
INTRODUCTION
While nasopharyngeal carcinoma (NPC) is rare in most populations worldwide, the incidence peaks in South China, where NPC occurs in 50 out of every 100,000 individuals [1, 36]. In many of these cases, Epstein-Barr virus (EBV) detection may be useful for detecting NPC and might be a prognostic marker [5]. Elevated IgA antibodies against viral capsid antigen (VCA), early antigen (EA) and nuclear antigens (EBNA1) are outstanding features of such NPC patients [4]. Compared with other antibodies, IgA-VCA is still the most sensitive serological EBV antibody [6], and is detectable even before the development of NPC [4, 37]. Recently, plasma EBV DNA, another biomarker in NPC patients, was used for diagnostic purposes [38] as well as for monitoring NPC patient response to therapy and risk of relapse [7, 8]. Regardless of the high sensitivity and specificity of IgA-VCA in the diagnosis of NPC, IgA-VCA was undetectable in 4 - 24% of patients [6, 9-12], and resulted in misdiagnosis. Quantitative polymerase chain reaction (qPCR) analysis of circulating EBV DNA resulted in tumor detection sensitivities of 22 - 86%, 48 - 95%, 74 - 100% and 79-100% in patients with stage I, II, III and IV disease, respectively [9]. Unlike patients with stage I NPC, which has a 5-year survival rate of 100% after treatment, and stage II, which has a 5-year survival rate of 95% after treatment, the prognosis of patients with stage IV disease is usually poor [9, 13]. Therefore, reliable diagnostic biomarkers to complement IgA-VCA or EBV DNA are required to improve diagnostic accuracy.
EBV infection of epithelial cells requires three viral glycoproteins, gB, gH, and gL, which are conserved among herpesviruses [14, 15]. EBV infection of B cells also requires the viral glycoprotein gp42, which binds the cell-surface major histocompatibility complex (MHC) class II molecules [14, 15]. Furthermore, EBV glycoproteins play an important role in the humoral immune response, and sera against these proteins often neutralize the virus [16]. Therefore, we propose that glycoproteins have the potential to serve as diagnostic biomarkers for NPC.
In the present study, we developed a novel ELISA using a baculovirus-expressed gH (aa 18-679) and gL (aa 24-137) protein complex to test the feasibility of antibodies against gH/gL in the detection of patients with NPC. We compared the sensitivities and specificities of these antibodies separately and in combination with IgA-VCA or circulating EBV DNA in the detection of NPC.
RESULTS
Production and purification of soluble EBV gH/gL protein
The stability and proper folding of gH requires gL. Based on preliminary experiments (data not shown), cells were infected with the baculovirus at an MOI of 5 and harvested 72 h post-infection. After purification with Ni chelate-Sepharose chromatography, proteins were quantified and purity was evaluated. SDS/PAGE gels revealed two major bands of 85 kDa and 26 kDa, which corresponded to the recombinant gH and gL proteins, respectively (Figure S1A). Immunoblotting with an anti-flag monoclonal antibody revealed specific bands of the same size (Figure S1B), which suggests that the soluble gH/gL proteins are the major components of the purified proteins. The observed molecular masses corresponded well to the combination of theoretical amino acid sequences and putative N-linked glycosylation. Thus, the purified proteins were considered to have specific antigenicity and could be used in a subsequent ELISA.
Elevated levels of gH/gL antibodies in NPC patients
To determine whether gH/gL could be applied for NPC diagnosis, we developed an ELISA to detect anti-gH/gL antibodies in sera from a training cohort (n = 208 patients with NPC and 198 healthy controls). A comparison of NPC patients with healthy controls (Table S1) showed that only family history (p < 0.001) was significant. Age (p = 0.973), sex (p = 0.388) and smoking history (p = 0.622) were not significant.
We evaluated the distribution of blood IgA antibodies against EBV gH/gL in patients using OD values (median ± IQR [IQR, interquartile range]). Antibody titers against gH/gL were elevated in a majority of patients with NPC compared to controls (Figure 1A). The median gH/gL OD value for NPC patients was 0.84±0.37, which was higher than that of the healthy controls (0.49±0.18) (p < 0.001).
The distribution of IgA-gH/gL levels according to the individual patient’s cancer stage is shown in Figure 1B. We found that the median IgA-gH/gL OD value for patients with stage IV NPC (0.96±0.35) was higher than that of early stage (I+II) (0.81±0.28) and stage III patients (0.79±0.35), but this was not statistically significant (I+II vs. III p = 0.284, I+II vs. IV p = 0.204, III vs. IV p = 0.671). Additionally, we did not observe correlations between antibody level and other patient clinical characteristics, such as age, gender, smoking history and IgA-VCA or EBV DNA status (Table 1).
Table 1: Associations of EBV IgA-gH/gL and NPC patient clinicopathological parameters in the training cohort.
