
INTRODUCTION
The EGFR family of receptor tyrosine kinases consists of four closely related family members: EGFR (Her1), ErbB2 (HER2), ErbB3 (Her3), and ErbB4 (Her4) [1]. These receptors are important regulators of normal growth and cell differentiation. Their gene amplification, overexpression or mutation is associated with tumor development and poor clinical prognosis in most of the human cancers [2]. These cell surface receptors are characterized by a composite extracellular domain which contains a well defined ligand-binding site in at least three of the four members (EGFR, ErbB3 and ErbB4), a single pass transmembrane domain, followed by an intracellular domain where a tyrosine kinase domain and a C-terminal non-catalytic signaling tail can be distinguished [3].
Signaling through these receptors is mediated by homodimerization or heterodimization with other family members, usually mediated by a rather promiscuous set of ligands. Multiple ligands are known to bind to the EGFR family. Some of them bind specifically to EGFR (EGF, TGF-α, and amphiregulin) or ErbB4 (neuregulin 3, neuregulin 4, and tomoregulin), whereas others have dual specificity (e.g., β-cellulin, epiregulin, and heparin-binding EGF-like growth factor for both EGFR and ErbB4, and neuregulins 1 and 2 for both ErbB3 and ErbB4) [4].
With the exception of HER2, the extracellular domains of the EGFR family members are unable to form stable homo- or heterodimers in the absence of ligand. Binding of the ligand with the extracellular domain of its corresponding receptor induces a structural reconfiguration, which promotes the exposure of the otherwise tethered dimerization domain; this allows the formation of dimers which results in receptor transphosphorylation of tyrosine residues within the activation loop leading to enhanced kinase activity with subsequent phosphorylation of tyrosine residues in the carboxy-terminal domain. This in turn enables recruitment of signaling molecules and activation of downstream intracellular signaling pathways [5] (Fig 1). The EGFR family members can also be activated through ligand-independent activation. These mechanisms include activation by non-physiological stimuli (e.g., oxidative stress, UV, and γ-irradiation) or by other receptor tyrosine kinases (notably MET, IGF-1R, or TRK-B). The importance of dimerization in EGFR family signaling is best illustrated by HER2 and ErbB3. HER2 is the preferred dimerization partner for all the EGFR family members. However, although HER2 has the strongest kinase activity, there is no known ligand to this receptor. Therefore, activation of HER2 is dependent upon dimerization with other family members [6]. In contrast, ErbB3 carries a well defined binding site for ligands but lacks intrinsic kinase activity and is therefore dependent upon heterodimerization to be phosphorylated in its signaling tail and to induce downstream signaling [6].
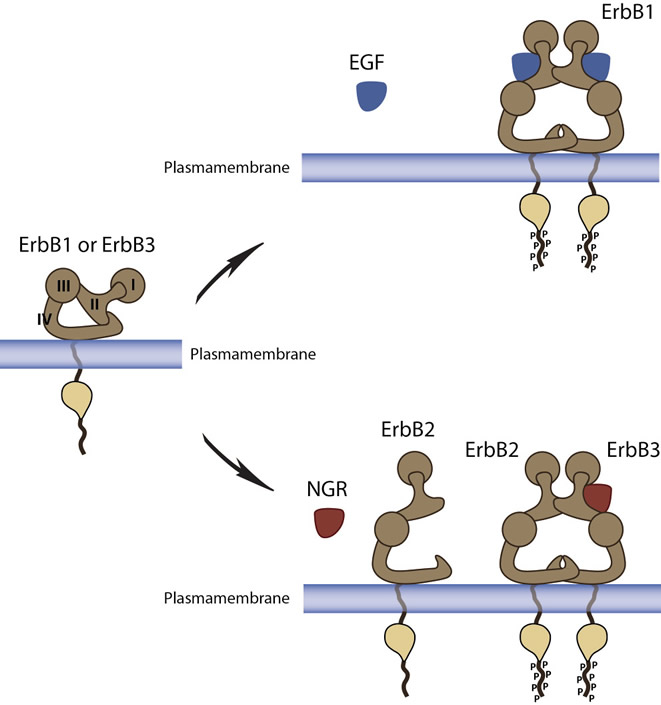
Figure 1: ErbB receptors ligand dependent change of conformation and signal transduction. In the absence of ligand, a direct intramolecular interaction between domains II and IV keeps ErbBs in a closed (locked or tethered) conformation that prevents interaction between domains I and III. This conformation disrupts the ligand-binding pocket and buries the dimerization arm of domain II. Upper panel: EGFR or ErbB1. Lower Panel: HER2 is inherently unable to dimerize because of a strong interaction between domains I and III which leads to a constitutively extended dimerization arm. HER2 is therefore constantly primed for interactions with ligand-bound receptors of the ErbB family. In the presence of NRG, the dimerization loop from domain II of ErbB3 extends to interact intramolecularly with a ligandless, primed HER2 monomer to form the oncogenic HER2-ErbB3 heterodimer.
Activated EGFR family members recruit various adaptors and signaling molecules through the phosphorylated cytoplasmic domain, which further leads to activation of a variety of downstream signaling pathways [7]. All of the EGFR family members activate the extracellular signal-regulated kinase (Erk)1/2 via recruitment of Grb2 or Shc adaptors [8, 9]. Activation of Erk1/2 has an important role in EGFR stimulated cell proliferation. Activation of another important signaling pathway for cell proliferation and survival, the phosphatidylinositol 3-kinase (PI3K)/Akt pathway, however, differs between the EGFR family members. Whereas ErbB3 and ErbB4 are capable of binding the p85 regulatory subunit of PI3K directly through their putative p85 binding sites (tyrosine-X-X-methionine), EGFR and HER2 bind indirectly to p85 through adaptors or through heterodimerization with ErbB3 or ErbB4 [10,11]. In addition to Erk1/2 and PI3K activation, phosphorylated EGFR family members can also activate a collection of transcription factors such as c-fos, c-jun, c-myc, signal transducer and activator of transcription, NF-kB, zinc-finger transcription factor, and Ets family members.
Two EGFR family receptor members, namely EGFR itself and HER2, have been amongst the most extensively studied targets for the therapy of cancer over the past twenty years and a wealth of drugs directed against them have been either already approved or are in advanced clinical development. EGFR is being targeted with the monoclonal antibodies cetuximab and panitumumab [12-17] and with two low molecular weight tyrosine kinase inhibitors, gefitinib and erlotinib [18-23] in several types of epithelial cancers, including head and neck, pancreatic, colorectal and a subset of non small cell lung cancers with mutant or highly expressed EGFR.
HER2 is targeted by the monoclonal antibody trastuzumab (Herceptin™) in breast cancers [24, 25], where it is overexpressed in approximately 20-25% of cases and this approach is now clinically approved for the treatment of breast cancer patients expressing HER2 at high levels (+3 herceptest™). However, trastuzumab had little or no effectiveness against cancers of the prostate [26], pancreas [27], colon and rectum [28] or lung epithelia [29], which also very often express HER2 at lower levels. Other antibodies targeting HER2 are Pertuzumab, a humanized antibody recognizing a different HER2 epitope involved in homo- or heterodimerization [30] which has very recently been approved by FDA in combination with trastuzumab and docetaxel for the treatment of patients with HER2-positive metastatic breast cancer (mBC) who have not received prior anti-HER2 therapy or chemotherapy for metastatic disease; and T-DM1, a drug conjugated version of trastuzumab, which has recently demonstrated superb clinical effects in a controlled Phase II trial conducted in Trastuzumab-resistant patients [31]. Finally, lapatinib, a small molecule inhibitor of both EGFR and HER2 has been developed [32], which is currently used in the treatment of HER2+ metastatic breast cancer resistant to trastuzumab. [33].
ErbB3 has been disregarded for several years as a cancer target, although the elevated expression of this receptor in several human cancers led in early times to postulate its involvement in tumor progression [34]. This low interest in ErbB3 was also due to the lack of detectable mutations in cancer samples and the absence of a strongly active tyrosine kinase in its intracellular domain [35]. However, during the past 5 years a mounting number of evidences have been accumulated pointing to a key role of this receptor in tumorigenesis and cancer progression and, above all in the establishment of resistance to therapies [36]. These evidences have triggered major efforts towards the development of anti-ErbB3 therapies. Because this receptor is devoid of strong intrinsic kinase activity, the major strategy in this case is the generation of monoclonal antibodies directed against the receptor. Some of these have already entered clinical development. It is reasonable to expect therefore, that in the next years if clinical proof-of-concept is achieved ErbB3 may become a next generation blockbuster target for cancer therapies.
Validation of ErbB3 as new cancer target
ErbB3 in Lung cancer
ErbB3 was shown to be highly expressed in lung adenocarcinomas and associated with poor prognosis as measured by immunohistochemistry [37]. Moreover quantitative real time indicated that high ErbB3 expression was associated with decreased survival in patients with early stage (I-IIIA) NSCLC [38]. ErbB3 was constitutively activated at high level in several lung adenocarcinoma cell lines, which also express HER2 [39]. Co-expression of ErbB3 with other ErbB family members was indicative of tumor recurrence [40]. The expression of the proliferation-associated marker Ki-67 at a higher frequency in ErbB3-positive NSCLC cases than in ErbB3-negative tumors was suggestive of a contribution of ErbB3 to an aggressive cancer behaviour [41]. Additional strong evidence for the importance of ErbB3 expression in lung tumorigenesis came from ErbB3 transgenic mice. These mice treated with the carcinogen methylnitrosourea developed a high incidence of lung tumors with reduced latency compared to wild type mice [42]. In addition mice double transgenic for human ErbB3 and rat HER2/neu had an incidence of spontaneous lung tumors similar to that of single transgenic mice, but with a shorter latency [42], thus demonstrating the relevance of HER2/ErbB3 heterodimers in lung tumorigenesis.
ErbB3 has also been shown to play a role in drug resistance: suboptimal pathway inhibition by tyrosine kinase inhibitors (TKIs) was shown to result in a compensatory shift in ErbB3 activation [43]. In this case ablation of ErbB3 expression by siRNAs restored efficacy and pro-apoptotic activities of TKIs [43]. Likewise, amplification of cMet was described to occur in cells resistant to TKI treatment and cMet transphosphorylation of ErbB3 was shown to be a mechanism whereby resistant cells can circumvent blockade of EGFR activity [44]. These data suggest that targeting ErbB3 could be a novel strategy to treat drug resistant tumors.
ErbB3 in Breast cancer
In breast cancer, increased ErbB3 expression is common. Indeed ErbB3 mRNA levels relative to normal gland and measured by real-time PCR are increased in 46% of breast cancers [45] and correlate positively with those for ErbB4 and negatively with EGFR mRNA [46, 47].
ErbB3 protein is detectable in 50–70% of human breast cancers by immunohistochemistry [48, 49]. Moreover, tyrosine-phosphorylated ErbB3 is frequently over-expressed in breast tumors that overexpress HER2 [48, 50]. This led to the suggestion that ErbB3 could be an important partner of HER2 in the development of breast tumors. It has been shown, in fact, in NIH 3T3 in vitro assays and in a transgenic mouse model that the heterodimer HER2-ErbB3 is able to transform cells and to induce mammary tumors [51]. Finally, Holbro and colleagues have demonstrated that HER2 inactivation blocked proliferation in HER2-overexpressing cells and this was associated with a decrease of ErbB3 phosphorylation [52].
On the other hand, there seems to be lack of clarity about the relationships between ErbB3 expression and estrogen receptor (ER). At the protein level, ErbB3 and ER did not correlate [50, 53] and a high percentage of ER-negative tumors were strongly positive for ErbB3 [50].
The ErbB3 receptor is emerging as a critical element not only in HER2-mediated transformation and tumor progression but also in drug resistance. In fact, inhibition of HER2 phosphorylation by TKIs targeting EGFR and HER2 in HER2+ breast cancer cells is followed by feedback upregulation of activated ErbB3 [54]. Other evidences underline the importance of ErbB3 in HER2 addicted breast cancers. In HER2-overexpressing cells, inhibitors of the PI3K pathway induce a compensatory up-regulation of the expression and phosphorylation of ErbB3 [55, 56]. Moreover, knocking down ErbB3 results in sensitization to PI3K inhibitors [57]. In breast cancer cell lines BTK474-HR20 and SKBR3-pool2 selected in vitro for resistance to trastuzumab a strong upregulation of ErbB3, pErbB3, IGF1R and pIGFR is observed which strongly contribute to cell proliferation [58]. Indeed in these cells destabilization of both ErBB3 and IGF1R by metformin exerts a strong anti-proliferative effect [58]. Finally in recent studies the group of Cook and collaborators [59, 60] has shown, using a variety of animal models, that ErbB3 ablation impairs PI3K/Akt-dependent mammary tumorigenesis and p44/42 phosphorylation in pre-neoplastic HER2-overexpressing mammary glands and tumors. This was associated with decreased growth of pre-existing HER2-overexpressing tumors and improved tumor responses to HER2 tyrosine kinase inhibitors. All together these findings suggest that ErbB3 cooperates with HER2 to induce changes in breast epithelium before, during and after tumor formation. Hence, therapeutic targeting of ErbB3 in combination with HER2 may be the most appropriate strategy to achieve full efficacy of anti-HER2 targeted therapy, in particular for breast cancer.
ErbB3 in Colorectal cancer
ErbB3 is occasionally mutated in colon carcinomas [61]. ErbB3 mRNA is overexpressed in the majority of human colon cancer cell lines and in human colorectal tumors [62]. Few studies have investigated the ErbB3 protein expression: the data obtained show that ErbB3 expression can be detected in 36-90% of colorectal cancer [63, 64]. ErbB3 is frequently co-expressed together with EGFR and HER2 [65]. In a recent study, colon carcinoma cells were transfected with a single chain antibody against HER2 and showed inactivation of HER2 tyrosine phosphorylation and reduced heterodimerization of HER2 with ErbB3. This reduction resulted in a increase of EGFR expression with the subsequent formation of heterodimers of EGFR with ErbB3 [66]. The signalling by EGFR-ErbB3 heterodimers may play an important role in colon cancer cell lines; in fact, inhibition of AKT phosphorylation and cell proliferation by the EGFR-specific inhibitor erlotinib (Tarceva) correlated with expression of ErbB3 [67]. As further evidence of ErbB3’s importance in this type of cancer, inhibition of proliferation and induction of apoptosis in HT-29 colon cancer cells by conjugated linoleic acid may be mediated by its ability to downregulate ErbB3 signaling and PI3K/ AKT pathway [68].
ErbB3 in Melanoma
ErbB3 is frequently expressed in human melanoma cell lines [69] and microarray analysis revealed that ErbB3 was one of a small number of genes whose upregulation is characteristic of melanoma [70]. Immuno-histochemical analysis showed high ErbB3 expression in melanoma metastases and suggested increased ErbB3 expression associated with disease progression [71]. Moreover, activated ErbB3 has been observed in primary melanocytic lesions and this suggests that activation of NRG1/ErbB3 signaling may contribute to the progression of melanoma from benign nevi to metastatic malignancies. The role of ErbB3 in melanoma can also be indirectly inferred from the evidence [72] that driver mutations are found in ErbB4, its preferred heterodimerizing partner in these cells, in a significant percentage of melanomas (approximately 20%). These mutations can be grouped into two main categories: a) kinase activating mutations similar to those found in EGFR in NSCLC and Head & Neck cancer; b) single aminoacid mutatations in domain II or IV of the extracellular domain which affect the intramolecular contact between these two domains and induce a shift towards a constitutively open conformation of the receptor [72]. Therefore, inhibition of ErbB3 with monoclonal antibodies is expected to impair the growth promoting effect of mutant ErbB4-ErbB3 heterodimers in melanomas.
Clear cell sarcoma of soft tissue (CCSST) is a rare lesion of the musculoskeletal system affecting children and young adults with melanocytic differentiation. ErbB3 is one of the most particularly up-regulated genes in these cancers. Cell lines derived from these tumors expressed ErbB3 protein and either HER2 or ErbB4 [73]; in half of the lines ErbB3 was constitutively activated by endogenous NRG1 expression; the others were responsive to added NRG1 [74].
ErbB3 in Ovarian Cancer
It has been reported that ErbB3 mRNA expression, as detected by RT-PCR, is increased in a proportion of ovarian cancers [75]. Of the ovarian carcinomas, 16% of the tumors overexpressed ErbB3 protein compared to normal ovarian [76]; moreover expression of NRG was detected in the majority of ovarian carcinomas and cell lines and this could be a potential for autocrine regulation of cell growth [77]. It has recently been confirmed that there was a direct correlation between ErbB3 overexpression and poor overall prognosis [78].
In OVCAR8 xenograft mouse model, down-regulation of ErbB3 by RNAi decreased tumor progression compared to controls, in the same mouse model treatment with a monoclonal anti-ErbB3 antibody also resulted in inhibition of tumour progression [79].
Signaling activation by ErbB3 in ovarian cancers seems to depend in some cases upon the formation of heterodimers with HER2 while in others, the activation of ErbB3 is independent of heterodimers with HER2 or EGFR. Interestingly, four alternate c-ErbB3 transcripts (1.6, 1.7, 2.1 and 2.3 kb) were isolated from an ovarian carcinoma-derived cell line. Fibroblasts transfected with all four transcripts express truncated ErbB-3 products. Three of these four products are soluble secreted proteins [80].
Targeting ErbB3: Anti-ErbB3 antibodies
A wealth of antibodies targeting human ErbB3 has been generated in the past years. In this section we will review their biochemical/biological properties and current status of development. Interestingly while the majority of these agents recognize only ErbB3, a subset of them has been engineered thanks to new technologies, to be able to bind not only ErbB3 but also an additional co-receptor. A comprehensive list of such antibodies is listed in table 1 and described in the following sections.
Monospecific anti-ErbB3 antibodies
MM-121
MM-121, a fully human IgG2 mAb that binds specifically to ErbB3, was isolated by selection from a phage display Fab library and screening for high-affinity binders by surface plasmon resonance [81]. MM-121 is being developed by Merrimack in partnership with Sanofi-Aventis. This antibody has a KD of approx 0.75 nM for human ErbB3, is cross reactive with mouse ErbB3 and competes for heregulin-1β binding to the receptor. The precise binding epitope of MM-121 to ErbB3 has not been disclosed, but the antibody is able, via a direct or indirect mechanism, to block ligand–induced receptor heterodimerization with HER2 (Fig.2). MM-121 inhibits HRG-1-induced ErbB3 and AKT phosphorylation with IC50 values of 2.4 and 6.0 nM respectively and completely inhibits in vivo tumor xenografts of renal ACHN cells. Further studies have subsequently shown through the analysis of a panel of several cell lines of various origin, that MM-121 is able to affect tumor growth only in a subset of them [82]. Sensitive tumors were further subdivided into strongly and partially responsive. In the attempt to identify biomarkers of responsiveness, the authors assessed several parameters such as ligands and ErbB receptor levels, and found that, although efficacy did not correlate with a single parameter, the best single feature capable to predict responsiveness was HRG-β1 expression. Therefore, MM-121 is mainly active in cancers with ligand-dependent activation. MM-121 effects were also assessed in a mouse model of lung cancer caused by doxycycline-induced over-expression of a double mutant EGFR T790M-L858R. This model mimics resistance to TKIs that develops in patients following the secondary mutation T790M and is characterized by increased heregulin expression and phosphorylation of ErbB3. MM-121, which is cross-reactive to the mouse homologue, was poorly active when administered as monotherapy, but exterted strong tumor growth inhibitory effect when given in combination with cetuximab [82]. MM-121 is currently being assessed in several Phase I and II clinical trials in breast cancer, non-small cell lung cancer, ovarian cancer and other solid tumor cancers.
Table 1: List and properties of anti-ErbB3 antibodies under development
Antibody name |
Company |
Isotype |
Affinity for human ErbB3 |
Other receptors recognized |
Receptor internalization and degradation |
ADCC Activity |
Stage of development |
Monospecific |
|||||||
AMG-888 |
U3 Pharma/Amgen/Daiichi-Sankyo |
ND - fully human |
ND |
ND |
ND |
Phase II |
|
MM-121 |
Merrimack/Sanofi Aventis |
IgG2 - fully human |
0.75 nM |
mErbB3 |
yes |
ND |
Phase II |
GE-huMab-HER3 |
Roche |
IgG1 - humanized |
ND |
ND |
Strong |
Phase I |
|
AV-203 |
Aveo Pharmaceuticals |
IgG1 - humanized |
0.076 nM |
cynoErbB3 |
ND |
ND |
Phase I |
TK-A3 |
Takis |
IgG1 - humanized |
7.2 nM |
mErbB3 |
yes |
ND |
PreC |
TK-A4 |
Takis |
IgG1 - humanized |
2.2 nM |
yes - strong |
ND |
PreC |
|
MP-RM-1 |
Mediapharma |
ND |
32 nM |
yes |
ND |
PreC |
|
LJM716 |
Novartis and Sanofi Aventis |
ND ? - fully human |
ND |
ND |
ND |
PreC |
|
REGN1400 |
Regereron |
ND? - fully human |
0.05 nM |
ND |
ND |
PreC |
|
Bispecific |
|||||||
MEHD7945A |
Genentech |
IgG1?- fully human |
0.4 nM |
hEGFR |
ND |
yes |
Phase I |
MM-111 |
Merrimack |
HSA linked to scFv |
10 nM |
hHER2 |
ND |
ND |
Phase I |
MM-141 |
Merrimack |
HAS linked to svFv |
ND |
hIGF1R |
ND |
ND |
PreC |
Legend: ND: Not disclosed;cyno, cynomologous monkey; m, mouse; h, human
AMG 888
AMG 888 (also known with the name U3-1287) is a fully human anti ErbB3 monoclonal isolated from U3 Pharma using Amgen’s Xenomouse® technology. It is currently being developed by Amgen in partnership with Daiichi–Sankyo. AMG 888 was reported to block ErbB-induced AKT and ERK signaling and to inhibit in vitro and in vivo growth of multiple tumor cell lines as single agent or, even better in combination with other ErbB family inhibitors, such as cetuximab. However, the exact biochemical properties of the antibody have not been disclosed in detail [83].
Also AMG 888 is currently undergoing clinical trials. Phase I data have been reported at the 2011 ASCO meeting [84]. The phase I trial had 2 parts: based on positive tolerability and Pharmacokinetic (PK) data in part 1, a part 2 dose-expansion was conducted on 31 cancer patients with advanced solid tumors the majority of whom (no.17) were NSCLC cancer patients, at doses of 9, 14 and 20 mpk. PK data showed that steady state was reached after 3 dosing cycles and Cmin was >10-fold greater than the threshold required to achieve 90% inhibition of ErbB3 phosphorylation in xenograft studies. Eight patients experienced stable disease. Of these one patients had approximately 26% tumor shrinkage and a partial metabolic response monitored by FDG-PET. Based on these highly encouraging data AMG 888 has been brought into Phase II clinical development.
TK-A3 and TK-A4
TK-A3 and TK-A4 are both mouse monoclonals of IgG1a isotype generated by Takis in collaboration with the University of Catanzaro (Italy), through muscle electroporation with the human ErbB3 cDNA carrying the H584F mutation which gives rise to a constitutively extended conformation of the receptor in the absence of ligand [85]. TK-A3 and A4 show an affinity for human ErbB3 of 7.2 and 2.2 nM respectively as measured by Biacore; they block ligand binding to the receptor with an IC50 of 1.5 and 2.2 nM respectively and inhibit pErbB3 ligand-dependent stimulation with an IC50 of 4 and 6 nM respectively. In vitro, these two anti-ErbB3 antibodies modulate the growth rate of cancer cells of different origins and in vivo they show antitumor properties in several xenograft models. TK-A3 but not TK-A4 is cross reactive against the murine ErbB3 receptor. Therefore its efficacy was assessed in the HER2-driven carcinogenesis genetically engineered BALB/neuT mouse model for mammary tumor. In this model TK-A3 significantly slowed down tumor growth, increase time-to-disease progression and downregulated in vivo ErbB3-mediated signaling [85]. TK-A3 and TK-A4 mechanism of action has been studied in greater detail in two melanoma cell lines. It was shown through a series of combined cell biology approaches that antibody efficacy strongly correlated with antibody-induced receptor internalization, degradation and inhibition of receptor recycling to the cell surface (Fig3) [86]. Epitope mapping studies using a peptide array scanning approach allowed to define the epitope bound by TK-A3 but not the one recognized by TK-A4. Interestingly, TK-A3 recognized the dimerization loop in domain II of the receptor extracellular region [86]. The availability of two monoclonals directed against different epitopes of the ErbB3 extracellular domain allows to assess the potential synergy of combined treatments. Indeed it has been previously shown that combination of pairs of mAbs recognizing distinct epitopes in HER2, exert a superior anti-tumor effect than the use of individual antibodies [87]. Since the effect was observed both in vivo and in vitro, it is likely that this is not due to the involvement of immunological mechanisms but to a more powerful downregulation of the signalling survival pathways of HER2 expressing cancer cells exerted by the antibodies combinations. Humanization of TK-A3 and TK-A4 has been achieved, with maintenance of similar affinity for ErbB3 receptor and biological properties such as inhibition of NRG induced signalling and cell proliferation (our unpublished data).
GE-huMab-HER3
GE-huMab-HER3 is a humanized and glycoengineered IgG1 antibody that is being developed by Roche [88]. Also this antibody prevents ligand binding and receptor heterodimerization. It inhibits ligand-dependent ErbB3 and Akt phosphorylation with an IC50 of approximately 0.1 µg/ml). GE-huMab-HER3 efficacy in vivo was assessed on a panel of primary NSCLC xenografts, where it caused >50% tumor growth inhibition in 10 out of 17 NSCLC cell lines and in some cases even resulted in complete tumor remission. A unique feature of GE-huMab-HER3 that differentiates it from all other anti-ErbB3 antibodies is its ability to bind to human FcγRIIIa on immune effector cells with a 50-fold higher affinity than standard IgG1 antibodies. Therefore, GE-huMab-HER3 has superior potency and efficacy in ADCC, both in in vitro assays and in in vivo models. This property may confer to GE-huMabHER3 superior clinical efficacy, but also potentially a higher degree of toxicity. Phase I clinical testing of this antibody has recently started.
AV-203
AV-203 is a humanized immunoglobulin IgG1 kappa antibody that is being developed by Aveo Pharmaceuticals [89]. AV-203 shows a very high affinity for human ErbB3 (Kd = 76 pM at 37°C), is cross-reactive to cynomolgous monkey ErbB3 but not to mouse ErbB3. AV-203 is a potent inhibitor of ligand-dependent ErbB3 activation and its downstream signaling molecule Akt, blocks ErbB3/HER2 heterodimerization and moderately down regulates ErbB3 receptor in vitro and in vivo. The antibody shows potent inhibition of tumor growth in a broad spectrum of xenograft models in which ErbB3 is activated by its ligand NRG1 or by HER2 overexpression. AV-203 has recently started Phase I clinical development in 2012.
MP-RM-1
MP-RM-1 is a mouse monoclonal of undisclosed isotype generated by Mediapharma through repeated mice immunizations with NIH-3T3 cells transfected with the human ErbB3 coding sequence [90]. In spite of the weak affinity for the receptor (Kd 32 nM) this antibody is able to fully inhibit ligand-induced ErbB3/Akt activation in several cell lines in a time and dose-dependent fashion. The antibody is also able to induce receptor internalization and degradation, although with a slower kinetic than signaling inhibition. MP-RM-1 has been shown to block formation of ligand-induced heterodimers of ErbB3 with HER2, but is unable to compete with the ligand for binding to the receptor [90]. In the absence of a precise knowledge about the epitope recognized by this antibody it is possible to speculate that its binding to the receptor hinders the dimerization epitope or impairs the ligand-induced conformational change needed to expose the dimerization interface. MP-RM-1 has been shown to slow down tumor growth approximately 50% when given as single agent in two xenograft models tested. Perhaps the most interesting feature of this antibody is its ability to suppress ligand-independent activation of ErbB3. This was assessed in MKN-45 human gastric cancer cells, which do not express NRG-1β and betacellulin and where ErbB3 constitutive phosphorylation is triggered by association with cMet. MP-RM-1 was shown to fully inhibit pErbB3 in a manner similar to a cMet inhibitor and in co-immunoprecipitation studies was shown to inhibit association of ErbB3 with cMet [90]. This finding has important implications for the treatment of tumors where activation of ErbB3 is not ligand-dependent.
LMJ716
LJM716 is a fully human antibody isolated from a Morphosys phage library and being developed by Novartis and Sanofi Aventis [91]. LJM716 potently inhibits ErbB3/ Akt phosphorylation and proliferation in both ligand dependent and HER2 amplified ligand-independent ErbB3 dependent cell lines in vitro; it induces tumor growth inhibition or even regression, both as single agent or in combination with other ErbB receptor family targeted agents. LJM716 does not compete with NRG for binding to ErbB3. The crystal structure LJM716 bound to the ErbB3 extra-cellular domain revealed binding to a conformational epitope contained within domains 2 and 4 and trapping ErbB3 in the inactive conformation. LJM716 therefore possesses a novel mechanism of action; it prevents the structural rearrangements required for ErbB3 activation induced by either HER2 or NRG.
REGN1400
REGN1400 is a fully-human anti-ErbB3 monoclonal antibody under development by Regeneron Pharmaceuticals Inc. [92]. The antibody shows a very high affinity for the receptor (Kd approx 50pM) and a potent inhibition of NRG binding (IC50 approx 30pM). REGN1400 inhibits phosphorylation of ErbB3 and Akt in multiple human tumor cell lines and growth of these cell lines in vitro. REGN1400 strongly inhibits the growth of tumor xenografts in a dose-dependent manner both as single agent or in combination with anti EGFR or anti HER2 antibodies.
Bispecific antibodies
MEHD7945A
MEHD7945A is a fully human dual specificity anti-ErbB3 and anti-EGFR monoclonal antibody under development by Genentech which was isolated from phage display Fab libraries [93, 95]. The objective of the authors was to obtain a wider spectrum monoclonal antibody capable of affecting ligand dependent proliferation of cancers which require signaling by EGFR or ErbB3 or both receptors simultaneously. The rationale behind the generation of this antibody was that, given similar structural configuration of the two receptors, their partial sequence homology and their similar mode of activation by ligand, it should be possible to obtain a normal IgG capable of interacting with either of the two antigens in their relevant ligand binding sites. The authors first isolated a monospecific anti-EGFR antibody from a phage library with diversity on the heavy chain CDRs. Then, by constructing a library with mutations in the light-chain CDRs, they were able to select clones that acquired binding to ErbB3 while maintaining binding to EGFR. MEHD7945A has a Kd of 1.9 nM for EGFR and of 0.4 nM for human ErbB3 respectively, is cross reactive with mouse EGFR but not with mouse ErbB3. MEHD7945A potently inhibits heregulin-induced pErbB3, pAkt and pERK 1/2 with IC50 values of 0.05, 0.19, and 1.13 µg/ml respectively. Likewise it inhibits TGFα induced pEGFR and pERK 1/2 with IC50 values of 0.03 and 0.16 µg/ml respectively. In in vitro proliferation assays in cell lines that are mainly dependent upon HER2-ErbB3 heterodimers (H1666) or upon EGFR-EGFR homodimers and EGFR heterodimers with other ErbB receptors, the antibody was more potent than monospecific antibodies such as cetuximab, pertuzumab or an anti-ErbB3 antibody.
The binding epitopes in EGFR and ErbB3 recognized by MEHD7945A were identified by X-ray crystallography of antibody receptor complexes. As expected they map in similar regions of domain 3, and overlap with the ligand binding site. However, the two epitopes are slightly shifted by a few Angstroms and are constituted by non homologous aminoacids. The binding mode of the antibody therefore suggests that its main mechanism of action is the block of ligand binding and fixing the receptors in a tethered conformation (Fig2).
MEHD7945A activity in vivo was tested in twelve xenograft models of various origin (NSCLC, ovarian, breast, pancreas, colorectal) in parallel with treatments with monospecific monoclonals against EGFR (cetuximab) or against ErbB3. In a subset of cases its activity was compared with the combination of the two monospecific antibodies. As expected from its mode of action MEHD7945A showed a broader profile and was efficacious in all lines tested: it had equal activity to cetuximab or to anti-ErbB3 in tumors which depend upon EGFR or ErbB3 respectively, and had higher potency than their combination in tumors which depend on both receptors. Interestingly, MEHD7945A is also able to induce antibody-dependent cellular cytotoxicity (ADCC) as shown in a series of in vitro and in vivo studies [93].
MM-111
MM-111 was rationally designed to be able to target tumor cells which acquire resistance to current HER2 inhibitors, such as trastuzumab or lapatinib. It has been observed that the mechanism of resistance is in most cases linked to ligand-dependent hyperactivation of ErbB3 in the presence of amplified-HER2 expression. In order to more efficiently target this type of tumors Merrimack pharmaceuticals recently generated MM-111 a bispecific antibody directed against both HER2 and ErBB3 [95]. MM-111 is made of two distinct single chain antibodies, B1D2 and H3 recognizing HER2 and ErbB3 with an affinity of 0.3 nM and 10 nM respectively, linked to a variant of recombinant human serum albumin (MHSA) with short connector linkers inserted at the amino and carboxy-termini of the molecule. MM-111 is able to engage simultaneously both receptors with the formation of inactive trimers and shows avidity binding to cells with an affinity for ErbB3 superior to that of the single sFv component (Fig.2). When tested in vitro in a panel of cell lines, MM-111 inhibits NRG induced pErbB3 and cell proliferation with an efficiency that correlates, as expected, with the degree of HER2 overexpression levels. MM-111 is stable in serum, in vivo upon injection in mice and shows prolonged terminal half-life. When tested as single agent in xenograft models its anti-tumor potency again correlated with the degree of HER2 expression, being most effective in HER2 +3 tumors at the dose of 30 mpk. Although MM-111 is directed against both HER2 and ErbB3 receptors, it is unable as single agent to fully block their activity. This can be indirectly inferred by the results of combination studies in vitro and in vivo with lapatinib or trastuzumab. Indeed combination of MM-111 with either lapatinib or trastuzumab in BT474 cells always showed superior efficacy. It is not clear why MM-111 alone is unable to fully inhibit HER2-ErbB3 activation, but one likely possibility is that this antibody does not trigger efficiently receptor internalization and degradation, This results in a residual receptor activation which needs additional targeted agents to be fully eliminated.
MM141
The evidence that a subset of cancer cell lines show simultaneous production of NRG and IGF1 and this gives rise to simultaneous activation of both receptors led Merrimack to generate a bispecific antibody directed against ErBB3 and IGF1R which was called MM-141 [96]. MM-141 blocks both NRG binding to ErbB3 and IGF-1/IGF-2 binding to IGF1R and inhibits downstream PI3K/Akt pathway activation (Fig.2). MM-141 may have the potential to be active on a wider spectrum of human tumors over a broad range of receptor profiles. More studies are however needed to precisely understand its superiority to monospecific anti-ErbB3 antibodies or dual specificity anti ErbB3-EGFR or anti-ErbB3-anti-HER2 antibodies.
CONCLUSIONS
In recent years ErbB3 has emerged as a key player in the establishment of malignancy. Disregarded for several years largely because of the absence of detectable ErbB3 mutations in cancer samples and for the lack of a functional kinase domain in its intra-cytoplasmic region, ErbB3 is instead one of the most potent activators of the PI3K/Akt axis and is upregulated and trans-phosphorylated in several forms of cancer, in particular following treatment with EGFR and HER2 inhibitors. Hence ever growing evidences point to ErbB3 as an important factor in the establishment of resistance to therapies.
The most effective strategy to specifically target ErbB3 is by use of monoclonal antibodies. The number of anti-ErbB3 antibodies in development is impressive and unprecedented (see Table 1). This clearly underscores the importance this target has reached during the past years and the high expectation about their therapeutic impact.
Most importantly anti-ErbB3 monoclonals may become “universal” ingredients in “combination cocktails” conceived to attack tumors resistant to small molecule inhibitors or monoclonal antibodies directed against other EGFR family members [97]. Indeed it has been shown that both in tumors resistant to trastuzumab or in tumor resistant to gefitinib, ErbB3 is upregulated and, most importantly pErbB3 is constitutively activated. Therefore in these cases anti-ErbB3 antibodies able to strongly induce receptor internalization and degradation and to affect simultaneously both PI3K and ERK pathways activation may significantly sensitize resistant tumors to the action of HER2 or EGFR inhibitors, respectively.
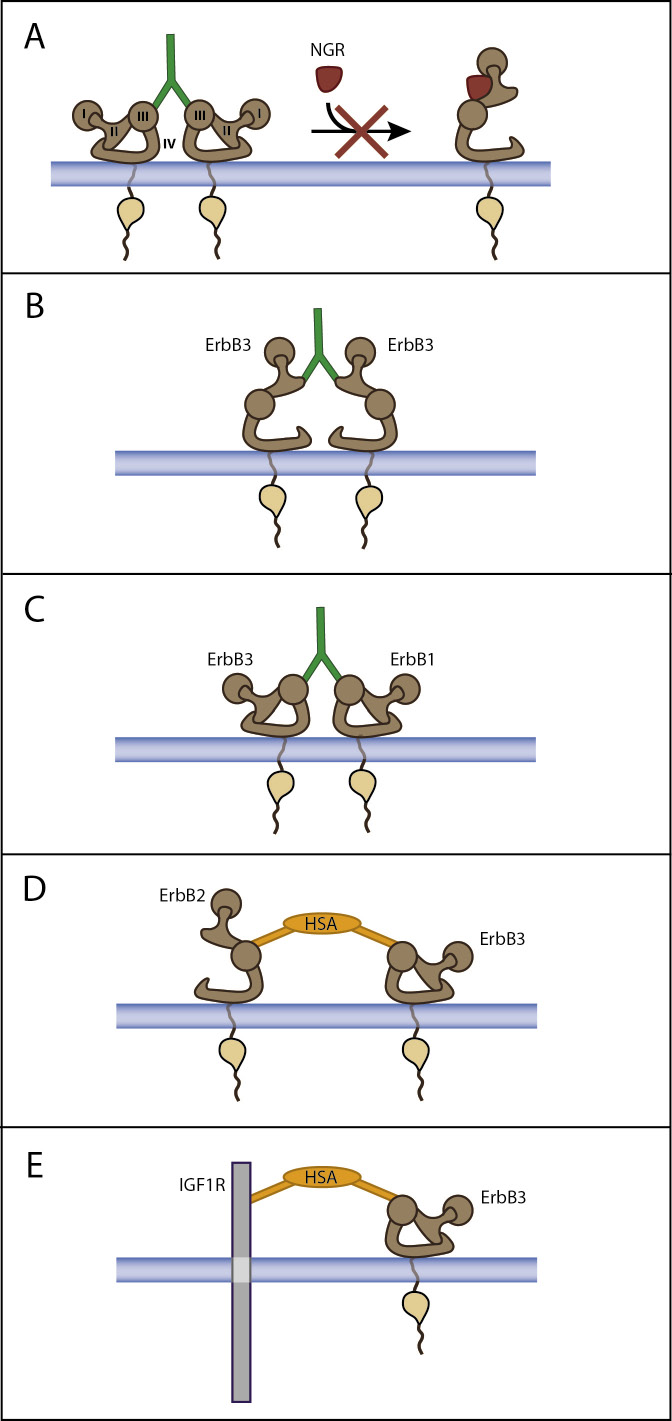
Figure 2: Mode of action of monospecific and dual specificity anti-ErbB3 antibodies. A) a monospecific antibody binding to domain III inhibits ligand binding and blocks the receptor in a tethered configuration unable to form heterodimers with other receptors. B) the antibody binds to the heterodimerization loop located in domain II. C) a dual specificity antibody binds to domain III of two distinct receptors, such as ErbB3 and EGFR. D) a dual specificity antibody interacts with its two scFv domains simultaneously with both HER2 and ErbB3. E) a dual specificity antibody against ErbB3 and IGF1R. HSA: human serum albumin.
These agents may have significantly different mode of binding, some of which are schematically represented in Figures 2 and 3. Figure 2 indicates the mode of action of some monospecific and dual specificity anti-ErbB3 antibodies. In Panel A a monospecific antibody binding to domain III inhibits ligand binding and blocks the receptor in a tethered configuration unable to form heterodimers with other receptors. In panel B the antibody (such as TK-A3) binds to the heterodimerization loop located in domain II. In panel C a dual specificity antibody such as MEHD7945A binds to domain III of two distinct receptors ErbB3 and EGFR. It is not excluded the possibility of formation of heterotrimers. In panel D a dual specificity antibody MM-111 interacts with its two scFv domains simultaneously with both HER2 and ErbB3. Panel E shows the mode of binding of a dual specificity antibody against ErbB3 and IGF1R such as MM-141.
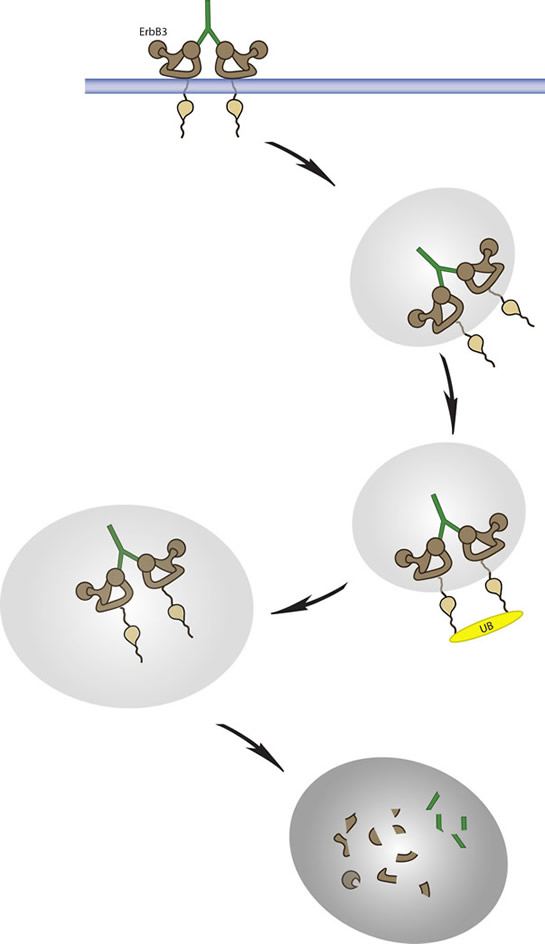
Figure 3: Internalization and degradation as therapeutic action by anti-ErbB3 antibodies. A hypothetical model of ErbB3 endocytosis and intracellular sorting mediated by anti-ErbB3 antibodies. Antibody binding to ErbB3 leads to receptor engagement, inhibition of phosphorylation and induced sorting of the ErbB3 receptor into lysosomes. ErbB3 receptor polyubiquitination and lysosomal degradation is the final step.
Figure 3 shows how antibodies capable of triggering strong receptor internalization and degradation can act a strong therapeutic agents by wiping out this key signaling receptor from cells. This is for example what we have observed with TK-A3 and TK-A4 [86]. Furthermore, in addition to these direct effects it must be considered the possible indirect contribution of the immune system through ADCC for which agents such as GE-huMab-HER3 and MEHD7945A have been optimized.
With some distinct features, all the antibodies possess the biochemical properties and the therapeutic effects in preclinical models desired for a anti-cancer product. Obviously it is difficult to believe that all these agents will become marketed agents. Therefore, the race to which of them will be the winner(s) is entirely open. This will certainly depend upon the speed in advancing these agents to the clinic. However, a major role will be played by the strategy adopted in clinical development:
- will it be possible to identify suitable biomarkers for the selection of responders?
- will it be more convenient to target ligand-dependent tumors or ligand-independent ones?
- and in both cases which will be the best combination agent and tumor type to target first?
These are still open questions which are the main focus of current research and for which we hope answers will be found soon.
CONFLICT OF INTEREST
Dr Aurisicchio, Dr Marra and Dr Roscilli are employed at Takis.
ACKOWLEDGEMENTS
We thank Cinzia Roffi and Manuela Emili for editorial assistance. This work was supported by grant AIRC IG10334 and MIUR to G. Ciliberto
Reference
1. Leahy DJ. Structure and function of the epidermal growth factor (EGF/ErbB) family of receptors. Adv Protein Chem. 2004;68:1-27.
2. Kolibaba KS, Druker BJ. Protein tyrosine kinases and cancer. Biochim Biophys Acta. 1997;1333(3):F217-48.
3. Yamamoto T, Ikawa S, Akiyama T, Semba K, Nomura N, Miyajima N, Saito T, Toyoshima K. Similarity of protein encoded by the human c-erb-B-2 gene to epidermal growth factor receptor. Nature 1986;319:230-234.
4. Burgess AW, Cho HS, Eigenbrot C, Ferguson KM, Garrett TP, Leahy DJ, Lemmon MA, Sliwkowski MX, Ward CW, Yokoyama S. Mol Cell 2003;12(3):541-52.
5. Schlessinger J. The epidermal growth factor receptor as a multifunctional allosteric protein. Biochemistry 1988;27(9):3119-23.
6. Citri A, Skaria KB, Yarden Y. The deaf and the dumb: the biology of ErbB-2 and ErbB-3. Exp Cell Res. 2003;284(1):54-65.
7. Yarden Y, Sliwkowski MX. Untangling the ErbB signaling network. Nat Rev Mol Cell Biol. 2001;2(2):127-37.
8. Brock R, Jovin TM. Heterogeneity of signal transduction at the subcellular level: microsphere-based focal EGF receptor activation and stimulation of Shc translocation. J Cell Sci 2001;114(Pt 13):2437-47.
9. Liebmann C. Regulation of MAP Kinase activity by peptide receptor signaling pathway: paradigms of multiplicity. Cell Signal. 2001;13(11):777-85.
10. Vivanco I, Sawyers CL. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev Cancer 2002;2(7):489-501.
11. Hemi R. Transactivation of ErbB2 and ErbB3 by tumor necrosis factor-alpha and anisomycin leads to impaired insulin signaling through serine/threonine phosphorylation of IRS proteins. J Biol Chem. 2002;277(11):8961-9.
12. Krempien R, Muenter MW, Huber PE, Nill S, Friess H, Timke C, Didinger B, Buechler P, Heeger S, Herfarth KK, Abdollahi A, Buchler MW, Debus J. Randomized phase II--study evaluating EGFR targeting therapy with cetuximab in combination with radiotherapy and chemotherapy for patients with locally advanced pancreatic cancer--PARC: study protocol [ISRCTN56652283]. BMC Cancer 2005;5:131.
13. Socinski MA. Antibodies to the epidermal growth factor receptor in non small cell lung cancer: current status of matuzumab and panitumumab.Clin Cancer Res. 2007;13(15 Pt 2):s4597-601.
14. Jonker DJ, O’Callaghan CJ, Karapetis CS, Zalcberg JR, Tu D, Au HJ, Berry SR, Krahn M, Price T, Simes RJ, Tebbutt NC, van Hazel G, Wierzbicki R, Langer C, Moore MJ. Cetuximab for the treatment of colorectal cancer. N Engl J Med 2007;357(20):2040-8.
15. Gholam D, Chebib A, Hauteville D, Bralet MP, Jasmin C. Combined paclitaxel and cetuximab achieved a major response on the skin metastases of a patient with epidermal growth factor receptor-positive, estrogen receptor-negative, progesterone receptor-negative and human epidermal growth factor receptor-2-negative (triple-negative) breast cancer. Anticancer Drugs.2007;18(7):835-7.
16. Giusti RM, Shastri KA, Cohen MH, Keegan P, Pazdur R. FDA drug approval summary: panitumumab (Vectibix). Oncologist. 2007;12(5):577-83.
17. Rosell R, Robinet G, Szczesna A, Ramlau R, Constenla M, Mennecier BC, Pfeifer W, O’Byrne KJ, Welte T, Kolb R, Pirker R, Chemaissani A, Perol M, Ranson MR, Ellis PA, Pilz K, et al. Randomized phase II study of cetuximab plus cisplatin/vinorelbine compared with cisplatin/vinorelbine alone as first-line therapy in EGFR-expressing advanced non-small-cell lung cancer. Ann Oncol. 2008;19(2):362-9.
18. Pollack VA, Savage DM, Baker DA, Tsaparikos KE, Sloan DE, Moyer JD, Barbacci EG, Pustilnik LR, Smolarek TA, Davis JA, Vaidya MP, Arnold LD, Doty JL, Iwata KK, Morin MJ. Inhibition of epidermal growth factor receptor-associated tyrosine phosphorylation in human carcinomas with CP-358,774: dynamics of receptor inhibition in situ and antitumor effects in athymic mice. J Pharmacol Exp Ther. 1999;291(2):739-48.
19. Baselga J, Averbuch SD. ZD1839 (‘Iressa’) as an anticancer agent. Drugs 2000;60 Suppl 1:33-40; discussion 41-2.
20. Lal A, Glazer CA, Martinson HM, Friedman HS, Archer GE, Sampson JH, Riggins GJ. Mutant epidermal growth factor receptor up-regulates molecular effectors of tumor invasion. Cancer Res. 2002;62(12):3335-9.
21. Fukuoka M, Yano S, Giaccone G, Tamura T, Nakagawa K, Douillard JY, Nishiwaki Y, Vansteenkiste J, Kudoh S, Rischin D, Eek R, Horai T, Noda K, Takata I, Smit E, Averbuch S, et al. Multi-institutional randomized phase II trial of gefitinib for previously treated patients with advanced non-small-cell lung cancer (The IDEAL 1 Trial) [corrected]. J Clin Oncol. 2003;21(12):2237-46.
22. Kris MG, Natale RB, Herbst RS, Lynch TJ Jr, Prager D, Belani CP, Schiller JH, Kelly K, Spiridonidis H, Sandler A, Albain KS, Cella D, Wolf MK, Averbuch SD, Ochs JJ, Kay AC. Efficacy of gefitinib, an inhibitor of the epidermal growth factor receptor tyrosine kinase, in symptomatic patients with non-small cell lung cancer: a randomized trial. JAMA. 2003;290(16):2149-58.
23. Bareschino MA, Schettino C, Troiani T, Martinelli E, Morgillo F, Ciardiello F. Erlotinib in cancer treatment. Annal Oncol. 2007;18 Suppl 6:vi35-41.
24. McNeil C. New Pap test technologies hit heavy seas but sales keep flying. J Natl Cancer Inst. 1998;90(18):1327-9.
25. ShaK S. Overview of the trastuzumab (Herceptin) anti-HER2 monoclonal antibody clinical program in HER2-overexpressing metastatic breast cancer. Herceptin Multinational Investigator Study Group. Semin Oncol. 1999;26(4 Suppl 12):71-7.
26. Ziada A, Barqawi A, Glode LM, Varella-Garcia M, Crighton F, Majeski S, Rosenblum M, Kane M, Chen L, Crawford ED. The use of trastuzumab in the treatment of hormone refractory prostate cancer; phase II trial. Prostate 2004;60(4):332-7.
27. Safran H, Iannitti D, Ramanathan R, Schwartz JD, Steinhoff M, Nauman C, Hesketh P, Rathore R, Wolff R, Tantravahi U, Hughes TM, Maia C, Pasquariello T, Goldstein L, King T, Tsai JY, et al. Herceptin and gemcitabine for metastatic pancreatic cancers that overexpress HER-2/neu. Cancer Invest. 2004;22(5):706-12.
28. Ramanathan RK, Hwang JJ, Zamboni WC, Sinicrope FA, Safran H, Wong MK, Earle M, Brufsky A, Evans T, Troetschel M, Walko C, Day R, Chen HX, Finkelstein S. Low overexpression of HER-2/neu in advanced colorectal cancer limits the usefulness of trastuzumab (Herceptin) and irinotecan as therapy. A phase II trial. Cancer Invest. 2004;22(6):858-65.
29. Clamon G, Herndon J, Kern J, Govindan R, Garst J, Watson D, Green M; Cancer and Leukemia Group B. Lack of trastuzumab activity in nonsmall cell lung carcinoma with overexpression of erb-B2: 39810: a phase II trial of Cancer and Leukemia Group B. Cancer. 2005;103(8):1670-5.
30. Nahta R, Hung MC, Esteva FJ. The HER-2-targeting antibodies trastuzumab and pertuzumab synergistically inhibit the survival of breast cancer cells. Cancer Res. 2004;64(7):2343-6.
31. Burris HA 3rd, Tibbitts J, Holden SN, Sliwkowski MX, Lewis Phillips GD. Trastuzumab emtansine (T-DM1): a novel agent for targeting HER2+ breast cancer. J Clin Oncol 2011;11(5):275-82.
32. Xia W, Mullin RJ, Keith BR, Liu LH, Ma H, Rusnak DW, Owens G, Alligood KJ, Spector NL. Anti-tumor activity of GW572016: a dual tyrosine kinase inhibitor blocks EGF activation of EGFR/erbB2 and downstream Erk1/2 and AKT pathways. Oncogene 2002;21(41):6255-63.
33. Blackwell KL, Pegram MD, Tan-Chiu E, Schwartzberg LS, Arbushites MC, Maltzman JD, Forster JK, Rubin SD, Stein SH, Burstein HJ. Single-agent lapatinib for HER2-overexpressing advanced or metastatic breast cancer that progressed on first- or second-line trastuzumab-containing regimens. Ann Oncol. 2009;20(6):1026-31.
34. Gullick WJ. The c-erbB3/HER3 receptor in human cancer. Cancer Surv. 1996;27:339-49.
35. Citri A, Yarden Y. EGF-ERBB signalling: towards the systems level. Nat Rev Mol Cell Biol. 2006;7(7):505-16.
36. Hsieh AC, Moasser MM. Targeting HER proteins in cancer therapy and the role of the non-target HER3. Br J Cancer. 2007;97(4):453-7.
37. Yi ES, Harclerode D, Gondo M, Stephenson M, Brown RW, Younes M, Cagle PT. High c-erbB-3 protein expression is associated with shorter survival in advanced non-small cell lung carcinomas. Mod. Pathol. 1997;10(2):142-8.
38. Müller-Tidow C, Diederichs S, Bulk E, Pohle T, Steffen B, Schwäble J, Plewka S, Thomas M, Metzger R, Schneider PM, Brandts CH, Berdel WE, Serve H. Identification of metastasis-associated receptor tyrosine kinases in non-small cell lung cancer. Cancer Res. 2005;65(5):1778-82.
39. Sithanandam G, Smith GT, Masuda A, Takahashi T, Anderson LM, Fornwald LW. Cell cycle activation in lung adenocarcinoma cells by the ErbB3/phosphatidylinositol 3-kinase/Akt pathway. Carcinogenesis 2003;24(10):1581-92.
40. Lai WW, Chen FF, Wu MH, Chow NH, Su WC, Ma MC, Su PF, Chen H, Lin MY, Tseng YL. Immunohistochemical analysis of epidermal growth factor receptor family members in stage I non-small cell lung cancer. Ann. Thorac. Surg. 2001;72(6):1868-76.
41. Hilbe W, Dirnhofer S, Oberwasserlechner F, Eisterer W, Ammann K, Schmid T, Hilbe G, Thaler J, Wöll E. Immunohistochemical typing of non-small cell lung cancer on cryostat sections: correlation with clinical parameters and prognosis. J. Clin. Pathol. 2003;56(10):736-41.
42. Zhou H, Liu L, Lee K, Qin X, Grasso AW, Kung HJ, Willis JE, Kern J, Wagner T, Gerson SL. Lung tumorigenesis associated with erb-B-2 and erb-B-3 overexpression in human erb-B-3 transgenic mice is enhanced by methylnitrosourea. Oncogene 2002;21(57):8732-40.
43. Sergina NV, Rausch M, Wang D, Blair J, Hann B, Shokat KM, Moasser MM. Escape from HER-family tyrosine kinase inhibitor therapy by the kinase-inactive HER3. Nature. 2007;445(7126):437-41.
44. Engelman JA, Zejnullahu K, Mitsudomi T, Song Y, Hyland C, Park JO, Lindeman N, Gale CM, Zhao X, Christensen J, Kosaka T, Holmes AJ, Rogers AM, Cappuzzo F, Mok T, Lee C, et al. MET amplification leads to gefitinib resistance in lung cancer by activating ERBB3 signaling. Science. 2007;316(5827):1039-43.
45. Li Q, Ahmed S, Loeb JA. Development of an autocrine neuregulin signaling loop with malignant transformation of human breast epithelial cells. Cancer Res 2004;64(19):7078-85.
46. PawLowski V, Révillion F, Hebbar M, Hornez L, Peyrat JP. Prognostic value of the type I growth factor receptors in a large series of human primary breast cancers quantified with a real-time reverse transcription-polymerase chain reaction assay. Clin Cancer Res 2000;6(11):4217-25.
47. Revillion F, Lhotellier V, Hornez L, Bonneterre J, Peyrat JP. ErbB/HER ligands in human breast cancer, and relationships with their receptors, the bio-pathological features and prognosis. Ann Oncol 2008;19(1):73-80.
48. Lemoine NR, Barnes DM, Hollywood DP, Hughes CM, Smith P, Dublin E, Prigent SA, Gullick WJ, Hurst HC. Expression of the ERBB3 gene product in breast cancer. Br J Cancer 1992;66(6):1116-21.
49. Witton CJ, Reeves JR, Going JJ, Cooke TG, Bartlett JM. Expression of the HER1-4 family of receptor tyrosine kinases in breast cancer. J Pathol. 2003;200(3):290-7.
50. Naidu R, Yadav M, Nair S, Kutty MK. Expression of c-erbB3 protein in primary breast carcinomas. Br J Cancer 1998;78(10):1385-90.
51. Alimandi M, Romano A, Curia MC, Muraro R, Fedi P, Aaronson SA, Di Fiore PP, Kraus MH. Cooperative signaling of ErbB3 and ErbB2 in neoplastic transformation and human mammary carcinomas. Oncogene 1995;10(9):1813-21.
52. Holbro T, Beerli RR, Maurer F, Koziczak M, Barbas CF 3rd, Hynes NE. The ErbB2/ErbB3 heterodimer functions as an oncogenic unit: ErbB2 requires ErbB3 to drive breast tumor cell proliferation. PNAS 2003;100(15):8933-8.
53. Travis A, Pinder SE, Robertson JF, Bell JA, Wencyk P, Gullick WJ, Nicholson RI, Poller DN, Blamey RW, Elston CW, Ellis IO. C-erbB-3 in human breast carcinoma: expression and relation to prognosis and established prognostic indicators. Br J Cancer 1996;74(2):229-33.
54. Sergina NV, Rausch M, Wang D, Blair J, Hann B, Shokat KM, Moasser MM. Escape from HER-family tyrosine kinase inhibitor therapy by the kinase-inactive HER3. Nature. 2007;445(7126):437-41.
55. Garrett JT, Olivares MG, Rinehart C, Granja-Ingram ND, Sánchez V, Chakrabarty A, Dave B, Cook RS, Pao W, McKinely E, Manning HC, Chang J, Arteaga CL. Transcriptional and posttranslational up-regulation of HER3 (ErbB3) compensates for inhibition of the HER2 tyrosine kinase. Proc Natl Acad Sci U S A. 2011;108(12):5021-6.
56. Chakrabarty A, Sánchez V, Kuba MG, Rinehart C, Arteaga CL. Feedback upregulation of HER3 (ErbB3) expression and activity attenuates antitumor effect of PI3K inhibitors. Proc Natl Acad Sci U S A. 2012;109(8):2718-23.
57. Serra V, Scaltriti M, Prudkin L, Eichhorn PJ, Ibrahim YH, Chandarlapaty S, Markman B, Rodriguez O, Guzman M, Rodriguez S, Gili M, Russillo M, Parra JL, Singh S, Arribas J, Rosen N, et al. PI3K inhibition results in enhanced HER signaling and acquired ERK dependency in HER2-overexpressing breast cancer. Oncogene. 2011;30(22):2547-57.
58. Liu B, Fan Z, Edgerton SM, Yang X, Lind SE, Thor AD. Potent anti-proliferative effects of metformin on trastuzumab-resistant breast cancer cells via inhibition of erbB2/IGF-1 receptor interactions. Cell Cycle. 2011 Sep 1;10(17):2959-66.
59. Cook RS, Garrett JT, Sánchez V, Stanford JC, Young C, Chakrabarty A, Rinehart C, Zhang Y, Wu Y, Greenberger L, Horak ID, Arteaga CL. ErbB3 ablation impairs PI3K/Akt-dependent mammary tumorigenesis. Cancer Res. 2011;71(11):3941-51.
60. Vaught DB, Stanford JC, Young C, Hicks DJ, Wheeler F, Rinehart C, Sánchez V, Koland J, Muller WJ, Arteaga CL, Cook RS. HER3 Is Required for HER2-Induced Preneoplastic Changes to the Breast Epithelium and Tumor Formation. Cancer Res. 2012;72(10):2672-82.
61. Jeong EG, Soung YH, Lee JW, Lee SH, Nam SW, Lee JY, Yoo NJ, Lee SH. ERBB3 kinase domain mutations are rare in lung, breast and colon carcinomas. Int J Cancer 2006;119(12):2986-7.
62. Ciardiello F, Kim N, Saeki T, Dono R, Persico MG, Plowman GD, Garrigues J, Radke S, Todaro GJ, Salomon DS. Differential expression of epidermal growth factor-related proteins in human colorectal tumors. PNAS 1991;88(17):7792-6.
63. Rajkumar T, Gooden CS, Lemoine NR, Gullick WJ, Goden CS. Expression of the c-erbB-3 protein in gastrointestinal tract tumours determined by monoclonal antibody RTJ1. J Pathol 1993;170(3):271-8.
64. Kountourakis P, Pavlakis K, Psyrri A, Rontogianni D, Xiros N, Patsouris E, Pectasides D, Economopoulos T. Prognostic significance of HER3 and HER4 protein expression in colorectal adenocarcinomas. BMC cancer 2006;6:46.
65. Venkateswarlu S, Dawson DM, St Clair P, Gupta A, Willson JK, Brattain MG. Autocrine heregulin generates growth factor independence and blocks apoptosis in colon cancer cells. Oncogene 2002;21(1):78-86.
66. Hu YP, Venkateswarlu S, Sergina N, Howell G, St Clair P, Humphrey LE, Li W, Hauser J, