualized using 3.3’-diaminobenzidine (DAB) and slides were counterstained with hematoxylin.
Evaluation of immunohistochemistry
Two observers blinded to clinicopathological features, MSI and JAK1 mutation status independently evaluated the stained slides. Expression of LMP7 and TAP1 was scored using a semiquantative scale as described previously [13, 28]. This score is based on the percentage of cells stained and the intensity of staining. The percentage of cells was scored on a 6 point scale with 0 for 0%, 1 for 1-5%, 2 for 5-25%, 3 for 25-50%, 4 for 50-75% and 5 for 75-100%. The intensity was scored on a 4 point scale with 0 indicating absence of staining and 3 indicating strong staining. The expression of LMP7, and TAP1 was categorized in impaired (score 0-2), normal (score 3-6) and upregulated (score 7-8) expression. For analysis of HLA class I expression, the percentage of tumor cells with membranous HCA2 and HC10 staining was quantified as previously described [16]. The expression of HLA class I was defined as follows: impaired HLA class I expression: less than 5% of tumor cells expressing both HCA2 and HC10, normal HLA class I expression; less than 5% of tumor cells expressing either of the markers, and upregulated HLA class I expression: 5% or more expressing both markers.
The number of CD8-positive T-cells was calculated using the average number of stained cells in 8 fields at 40x magnification. The average was calculated for four locations: intraepithelial at the tumor center, intraepithelial at the invasive margin, intrastromal in the tumor center and intrastromal at the invasive margin. For statistical analysis values for CD8 were dichotomized using the median as a cut off.
The cancer genome atlas (TCGA) RNAseq data
Details of the TCGA RNAseq analysis have been previously reported [1]. Level 3 RSEM normalized RNA data profiled using the Illumina HiSeq RNAseq v2 were retrieved at the TCGA data portal. MSI events, differences in length of microsatellites, in 30 MSI EC patients were reported by Kim et al [4]. In total, 25 MSI endometrial cancers with both RSEM normalized and DNA slippage event data were informative for analysis.
Statistical analysis
JAK1 mutation status was compared between cases with and without microsatellite instability using Chi-square tests. Similarly, Chi-square tests were used to detect differences in expression of LMP7, TAP1, HLA class I and number of CD8-positive T-cells below or above the median for cases with and without JAK1 mutation. The non-parametric Mann-Whitney test was used for all comparisons of continuous data and Spearman’s rho to analyze correlation between variables. RNAseq data was visualized by unsupervised clustering using RStudio.
To evaluate the impact of JAK1 mutation status on survival in MSI endometrial cancer patients that participated in the randomized PORTEC-1 and -2 clinical trials, time-to-event analyses were calculated from the date of randomization to date of recurrence (vaginal, pelvic and/or distant recurrence) or to date of death (overall survival); patients who were alive and without recurrence were censored at the date of last follow-up. Survival curves were calculated using the Kaplan—Meier method with log-rank test.
Analyses were performed using SPSS (v20, IBM statistics, Chicago, IL, USA).
ACKNOWLEDGMENTS
This study was supported by the Dutch Cancer Society (UL2012-5719). The PORTEC-1 and -2 trials were also supported by Dutch Cancer Society grants. We acknowledge all members of the PORTEC study group and the many patients who participated in the PORTEC trials.
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
FUNDING
Dutch Cancer Society.
REFERENCES
1. Kandoth C, Schultz N, Cherniack AD, Akbani R, Liu Y, Shen H, Robertson AG, Pashtan I, Shen R, Benz CC, Yau C, Laird PW, Ding L, et al. Integrated genomic characterization of endometrial carcinoma. Nature. 2013; 497:67-73.
2. Imai K and Yamamoto H. Carcinogenesis and microsatellite instability: the interrelationship between genetics and epigenetics. Carcinogenesis. 2008; 29:673-80.