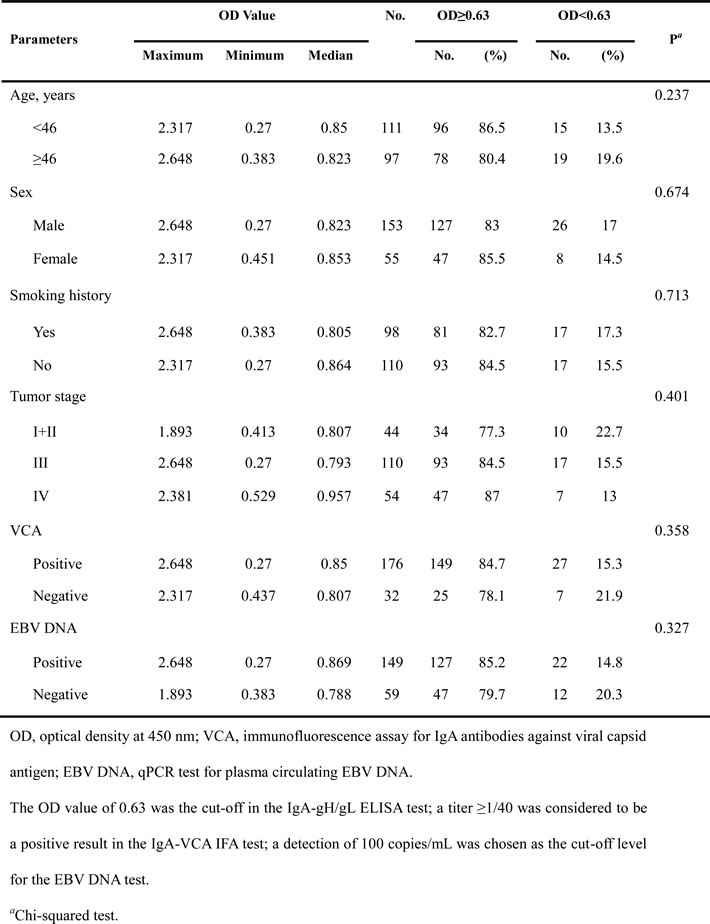
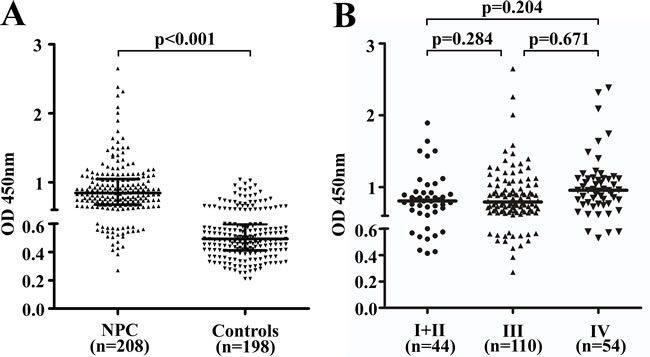
Figure 1: Characteristics and diagnostic values of IgA-gH/gL in the training cohort. Scatter plots of the distribution of IgA-gH/gL ELISA results for NPC cases (n = 208) and healthy controls (n = 198) A. Black horizontal lines are medians. The upper black horizontal line indicates the 75th percentile of the data set, and the lower line indicates the 25th percentile. ELISA results for pretreatment serum of NPC patients with stage I (n = 10), II (n = 34), III (n = 110) or IV (n = 54) disease B. Black horizontal lines are medians. The IgA-gH/gL OD value distributions were not significantly different between stages.
Diagnostic values of IgA-gH/gL for NPC in the training cohort
IgA-gH/gL was evaluated as a potential marker for the diagnosis of NPC using ROC (receiver operating characteristic) analysis based on OD values (Figure 2A, Table 2). Using a cut-off OD value of 0.63 for the gH/gL test, the IgA-gH/gL ELISA had a sensitivity of 83.7%, specificity of 82.3%, positive predictive value (PPV) of 83.3%, negative predictive value (NPV) of 82.7%, and AUC (area under the curve) of 0.893 (95% CI, 0.862-0.924). Compared to IgA-gH/gL, the IgA-VCA IFA had a similar sensitivity of 84.6% (p = 0.89), higher specificity of 93.4% (p = 0.002) and similar AUC of 0.933 (95% CI, 0.906 - 0.959) (p = 0.053). However, circulating EBV DNA had the lowest sensitivity at 71.6% (p = 0.004), AUC of 0.849 (95% CI, 0.810 - 0.889) (p = 0.046) and highest specificity of 94.9% (p < 0.001). These results showed that the diagnostic capacity of gH/gL was comparable to that of the other two EBV markers.
Table 2: AUC, sensitivity, specificity, PPV and NPV of IgA-gH/gL, IgA-VCA, EBV DNA and their combinations for detection of NPC in the training cohort.
Establishment of a logistic regression model combining IgA-gH/gL and IgA-VCA
Because IgA-gH/gL alone did not show an advantage in the detection of NPC in comparison to IgA-VCA, we used a binary logistic regression model to assess whether a combination of EBV markers could improve diagnostic efficiency (Figure 2B, Table 2). The combination of IgA-gH/gL and IgA-VCA achieved similar sensitivity (88.5%), specificity (97%), PPV (96.8%) and NPV (88.9%) and an AUC of 0.980 (95% CI, 0.970 - 0.991) as the combination of IgA-VCA and EBV DNA (87.5%, 97.4%, 97.3%, 88.1%, and 0.967 [95% CI, 0.949 - 0.985], respectively), which was superior to the combination of IgA-gH/gL and EBV DNA (87.5%, 89.4%, 89.7%, 87.2%, and 0.941 [95% CI, 0.919 - 0.963], respectively). We selected the logistic regression model that combines IgA-VCA with the IgA-gH/gL ELISA as the new optimal combination for NPC detection. The following formula was established:
Log (![]() ) = -7.36 + 0.055 × VCA + 8.133 × gH/gL
) = -7.36 + 0.055 × VCA + 8.133 × gH/gL
where VCA represents the reciprocal transformation of the titer of IgA to VCA and gH/gL represents the OD at a wave length of 450 nm of IgA to gH/gL.
