
INTRODUCTION
Telomerase reverse transcriptase (TERT) is the catalytic subunit of telomerase [1], which is an essential enzyme for elongating telomeres at the end of chromosomes [2]. TERT expression levels are highly associated with cancer risk in various human cancers [1–3]. Cancer risk is associated with common single-nucleotide polymorphisms (SNPs), including the TERT gene variant, rs2853669 (-245T > C) [3]. Although rs2853669 increases lung cancer risk [4], it contributes to a lower breast cancer risk [3]. In studies of hepatocellular carcinoma (HCC), which accounts for more than 85% of liver cancers [5], no evidence has been reported on the significant association between the rs2853669 variant and HCC risk [6].
Telomerase activation is responsible for circumventing cellular senescence or cell death caused by telomere shortening in cells [7–9]. TERT expression, which is necessary for telomerase activity, is greater in various human tumors than in normal organs [2]. Recently, -124C > T and -146C > T somatic mutations at the TERT promoter were discovered in melanoma; a reporter assay showed that these mutations increased TERT transcription activity by creating a binding motif for transcription factor ETS2 in multiple cell lines [10–12]. These -124C > T and -146C > T somatic mutations are present in various human tumors, including HCC [1]. However, a number of previous reports disagree on whether TERT promoter mutations are responsible for elevated TERT expression or low patient survival rate in a variety of cancers, including, but not limited to, cutaneous melanoma [13, 14], thyroid cancer [15], bladder cancer [16–18], and HCC [19, 20]. Although TERT expression is greater in cirrhotic preneoplastic lesions (a HCC precursor) with the TERT promoter mutations compared with that of lesions that do not include the mutations [19], it is yet unclear whether somatic mutations at the TERT promoter affect the TERT expression levels or patient survival rates in HCC [19, 20].
Several lines of research suggest that rs2853669 suppresses the TERT promoter mutation-mediated TERT expression regulation and mortality or recurrence rates for bladder cancer [17], gliomas [21, 22], and renal cell cancer [23]; however, the mechanism by which TERT transcription is regulated remains unknown. So far, no studies have considered effect of rs2853669 combined with somatic mutations at the TERT promoter on liver cancer. Here, we aim to discover a novel role for the rs2853669 variant and a mechanism for regulating rs2853669 variant-dependent TERT promoter activity in liver cancer.
RESULTS
The combination of the rs2853669 variant and the TERT promoter mutation increases mortality and cancer recurrence rates in HCC patients
To determine the combined effect of the rs2853669 (-245T > C) and TERT promoter mutation (-124C > T or -146C > T) on liver cancer survival rates, we first analyzed the overall survival rates of a Korean HCC patient cohort (SMH cohort, n = 93; Figure 1A, 1B). The combination was correlated with a low overall survival rate (Log-rank test, P = 0.0055; Figure 1B) and a high significant risk of HCC-related death, as evidenced by the hazard ratio of 5.259 (95% CI = 1.42–19.48, P = 0.013; Supplementary Table 1). We did not observe an association between the overall survival rate and the TERT promoter mutation in HCC patients lacking rs2853669 (Figure 1B). Moreover, no significant difference in the survival rates was observed among the HCC patients with or without the TERT promoter mutation (Figure 1B), which is consistent with previous reports [19, 20]. Therefore, the mutation alone does not lead to high mortality rates in HCC patients while we concluded that the TERT promoter mutation and rs2853669, when in combination, does.
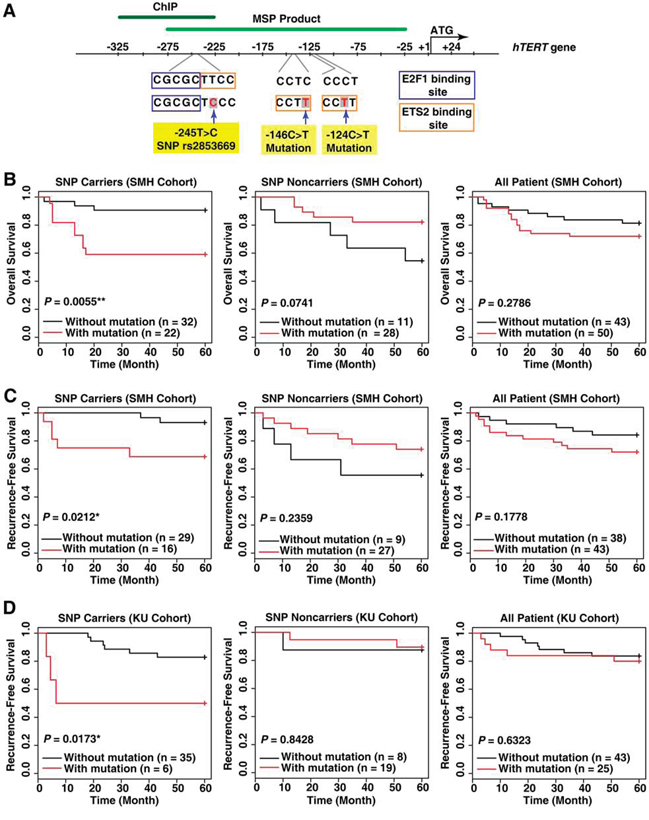
Figure 1: The variant rs2853669 at the TERT promoter is associated with a high risk of death and cancer recurrence in hepatocellular carcinoma (HCC) patients with a TERT promoter mutation. A. A representative map describing the SNP rs2853669 (-245T > C), -146C > T mutation, and -124C > T mutation in the human TERT gene. ChIP, Chromatin immunoprecipitation; MSP product, methylation specific PCR product. B. Kaplan–Meier analysis of differences in overall survival and C, D. recurrence-free survival based on the presence of the rs2853669 variant and TERT promoter mutation status. HCC patients included the SMH cohort (B, C) and KU cohort (D) *P < 0.05; **P < 0.01.
Next, we examined whether rs2853669 and the TERT promoter mutation combination influenced the risk of HCC recurrence in two independent Korean HCC patient cohorts (n = 93 for the SMH cohort; n = 72 for the KU cohort). The data show that rs2853669 is associated with poor recurrence-free survival rates and a significant risk of HCC recurrence in patients with the TERT promoter mutation (Log-rank test, P = 0.02119; hazard ratio = 5.5611, 95% CI = 1.076–28.75, P = 0.0406 for the SMH cohort; and Log-rank test, P = 0.0173; hazard ratio = 4.6639, 95% CI = 1.157–18.8, P = 0.0304 for the KU cohort; Figure 1C, 1D and Supplementary Table 1). In accordance with the results of the overall survival analysis (Figure 1B), analysis of the recurrence-free survival rates revealed that in patients without the rs2853669 or in all patients (with and without the SNP), there was no significant difference in the recurrence rate in response to the TERT promoter mutation (Figure 1C, 1D). Furthermore, both the overall survival and recurrence-free survival rates did not differ significantly between the patients with and without the rs2853669 alone (Supplementary Figure 1). Overall, only the combination of the rs2853669 and the TERT promoter mutation contributed to a high risk of HCC recurrence.
The combination of the rs2853669 variant and the TERT promoter mutation increases TERT transcription activity in HCC cell lines and HCC tumors
The luciferase promoter activity of the TERT reporter vector with rs2853669 (-245T > C) was increased in all four HCC cell lines examined (t-test, P < 0.001 for all cell lines; Figure 2A). Furthermore, the luciferase promoter activity of the TERT reporter vector with both rs2853669 and -124C > T was significantly greater than that of vectors with the -124C > T mutation only (t-test, P = 0.002 for Huh7, P = 0.012 for HepG2, P < 0.001 for Hep3B, and P < 0.001 for SNU-449; Figure 2A), which indicates a marked impact by the combination on TERT expression up-regulation.
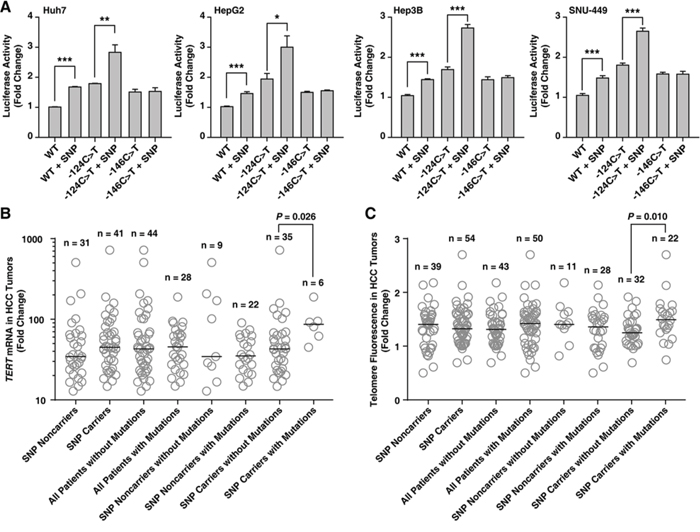
Figure 2: The variant rs2853669 at the TERT promoter is associated with an increased TERT promoter activity in HCC cell lines, and a high TERT mRNA expression level and long telomere lengths in HCC tumors. A–C. Quantification of TERT promoter activity in hepatocellular carcinoma (HCC) cell lines (A), TERT expression (B) and telomere fluorescence levels (C) in HCC tumor tissues based on the presence of the variant rs2853669 (single-nucleotide polymorphism (SNP)) only, the mutation only (-124C > T or -146C > T), and a combination of both (-124C > T + SNP or -146C > T + SNP). The data in A are presented as the means ± SEM (n = 4). The horizontal bar in B and C show the median. *P < 0.05, **P < 0.01, and ***P < 0.001.
Lastly, we examined whether the TERT mRNA expression level positively associate with the rs2853669 and the mutation combination. The TERT mRNA expression level was greater in HCC tumors with the combination than in tumors with rs2853669 only (Mann–Whitney test, P = 0.026; Figure 2B). Furthermore, HCC tumors with the combination had longer telomeres, which correlates with high TERT mRNA levels in HCC [24], than tumors with the rs2853669 polymorphism alone (Mann–Whitney test, P = 0.010; Figure 2C and Supplementary Figure 2). These data show rs2853669 variant and the TERT promoter mutation combination is positively associated with HCC tumor TERT mRNA expression levels and telomere lengths.
Inhibiting E2F1 binding to the TERT promoter increases TERT transcription levels in Huh7 cells without the rs2853669 variant
We further investigated the mechanism underlying the role of the rs2853669 in the TERT transcription activation as observed in the HCC tumors with the rs2853669 variant and the mutation combination. The rs2853669 variant site is close to (2 bp downstream) the binding site of E2F transcription factor 1 (E2F1) (Figure 1A). Therefore, we evaluated whether the rs2853669 variant can disrupt a preexisting E2F1 binding site at the TERT promoter and inhibit the E2F1 function as a TERT transcription repressor. First, we tested whether the E2F1 protein was involved in the lower TERT gene expression levels in Huh7 cells, which are HCC cells without the rs2853669 variant (Supplementary Figure 3). As a result, the E2F1 wild-type (WT) overexpression decreased the TERT mRNA and TERT protein expression levels as well as the TERT gene promoter activity in Huh7 cells (Figure 3A). In contrast, E2F1-Eco132 ectopic expression, which is due to a dominant-negative E2F1 mutant [25], increased the TERT mRNA, protein expression levels, and TERT promoter activity (Figure 3A), suggesting that E2F1 is a TERT transcriptional repressor in Huh7 cells.
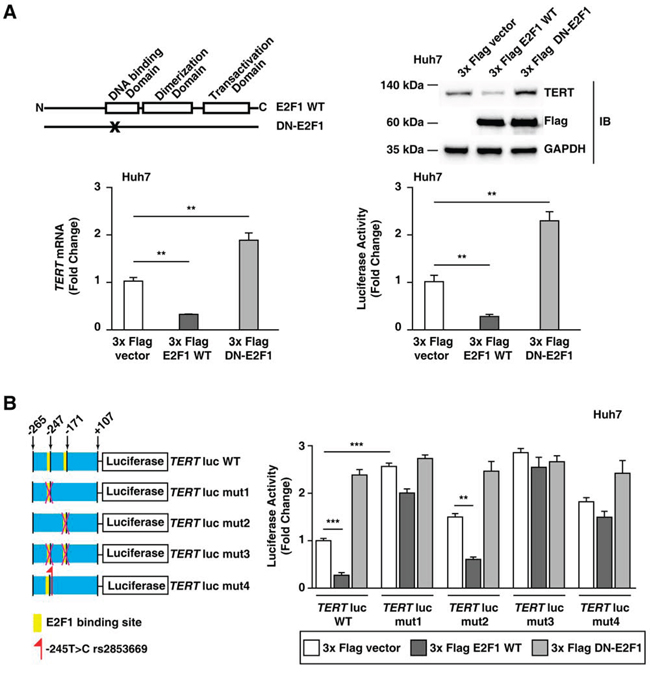
Figure 3: E2F1 is a TERT transcriptional repressor in Huh7 cells without rs2853669. A. Domain structure of wild-type E2F1 (E2F1 WT) and dominant-negative E2F1 (DN-E2F1); the immunoblot assay, qPCR, and the luciferase assay using a 3x Flag pCMV-10 empty vector, 3x Flag E2F1, and 3x Flag DN-E2F1. DN-E2F1 is mutated in the DNA binding domain. B. Luciferase assay using the WT TERT promoter (TERT-WT-luc), TERT promoter with a mutated E2F1-binding site (-247 bp upstream of ATG) (TERT-mut1-luc), TERT promoter with a mutated E2F1-binding site (-171 bp upstream of ATG) (TERT-mut2-luc), TERT promoter with mutated E2F1-binding sites (-247 bp upstream and -171 bp upstream of ATG) (TERT-mut3-luc), and TERT promoter with rs2853669 (-245 bp upstream of ATG) (TERT-mut4-luc). The data are shown as the mean ± SEM, **P < 0.01, *** P < 0.005.
We confirmed whether the effect of the rs2853669 variant (-245T > C) on TERT promoter activity was associated with the function of E2F1 as a transcriptional repressor. A previous study showed that the two E2F1 binding sites at -247 bp and -171 bp relative to ATG can regulate TERT promoter activity [26]. As demonstrated by the luciferase assay, the E2F1-binding site mutation at -247 bp relative to ATG (TERT-mut1-luc) compared to the E2F1-binding site mutation at -171 bp relative to ATG (TERT-mut2-luc) had a greater impact on the TERT promoter activity (Figure 3B). In addition, TERT-mut2-luc luciferase activity decreased after E2F1 overexpression (P < 0.01); however, TERT-mut1-luc luciferase activity was not affected by E2F1 overexpression (Figure 3B). Moreover, a subsequent luciferase assay demonstrated that the increased E2F1 level did not influence the luciferase activity of the TERT promoter with the rs2853669 variant (TERT-mut4-luc), similar to that demonstrated for the TERT promoter with a mutation at the E2F1-binding site (TERT-mut1-luc) (Figure 3B). These results suggested that rs2853669 inhibited E2F1 binding to the TERT promoter. Taken together, the data suggest that the rs2853669 variant induces TERT transcription levels by blocking E2F1 binding to its promoter.
DNA methyltransferase 1 and histone deacetylase 1 are involved in E2F1-mediated down-regulation of TERT transcription levels in the absence of the rs2853669 variant
E2F1 represses gene transcription levels by recruiting DNA methyltransferase 1 (DNMT1) [27]. Because DNMT1 acetylation often induces DNMT1 degradation, histone deacetylase 1 (HDAC1) stabilizes DNMT1 by interacting with DNMT1 [27, 28]. Immunostaining analyses revealed that E2F1 can interact with DNMT1, and the co-localization of DNMT1 and E2F1 was clearly observed in Huh7 cell nuclei (Figure 4A). In addition, co-immunoprecipitation analyses further demonstrated that both DNMT1 and HDAC1 [27, 28] interact with E2F1 (Figure 4B). These data suggest that the DNMT1, HDAC1, and E2F1 work in concert to facilitate TERT transcription level repression.
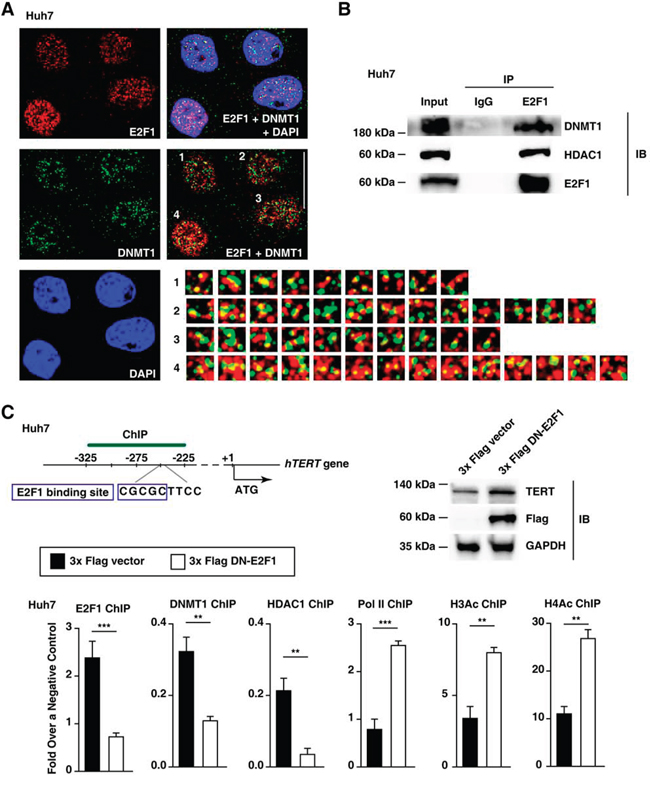
Figure 4: Epigenetic changes in the TERT promoter are involved in increased TERT transcription levels in Huh7 cells. A. Immunofluorescence staining of E2F1 (red), DNMT1 (green), and DAPI (blue) in Huh7 cell lines. Scale bar, 20 μm. The number-labeled boxes indicate the areas of protein co-localization (yellow). DAPI, 4′,6-diamidino-2-phenylindole. B. Huh7 cell nuclear extracts or immunoprecipitated products (IP) generated using control IgG and E2F1 antibodies were subjected to immunoblot analysis (IB) using the antibodies indicated on the right. C. Immunoblot assay and ChIP experiments using the TERT promoter from each 3x Flag empty vector- or 3x Flag DN-E2F1-ectopic expressed Huh7 cell line. Pol II, RNA polymerase II; H3Ac and H4Ac, acetylated histones H3 and H4. The data are shown as the mean ± SEM, **P < 0.01, *** P < 0.005.
Our ChIP data demonstrated that reducing endogenous E2F1 expression levels via a dominant-negative E2F1 (DN-E2F1) decreases both DNMT1 and HDAC1 binding to the TERT promoter in Huh7 cells (Figure 4C). On the contrary, the binding of the transcription activation-associated factors, RNA polymerase II (Pol II) and acetylated histones H3 and H4 (H3Ac and H4Ac), increased at the TERT promoter region 318–227 bp upstream of ATG after the dominant-negative-mediated inhibition of E2F1 (Figure 4C). Taken together, E2F1 binding to the TERT promoter enhanced DNMT1 and HDAC1 recruitment in Huh7 cells.
Inhibiting E2F1 binding to the TERT promoter does not increase TERT transcription levels in HepG2 cells with the rs2853669 variant
To examine whether the rs2853669 variant (-245T > C) affects E2F1-mediated DNMT1 and HDAC1-binding to the TERT promoter, we analyzed the interaction between E2F1 and the rs2853669 variant in HepG2 cells, which is a HCC cell line with the rs2853669 variant (Supplementary Figure 3). The TERT promoter sequence (from 367 bp upstream of ATG to 35 bp downstream of ATG [26]; Supplementary Figure 3) does not differ between HepG2 and Huh7 cells, except that the former includes the rs2853669 variant, and the latter does not. ChIP analyses after ectopic expression of WT E2F1 (increased E2F1) or DN-E2F1 (decreased E2F1) showed no difference in E2F1-binding to the TERT promoter compared with ectopic expression of an empty vector in HepG2 cells (Figure 5A). The difference in DNMT1 or HDAC1 binding to the TERT promoter was also negligible between E2F1-up-regulated HepG2 cells and E2F1-down-regulated HepG2 cells (Figure 5A). This result is consistent with the observation that the luciferase activity of the TERT promoter with the rs2853669 variant did not exhibit a statistically significant difference when E2F1 expression was up-regulated and when E2F1-binding to the TERT promoter was inhibited (Figure 5B). As expected, the TERT mRNA expression levels in HepG2 did not increase even though endogenous E2F1 activity was inhibited by DN-E2F1 mutant overexpression (Figure 5C), which indicates that rs2853669 is responsible for evading E2F1-induced down-regulation of TERT expression levels.
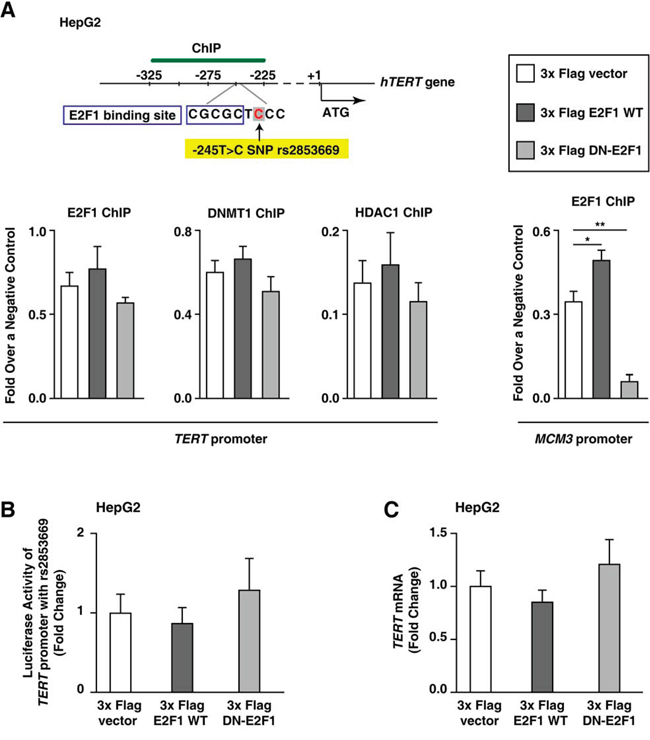
Figure 5: E2F1 does not repress TERT transcription in HepG2 cells with rs2853669. A. ChIP experiments using TERT promoter from each 3x Flag empty vector–, 3x Flag E2F1–, or 3x Flag DN-E2F1–ectopic expressed HepG2 cell line. Primers for Minichromosome maintenance complex component 3 (MCM3) were used as an internal control. B, C. Luciferase assay (B) and qPCR (C) using 3x Flag empty vector–, 3x Flag E2F1–, or 3x Flag DN-E2F1–ectopic expressed HepG2 cells. The data are shown as the mean ± SEM, *P < 0.05, ** P < 0.01.
The rs2853669 variant is associated with a methylated TERT promoter in HCC cell lines and HCC tumors
Next, we validated that the role of E2F1 as a TERT transcription repressor was dependent on DNMT1 activity. We found that 5-aza-2-deoxycytidine (5-aza-dC)-treated Huh7 cells had lower TERT promoter methylation levels and increased TERT transcription levels, despite an increase in the binding of E2F1 to the TERT promoter (Figure 6A). Since previous studies show that 5-aza-dC inhibits DNMT1 activity [29] and that TERT promoter methylation can inhibit the binding of TERT repressors [30], it is possible that DNMT1 regulates the action of E2F1 as a TERT transcription repressor in Huh7 cells. We further examined whether E2F1 localized DNMT1 to the TERT promoter containing the site at -245 bp relative to ATG. E2F1 overexpression increased DNMT1 binding to the TERT promoter and decreased the TERT transcription level; however, concurrent 5-aza-dC treatment and E2F1 overexpression decreased DNMT1-binding to the TERT promoter and increased TERT transcription levels (Figure 6B). The rs2853669 variant disrupted the E2F1 binding-site, which was demonstrated through luciferase reporter and ChIP assays (Figures 3 and 5). Collectively, the rs2853669 variant aids in stimulating TERT transcription levels by preventing E2F1-mediated DNMT1 localization to the TERT promoter.
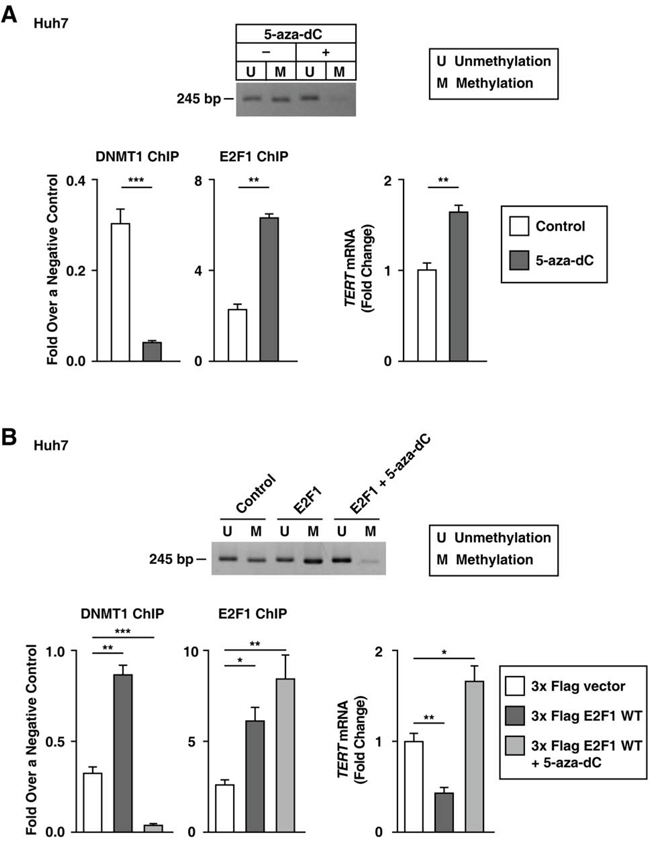
Figure 6: Blocking DNMT1 activity increases TERT transcription levels in Huh7 cells without rs2853669. A, B. MS-PCR using the primer for the TERT CpG island MSP targeting site, qPCR, and the ChIP experiment using 5-aza-dC-treated Huh7 cells (A), and a 3x Flag empty vector–, 3x Flag E2F1–, or 3x Flag E2F1 combined with 5-aza-dC-treated Huh7 cells (B) The data are shown as the mean ± SEM, *P < 0.05, ** P < 0.01, *** P < 0.005. MS-PCR, methylation specific PCR.
The rs2853669 variant (the -245T > C) was not only located near the E2F1 binding site, but also overlapped with an ETS2 binding site at the TERT promoter [10] (Figure 1A). We first examined whether E2F1 affected ETS2 binding to the TERT promoter by ChIP assay (318–227 bp upstream of the ATG start site). The level of binding between ETS2 and the TERT promoter decreased in E2F1 overexpressed–Huh7 cells, whereas the level remained unchanged in E2F1-overexpressed HepG2 cells (Figure 7A). Moreover, in HepG2 cells, the level of ETS2 binding to the TERT promoter did not change when E2F1 binding to the TERT promoter was inhibited (Figure 7B). Interestingly, in Huh7 cells, deficient E2F1 activity increased ETS2 binding to the TERT promoter (Figure 7B). The rs2853669 variant may disrupt both E2F1 and ETS2 binding sites, and thus interfere with both E2F1 and ETS2 binding to the TERT promoter in HepG2 cells. Consistent with the previous studies [12], silencing ETS2 by siRNA decreased TERT transcription levels in Huh7 and HepG2 cells (Figure 7C), which demonstrates that the ETS2 effects on TERT transcription activation differs from E2F1. Using a luciferase assay, we show that the rs2853669 variant increased TERT transcription activity in HCC cell lines (Figure 2A), which suggests that E2F1, not ETS2, dominantly affects the TERT promoter region containing the site at -245 bp relative to ATG. Altogether, E2F1 may occupy the TERT promoter region containing the variant site and then interfere with ETS2 binding to the TERT promoter.
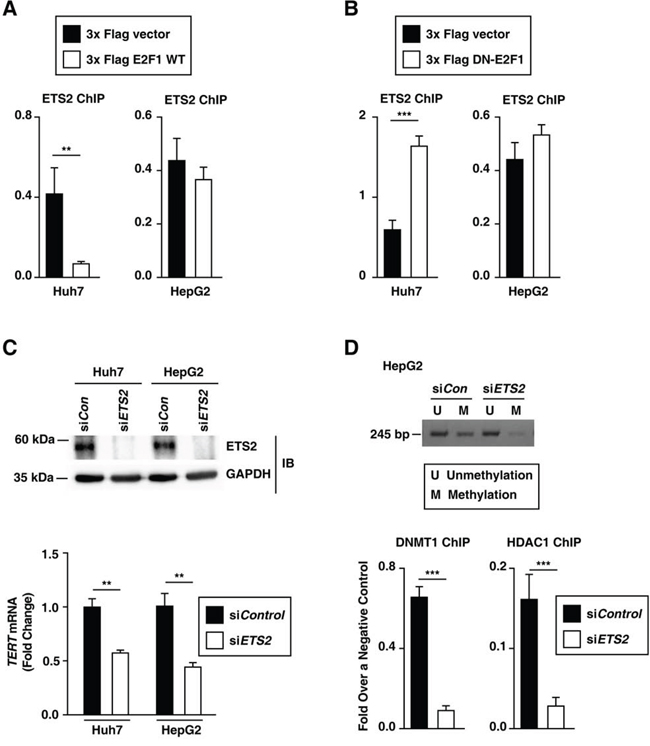
Figure 7: E2F1 and ETS2 regulate the rs2853669 variant-mediated TERT expression in HCC cells. A, B. ETS2 ChIP experiments using the TERT promoter after ectopic expression of each 3x Flag empty vector and 3x Flag E2F1 (A) and a 3x Flag empty vector and 3x Flag DN-E2F1 (B) in Huh7 and HepG2 cells. C. Immunoblot assay and qPCR after siRNA-mediated ETS2 knockdown in Huh7 and HepG2 cells. D. MS-PCR using the primer for the TERT CpG island MSP targeting site and ChIP experiment using siControl (siCon)- and siETS2-treated HepG2 cells. The data are shown as the mean ± SEM, **P < 0.01, *** P < 0.001. MS-PCR, methylation specific PCR.
Both HepG2 cells and Huh7 cells exhibited a -124C > T mutation [19], which creates an ETS2 binding site. Interestingly, ETS2 knockdown decreased both DNMT1 and HDAC1 binding to the TERT promoter and further decreased TERT promoter methylation in HepG2 cells (Figure 7D). Blocking E2F1 alone failed to decrease DNMT1 binding to the TERT promoter in HepG2 cells (Figure 5A), which indicates that ETS2 is required for methylation of the TERT promoter region containing the rs2853669 variant (-245T > C). In Huh7 cells, blocking both E2F1 and ETS2 also decreased both DNMT1 and HDAC1 binding to the TERT promoter and decreased TERT promoter methylation Supplementary Figure 4). Thus, we conclude that both the rs2853669 variant (the co-target of E2F1 and ETS2) and -124C > T mutation (the ETS2 target) are involved in the modulation of methylation at the TERT promoter and further increased the TERT expression in HCC cells.
We previously demonstrated that methylation in the TERT promoter region spanning from -270 bp to -31 bp upstream of the ATG start site correlates with high TERT expression levels and poor recurrence-free survival rates in HCC patients [31]. The present findings show that the rs2853669 variant (-245T > C) and -124C > T mutation combination increase TERT expression, which is involved in TERT promoter methylation as described in our previous report [31] (Figures 2 and 7). We propose that TERT expression is increased by TERT promoter methylation in HCC tumors with the rs2853669 variant and -124C > T mutation combination. The data show that the TERT promoter methylation is positively associated with TERT mRNA expression and TERT protein expression in HCC tumors (Figure 8A, 8B and Supplementary Figure 5), which is consistent with our previous data [31]. To examine whether the rs2853669 variant and -124C > T mutation combination is related to methylation of the TERT promoter containing the site at -245 bp relative to ATG, we quantified TERT promoter methylation levels in the four cases of HCC tumors; with rs2853669 variant alone, with the -124C > T mutation alone, with both the variant and mutation combination, and without neither the variation nor the mutation. We found that the TERT promoter methylation levels were greater in HCC tumors with the rs2853669 variant and -124C > T mutation combination compared with the HCC tumors without both the variation and mutation (P = 0.0009), the HCC tumors the with rs2853669 variant alone (P = 0.0003), and the HCC tumors with the -124C > T mutation alone (P < 0.0001) (Figure 8C). A positive correlation between the TERT promoter methylation level (270–31 bp upstream of the ATG start site) and the TERT mRNA expression level was also confirmed by linear regression analysis (P < 0.0001; Figure 8D). We showed that a combination of the rs2853669 variant (-245T > C) and -124C > T mutation were associated with poor survival rate in HCC patients (Figure 1). Taken together, these results suggest that this combination contributes to the poor survival rate of HCC through TERT promoter methylation-mediated TERT transcriptional activation (Figure 8D).
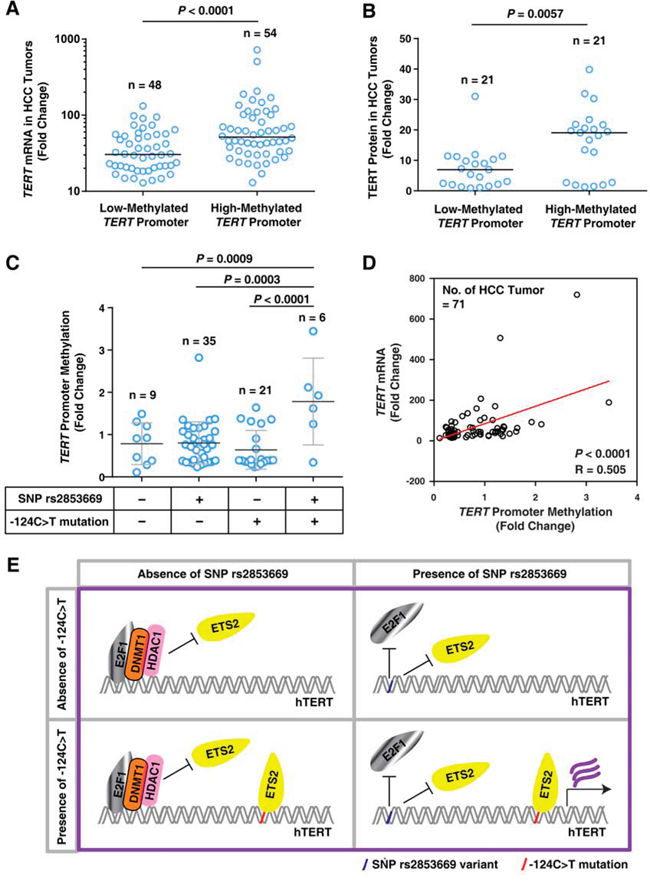
Figure 8: The rs2853669 variant combined with the -124C>T mutation is associated with TERT promoter methylation in HCC tumors. A. Quantification of TERT mRNA levels in HCC tumors with a low-methylated- or high-methylated-TERT promoter. Horizontal bars indicate the median value. Samples were separated into 2 groups based on the median methylation level of TERT promoter. n = 102. B. Quantification of TERT protein expression in HCC tumors with a low-methylated- or high-methylated-TERT promoter. Horizontal bars indicate the median value. Samples were separated into 2 groups based on the median methylation level of TERT promoter. n = 42. C. Quantitation of TERT promoter methylation in HCC tumors without rs2853669 (-245T > C) and the -124C > T mutation or with the SNP rs285366 only, the -124C > T mutation only, or combination of both (-124C > T + SNP). D. Linear regression analysis of TERT promoter methylation levels (%) and TERT mRNA levels. R, Spearman's rank correlation coefficient. E. Molecular model illustrating the potential function of rs2853669 and -124C > T mutation in regulating TERT transcription.
