
Introduction
Most recurrent somatic mutations recently evidenced in myeloid malignancies, including myelodysplastic syndromes (MDS), myeloproliferative neoplasms (MPN), and acute myeloid leukemias (AML), affect genes implicated in the regulation of gene expression (RUNX1, TEL/ETV6, TP53, TET2, ASXL1, EZH2, IDH1/2, DNMT3A). In MDS, high throughput sequencing comparing DNA from bone marrow mononuclear cells to germ-line DNA, which identified around 10 acquired mutations per patient sample, suggested that alterations in the control of translation may be implicated in the pathogenesis of myeloid disorders [1, 2, 3]. Impaired protein translation had been identified previously in the erythroid lineage of patients with a 5q- syndrome, a particular MDS subtype characterized by the haplo-insufficiency of RPS14 gene encoding a ribosomal protein of the small ribosome subunit [4]. Exome sequencing of MDS samples has shown that altered translation could be related also to recurrent mutations in spliceosomal protein genes (SF3B1, SRSF2, ZRSR2, U2AF35) whose products control the mechanism of pre-messenger RNA (pre-mRNA) splicing. Similar analyses conducted in chronic lymphocytic leukemia (CLL), where DNA from tumor CD19+CD5+ lymphocytes and non-tumor cells were compared, also identified mutations affecting the control of splicing mechanisms [5,6] indicating that similar mutations could be observed in both myeloid and lymphoid neoplasms [7, 8]. In addition to myeloid and lymphoid neoplasms, splice gene mutations and amplifications were identified in solid tumors such as breast and lung cancers [9]. Altogether, these observations suggest that aberrations in genes of the spliceosome machinery could contribute to the onset of cancers [10, 11].
Spliceosome functioning
Gene expression in eukaryotic cells includes the processing of pre-mRNA into mature forms of mRNA through the splicing of introns and ligation of exons in the nucleus [for review, ref. 12]. Most of the human genes are processed to produce two or more transcripts by a mechanism of alternative splicing ie the alternative inclusion or exclusion of coding exons, or part of coding exons, in a cell type-specific manner. This mechanism generates a large diversity of mRNA species and is submitted to quality check by a “nuclear surveillance”. When 3’ processing/ polyadenylation is inefficient or compromised by gene mutations, then the nuclear exosome is recruited to degrade aberrantly spliced and read-through pre-mRNA [13, 14]. The spliceosomes are made up of multiple and large small nuclear ribonucleoprotein (RNP) complexes that catalyze the splicing reaction. The vast majority of introns (~99%) are spliced by a so-called “U2-dependent spliceosome” [15]. The major U2-dependent spliceosome contains more than 150 proteins and 5 small nuclear (sn) RNAs (U1, U2, U4/U6, and U5). Early steps include the recruitment of snRNP complexes to newly transcribed pre-mRNA for the recognition of 5’ and 3’ exon/intron junctions and later steps include the interaction between the 5’ and 3’ complexes to catalyse the excision of introns.
RNA splicing is initiated by the recognition of 5’ splice site by an U1 snRNP complex. The splicing factor 1 (SF1) complex, via its component SF3B1, binds to the branchpoint sequence located upstream of the 3’ end of the intron to protect the region before the splicing reaction. A complex that contains an U2 auxiliary factor (AF) 35/65 heterodimer, ZRSR2, and one of the serine-arginine (SR)-rich splicing factors, SRSF1 or SRSF2, is recruited to the polypyrimidine tract located between the branchpoint and the 3’ splice site. SRSF1 and SRSF2 play a role in preventing exon skipping, thus regulating alternative splicing. U2AF65 binds the polypyrimidine tract, while U2AF35, also known as U2AF1, interacts with the AG splice acceptor dinucleotide of the target intron at the 3’ splice site. ZRSR2 selectively binds to the 3’ splice site of U2 dependent pre-mRNA. Together with U2AF heterodimer, SF1 participates to the establishment of the E splicing complex. Then, the U2 snRNP complex, which contains the SF3b subcomplex made of SF3B1 and SF3A1 proteins, displaces SF1 to generate the A splicing complex. This step is followed by the incorporation of the U4/U6-U5 snRNP complexes. Lastly, the release of U1 and U4 snRNP changes the conformation of the complex for the spliceosome to become catalytically competent [16]. The E/A complexes are involved in the recognition of pre-mRNA during the very first steps of splicing, more precisely to the recognition of the polypyrimidine track and the acceptor splice site and also the exon splice enhancer (Figure 1).
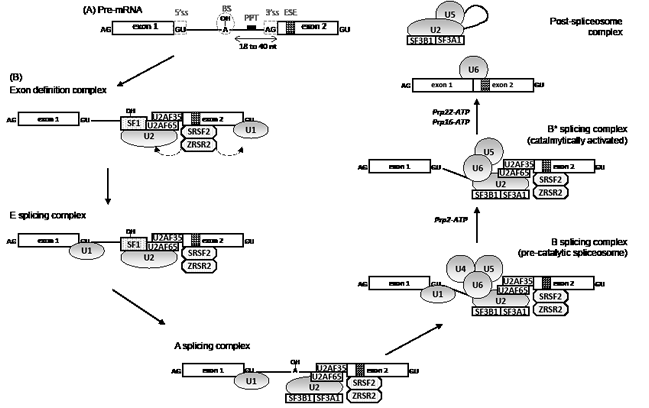
Figure 1: Pre-mRNA splicing mechanism by the U2-type spliceosome. (A) A pre-mRNA contains conserved sequences in intron and exon which are necessary for splicing. The 5’splicing site (ss) and the 3’ splicing site define the intron and participate in the assembly of the spliceosome together with the branch site (BS) and the polypyrimidine tract (PPT). Within exons, sequence like the exon splicing enhancer (ESE) also participates to the splicing process. (B) RNA splicing is initiated by the recognition of 5’ splice site by an U1 snRNP complex. The splicing factor 1 (SF1) binds to the branchpoint sequence located upstream of the 3’ end of the intron and a complex which contains an U2 auxiliary factor (AF) 35/65 heterodimer binds respectively to the polypyrimidine tract and the 3’splicing site. This complex is stabilized by ZRSR2 and one protein of the serine-arginine (SR)-rich domain family proteins, SRSF1 or SRSF2. A poorly understood mechanism leads to a switch from an exon-defined to intron-defined splicing complex which forms the E complex. Then, the U2 snRNP complex which contains the SF3b subcomplex, made of SF3B1 and SF3A1 proteins, displaces SF1 to generate the A splicing complex. This step is followed by the integration of the U4/U6-U5 snRNP complexe leading to the formation of the pre-catalytic spliceosome. Lastly, the conformation changes with the release of U1 and U4 snRNP for the spliceosome to become catalytically competent with the help of Prp2, Prp22 and Prp16 DExHD-box helicases. After catalysis, the spliceosome disassembles and is recycled.