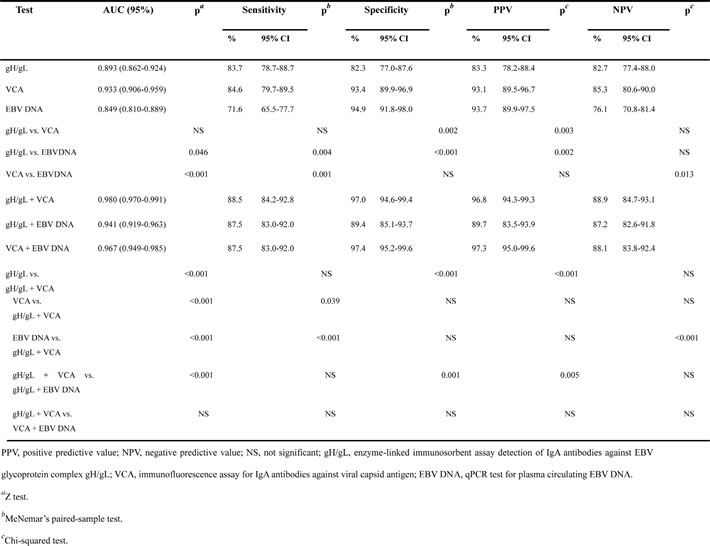
In this model, the ROC analysis (Figure 2B) showed that testing of both VCA and gH/gL increased the NPC diagnostic capacity (AUC, 0.98; 95% CI, 0.97 - 0.991) compared to either individual test. A total of 201 (96.6%) of the 208 NPC patients had positive results when VCA and gH/gL were tested together (Figure 2D). Furthermore, 25 (78.1%) of 32 VCA-negative patients with NPC had positive gH/gL results (Figure 2E). The rate was similar (150 [85.2%] of 176) to that observed in the VCA-positive patients (Figure 2E). Fifteen IgA-VCA-negative NPC patients were at stage I or II and the rate of positive gH/gL among them was 66.7%. Seventeen IgA-VCA-negative NPC patients were at stage III or IV and the rate of positive gH/gL among them was 88.2%. However, we did not find any significant difference among different clinical stages (p = 0.337). The ROC curves for gH/gL indicated a diagnosis of NPC in patients with negative VCA (Figure 2C), with a sensitivity of 78.1% and AUC of 0.879 (95% CI, 0.820 - 0.937). In the case of EBV DNA, the most specific assay with the highest positive predictive value was the combination of IgA-VCA and EBV DNA, but only 17 (53.1%) of the 32 VCA-negative patients with NPC had positive EBV DNA results (Figure 2E). This rate was lower than that of the VCA-positive patients (132 [75%] of 176). EBV DNA had a sensitivity of 53.1% and AUC of 0.750 (95% CI, 0.637 - 0.863); these values were inferior to those of gH/gL (Table 3).
Table 3: AUC, sensitivity, specificity, PPV and NPV of IgA-gH/gL and EBV DNA in IgA-VCA-negative NPC patients in the training cohort.
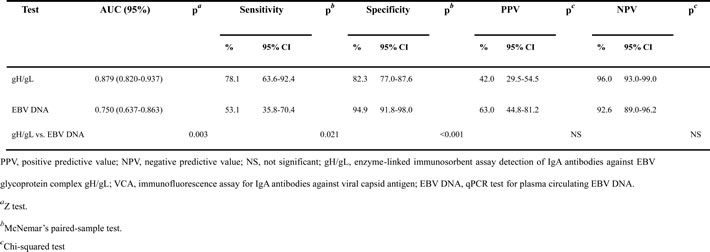
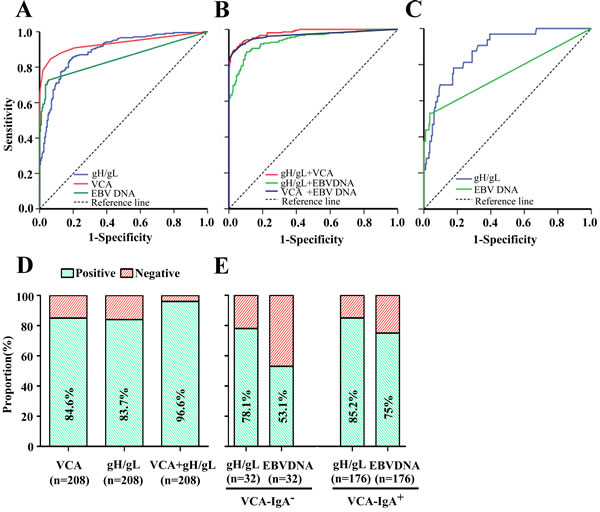
Figure 2: Diagnostic outcomes of gH/gL, VCA, EBV DNA and their combinations for detection of NPC in the training cohort. ROC curve for gH/gL, VCA or EBV DNA for NPC patients vs. controls A. ROC curves for the combination of gH/gL and VCA, gH/gL and EBV DNA, or VCA and EBV DNA for NPC patients vs. controls B. ROC analyses of antibody responses to gH/gL or plasma EBV DNA concentrations in VCA-negative NPC patients, showing an AUC of 0.879 (95% CI, 0.820-0.937) for IgA-gH/gL and of 0.750 (95% CI, 0.637-0.863) for EBV DNA C. Rates of correct NPC diagnosis for VCA, gH/gL or both in NPC patients D. Rates of correct NPC diagnosis for gH/gL and EBV DNA in VCA-negative or VCA-positive patients E.
Validating the logistic regression model combining IgA-gH/gL and IgA-VCA
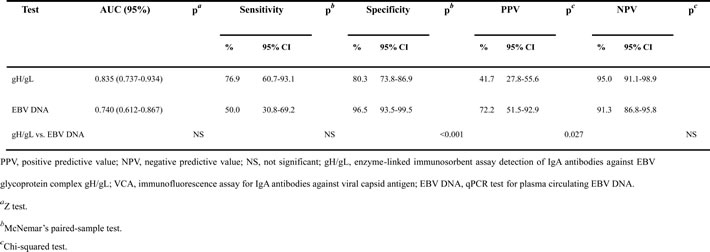
To further verify the efficacy of the regression model estimated from the training data set, we applied it in an independent validation cohort to diagnosis NPC (n = 137 patients with NPC and 142 healthy controls). With a cut-off OD value of 0.63 for the IgA-gH/gL alone, we observed similar results in the validation cohort as in the training cohort (Figure 3A, Table 4). IgA-gH/gL ELISA had good diagnostic capacity with a sensitivity of 86.9%, specificity of 80.3%, positive predictive value (PPV) of 81%, negative predictive value (NPV) of 86.3%, and AUC of 0.912 (95% CI, 0.878 - 0.947). Additionally, the validation cohort confirmed the ability of IgA-gH/gL to diagnose NPC in VCA-negative patients, with a sensitivity of 76.9% and AUC of 0.835 (95% CI, 0.737 - 0.934) (Table 5, Figure 3C). Twenty (76.9%) of 26 VCA-negative patients with NPC had positive gH/gL results (Figure 3E). This rate was similar to that observed in the VCA-positive patients (99 [89.2%] of 111) (Figure 3E). Nine IgA-VCA-negative NPC patients were at stage I or II and the rate of positive gH/gL among them was 88.9%. The improvement in NPC diagnosis resulting from measurement of IgA-gH/gL and IgA-VCA together was also observed in the validation cohort (Table 4, Figure 3B). ROC analysis showed that the newly established regression model had a sensitivity of 91.2%, specificity of 96.5%, PPV of 96.2%, NPV of 91.9%, and AUC of 0.97 (95% CI, 0.951 - 0.988). A total of 131 (95.6%) of 137 NPC patients were diagnosed correctly when VCA and gH/gL were tested together (Figure 3D).
Table 4: AUC, sensitivity, specificity, PPV and NPV of IgA-gH/gL, IgA-VCA, EBV DNA and their combinations for detection of NPC in the validation cohort.
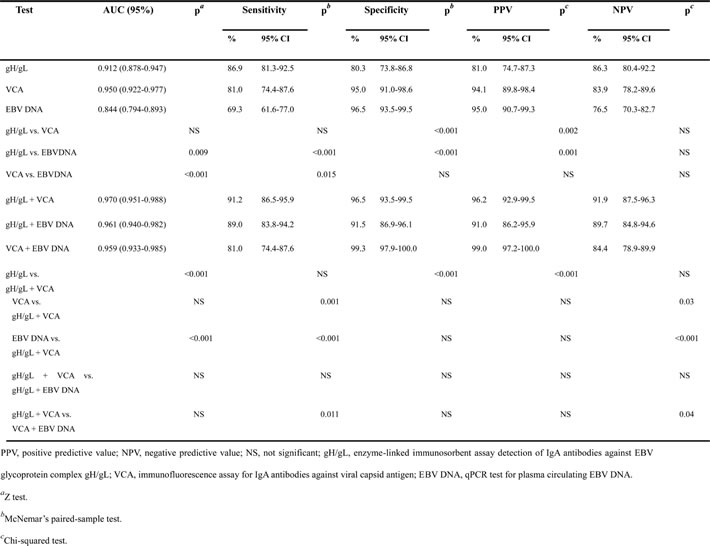
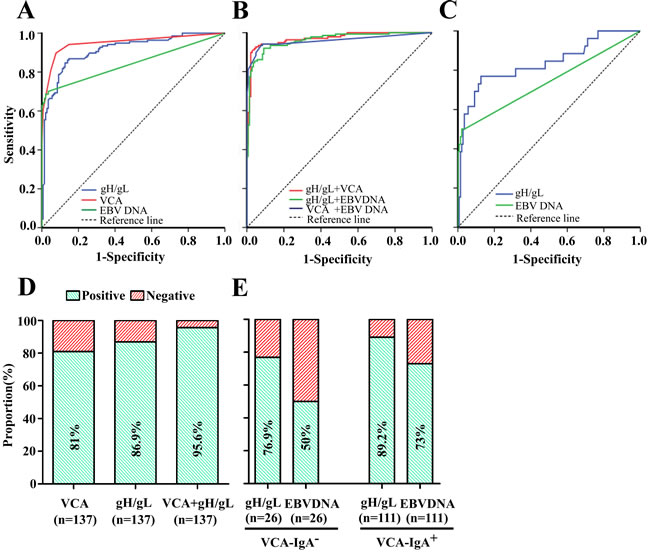
Figure 3: Diagnostic outcomes of gH/gL, VCA, EBV DNA and their combinations for detection of NPC in the validation cohort. ROC curve for gH/gL, VCA or EBV DNA for NPC patients vs. controls A. ROC curves for the combination of gH/gL and VCA, gH/gL and EBV DNA or VCA and EBV DNA NPC patients vs. controls B. ROC analyses of antibody responses to gH/gL or plasma EBV DNA concentrations in VCA-negative NPC patients, showing an AUC of 0.835 (95% CI, 0.737-0.934) for IgA-gH/gL and of 0.740 (95% CI, 0.612-0.867) for EBV DNA C. Rates of correct NPC diagnosis for VCA, gH/gL or both in NPC patients D. Rates of correct NPC diagnosis for gH/gL and EBV DNA in VCA-negative or VCA-positive patients E.
DISCUSSION
In this study, we found that IgA-gH/gL had a similar sensitivity for the diagnosis of NPC compared with IgA-VCA IFA (p = 0.89), but was more sensitive than plasma EBV DNA (p = 0.004). IgA-gH/gL had an AUC of 0.893 (95% CI, 0.862-0.924), which was better than that of EBV DNA (p < 0.05). The combination of IgA-gH/gL and IgA-VCA could improve the detection of NPC in patients.
Table 5: AUC, sensitivity, specificity, PPV and NPV of IgA-gH/gL and EBV DNA in IgA-VCA-negative NPC patients in the validation cohort.