DISCUSSION
Methylation at the TERT promoter has been observed in various tumor tissues and transformed cell lines [32]. Furthermore, it exhibits a positive association with high TERT expression levels and poor survival rates in patients with childhood brain tumors [30]. Our previous data show that TERT promoter methylation contributes to HCC progression by increasing TERT expression [31]. Here, our study demonstrates that TERT promoter methylation was induced by decreasing E2F1 binding (in the presence of rs2853669 (-245T > C)) combined with increasing ETS2 binding at the TERT promoter (in the presence of the -124C > T mutation). Notably, we found that the rs2853669 variant and -124C > T mutation combination is markedly associated with high TERT expression levels, poor overall survival rates, and poor recurrence-free survival rates in HCC patients. HCC patients with the combination also show greater TERT promoter methylation levels compared with HCC patients with the variant alone or mutation alone and HCC patients without both the variant and mutation, indicating that the variant combined with the mutation contributes to TERT promoter methylation, which leads to increased TERT expression, HCC mortality, and HCC recurrence.
The level of TERT expression is increased by the inhibition of E2F1 as a transcriptional repressor through the rs2853669 variant (-245T > C); however, the concurrent stimulation of ETS2 as a transcriptional activator through the -124C > T mutation is also required for increased TERT expression (Figure 8E). The ETS2 binding to the site adjacent to the variant site have been reported to induce c-MYC binding to the TERT promoter, thereby leading to an increase in TERT expression [12]. Previous reports suggested that rs2853669 disrupts the ETS2 binding site, thus reducing the c-MYC-induced TERT expression regulation [17, 33]. However, a recent finding showed that c-MYC knockdown can instead activate TERT promoter activity through enhanced binding of multiple transcription activators to the TERT promoter [34]. As the binding of a transcription factor to its promoter can lead to local structural modification, causing the removal of a preexisting component or recruitment of a new component, the binding of ETS2 to the TERT promoter exhibits the modulation of local structure in the promoter region [12]. A recent study by Bell RJ et al. also suggests that TERT promoter mutations (-124C > T and -146C > T) cooperate with native ETS sites to form high-order structures such as G-quadruplexes; as a result, these structures contribute to the recruitment of the multimeric GA-binding protein (GABP) transcription factor and to the up-regulation of TERT expression [35]. The -245T region is located on a ETS2 binding site, but this ETS2 binding site does not belong to the native ETS sites reported by Bell RJ. et al. [35]. This suggests that TERT expression is regulated by the cooperation of TERT transcription factor binding sites, including TERT promoter mutations, native ETS sites, and SNPs. Thus, it is possible that the rs2853669 and -124C > T mutation combination alters the structural modification of the TERT promoter to improve its activity.
Several SNPs in the telomere maintenance genes are highly associated with the survival rates in HCC patients [6]. According to the data regarding the association between HCC risk and SNPs in the TERT gene, the SNP rs13167280 (IVS3–24 C > T), located on the third intron of the TERT gene, is associated with a decreased risk of HCC progression, whereas no significant association is reported between rs2853669 (-245T > C) and HCC risk [6]. However, our current data demonstrate that the rs2853669 variant combined with the -124C > T mutation at the TERT promoter increases TERT expression, telomere length, and HCC mortality and recurrence rates. Thus, in order to better predict the prognosis of HCC patients, we suggest that studies of SNPs expand to incorporate the relationships and the clinical implications of SNPs when coupled with other SNPs or genetic alterations.
We show that rs2853669 (-245T > C) does not affect the luciferase promoter activity of the TERT reporter vector with the -146C > T mutation (Figure 2A) which is a rare mutation in our HCC patient cohorts (0 case out of 93 cases for the SMH cohort, and 1 case out of 72 cases for the KU cohort; Supplementary Tables 2 and 3). Therefore, the rs2853669 and -124C > T combination likely increases telomerase activity by elevating TERT expression levels, thereby elongating telomere length in HCC. In another study, this combination decreased TERT promoter activity, as assessed by luciferase activity, in urothelial carcinoma cell lines (T24 cell line and CLS-439 cell line) [17], thereby showing that rs2853669 may modulate the TERT promoter activity in a cell type-specific manner. Different cancer types have various TERT promoter mutation frequencies [10, 11, 17, 19, 20]. While 81.8% of urothelial carcinomas carry a -124C > T mutation and 17.8% a -146C > T mutation [17], HCC showed a -124C > T mutation in 93–100% of the cases and a -146C > T mutation in 6–10% of the cases in our study (Supplementary Tables 2 and 3) as well as in other reports [19, 20]. The interaction of TERT promoter mutations and ETS binding sites [35] can regulate the expression of TERT. On the same line, the binding sites for TERT transcription factors can cooperate with SNPs, leading to the activation of TERT via the recruitment of multiple TERT transcription factors. This is probably the reason for cancer-specific effects of SNPs on TERT expression. The rs2853669 variant and -124C > T mutation combination increased TERT promoter activity in four HCC cell lines, which indicates that rs2853669 is involved in regulating TERT promoter activity in cell types that are originated from HCC.
TERT promoter mutations are major genomic alterations in the step-by-step hepatocarcinogenic process, which is involved in HCC developed from chronic liver disease as well as from hepatocellular adenomas (HCA) [36, 37]. A recent report also showed that TERT promoter mutations are key determinants for HCC, as they are observed in low- or high- grade dysplastic nodules (LGDNs, HGDNs), while other mutations in ten cancer genes (CTNNB1, TP53, ARID1A, ARID2, NFE2L2, AXIN1, PIK3CA, KEAP1, RPS6KA3, and CDKN2A) have not been observed in cirrhotic livers, LGDNs, and or HGDNs [38]. Although a number of studies indicate that TERT promoter mutations may be a determinant for HCC, HCC tumors displaying increased TERT expression independent of the TERT mutation status also exist [37]. We showed that the rs2853669 polymorphism combined with TERT promoter mutations increased TERT expression compared to TERT promoter mutations only (Figure 2). Thus, it is possible that the SNP rs2853669 polymorphism combined with TERT promoter mutations, as well as alternative causes, are part of the mechanism responsible for the increase in TERT expression.
In conclusion, we report for the first time that a common variant of the TERT gene, rs2853669, is significantly associated with a high risk of death and cancer recurrence in patients with liver cancer, and that the rs2853669 variant (-245T > C), combined with the -124C > T mutation, mediates TERT transcriptional activity by modulating the binding of both E2F1 and ETS2, which is responsible for the high risk of HCC. Our study suggests that the rs2853669 variant combined with the -124C > T mutation in the TERT promoter is a novel risk factor for poor prognosis in liver cancer.
MATERIALS AND METHODS
Human samples
The Seoul National University Institutional Review Board (SNUIRB No. E1308/001-035) approved this study (Supplementary Materials).
DNA sequencing
Genomic DNA was extracted from HCC tumors and corresponding non-tumorous tissues. The genomic DNA samples from paraffin-embedded tissues and frozen tissues were isolated using the Arcturus PicoPure DNA Extraction Kit (Applied Biosciences, Foster City, CA, USA) and NucleoSpin® TriPrep Kit (Macherey-Nagel, Düren, Germany; 740966.250), respectively, in accordance with the instructions of each manufacturer (Supplementary Materials).
Quantification of telomere fluorescence levels using immunoFISH
An immunoFISH protocol [39, 40] was used with modifications (Supplementary Materials).
Quantification of TERT mRNA expression levels by quantitative real-time PCR
The total RNA was isolated using the NucleoSpin® TriPrep Kit (Macherey-Nagel, Düren, Germany; 740966.250), and cDNA was synthesized using TOPscript™ RT Drymix (dT18) (Enzynomics, Daejeon, Korea; RT200) (Supplementary Materials).
Cell culture and treatment
The HCC cell lines (Huh7, Hep3B, HepG2, and SNU-449) were obtained from the Korean Cell Line Bank (KCLB, Seoul, Korea) (Supplementary Materials).
Luciferase reporter assay
For the luciferase reporter assay, 1.5 × 105 HCC cells were seeded onto 6-well plates and transfected using Fugene® 6 (Roche, Basel, Switzerland) and 1 μg of the wild-type TERT promoter-luciferase construct with or without the variant rs2853669 (-245T > C) or TERT promoter mutation (-124C > T or -146C > T). The wild-type (WT) TERT promoter was subcloned into the pGL3 luciferase empty vector (Promega, Madison, WI, USA) (Supplementary Materials).
Immunofluorescence assay
The cells were fixed using 4% paraformaldehyde, permeabilized using 0.5% Triton X-100 in PBS, blocked with 20% NGS (Normal Goat Serum) and treated with a mouse monoclonal anti-DNMT1 antibody (1:500, Abcam, Cambridge, MA, USA; ab13537) as well as a rabbit polyclonal anti-E2F1 antibody (1:500, ab6302; Cell Signaling) overnight at 4°C. After washing and incubating in a secondary antibody for 1 hour, the slides were mounted using a medium containing DAPI (Vector Laboratories). The images were collected using a confocal microscope (LSM 700; Carl Zeiss, Oberkochen, Germany). The image analyses were performed using Image-Pro plus 6.0 software (Media Cybernetics, Inc., Rockville, MD, USA).
Immunoblot analysis
A total of 2 × 105 cells were boiled for 5 minutes in 2× SDS sample buffer (100 mM Tris–HCl [pH 6.8], 4% SDS, 0.2% bromophenol blue, 20% glycerol, and 200 mM β-mercaptoethanol) and subjected to SDS–PAGE as well as western blotting (Supplementary Materials).
Chromatin immunoprecipitation
Chromatin immunoprecipitation (ChIP) experiments were performed as previously described [29] with certain modifications (Supplementary Materials).
TERT promoter methylation assay
Two micrograms of genomic DNA was treated with sodium bisulfite, and the bisulfite-converted DNA was purified using an EpiTect Bisulfite Kit (Qiagen) in accordance with the manufacturer's instructions (Supplementary Materials).
Statistical analysis
The data were analyzed using R software (www.r-project.org) and GraphPad Software version 4.0 (GraphPad Software Inc., San Diego, CA, USA). The survival data were estimated using the Kaplan–Meier method, and the differences in the survival rates were compared using the log-rank test. The mRNA expression levels and telomere lengths were analyzed using the Mann–Whitney rank sum test, and the promoter activity and ChIP assay were analyzed using a two-tailed t test. The experiments were independently repeated at least three times. The significance values were *P < 0.05, **P < 0.01, and ***P < 0.001.
ACKNOWLEDGMENTS
The biospecimens for this study were provided by Seoul St. Mary's Hospital of the Catholic University of Korea and the Korea University Guro Hospital of National Biobank, a member of the National Biobank of Korea.
CONFLICTS OF INTEREST
The authors have no conflicts of interest to disclose.
GRANT SUPPORT
Funding was provided by the Korea Healthcare Technology Research and Development Project of the Ministry of Health and Welfare of the Republic of Korea (HI14C2094).
Editorial note
This paper has been accepted based in part on peer-review conducted by another journal and the author’s response and revisions as well as expedited peer-review in Oncotarget.
REFERENCES
1. Vinagre J, Pinto V, Celestino R, Reis M, Populo H, Boaventura P, Melo M, Catarino T, Lima J, Lopes JM, Maximo V, Sobrinho-Simoes M and Soares P. Telomerase promoter mutations in cancer: an emerging molecular biomarker? Virchows Arch. 2014; 465:119-133.
2. Daniel M, Peek GW and Tollefsbol TO. Regulation of the human catalytic subunit of telomerase (hTERT). Gene. 2012; 498:135-146.
3. Bojesen SE, Pooley KA, Johnatty SE, Beesley J, Michailidou K, Tyrer JP, Edwards SL, Pickett HA, Shen HC, Smart CE, Hillman KM, Mai PL, Lawrenson K, Stutz MD, Lu Y, Karevan R, et al. Multiple independent variants at the TERT locus are associated with telomere length and risks of breast and ovarian cancer. Nat Genet. 2013; 45:371-384, 384e371-372.
4. Zhong R, Liu L, Zou L, Zhu Y, Chen W, Zhu B, Shen N, Rui R, Long L, Ke J, Lu X, Zhang T, Zhang Y, Wang Z, Sun Y, Cheng L, et al. Genetic variations in TERT-CLPTM1L locus are associated with risk of lung cancer in Chinese population. Mol Carcinog. 2013; 52:E118-126.
5. El-Serag HB and Rudolph KL. Hepatocellular carcinoma: epidemiology and molecular carcinogenesis. Gastroenterology. 2007; 132:2557-2576.
6. Jung SW, Park NH, Shin JW, Park BR, Kim CJ, Lee JE, Shin ES, Kim JA and Chung YH. Prognostic impact of telomere maintenance gene polymorphisms on hepatocellular carcinoma patients with chronic hepatitis B. Hepatology. 2014; 59:1912-1920.
7. Satyanarayana A, Manns MP and Rudolph KL. Telomeres and telomerase: a dual role in hepatocarcinogenesis. Hepatology. 2004; 40:276-283.
8. Shay JW. Determining if telomeres matter in colon cancer initiation or progression. J Natl Cancer Inst. 2013; 105:1166-1168.
9. Hahn WC, Stewart SA, Brooks MW, York SG, Eaton E, Kurachi A, Beijersbergen RL, Knoll JH, Meyerson M and Weinberg RA. Inhibition of telomerase limits the growth of human cancer cells. Nat Med. 1999; 5:1164-1170.
10. Horn S, Figl A, Rachakonda PS, Fischer C, Sucker A, Gast A, Kadel S, Moll I, Nagore E, Hemminki K, Schadendorf D and Kumar R. TERT promoter mutations in familial and sporadic melanoma. Science. 2013; 339:959-961.
11. Huang FW, Hodis E, Xu MJ, Kryukov GV, Chin L and Garraway LA. Highly recurrent TERT promoter mutations in human melanoma. Science. 2013; 339:957-959.
12. Xu D, Dwyer J, Li H, Duan W and Liu JP. Ets2 maintains hTERT gene expression and breast cancer cell proliferation by interacting with c-Myc. J Biol Chem. 2008; 283:23567-23580.
13. Heidenreich B, Nagore E, Rachakonda PS, Garcia-Casado Z, Requena C, Traves V, Becker J, Soufir N, Hemminki K and Kumar R. Telomerase reverse transcriptase promoter mutations in primary cutaneous melanoma. Nat Commun. 2014; 5:3401.
14. Populo H, Boaventura P, Vinagre J, Batista R, Mendes A, Caldas R, Pardal J, Azevedo F, Honavar M, Guimaraes I, Manuel Lopes J, Sobrinho-Simoes M and Soares P. TERT promoter mutations in skin cancer: the effects of sun exposure and X-irradiation. J Invest Dermatol. 2014; 134:2251-2257.
15. Melo M, da Rocha AG, Vinagre J, Batista R, Peixoto J, Tavares C, Celestino R, Almeida A, Salgado C, Eloy C, Castro P, Prazeres H, Lima J, Amaro T, Lobo C, Martins MJ, et al. TERT promoter mutations are a major indicator of poor outcome in differentiated thyroid carcinomas. J Clin Endocrinol Metab. 2014; 99:E754-765.
16. Allory Y, Beukers W, Sagrera A, Flandez M, Marques M, Marquez M, van der Keur KA, Dyrskjot L, Lurkin I, Vermeij M, Carrato A, Lloreta J, Lorente JA, Carrillo-de Santa Pau E, Masius RG, Kogevinas M, et al. Telomerase reverse transcriptase promoter mutations in bladder cancer: high frequency across stages, detection in urine, and lack of association with outcome. Eur Urol. 2014; 65:360-366.
17. Rachakonda PS, Hosen I, de Verdier PJ, Fallah M, Heidenreich B, Ryk C, Wiklund NP, Steineck G, Schadendorf D, Hemminki K and Kumar R. TERT promoter mutations in bladder cancer affect patient survival and disease recurrence through modification by a common polymorphism. Proc Natl Acad Sci U S A. 2013; 110:17426-17431.
18. Borah S, Xi L, Zaug AJ, Powell NM, Dancik GM, Cohen SB, Costello JC, Theodorescu D and Cech TR. Cancer. TERT promoter mutations and telomerase reactivation in urothelial cancer. Science. 2015; 347:1006-1010.
19. Nault JC, Mallet M, Pilati C, Calderaro J, Bioulac-Sage P, Laurent C, Laurent A, Cherqui D, Balabaud C and Zucman-Rossi J. High frequency of telomerase reverse-transcriptase promoter somatic mutations in hepatocellular carcinoma and preneoplastic lesions. Nat Commun. 2013; 4:2218.
20. Chen YL, Jeng YM, Chang CN, Lee HJ, Hsu HC, Lai PL and Yuan RH. TERT promoter mutation in resectable hepatocellular carcinomas: a strong association with hepatitis C infection and absence of hepatitis B infection. Int J Surg. 2014; 12:659-665.
21. Labussiere M, Di Stefano AL, Gleize V, Boisselier B, Giry M, Mangesius S, Bruno A, Paterra R, Marie Y, Rahimian A, Finocchiaro G, Houlston RS, Hoang-Xuan K, Idbaih A, Delattre JY, Mokhtari K, et al. TERT promoter mutations in gliomas, genetic associations and clinico-pathological correlations. Br J Cancer. 2014; 111:2024-2032.
22. Park CK, Lee SH, Kim JY, Kim JE, Kim TM, Lee ST, Choi SH, Park SH and Kim IH. Expression level of hTERT is regulated by somatic mutation and common single nucleotide polymorphism at promoter region in glioblastoma. Oncotarget. 2014; 5:3399-3407.
23. Hosen I, Rachakonda PS, Heidenreich B, Sitaram RT, Ljungberg B, Roos G, Hemminki K and Kumar R. TERT promoter mutations in clear cell renal cell carcinoma. Int J Cancer. 2015; 136:2448-2452.
24. Oh BK, Kim H, Park YN, Yoo JE, Choi J, Kim KS, Lee JJ and Park C. High telomerase activity and long telomeres in advanced hepatocellular carcinomas with poor prognosis. Lab Invest. 2008; 88:144-152.
25. Taubert S, Gorrini C, Frank SR, Parisi T, Fuchs M, Chan HM, Livingston DM and Amati B. E2F-dependent histone acetylation and recruitment of the Tip60 acetyltransferase complex to chromatin in late G1. Mol Cell Biol. 2004; 24:4546-4556.
26. Crowe DL, Nguyen DC, Tsang KJ and Kyo S. E2F-1 represses transcription of the human telomerase reverse transcriptase gene. Nucleic Acids Res. 2001; 29:2789-2794.
27. Robertson KD, Ait-Si-Ali S, Yokochi T, Wade PA, Jones PL and Wolffe AP. DNMT1 forms a complex with Rb, E2F1 and HDAC1 and represses transcription from E2F-responsive promoters. Nat Genet. 2000; 25:338-342.
28. Bird A. DNA methylation patterns and epigenetic memory. Genes Dev. 2002; 16:6-21.
29. Lim SO, Gu JM, Kim MS, Kim HS, Park YN, Park CK, Cho JW, Park YM and Jung G. Epigenetic changes induced by reactive oxygen species in hepatocellular carcinoma: methylation of the E-cadherin promoter. Gastroenterology. 2008; 135:2128-2140, 2140 e2121-2128.
30. Castelo-Branco P, Choufani S, Mack S, Gallagher D, Zhang C, Lipman T, Zhukova N, Walker EJ, Martin D, Merino D, Wasserman JD, Elizabeth C, Alon N, Zhang L, Hovestadt V, Kool M, et al. Methylation of the TERT promoter and risk stratification of childhood brain tumours: an integrative genomic and molecular study. Lancet Oncol. 2013; 14:534-542.
31. Ko E, Jung ES and Jung G. Telomerase reverse transcriptase promoter methylation is related to a risk of recurrence in hepatocellular carcinoma. Hepatology. 2015. doi: 10.1002/hep.27833.
32. Dessain SK, Yu H, Reddel RR, Beijersbergen RL and Weinberg RA. Methylation of the human telomerase gene CpG island. Cancer Res. 2000; 60:537-541.
33. Hsu CP, Hsu NY, Lee LW and Ko JL. Ets2 binding site single nucleotide polymorphism at the hTERT gene promoter—effect on telomerase expression and telomere length maintenance in non-small cell lung cancer. Eur J Cancer. 2006; 42:1466-1474.
34. Zhao Y, Cheng D, Wang S and Zhu J. Dual roles of c-Myc in the regulation of hTERT gene. Nucleic Acids Res. 2014; 42:10385-10398.
35. Bell RJ, Rube HT, Kreig A, Mancini A, Fouse SD, Nagarajan RP, Choi S, Hong C, He D, Pekmezci M, Wiencke JK, Wrensch MR, Chang SM, Walsh KM, Myong S, Song JS, et al. Cancer. The transcription factor GABP selectively binds and activates the mutant TERT promoter in cancer. Science. 2015; 348:1036-1039.
36. Pilati C, Letouze E, Nault JC, Imbeaud S, Boulai A, Calderaro J, Poussin K, Franconi A, Couchy G, Morcrette G, Mallet M, Taouji S, Balabaud C, Terris B, Canal F, Paradis V, et al. Genomic profiling of hepatocellular adenomas reveals recurrent FRK-activating mutations and the mechanisms of malignant transformation. Cancer Cell. 2014; 25:428-441.
37. Pinyol R, Tovar V and Llovet JM. TERT promoter mutations: gatekeeper and driver of hepatocellular carcinoma. J Hepatol. 2014; 61:685-687.
38. Nault JC, Calderaro J, Di Tommaso L, Balabaud C, Zafrani ES, Bioulac-Sage P, Roncalli M and Zucman-Rossi J. Telomerase reverse transcriptase promoter mutation is an early somatic genetic alteration in the transformation of premalignant nodules in hepatocellular carcinoma on cirrhosis. Hepatology. 2014; 60:1983-1992.
39. Plentz RR, Park YN, Lechel A, Kim H, Nellessen F, Langkopf BH, Wilkens L, Destro A, Fiamengo B, Manns MP, Roncalli M and Rudolph KL. Telomere shortening and inactivation of cell cycle checkpoints characterize human hepatocarcinogenesis. Hepatology. 2007; 45:968-976.
40. Ko E and Jung G. Positive association of long telomeres with the invasive capacity of hepatocellular carcinoma cells. Biochem Biophys Res Commun. 2014; 447:358-363.