3. Duval A, Reperant M, Compoint A, Seruca R, Ranzani GN, Iacopetta B and Hamelin R. Target gene mutation profile differs between gastrointestinal and endometrial tumors with mismatch repair deficiency. Cancer Res. 2002; 62:1609-12.
4. Kim TM, Laird PW and Park PJ. The landscape of microsatellite instability in colorectal and endometrial cancer genomes. Cell. 2013; 155:858-68.
5. Ferreira AM, Tuominen I, Sousa S, Gerbens F, van Dijk-Bos K, Osinga J, Kooi KA, Sanjabi B, Esendam C, Oliveira C, Terpstra P, Hardonk M, van der Sluis T, et al. New target genes in endometrial tumors show a role for the estrogen-receptor pathway in microsatellite-unstable cancers. Hum Mutat. 2014; 35:1514-23.
6. Myeroff LL, Parsons R, Kim SJ, Hedrick L, Cho KR, Orth K, Mathis M, Kinzler KW, Lutterbaugh J and Park K. A transforming growth factor beta receptor type II gene mutation common in colon and gastric but rare in endometrial cancers with microsatellite instability. Cancer Res. 1995; 55:5545-47.
7. Thomas SJ, Snowden JA, Zeidler MP and Danson SJ. The role of JAK/STAT signalling in the pathogenesis, prognosis and treatment of solid tumours. Br J Cancer. 2015; 113:365-71.
8. Aaronson DS and Horvath CM. A road map for those who don’t know JAK-STAT. Science. 2002; 296: 1653-55.
9. Murray PJ. The JAK-STAT signaling pathway: input and output integration. J Immunol. 2007; 178:2623-29.
10. Ren Y, Zhang Y, Liu RZ, Fenstermacher DA, Wright KL, Teer JK and Wu J. JAK1 truncating mutations in gynecologic cancer define new role of cancer-associated protein tyrosine kinase aberrations. Sci Rep. 2013; 3: 3042.
11. Vesely MD, Kershaw MH, Schreiber RD and Smyth MJ. Natural innate and adaptive immunity to cancer. Annu Rev Immunol. 2011; 29: 235-71.
12. Schreiber RD, Old LJ and Smyth MJ. Cancer immunoediting: integrating immunity’s roles in cancer suppression and promotion. Science. 2011; 331:1565-70.
13. Bijen CB, Bantema-Joppe EJ, de Jong RA, Leffers N, Mourits MJ, Eggink HF, van der Zee AG, Hollema H, de Bock GH and Nijman HW. The prognostic role of classical and nonclassical MHC class I expression in endometrial cancer. Int J Cancer. 2010; 126:1417-27.
14. de Jong RA, Boerma A, Boezen HM, Mourits MJ, Hollema H and Nijman HW. Loss of HLA class I and mismatch repair protein expression in sporadic endometrioid endometrial carcinomas. Int J Cancer. 2012; 131:1828-36.
15. Yakabe K, Murakami A, Nishimoto Y, Kajimura T, Sueoka K and Sugino N. Clinical implications of human leukocyte antigen class I expression in endometrial cancer. Mol Clin Oncol. 2015; 3:1285-90.
16. van Gool IC, Eggink FA, Freeman-Mills L, Stelloo E, Marchi E, de Bruyn M, Palles C, Nout RA, de Kroon CD, Osse EM, Klenerman P, Creutzberg CL, Tomlinson IP, et al. POLE Proofreading Mutations Elicit an Antitumor Immune Response in Endometrial Cancer. Clin Cancer Res. 2015; 21:3347-55.
17. Le DT, Uram JN, Wang H, Bartlett BR, Kemberling H, Eyring AD, Skora AD Luber BS, Azad NS, Laheru D, Biedrzycki B, Donehower TC, et al. PD-1 Blockade in Tumors with Mismatch-Repair Deficiency. N Engl J Med. 2015; 372:2509-20.
18. Shin D, Garcia-Diaz A, Zaretsky J, Escuin-Ordinas H, Hu-Lieskovan S, Berent-Maoz B, Comin-Anduix B and Ribas A. Innate resistance of PD-1 blockade through loss of function mutations in JAK resulting in inability to express PD-L1 upon interferon exposure. J Immunother Cancer. 2015; 3: P311.
19. Creutzberg CL, van Putten WL, Koper PC, Lybeert ML, Jobsen JJ, Warlam-Rodenhuis CC, De Winter KA, Lutgens LC, van den Bergh AC, Steen-basanik E, Beerman H and van Lent M. Surgery and postoperative radiotherapy versus surgery alone for patients with stage-1 endometrial carcinoma: multicentre randomised trial. PORTEC Study Group. Post Operative Radiation Therapy in Endometrial Carcinoma. Lancet. 2000; 355:1404-11.
20. Nout RA, Smit VT, Putter H, Jurgenliemk-Schulz IM, Jobsen JJ, Lutgens LC, van der Steen-Banasik EM, Mens JW, Slot A, Kroese MC, van Bunningen BN, Ansink AC, van Putten WL, et al. Vaginal brachytherapy versus pelvic external beam radiotherapy for patients with endometrial cancer of high-intermediate risk (PORTEC-2): an open-label, non-inferiority, randomised trial. Lancet. 2010; 375:816-23.
21. Cabrera CM, Jimenez P, Cabrera T, Esparza C, Ruiz-Cabello F and Garrido F. Total loss of MHC class I in colorectal tumors can be explained by two molecular pathways: beta2-microglobulin inactivation in MSI-positive tumors and LMP7/TAP2 downregulation in MSI-negative tumors. Tissue Antigens. 2003; 61:211-19.
22. Kang JK, Yoon SJ, Kim NK and Heo DS. The expression of MHC class I, TAP1/2, and LMP2/7 gene in human gastric cancer cell lines. Int J Oncol. 2000;16:1159-63.
23. Corthay A, Skovseth DK, Lundin KU, Rosjo E, Omholt H, Hofgaard PO, Haraldsen G and Bogen B. Primary antitumor immune response mediated by CD4+ T cells. Immunity. 2005; 22:371-83.
24. Kloor M, Michel S and von Knebel DM. Immune evasion of microsatellite unstable colorectal cancers. Int J Cancer. 2010; 127:1001-10.
25. Xiong H, Zhang ZG, Tian XQ, Sun DF, Liang QC, Zhang YJ, Lu R, Chen YX and Fang JY. Inhibition of JAK1, 2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces tumor cell invasion in colorectal cancer cells. Neoplasia. 2008; 10:287-97.
26. Stelloo E, Nout RA, Osse EM, Jurgenliemk-Schulz IJ, Jobsen JJ, Lutgens LC, van der Steen-Basanik EM, Nijman HW, Putter H, Bosse T, Creutzberg CL, Smit VT. Improved risk assessment by integrating molecular and clinicopathological factors in early-stage endometrial cancer - combined analysis of PORTEC cohorts. Clin cancer res. 2016.
27. Stelloo E, Bosse T, Nout RA, MacKay HJ, Church DN, Nijman HW, Leary A, Edmondson RJ, Powell ME, Crosbie EJ, Kitchener HC, Mileshkin L, Pollock PM, et al. Refining prognosis and identifying targetable pathways for high-risk endometrial cancer; a TransPORTEC initiative. Mod Pathol. 2015; 28:836-44.
28. Ruiter DJ, Ferrier CM, van Muijen GN, Henzen-Logmans SC, Kennedy S, Kramer MD, Nielsen BS and Schmitt M. Quality control of immunohistochemical evaluation of tumour-associated plasminogen activators and related components. European BIOMED-1 Concerted Action on Clinical Relevance of Proteases in Tumour Invasion and Metastasis. Eur J Cancer. 1998; 34:1334-40.