We chose IgA-VCA as the reference marker because it has been widely used in clinical diagnosis and population screening studies due to its relatively high sensitivity and specificity. However, the EBV serological spectrum differs among individuals, and 4 - 24% of patients with NPC remain negative for VCA [6, 9-12]. Consequently, a combination of different markers, especially novel markers with high diagnostic capacity among VCA-negative patients, is extremely important for diagnostic accuracy. A combination of circulating EBV DNA and IgA-VCA, studied by Leung, et al. [17], showed that seven NPC patients who had false negative results for EBV DNA did not overlap with the 27 NPC patients who had false negative results for IgA-VCA, except in two cases. Similarly, Dardari, et al. [18] reported that 89% of sera from young Maghrebian NPC patients that were negative for IgA-VCA were positive at very high titers for IgG antibodies to the EBV transactivator protein (ZEBRA). More recently, positive IgA/VCA-p40+18 antibody ELISA results in 63.6% of nasopharyngeal carcinoma samples were missed entirely by peptide-based IgA/EBV-ELISAs (peptides derived from immunodominant epitopes of EBNA1 and VCA-p18) [19].
In our report, the sensitivity of IgA-gH/gL was close to that of IgA-VCA. Of note, 15.4% of all NPC patients in the training cohort were IgA-VCA negative; however, gH/gL maintained diagnostic capacity (25 of the 32 VCA-negative NPC patients had positive gH/gL results), and 201 (96.6%) of the 208 NPC patients were diagnosed correctly when VCA and gH/gL were tested together. The logistic regression analysis showed that IgA-VCA combined with IgA-gH/gL ELISA was an effective method for differentiating between NPC patients and controls, and these results were validated in a second independent cohort. Therefore, IgA-gH/gL detection complemented the IgA-VCA test for NPC diagnosis.
Plasma EBV DNA, which is another new non-protein serum marker for NPC screening, has also been reported to perform well in diagnostic tests [17, 20]. In our study, EBV DNA had the highest specificity (94.9%) and positive predictive value (93.7%) among the three biomarkers. The combination of EBV DNA and IgA-VCA showed excellent diagnostic performance, especially in specificity (97.4%) and positive predictive value (97.3%). However, EBV DNA might be more attractive as an independent prognostic factor for relapse risk or overall patient survival, because DNA load corresponded to NPC progression and remission [21].
EBV gH, also known as gp85, is an EBV membrane antigen encoded by the EBV late lytic gene BXLF2, which corresponds to the EBV lytic-cycle product [4]. The gH and gL proteins associate to form a heterodimer necessary for efficient membrane fusion [22]. Khanna, et al. [23] demonstrated that cytotoxic t-lymphocytes (CTLs) from acute infectious mononucleosis patients displayed strong ex vivo reactivity against EBV gH, and suggested that CTL epitopes from EBV gH could be exploited in the development of an EBV vaccine. Urquiza, et al. [24] found that peptide 11438 from the EBV gH host cell-binding region inhibited EBV infection of peripheral blood mononuclear cells (PBMCs). The peptide induced production of antibodies that recognize EBV and EBV-infected cells.
Additional studies have associated a host immune response with other types of herpesvirus gH. For example, varicella-zoster virus gH/gL is reactive with the anti-human gamma chain of IgG near the glycosylation site [25], and baculovirus expression of recombinant chelonid fibropapillomatosis-associated herpesvirus (CFPHV) gH has been successfully applied to detect turtle antibodies to recombinant CFPHV gH using ELISA [26]. The application of EBV glycoproteins in NPC screening has also been previously reported. gp350/220 induces a strong immune response in EBV-infected individuals, and several studies have detected IgA-gp350/220 in NPC patients at higher titers than in healthy controls [27, 28]. Although the function of gp78 remains unknown, the sensitivities and specificities of IgA-gp78 and IgG-gp78 have both exceeded 70% as reported by Gu, et al. [29]. The VCA complex protein, the antigen most commonly used for serological diagnosis, is composed of several different proteins. One of these, gB (gp125), is believed to be a dominant immunogen of the VCA complex [30, 31], and several ELISA kits based on VCA-gp125 have been commercialized for NPC [6].
Our study had several limitations. The sample size was small, and to confirm the efficacy of IgA-gH/gL, a multi-center study is required. Additionally, detection of IgA antibodies against the EBV early antigen in sera was demonstrated to be a strong indicator of NPC [6, 9]; whether its combination with IgA-gH/gL could achieve a better diagnostic outcome warrants further exploration.
MATERIALS AND METHODS
Study population
A total of 208 serum samples in a training cohort were collected from histologically confirmed NPC patients before treatment. All patients were histologically diagnosed with WHO type II or III NPC [32] at the Sun Yat-sen University Cancer Center (SYSUCC), and had definite clinical stage NPC according to the seventh American Joint Committee on Cancer TNM staging manual [33]. We randomly selected 198 healthy controls, including 138 males and 60 females ranging in age from 30 to 60 years (mean age = 44.1 years), from the hospital staff who participated in the medical examination projects. A validation cohort comprising 137 patients with NPC and 142 healthy controls was also recruited from SYSUCC. We matched the groups in the two cohorts for age and sex as best as possible. The characteristics of the patients are presented in the Table S1. This blinded study was approved by the Clinical Research Ethics Committee of the SYSUCC, and all of the participants provided written informed consent.
Cell culture
High FiveTM insect cells (a gift from Yun Wang, Wuhan Institute of Virology, Chinese Academy of Sciences, Hubei, China, [email protected]) were used for protein expression and were grown in shaker flasks in Express Five serum-free medium (Invitrogen, Grand Island, N Y). Sf9 insect cells used for baculovirus production were grown in 150-cm2T flasks in Sf-900 II serum-free medium (Invitrogen). All media contained penicillin-streptomycin and amphotericin B (Sigma-Aldrich, St Louis, MO, USA).
EBV gH, EBV gL constructs
P2089 EBV plasmid (gift from Wolfgang Hammerschmidt, German Research Center for Environmental Health, Munich, Germany, [email protected]) was used as the template to PCR-amplify EBV gH/gL. The construction of soluble EBV gH/gL has been described previously [14]. In the expression constructs, gH (aa 18-679) [GenBank: NC009334] and gL (aa 24-137) [GenBank: YP001129472] were fused to the baculovirus gp64 signal sequence, and a flag epitope tag was inserted into the carboxyl termini of each protein. The related primers and gp64 are listed in Table S2. pFastBacHTB and DH10BacTM E. coli were kindly provided by Yun Wang.
Protein expression and purification
One liter of High FiveTM insect cells at a density of 2×106 cells/mL in shaker flasks was infected with the P3 gH/gL baculovirus at 27°C with stirring at 100 rpm. Supernatants were harvested at 72 h post-infection under the optimal experimental conditions. The supernatants were concentrated and sterile filtered. The expressed six-His-tagged protein was purified from supernatants by Ni chelate-Sepharose (Qiagen-Sample & Assay Technologies, Hilden, Germany) chromatography according to the manufacturer’s protocol. After washing the combined agarose with wash buffer (50 mM NaH2PO4, 300 mM NaCl, 20 mM imidazole and pH 8.0), purified recombinant proteins were eluted with elution buffer (50 mM NaH2PO4, 300 mM NaCl, 300 mM imidazole and pH 7.5). Eluted proteins were exchanged into PBS by ultrafiltration. Purified proteins were concentrated 10-fold, dialyzed and stored at -80°C.
SDS/PAGE and western blot analysis of the EBV gH/gL
Ten µL of eluted protein was denatured in 10 µL 2x sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS/PAGE) loading buffer (0.2% Bromophenol Blue, 20% (v/v) glycerol, 4% (w/v) SDS, 100 mmol/l Tris/HCl, pH 6.8, and 200 mmol/L dithiothreitol) by boiling at 98°C for 10 min. The samples were subjected to 10.5% acrylamide resolving gels, stained with Coomassie Blue for at least 30 min and then destained in methanol-acetic acid. Proteins were transferred onto PVDF membranes at 200 mA for 3 h. The membranes were blocked for 45 min at room temperature with 5% nonfat milk in PBS-T buffer (containing 0.1% Tween 20). Membranes were then incubated with primary antibody (1:1000; anti-flag tag mouse polyclonal antibody, flag-M2, Sigma-Aldrich) at 4°C overnight, followed by secondary antibody (anti-mouse IgG) coupled to horseradish peroxidase (HRP) for at least 45 min.
ELISA detection of antibodies to recombinant gH/gL
The purified recombinant gH/gL was dissolved in 10 mM PBS at pH 7.4. The 96-well microtiter plates were coated with 450 ng of gH/gL protein per well at 37°C for 2 h. After incubation, unoccupied sites were blocked with 3% BSA in PBS. The plates were washed five times with PBS containing 0.1% Tween-20 (PBS-T). After washing, serum samples (1:100 in PBST containing 3% BSA) were added and incubated for 1 h at 37°C. After five washes, HRP-labelled goat anti-human IgA antibodies (1:6000; Boster corporation, Wuhan, China) for gH/gL in PBS-T were added. The plates were incubated at 37°C for 30 min and washed before adding a tetramethylbenzidine reagent (Sigma-Aldrich, St Louis, MO, USA) for 10 min at 37°C. The reaction was stopped with 2 M H2SO4 and the optical density (OD) at 450 nm was determined using an ELISA reader.
IgA-VCA immunofluorescence assay and plasma EBV assay
The EBV-specific IgA-VCA antibody was assessed using a previously described immunofluorescent method [2, 6]; a titer ≥1/40 was considered to be positive. The procedures for real-time quantitative PCR (RT-qPCR) and measurement of plasma EBV DNA were described in previous studies [11, 34]. The RT-qPCR system was developed for plasma EBV DNA detection toward the BamHI-W region. The system consisted of the amplification primers W-44F (5’-AGT CTC TGC CTC AGG GCA-3’) and W-119R (5’-ACA GAG GGC CTG TCC ACCG-3’) and the dual-labeled fluorescent probe W-67T (5’-[FAM] CAC TGT CTG TAA AGT CCA GCC TCC [TAMRA]-3’). A detection level of 100 copies/mL was chosen as the cut-off level.
Statistical analysis
The results were analyzed using the statistical software SPSS for Windows (version 16.0), MedCalc (version 13.0.2.0) and GraphPad Prism (version 5.0). Participants’ characteristics and NPC risk factors were compared using chi-squared tests. The Mann-Whitney U test was used to analyze the differences between patients with NPC and healthy controls. ROC curves were constructed to assess sensitivity, specificity and area under the curve (AUCs) using 95% CI. The cut-off value for the biomarker was defined as the value with the highest sensitivity and specificity selected from the respective ROCs [35]. Chi-squared tests were used to compare the mean IgA-gH/gL OD among patients at different cancer stages. McNemar’s paired-sample test or chi-squared test was applied to determine whether the sensitivity, specificity, positive predictive values and negative predictive values of gH/gL, VCA, EBV DNA and their combinations were significantly different, and AUC comparisons were assessed using Z tests. Binary logistic regression models of all possible combinations of biomarkers were used to select the optimal combination for NPC diagnosis. P < 0.05 was considered to indicate statistical significance, and all of the statistical tests were two-sided.
ACKNOWLEDGMENTS
We are grateful to Professor Yun Wang for providing us with several important reagents for the baculovirus expression system.
GRANT SUPPORT
This study was supported by grants from the Ministry of Science and Technology of China (No. 2012CB967003 and 2011BAZ02879 to Mu-Sheng Zeng), grants from the National Natural Science Foundation of China (81520108022 to Mu-Sheng Zeng; 81272950 to Xiao-Ming Huang) and Health & Medical Collaborative Innovation Project of Guangzhou City, China (No. 201400000001 to Mu-Sheng Zeng).
CONFLICTS OF INTEREST
The authors declare that they have no conflict of interest.
REFERENCES
1. Wei KR, Zheng RS, Zhang SW, Liang ZH, Ou ZX and Chen WQ. Nasopharyngeal carcinoma incidence and mortality in China in 2010. Chin J Cancer. 2014; 33:381-387.
2. Xu FH, Xiong D, Xu YF, Cao SM, Xue WQ, Qin HD, Liu WS, Cao JY, Zhang Y, Feng QS, Chen LZ, Li MZ, Liu ZW, et al. An epidemiological and molecular study of the relationship between smoking, risk of nasopharyngeal carcinoma, and Epstein-Barr virus activation. J Natl Cancer Inst. 2012; 104:1396-1410.
3. Fang W, Zhang J, Hong S, Zhan J, Chen N, Qin T, Tang Y, Zhang Y, Kang S, Zhou T, Wu X, Liang W, Hu Z, et al. EBV-driven LMP1 and IFN-gamma up-regulate PD-L1 in nasopharyngeal carcinoma: Implications for oncotargeted therapy. Oncotarget. 2014; 5:12189-12202. doi: 10.18632/oncotarget.2608.
4. Chiu SH, Wu CC, Fang CY, Yu SL, Hsu HY, Chow YH and Chen JY. Epstein-Barr virus BALF3 mediates genomic instability and progressive malignancy in nasopharyngeal carcinoma. Oncotarget. 2014; 5:8583-8601. doi: 10.18632/oncotarget.2323.
5. Young LS and Dawson CW. Epstein-Barr virus and nasopharyngeal carcinoma. Chin J Cancer. 2014; 33:581-590.
6. Liu Y, Huang Q, Liu W, Liu Q, Jia W, Chang E, Chen F, Liu Z, Guo X, Mo H, Chen J, Rao D, Ye W, et al. Establishment of VCA and EBNA1 IgA-based combination by enzyme-linked immunosorbent assay as preferred screening method for nasopharyngeal carcinoma: a two-stage design with a preliminary performance study and a mass screening in southern China. Int J Cancer. 2012; 131:406-416.
7. Wang WY, Twu CW, Chen HH, Jan JS, Jiang RS, Chao JY, Liang KL, Chen KW, Wu CT and Lin JC. Plasma EBV DNA clearance rate as a novel prognostic marker for metastatic/recurrent nasopharyngeal carcinoma. Clin Cancer Res. 2010; 16:1016-1024.
8. Li HP, Peng CC, Chung IC, Huang MY, Huang ST, Chen CC, Chang KP, Hsu CL and Chang YS. Aberrantly hypermethylated Homeobox A2 derepresses metalloproteinase-9 through TBP and promotes invasion in Nasopharyngeal carcinoma. Oncotarget. 2013; 4:2154-2165. doi: 10.18632/oncotarget.1367.
9. Yip TT, Ngan RK, Fong AH and Law SC. Application of circulating plasma/serum EBV DNA in the clinical management of nasopharyngeal carcinoma. Oral Oncol. 2014; 50:527-538.
10. Gu AD, Mo HY, Xie YB, Peng RJ, Bei JX, Peng J, Li MY, Chen LZ, Feng QS, Jia WH and Zeng YX. Evaluation of a multianalyte profiling assay and an enzyme-linked immunosorbent assay for serological examination of Epstein-Barr virus-specific antibody responses in diagnosis of nasopharyngeal carcinoma. Clin Vaccine Immunol. 2008; 15:1684-1688.
11. Shao JY, Li YH, Gao HY, Wu QL, Cui NJ, Zhang L, Cheng G, Hu LF, Ernberg I and Zeng YX. Comparison of plasma Epstein-Barr virus (EBV) DNA levels and serum EBV immunoglobulin A/virus capsid antigen antibody titers in patients with nasopharyngeal carcinoma. Cancer. 2004; 100:1162-1170.
12. Chan KH, Gu YL, Ng F, Ng PS, Seto WH, Sham JS, Chua D, Wei W, Chen YL, Luk W, Zong YS and Ng MH. EBV specific antibody-based and DNA-based assays in serologic diagnosis of nasopharyngeal carcinoma. Int J Cancer. 2003; 105:706-709.
13. Cai L, Li J, Zhang X, Lu Y, Wang J, Lyu X, Chen Y, Liu J, Cai H, Wang Y and Li X. Gold nano-particles (AuNPs) carrying anti-EBV-miR-BART7-3p inhibit growth of EBV-positive nasopharyngeal carcinoma. Oncotarget. 2015; 6:7838-7850. doi: 10.18632/oncotarget.3046.
14. Kirschner AN, Omerovic J, Popov B, Longnecker R and Jardetzky TS. Soluble Epstein-Barr virus glycoproteins gH, gL, and gp42 form a 1:1:1 stable complex that acts like soluble gp42 in B-cell fusion but not in epithelial cell fusion. J Virol. 2006; 80:9444-9454.
15. Plate AE, Reimer JJ, Jardetzky TS and Longnecker R. Mapping regions of Epstein-Barr virus (EBV) glycoprotein B (gB) important for fusion function with gH/gL. Virology. 2011; 413:26-38.
16. Sashihara J, Burbelo PD, Savoldo B, Pierson TC and Cohen JI. Human antibody titers to Epstein-Barr Virus (EBV) gp350 correlate with neutralization of infectivity better than antibody titers to EBV gp42 using a rapid flow cytometry-based EBV neutralization assay. Virology. 2009; 391:249-256.
17. Leung SF, Tam JS, Chan AT, Zee B, Chan LY, Huang DP, Van Hasselt A, Johnson PJ and Lo YM. Improved accuracy of detection of nasopharyngeal carcinoma by combined application of circulating Epstein-Barr virus DNA and anti-Epstein-Barr viral capsid antigen IgA antibody. Clin Chem. 2004; 50:339-345.
18. Dardari R, Khyatti M, Benider A, Jouhadi H, Kahlain A, Cochet C, Mansouri A, El GB, Benslimane A and Joab I. Antibodies to the Epstein-Barr virus transactivator protein (ZEBRA) as a valuable biomarker in young patients with nasopharyngeal carcinoma. Int J Cancer. 2000; 86:71-75.
19. Fachiroh J, Stevens SJ, Haryana SM and Middeldorp JM. Combination of Epstein-Barr virus scaffold (BdRF1/VCA-p40) and small capsid protein (BFRF3/VCA-p18) into a single molecule for improved serodiagnosis of acute and malignant EBV-driven disease. J Virol Methods. 2010; 169:79-86.
20. Lin JC, Wang WY, Chen KY, Wei YH, Liang WM, Jan JS and Jiang RS. Quantification of plasma Epstein-Barr virus DNA in patients with advanced nasopharyngeal carcinoma. N Engl J Med. 2004; 350:2461-2470.
21. Ng WT, Yuen KT, Au KH, Chan OS and Lee AW. Staging of nasopharyngeal carcinoma—the past, the present and the future. Oral Oncol. 2014; 50:549-554.
22. Matsuura H, Kirschner AN, Longnecker R and Jardetzky TS. Crystal structure of the Epstein-Barr virus (EBV) glycoprotein H/glycoprotein L (gH/gL) complex. Proc Natl Acad Sci U S A. 2010; 107:22641-22646.
23. Khanna R, Sherritt M and Burrows SR. EBV structural antigens, gp350 and gp85, as targets for ex vivo virus-specific CTL during acute infectious mononucleosis: potential use of gp350/gp85 CTL epitopes for vaccine design. J Immunol. 1999; 162:3063-3069.
24. Urquiza M, Suarez J, Lopez R, Vega E, Patino H, Garcia J, Patarroyo MA, Guzman F and Patarroyo ME. Identifying gp85-regions involved in Epstein-Barr virus binding to B-lymphocytes. Biochem Biophys Res Commun. 2004; 319:221-229.
25. Yokoyama T, Ayabe S, Miyagi H, Sugano T, Otsu A, Sato H, Kageyama S, Fujii T and Shiraki K. Varicella-zoster virus gH:gL contains a structure reactive with the anti-human gamma chain of IgG near the glycosylation site. J Gen Virol. 2001; 82:331-334.
26. Herbst LH, Lemaire S, Ene AR, Heslin DJ, Ehrhart LM, Bagley DA, Klein PA and Lenz J. Use of baculovirus-expressed glycoprotein H in an enzyme-linked immunosorbent assay developed to assess exposure to chelonid fibropapillomatosis-associated herpesvirus and its relationship to the prevalence of fibropapillomatosis in sea turtles. Clin Vaccine Immunol. 2008; 15:843-851.
27. Xu J, Ahmad A, Blagdon M, D’Addario M, Jones JF, Dolcetti R, Vaccher E, Prasad U and Menezes J. The Epstein-Barr virus (EBV) major envelope glycoprotein gp350/220-specific antibody reactivities in the sera of patients with different EBV-associated diseases. Int J Cancer. 1998; 79:481-486.
28. Littler E, Baylis SA, Zeng Y, Conway MJ, Mackett M and Arrand JR. Diagnosis of nasopharyngeal carcinoma by means of recombinant Epstein-Barr virus proteins. Lancet. 1991; 337:685-689.
29. Gu AD, Xie YB, Mo HY, Jia WH, Li MY, Li M, Chen LZ, Feng QS, Liu Q, Qian CN and Zeng YX. Antibodies against Epstein-Barr virus gp78 antigen: a novel marker for serological diagnosis of nasopharyngeal carcinoma detected by xMAP technology. J Gen Virol. 2008; 89:1152-1158.
30. Gu AD, Lu LX, Xie YB, Chen LZ, Feng QS, Kang T, Jia WH and Zeng YX. Clinical values of multiple Epstein-Barr virus (EBV) serological biomarkers detected by xMAP technology. J Transl Med. 2009; 7:73.
31. Sanchez-Martinez D, Patton JL, Stewart JA and Pellett PE. Detection of Epstein-Barr virus-specific antibodies by means of baculovirus-expressed EBV gp125. J Virol Methods. 1995; 52:145-153.
32. Shanmugaratnam K and Sobin LH. The World Health Organization histological classification of tumours of the upper respiratory tract and ear. A commentary on the second edition. Cancer. 1993; 71:2689-2697.
33. Edge SB, Byrd DR, Compton CC, Fritz AG, Greene FL, Trotti A, editors. AJCC cancer staging manual (7th ed). NewYork: Springer; 2009.
34. Lo YM, Chan LY, Lo KW, Leung SF, Zhang J, Chan AT, Lee JC, Hjelm NM, Johnson PJ and Huang DP. Quantitative analysis of cell-free Epstein-Barr virus DNA in plasma of patients with nasopharyngeal carcinoma. Cancer Res. 1999; 59:1188-1191.
35. Shen Q, Fan J, Yang XR, Tan Y, Zhao W, Xu Y, Wang N, Niu Y, Wu Z, Zhou J, Qiu SJ, Shi YH, Yu B, et al. Serum DKK1 as a protein biomarker for the diagnosis of hepatocellular carcinoma: a large-scale, multicentre study. Lancet Oncol. 2012; 13:817-826.
36. Cao SM, Xu YJ, Lin GZ, Huang QH, Wei KR, Xie SH and Liu Q. Estimation of cancer burden in Guangdong Province, China in 2009. Chin J Cancer. 2015; 34:58.
37. Chien YC, Chen JY, Liu MY, Yang HI, Hsu MM, Chen CJ and Yang CS. Serologic markers of Epstein-Barr virus infection and nasopharyngeal carcinoma in Taiwanese men. N Engl J Med. 2001; 345:1877-1882.
38. Chan KC. Plasma Epstein-Barr virus DNA as a biomarker for nasopharyngeal carcinoma. Chin J Cancer. 2014; 33:598-603.