Most pre-RNA splicing events occur co-transcriptionally. The recruitment of spliceosome complexes to actively transcribing genes is finely ordered starting with the association of U1 snRNP to a newly formed 5’ splice site followed by U2 and U5 snRNP when the intron is fully synthesized. The direct interaction between U1 snRNP and/or splicing regulatory proteins and the DNA polymerase II suggests an interconnection between transcription and mRNA splicing [17]. There is also a link between splicing and mRNA export from the nucleus and translation, which is mediated by SR family proteins. These proteins are recruited to pre-mRNA for splicing in a hyperphosphorylated state and become partially dephosphorylated as the splicing reaction proceeds. They also function as adaptors for spliced mRNA export when in hypophosphorylated state [18]. The SR protein, SF2, also enhances translation initiation through the recruitment of 40S ribosomal S6 kinase 1, which activates translation initiation when activated by the TOR signaling cascade.
Because introns often contain termination codons in frame with the upstream protein coding sequences, a leakage of unspliced pre-mRNA into the cytoplasm could occur. To prevent the production of aberrantly truncated proteins, mRNA species containing premature termination codon are degraded through the nonsense-mediated mRNA decay (NMD), a translation-dependent degradation pathway in the cytoplasm. This pathway is also a key regulator of pre-mRNA splicing factors cell content. When intracellular concentrations become too high, splicing factors regulate their own expression by targeting their mRNAs to NMD. Again, this establishes a link between splicing factors and NMD [19]. In addition, the SF3b subcomplex in the U2 snRNP is involved in the retention of aberrant pre-mRNA in the nucleus. Thus, splicing machinery components participate to the quality check allowing the translation of well-processed mature mRNA in the cytoplasm.
Recent studies have identified striking differences of alternative splicing between embryonic stem cells and differentiated cell populations [20, 21, 22]. For instance, an embryonic stem cell (ESC)-specific alternative splicing event changes the DNA-binding preference of the Forkhead family transcription factor FOXP1, stimulating the expression of transcription factor genes required for the maintenance of pluri-potency, and repressing the expression of genes driving ESC differentiation [22]. Thus, the splicing needs to be tightly regulated to avoid the deregulation of cell differentiation processes.
Consequences of spliceosome alterations
Most of the splice factors can be mutated in hematological malignancies or solid tumors, some of them being implicated in splicing or in alternative splicing, and also in some extra-splicing functions. Knockdown of these factors alters the splicing pattern of a subset of genes, and changes in their expression level have been linked also to the pathogenesis of cancer [9, 23]. For instance, the embryonic M2 isoform of the pyruvate kinase is re-expressed through alternative splicing in lung cancers, thus promoting the aerobic glycolysis in tumor cells, also known as the Warburg effect. The mechanism of this re-expression involves three heterogeneous nuclear RNP, the polypyrimidine tract binding protein PTB, hnRNPA1 and hnRNPA2, which bind to flanking sequences of exon 9 and repress the exclusion of exon 10 [24]. Expression of the three hnRNPs can be driven by the c-Myc oncogene that upregulates their transcription [25]. During c-Myc-driven cell transformation, the upregulation of splicing factor SRSF1 also results in aberrant alternative splicing events in cell cycle, apoptosis and signalling regulatory genes BCL2L11, BIN1, MNK2, S6K1, CCND1, RAC1, and RON [26, 27].
Given that splicing is often tightly coupled to transcription [28], alternative splicing might be affected by chromatin structure and histone modification, for instance through the direct recruitment of the splice factor MRG15/MORF4L1 to the H3K36me3 chromatin [29]. Conversely, SF3B1 interacts with the Polycomb repressive complex (PRC), and the lack of SF3B1 impairs PRC function, which may influence the chromatin structure and the transcription of HOX genes [30]. In addition, mutations affecting the nuclear degradation of non-coding RNA by exosome are likely involved in cell transformation. Inactivating mutations in the exonuclease gene DIS3, which encodes the catalytic subunit of the exosome [31] or in the exoribonuclease gene SEP1/KEM1, could lead to an accumulation of non-coding RNA species and interfere with transcription [32]. DIS3 mutations have already been reported in AML [33]. Changes targeting “splice genes” may therefore alter the maturation of pre-mRNA by affecting the spliceosome functions, and also the efficiency of transcription and the degradation of non-coding RNA [14]. This may lead to (i) degradation of unstable mRNA mimicking a loss of tumor suppressor gene or (ii) aberrant alternative splicing increasing the oncogenic potential of the defective proteins with a dominant negative effect. The role of alternative splicing in cell differentiation has been well established, and deregulation of these processes may also be involved in cell transformation [22]. Thus, incidence of splice gene mutations could depend on whether they occur in the hematopoietic stem cell or in more committed progenitors.
Frequence of splice gene mutations in MDS and other myeloid malignancies
Mutations in SF3B1 gene were initially identified by whole exome sequencing in 6 cases of 9 MDS, 8 of them being refractory anemia with ring sideroblasts, and a large scale mutational analysis confirmed the high frequency of SF3B1 mutations in MDS (20%) compared to other myeloid neoplasms (AML 5%; MPN 3%).[3] Yoshida and colleagues analyzed 29 MDS cases and identified multiple mutations of the RNA splicing machinery including SF3B1, U2AF35, SRSF2, ZRSR2, SF3A1 and PRPF40B in 16 cases [1]. Those mutations were frequent in MDS (45-87%), CMML (55%), and secondary AML (26%), but rare in de novo AML (7%), MPN (9%), primary myelofibrosis (6.5%), and pediatric myeloid neoplasms including JMML [34-41]. We reported splice gene mutations in 95/221 MDS patients with a frequence of 16, 11, 11 and 5% for SF3B1, SRSF2, ZRSR2 and U2AF35 mutations, respectively [34]. In CMML, mutations in SRSF2 gene, which are frequently associated to mutations in TET2 gene, are identified in ~50% of the patients whereas mutations in other splice genes are found in an additional 10% of these patients [42-43]. The frequence of SF3B1 mutations is equivalent in de novo and secondary AML while that of U2AF35 mutations is higher in secondary AML than in de novo diseases (Table 1).
Study |
Ref |
SF3B1 |
SRSF2 |
ZRSR2 |
U2AF35 / U2AF1 |
Prognosis |
MDS |
||||||
Yoshida et al, 2011 |
1 |
75.3% (55/73) RS patients |
5,50% |
1,40% |
0 |
|
1 |
6.5% (10/155) non RS patients |
11,60% |
7,70% |
11,60% |
|
|
Papaemmanuil et al, 2011 |
3 |
20.3% (72/354) |
NA |
NA |
NA |
Favorable |
Damm et al, 2012a |
48 |
14.7% (47/317) |
NA |
NA |
NA |
No |
Patnaik et al, 2011 |
44 |
49.5% (53/107) RS patients only |
NA |
NA |
NA |
Favorable |
Malcovati et al, 2011 |
45 |
29,4% (162/551) |
NA |
NA |
NA |
Favorable |
Visconte et al, 2012a |
41 |
68.8% (22/32) RS patients |
NA |
NA |
NA |
|
41 |
0% (0/24) non RS patients |
NA |
NA |
NA |
|
|
Thol et al, 2012 |
35 |
14,5% (28/193) |
12,1% (24/193) |
3,1% (6/193) |
7,3% (14/193) |
Poor |
Damm et al, 2012b |
34 |
16,4% (37/221) |
11,1% (25/221) |
11,1% (25/221) |
5,4% (12/221) |
Poor |
Bejar et al, 2012 |
36 |
22% (64/288) |
13% (36/288) |
NA |
16% (46/288) |
No |
Cui et al, 2012 |
47 |
53% (55/104) MDS-RS |
NA |
NA |
NA |
Favorable |
Jeromin et al, 2012 |
39 |
87,2% (41/47) MDS-RS |
NA |
NA |
NA |
|
Makishima et al, 2012 |
37 |
28,4% (25/88, RS and non RS) |
10,2% (9/88) |
NA |
9,0% (8/88) |
Poor (SRSF2, U2AF35) |
Visconte et al, 2012b |
46 |
39% (37/93), RARS: 68% (13/19) |
NA |
NA |
NA |
Favorable |
Graubert T et al, 2011 |
2 |
NA |
NA |
NA |
8,7% (13/150) |
Poor |
Qian et al, 2012 |
52 |
NA |
NA |
NA |
6,3% (6/96) |
No |
Wu et al, 2012 |
53 |
NA |
14,6% (34/233) |
NA |
NA |
Poor |
MPN |
||||||
Yoshida et al, 2011 |
1 |
0% (0/53) |
1,90% |
1,90% |
1,90% |
|
Papaemmanuil et al, 2011 |
3 |
ET: 3% (6/189) - PMF: 4% (6/136) - PV: 0/95 |
NA |
NA |
NA |
|
Zhang et al, 2012 |
38 |
PMF : 5,3% (2/38) |
2,6% (1/38) |
5,6% (2/38) |
2,6% (1/38) |
|
Lasho et al, 2012a |
49 |
PMF : 6.5% (10/155) |
NA |
NA |
NA |
No |
Lasho et al, 2012b |
55 |
PMF : 7% (12/187) |
17% (32/187) |
NA |
NA |
Poor |
CMML |
||||||
Yoshida et al, 2011 |
1 |
4.5% (4/88) |
28,40% |
8% |
8% |
|
Papaemmanuil et al, 2011 |
3 |
4.7% (5/106) |
NA |
NA |
NA |
|
Makishima et al, 2012 |
37 |
3,0% (2/66) |
21,2% (14/66) |
NA |
7,5% (5/66) |
|
Visconte et al, 2012a |
41 |
8,3% (5/60) |
NA |
NA |
NA |
|
Meggendorfer et al, 2012 |
42 |
NA |
47% (129/275) |
NA |
NA |
No |
Malcovati et al, 2011 |
45 |
6.5% (4/62) |
NA |
NA |
NA |
|
Abu Kar et al, 2012 |
54 |
6% (5/87) |
32% (28/87) |
NA |
13% (11/87) |
Poor |
JMML |
||||||
Hirabayashi et al, 2012 |
40 |
0 |
2/116 |
NA |
0 |
|
Abu Kar et al, 2012 |
54 |
0 |
0 |
NA |
0 |
|
AML |
||||||
Yoshida et al, 2011 |
1 |
4,8% (3/62) sAML |
6,5% (4/62) |
1,6% (1/62) |
9,7% (5/62) |
|
Yoshida et al, 2011 |
1 |
2,6% (7/151) nAML |
0,7% (1/151) |
0 |
1,3% (2/151) |
|
Papaemmanuil et al, 2011 |
3 |
5.3% (3/57) nAML |
NA |
NA |
NA |
|
Makishima et al, 2012 |
37 |
3,7% (2/54) sAML |
13% (7/54) |
NA |
9,2% (5/54) |
|
Makishima et al, 2012 |
37 |
7,2% (4/55) nAML |
0 |
NA |
10,9% (6/55) |
|
Zhang et al, 2012 |
38 |
3,7% (2/54) nAML |
5,6% (3/54) |
5,6% (3/54) |
1,9% (1/54) |
|
Zhang et al, 2012 |
38 |
8,6% (8/95) sAML |
23,7% (22/95) |
1,8% (2/95) |
5,7% (5/95) |
Poor |
Visconte et al, 2012a |
41 |
4,7% (2/44) nAML |
NA |
NA |
NA |
|
Visconte et al, 2012a |
41 |
5,9% (3/50) sAML |
NA |
NA |
NA |
|
Malcovati et al, 2011 |
45 |
5.3% (2/38) sAML |
NA |
NA |
NA |
|
Qian et al, 2012 |
52 |
NA |
NA |
NA |
2,5% (7/275) nAML |
|
CLL |
||||||
Wang et al, 2011 |
5 |
15% (14/91) |
NA |
NA |
NA |
Poor |
Quesada et al, 2011 |
6 |
9.7% (27/279) |
NA |
NA |
NA |
Poor |
Rossi et al, 2011 |
50 |
7.5% (27/360) |
NA |
NA |
NA |
Poor |
Oscier et al, 2012 |
51 |
17% (84/494) |
NA |
NA |
NA |
Poor |
This table summarizes the main studies establishing the rate (%) of somatic mutations of SF3B1, SRSF2, ZRSR2 and U2AF35/U2AF1 in haematological malignancies. The number of positives cases among the global cohort is indicated in parenthesis for each study. When available, the impact on the prognosis (poor, favourable or no impact) is indicated in the last column.
For MDS, authors may have distinguished patients with or without Ring Sideroblasts (RS).
In some cases, secondary AML (sAML) and de novo AML (nAML) have been studied separately. NA: Not Available, RS: Ring Sideroblasts, ET: Essential Thrombocytemia, PMF: Primary MyeloFibrosis, PV: Polycythemia Vera, MDS: myelodysplastic syndromes, CMML : chronic myelomonocytic luekemia, MPN: myeloproliferative neoplasms, JMML : juvenile myelomonocytic leukemia; CLL: chronic lymphocytic leukemia.
SF3B1 gene is located on 2q33.1, SRSF2 on 17q25.2, and U2AF35 on 21q22 chromosomes. With the exception of ZRSR2, which is located on the Xp22.2 chromosome, thus can be fully inactivated by heterozygous mutations in males, the other spliceosome mutations are missense mutations, often recurrently targeting a single amino acid. For example, 50% of SF3B1 mutations target the K700 amino-acid, most of the SRSF2 mutations are missense mutations or deletions at P95, while U2AF35 mutations in S34 and Q157 affect the two canonical zinc finger domains. In most cases, splice gene mutations are mutually exclusive. This type of mutational profile often indicates a gain of function that could possibly alter spliceosome functioning. As a result, the expression of many genes could be affected because of intron presence or the omission of exons in the mature transcript, or the deregulation of alternative splicing. In vitro, cells transfected with mutant U2AF35 present with an increase in exon skipping, thus confirming a gain of function, leading to a decrease in cell proliferation capacities and to a lower reconstitution capacity to competitive assay in mice [1,2]. Interestingly, whole mRNA deep sequencing comparing patients with a splice gene mutation to a patient without mutation revealed no genome-wide increase in intron retention, but an alternative splicing pattern in specific genes, including TET2 and RUNX1 [37].
Impact of splice gene mutations on MDS phenotype and clinical outcome
In MDS, mutations in SF3B1 are associated with lower hemoglobin levels than other splice gene mutations and cluster with the presence of ringed sideroblasts and with DNMT3A mutations [1, 3, 34-38, 44-48]. SF3B1 mutations are strongly predictive of the presence of ringed sideroblasts, whatever the WHO subtype [37, 45]. Sf3b1+/- mice also show an excess of ring sideroblasts in the bone marrow, without features of anemia, while the homozygous knockdown of Sf3b1 is lethal in embryos [30, 46].
Splice gene mutations correlate with distinct clinical phenotypes. For example, mutations in SRSF2 and U2AF35 are frequently detected in patients with advanced stages of MDS [34, 35]. SRSF2 mutated patients exhibit pronounced thrombocytopenias, while ZRSR2 mutated patients often display isolated neutropenias. The two mutations are more frequently associated with TET2 mutations than by chance [35]. SRSF2 mutations are more frequent in CMML (up to ~50%) than in MDS (~10%) [34, 43]. Also, patients with U2AF35 mutations, who have an increased prevalence of chromosome 20 deletions and ASXL1 mutations, have more frequently with advanced stages and sAML.
SF3B1 mutations have been associated with a good impact on overall survival and disease progression to AML in large series of patients, including a majority of refractory anemia with RS (RARS) [3, 44-47]. However, the good prognostic of RARS could induce a bias in these analyses and the subtype-independent prognostic impact of the mutation in MDS is still a matter of debate [34-37, 48]. By contrast, SF3B1 mutations have no impact on primary myelofibrosis and support a poor prognosis on overall survival in CLL [49, 50, 51].
The prognostic impact of U2AF35 mutations is also a controversial issue in myeloid malignancies [2, 34-37, 52], while SRSF2 mutations have a negative impact on MDS, MDS/MPN or MPN survival and MDS disease progression [34, 35, 37, 38, 53-55]. This is less clear in CMML in which the prognosis could depend on the combined mutations, i.e. the poor prognostic value of the mutations disappears when combined with a mutation in RUNX1 [43]. Thus, the frequent combination of SRSF2 mutations with ASXL1, TET2 and RUNX1 mutations could blur the picture [43]. For example, a multivariate analysis conducted in a cohort of 221 MDS patients whose mutational status was determined for 16 different genes, an inferior overall survival and a higher AML transformation rate was found for the genotype ZRSR2mut/TET2wt [34] (Table 1).
Targeting the splice
Several anticancer drugs are spliceosome inhibitors. Heterozygous splice gene mutations mainly result in a gain of function of the protein that has to be inhibited. Given that homozygous inactivation of splice genes is lethal in mice, spliceosome inhibitors have to be more toxic to heterozygous mutant cells than normal ones.
Such a therapeutic avenue could exist for FR901464, a natural bacterial product that was causes cell cycle arrest at G1 and G2/M phases in tumor cells. Recent evidence demonstrates that this compound and its methylated derivative spliceostatin A inhibits pre-mRNA splicing by non convalent binding to the SF3b complex in the U2 snRNP, leading to the leak of unspliced mRNA to be translated in the cytoplasm, and a partial accumulation of mRNA-polyA in the nucleus [56]. The stable derivative of FR901464, known as meayamycin, produces ring sideroblasts in normal erythroid cell cultures [46] suggesting that, if it efficiently reduces tumor cell growth, it may also alter residual normal erythropoiesis.
Another anti-tumor compound is the biflavonoid isogenkgetin, which prevents the stable recruitment of the U4-U6/U5 tri-snRNP to the pre-mRNA, resulting in the accumulation of pre-complex A [57]. Pladienolide is a natural macrolide with antitumor activity that binds the SF3b complex and was shown to inhibit mRNA splicing and cell proliferation in colon cancer cells but a drug resistance mechanism has already be identified, i.e. a mutation at Arg 1074 in the SF3B1 gene decreases the binding affinity of pladienolide to its target [58]. Additional in vitro studies on clonal and normal hematopoiesis and pre-clinical studies are ongoing to determine if these compounds deserve to be tested in MDS.
Some genes can produce both oncogenic and tumor suppressor proteins, based on the inclusion or exclusion of specific exons. In most cancers, alternative splicing of many genes is deregulated. Thus, another therapeutic approach that has been envisioned is the modulation of alternative splicing to prevent the generation of oncogenic forms of some proteins. Such an approach could apply to the alternative splicing of genes encoding pro-angiogenic (VEGF), signalling (STAT3), or pro-apoptotic (Bax) proteins. For example, borrelin-1, which was shown to be a potent inhibitor of angiogenesis, targets a spliceosome-associated protein, FBP21, leading to a modification of the ratio between VEGF isoforms in favour of the anti-angiogenic isoform [59]. The STAT3 transcription factor, which is constitutively activated in a number of human cancers, exists as a full-length STAT3a isofrom and a shorter STAT3b that lacks part of exon 23, due to an alternative splicing event. The short isoform lacks the transactivation domain but still dimerizes with STAT3a and binds to DNA without inducing transcription, thus may act in a dominant-negative manner to induce apoptosis and inhibit tumor growth. An antisense oligonucleotide (AON) targeting the acceptor splice site of STAT3a exon 23 induces a switch from STAT3a to STAT3b in cultured tumor cells and increases cell death, whereas in vivo injections of the morpholino counterparts into implanted tumors in mice results in tumor regression [60].
Conclusion and perspectives
Mutations targeting “splice genes” appear to be deleterious to cell growth. Such a detrimental mutation could hitchhike on a previous mutation to become advantageous, a scenario called “epistasis” [61]. An epistatic relation could exist, for example, between SRSF2 and TET2 genes, with SRSF2 mutation being detrimental in the context of a TET2-wildtype cell, and advantageous to the TET2-mutated clone, thus inducing a TET2/SRSF2 synthetic viability and generating a MDS or a CMML phenotype [34, 62]. According to this scenario, the prognostic significance of SRSF2 mutations could depend on the genetic background on which the mutation occurs. Very few studies have examined the specific targets of deregulated spliceosome and the mechanisms of action of these mutations, which will require the development of sophisticated models. Extensive clinical studies will also contribute to decipher the respective contribution of mutations in splice genes, epigenetic regulators, signaling molecules, and their combinations, leading to a refined molecular classification of myeloid malignancies and to the development of targeted therapies.
Acknowledgements:
Teams were supported by the Direction de la recherche clinique AP-HP (PHRC MDS-04), INCa, INSERM, association Laurette Fugain and labels from Ligue Nationale Contre le Cancer (LNCC). V. C. is the recipient of a grant from the Ministère de la Recherche et de l’enseignement supérieur. F. D. is supported by a grant from the Deutsche Krebshilfe. R.I. was the recipient of a poste d’accueil INSERM and a grant from INCa.
The authors have no conflict of interest to disclose
Reference
1. Yoshida K, Sanada M, Shiraishi Y, Nowak D, Nagata Y, Yamamoto R, Sato Y, Sato-Otsubo A, Kon A, Nagasaki M, Chalkidis G, Suzuki Y, Shiosaka M, Kawahata R, Yamaguchi T, Otsu M et al. Nature 2011; 478 : 64-69.
2. Graubert TA, Shen D, Ding L, Okeyo-Owuor T, Lunn CL, Shao J, Krysiak K, Harris CC, Koboldt DC, Larson DE, McLellan MD, Dooling DJ, Abbott RM, Fulton RS, Schmidt H, Kalicki-Veizer J, et al. Recurrent mutations in the U2AF1 splicing factor in myelodysplastic syndromes. Nat Genet 2011; 44 : 53-57.
3. Papaemmanuil E, Cazzola M, Boultwood J, Malcovati L, Vyas P, Bowen D, Pellagatti A, Wainscoat JS, Hellstrom-Lindberg E, Gambacorti-Passerini C, Godfrey AL, Rapado I, Cvejic A, Rance R, McGee C, Ellis P, et al. Somatic SF3B1 Mutation in Myelodysplasia with Ring Sideroblasts. N Engl J Med 2011; 365: 1384-1395.
4. Ebert BL, Pretz J, Bosco J, Chang CY, Tamayo P, Galili N, Raza A, Root DE, Attar E, Ellis SR, Golub TR. Identification of RPS14 as a 5q- syndrome gene by RNA interference screen. Nature. 2008; 451: 335-339.
5. Wang L, Lawrence MS, Wan Y, Stojanov P, Sougnez C, Stevenson K, Werner L, Sivachenko A, DeLuca DS, Zhang L, Zhang W, Vartanov AR, Fernandes SM, Goldstein NR, Folco EG, Cibulskis K, et al. SF3B1 and other novel cancer genes in chronic lymphocytic leukemia. N Engl J Med 2011; 365: 2497-2506.
6. Quesada V, Conde L, Villamor N, Ordóñez GR, Jares P, Bassaganyas L, Ramsay AJ, Beà S, Pinyol M, Martínez-Trillos A, López-Guerra M, Colomer D, Navarro A, Baumann T, Aymerich M, Rozman M, et al. Exome sequencing identifies recurrent mutations of the splicing factor SF3B1 gene in chronic lymphocytic leukemia. Nat Genet 2011; 44: 47-52.
7. Delhommeau F, Dupont S, Della Valle V, James C, Trannoy S, Massé A, Kosmider O, Le Couedic JP, Robert F, Alberdi A, Lécluse Y, Plo I, Dreyfus FJ, Marzac C, Casadevall N, Lacombe C, et al. Mutation in TET2 in myeloid cancers. N Engl J Med. 2009;360:2289-2301.
8. Quivoron C, Couronné L, Della Valle V, Lopez CK, Plo I, Wagner-Ballon O, Do Cruzeiro M, Delhommeau F, Arnulf B, Stern MH, Godley L, Opolon P, Tilly H, Solary E, Duffourd Y, Dessen P, et al. TET2 inactivation results in pleiotropic hematopoietic abnormalities in mouse and is a recurrent event during human lymphomagenesis. Cancer Cell 2011; 20: 25-38.
9. Gout S, Brambilla E, Boudria A, Drissi R, Lantuejoul S, Gazzeri S, Eymin B. Abnormal Expression of the Pre-mRNA Splicing Regulators SRSF1, SRSF2, SRPK1 and SRPK2 in Non Small Cell Lung Carcinoma. PLoS One. 2012; 7: e46539.
10. Imielinski M, Berger AH, Hammerman PS, Hernandez B, Pugh TJ, Hodis E, Cho J, Suh J, Capelletti M, Sivachenko A, Sougnez C, Auclair D, Lawrence MS, Stojanov P, Cibulskis K, Choi K, et al. Mapping the hallmarks of lung adenocarcinoma with massively parallel sequencing. Cell. 2012; 150:1107-1120.
11. Ellis MJ, Ding L, Shen D, Luo J, Suman VJ, Wallis JW, Van Tine BA, Hoog J, Goiffon RJ, Goldstein TC, Ng S, Lin L, Crowder R, Snider J, Ballman K, Weber J, et al. Whole-genome analysis informs breast cancer response to aromatase inhibition. Nature 2012; 486: 353-360.
12. Wahl MC, Will CL, Luhrmann R. The spliceosome: design principles of a dynamic RNP machine. Cell. 2009; 136: 701-718.
13. Moore M and Proudfoot NJ. Pre-mRNA processing reaches back to transcription and ahead to translation. Cell. 2009; 136: 688-700
14. Lemieux C, Marguerat S, Lafontaine J, Barbezier N, Bahler J, Bachand F. A Pre-mRNA degradation pathway that selectively targets intron-containing genes requires the nuclear poly(A)-binding protein. Mol Cell. 2011; 44: 108-119.
15. Wachtel C, Manley JL. Splicing of mRNA precursors: the role of RNAs and proteins in catalysis. Mol Biosyst. 2009; 5: 311-316.
16. Will CL, Luhrmann R. Spliceosome structure and function. Cold Spring Harb Perspect Biol 2011 ; 3(7).
17. Das R, Yu J, Zhang Z, Gygi MP, Krainer AR, Gygi SP, Reed R. SR proteins function in coupling RNAP II transcription to pre-mRNA splicing. Mol Cell. 2007; 26: 867-881.
18. Huang Y, Steitz JA. SRprises along a messenger’s journey. Mol Cell. 2005; 17: 613-615.
19. McGlincy NJ, Smith CW. Alternative splicing resulting in nonsense-mediated mRNA decay: what is the meaning of nonsense? Trends Biochem Sci. 2008; 33: 385-393.
20. Atlasi Y, Mowla SJ, Ziaee SA, Gokhale PJ, Andrews PW. OCT4 spliced variants are differentially expressed in human pluripotent and nonpluripotent cells. Stem Cells. 2008; 26: 3068-3074.
21. Rao S, Zhen S, Roumiantsev S, McDonald LT, Yuan GC, Orkin SH. Differential roles of Sall4 isoforms in embryonic stem cell pluripotency. Mol Cell Biol. 2010; 30: 5364-5380.
22. Gabut M, Samavarchi-Tehrani P, Wang X, Slobodeniuc V, O’Hanlon D, Sung HK, Alvarez M, Talukder S, Pan Q, Mazzoni EO, Nedelec S, Wichterle H, Woltjen K, Hughes TR, Zandstra PW, Nagy A, et al. An alternative splicing switch regulates embryonic stem cell pluripotency and reprogramming. Cell. 2011; 147: 132-146.
23. Pal S, Gupta R, Davuluri RV. Alternative transcription and alternative splicing in cancer. Pharmacol Ther. 2012; 136: 283-294.
24. Christofk HR, Vander Heiden MG, Harris MH, Ramanathan A, Gerszten RE, Wei R, Fleming MD, Schreiber SL, Cantley LC. The M2 splice isoform of pyruvate kinase is important for cancer metabolism and tumour growth. Nature. 2008; 452: 230-233.
25. David CJ, Chen M, Assanah M, Canoll P, Manley JL. HnRNP proteins controlled by c-Myc deregulate pyruvate kinase mRNA splicing in cancer. Nature. 2010; 463: 364-368.
26. Das S, Anczuków O, Akerman M, Krainer AR. Oncogenic splicing factor SRSF1 is a critical transcriptional target of MYC. Cell Rep. 2012; 1: 110-117.
27. Anczuków O, Rosenberg AZ, Akerman M, Das S, Zhan L, Karni R, Muthuswamy SK, Krainer AR. The splicing factor SRSF1 regulates apoptosis and proliferation to promote mammary epithelial cell transformation. Nat Struct Mol Biol. 2012; 19: 220-228.
28. Alexander RD, Innocente SA, Barrass JD, Beggs JD. Splicing-dependent RNA polymerase pausing in yeast. Mol Cell. 2010; 40: 582-593.
29. Luco RF, Pan Q, Tominaga K, Blencowe BJ, Pereira-Smith OM, Misteli T. Regulation of alternative splicing by histone modifications. Science. 2010; 327: 996-1000.
30. Isono K, Mizutani-Koseki Y, Komori T, Schmidt-Zachmann MS, Koseki H. Mammalian polycomb-mediated repression of Hox genes requires the essential spliceosomal protein Sf3b1. Genes Dev. 2005; 19: 536-541.
31. Schaeffer D, Tsanova B, Barbas A, Reis FP, Dastidar EG, Sanchez-Rotunno M, Arraiano CM, van Hoof A. The exosome contains domains with specific endoribonuclease, exoribonuclease and cytoplasmic mRNA decay activities. Nat Struct Mol Biol. 2009; 16: 56-62.
32. van Dijk EL, Chen CL, d’Aubenton-Carafa Y, Gourvennec S, Kwapisz M, Roche V, Bertrand C, Silvain M, Legoix-Né P, Loeillet S, Nicolas A, Thermes C, Morillon A. XUTs are a class of Xrn1-sensitive antisense regulatory non-coding RNA in yeast. Nature. 2011; 475: 114-117.
33. Ding L, Ley TJ, Larson DE, Miller CA, Koboldt DC, Welch JS, Ritchey JK, Young MA, Lamprecht T, McLellan MD, McMichael JF, Wallis JW, Lu C, Shen D, Harris CC, Dooling DJ et al. Clonal evolution in relapsed acute myeloid leukaemia revealed by whole-genome sequencing. Nature. 2012; 481: 506-510.
34. Damm F, Kosmider O, Gelsi-Boyer V, Renneville A, Carbuccia N, Hidalgo-Curtis C, Della Valle V, Couronné L, Scourzic L, Chesnais V, Guerci-Bresler A, Slama B, Beyne-Rauzy O, Schmidt-Tanguy A, Stamatoullas-Bastard A, Dreyfus F et al, Mutations affecting mRNA splicing define distinct clinical phenotypes and correlate with patient outcome in myelodysplastic syndromes. Blood. 2012; 119: 3211-3218.
35. Thol F, Kade S, Schlarmann C, Löffeld P, Morgan M, Krauter J, Wlodarski MW, Kölking B, Wichmann M, Görlich K, Göhring G, Bug G, Ottmann O, Niemeyer CM, Hofmann WK, Schlegelberger B, et al. Frequency and prognostic impact of mutations in SRSF2, U2AF1, and ZRSR2 in patients with myelodysplastic syndromes. Blood. 2012; 119: 3578-3584
36. Bejar R, Stevenson KE, Caughey BA, Abdel-Wahab O, Steensma DP, Galili N, Raza A, Kantarjian H, Levine RL, Neuberg D, Garcia-Manero G, Ebert BL. Validation of a prognostic model and the impact of mutations in patients with lower-risk myelodysplastic syndromes. J Clin Oncol. 2012; 30: 3376-3382.
37. Makishima H, Visconte V, Sakaguchi H, Jankowska AM, Abu Kar S, Jerez A, Przychodzen B, Bupathi M, Guinta K, Afable MG, Sekeres MA, Padgett RA, Tiu RV, Maciejewski JP. Mutations in the spliceosome machinery, a novel and ubiquitous pathway in leukemogenesis. Blood. 2012; 119: 3203-3210.
38. Zhang SJ, Rampal R, Manshouri T, Patel J, Mensah N, Kayserian A, Hricik T, Heguy A, Hedvat C, Gönen M, Kantarjian H, Levine RL, Abdel-Wahab O, Verstovsek S. Genetic analysis of patients with leukemic transformation of myeloproliferative neoplasms shows recurrent SRSF2 mutations that are associated with adverse outcome. Blood. 2012; 119: 4480-4485.
39. Jeromin S, Haferlach T, Grossmann V, Alpermann T, Kowarsch A, Haferlach C, Kern W, Schnittger S. High frequencies of SF3B1 and JAK2 mutations in refractory anemia with ring sideroblasts associated with marked thrombocytosis strengthen the assignment to the category of myelodysplastic/myeloproliferative neoplasms. Haematologica. 2012 Aug 28.
40. Hirabayashi S, Flotho C, Moetter J, Heuser M, Hasle H, Gruhn B, Klingebiel T, Thol F, Schlegelberger B, Baumann I, Strahm B, Stary J, Locatelli F, Zecca M, Bergstraesser E, Dworzak M et al. Spliceosomal gene aberrations are rare, coexist with oncogenic mutations, and are unlikely to exert a driver effect in childhood MDS and JMML. Blood. 2012 Mar 15; 119: e96-99.
41. Visconte V, Makishima H, Jankowska A, Szpurka H, Traina F, Jerez A, O‘Keefe C, Rogers HJ, Sekeres MA, Maciejewski JP, Tiu RV. SF3B1, a splicing factor is frequently mutated in refractory anemia with ring sideroblasts. Leukemia. 2012; 26: 542-545.
42. Meggendorfer M, Roller A, Haferlach T, Eder C, Dicker F, Grossmann V, Kohlmann A, Alpermann T, Yoshida K, Ogawa S, Koeffler HP, Kern W, Haferlach C, Schnittger S. SRSF2 mutations in 275 cases with chronic myelomonocytic leukemia (CMML). Blood. 2012; 120: 3080-3088.
43. Itzykson R, Kosmider O, Renneville A, Gelsi-Boyer V, Meggendorfer M, Morabito M, Berthon C, Ades L, Fenaux P, Beyne-Rauzy O, Vey N, Braun T, Haferlach T, Dreyfus F, Cross N, Preudhomme C, et al. A prognostic score including gene mutations in chronic myelomonocytic leukemia. J Clin Oncol 2012, in revision.
44. Patnaik MM, Lasho TL, Hodnefield JM, Knudson RA, Ketterling RP, Garcia-Manero G, Steensma DP, Pardanani A, Hanson CA, Tefferi A. SF3B1 mutations are prevalent in myelodysplastic syndromes with ring sideroblasts but do not hold independent prognostic value. Blood. 2012; 119: 569-572.
45. Malcovati L, Papaemmanuil E, Bowen DT, Boultwood J, Della Porta MG, Pascutto C, Travaglino E, Groves MJ, Godfrey AL, Ambaglio I, Gallì A, Da Vià MC, Conte S, Tauro S, Keenan N, Hyslop A et al. Clinical significance of SF3B1 mutations in myelodysplastic syndromes and myelodysplastic/myeloproliferative neoplasms. Blood. 2011; 118: 6239-6246.
46. Visconte V, Rogers HJ, Singh J, Barnard J, Bupathi M, Traina F, McMahon J, Makishima H, Szpurka H, Jankowska A, Jerez A, Sekeres MA, Saunthararajah Y, Advani AS, Copelan E, Koseki H et al. SF3B1 haploinsufficiency leads to formation of ring sideroblasts in myelodysplastic syndromes. Blood. 2012; 120: 3173-3186.
47. Cui R, Gale RP, Xu Z, Qin T, Fang L, Zhang H, Pan L, Zhang Y, Xiao Z. Clinical importance of SF3B1 mutations in Chinese with myelodysplastic syndromes with ring sideroblasts. Leuk Res. 2012; 36: 1428-1433.
48. Damm F, Thol F, Kosmider O, Kade S, Löffeld P, Dreyfus F, Stamatoullas-Bastard A, Tanguy-Schmidt A, Beyne-Rauzy O, de Botton S, Guerci-Bresler A, Göhring G, Schlegelberger B, Ganser A, Bernard OA, Fontenay M et al. SF3B1 mutations in myelodysplastic syndromes: clinical associations and prognostic implications. Leukemia. 2012; 26: 1137-1140.
49. Lasho TL, Finke CM, Hanson CA, Jimma T, Knudson RA, Ketterling RP, Pardanani A, Tefferi A. SF3B1 mutations in primary myelofibrosis: clinical, histopathology and genetic correlates among 155 patients. Leukemia. 2012; 26: 1135-1137.
50. Rossi D, Bruscaggin A, Spina V, Rasi S, Khiabanian H, Messina M, Fangazio M, Vaisitti T, Monti S, Chiaretti S, Guarini A, Del Giudice I, Cerri M, Cresta S, Deambrogi C, Gargiulo E et al. Mutations of the SF3B1 splicing factor in chronic lymphocytic leukemia: association with progression and fludarabine-refractoriness. Blood. 2011; 118: 6904-6908.
51. Oscier DG, Rose-Zerilli MJ, Winkelmann N, Gonzalez de Castro D, Gomez B, Forster J, Parker H, Parker A, Gardiner A, Collins A, Else M, Cross NC, Catovsky D, Strefford JC. The clinical significance of NOTCH1 and SF3B1 mutations in the UK LRF CLL4 trial. Blood 2012 Oct 18.
52. Qian J, Yao DM, Lin J, Qian W, Wang CZ, Chai HY, Yang J, Li Y, Deng ZQ, Ma JC, Chen XX. U2AF1 Mutations in Chinese Patients with Acute Myeloid Leukemia and Myelodysplastic Syndrome. PLoS One. 2012; 7: e45760.
53. Wu SJ, Kuo YY, Hou HA, Li LY, Tseng MH, Huang CF, Lee FY, Liu MC, Liu CW, Lin CT, Chen CY, Chou WC, Yao M, Huang SY, Ko BS, Tang JL et al. The clinical implication of SRSF2 mutation in patients with myelodysplastic syndrome and its stability during disease evolution. Blood. 2012; 120: 3106-3111.
54. Abu Kar S, Jankowska AM, Makishima H, Visconte V, Jerez A, Sugimoto Y, Muramatsu H, Traina F, Afable M, Guinta K, Tiu RV, Przychodzen B, Sakaguchi H, Kojima S, Sekeres MA, List AF et al. Spliceosomal gene mutations are frequent events in the diverse mutational spectrum of chronic myelomonocytic leukemia but largely absent in juvenile myelomonocytic leukemia. Haematologica. 2012 Jul 6.
55. Lasho TL, Jimma T, Finke CM, Patnaik M, Hanson CA, Ketterling RP, Pardanani A, Tefferi A. SRSF2 mutations in primary myelofibrosis: significant clustering with IDH mutations and independent association with inferior overall and leukemia-free survival. Blood. 2012 Sep 11.
56. Kaida D, Motoyoshi H, Tashiro E, Nojima T, Hagiwara M, Ishigami K, Watanabe H, Kitahara T, Yoshida T, Nakajima H, Tani T, Horinouchi S, Yoshida M. Spliceostatin A targets SF3b and inhibits both splicing and nuclear retention of pre-mRNA. Nat Chem Biol. 2007; 3: 576-583.
57. O’Brien K, Matlin AJ, Lowell AM, Moore MJ. The biflavonoid isoginkgetin is a general inhibitor of Pre-mRNA splicing. J Biol Chem. 2008; 283: 33147-33154.
58. Yokoi A, Kotake Y, Takahashi K, Kadowaki T, Matsumoto Y, Minoshima Y, Sugi NH, Sagane K, Hamaguchi M, Iwata M, Mizui Y. Biological validation that SF3b is a target of the antitumor macrolide pladienolide. FEBS J 2011; 278: 4870-4880.
59. Woolard J, Vousden W, Moss SJ, Krishnakumar A, Gammons MV, Nowak DG, Dixon N, Micklefield J, Spannhoff A, Bedford MT, Gregory MA, Martin CJ, Leadlay PF, Zhang MQ, Harper SJ, Bates DO, Wilkinson B. Borrelidin modulates the alternative splicing of VEGF in favour of anti-angiogenic isoforms. Chem Sci. 2011; 2011: 273-278.
60. Zammarchi F, de Stanchina E, Bournazou E, Supakorndej T, Martires K, Riedel E, Corben AD, Bromberg JF, Cartegni L. Antitumorigenic potential of STAT3 alternative splicing modulation. Proc. Natl. Acad. Sci. USA 201; 108: 17779–17784.
61. Ashworth A, Lord CJ, Reis-Filho JS. Genetic interactions in cancer progression and treatment. Cell. Cell. 2011;145:30-38.
62. Itzykson R, Kosmider O, Renneville A, Morabito M, Preudhomme C, Berthon C, Ades L, Fenaux P, Platzbecker U, Gagey O, Rameau P, Meurice G, Orear C, Delhommeau F, Bernard OA, Fontenay M, et al. Clonal architecture of chronic myelomonocytic leukemias. Blood 2012, in press.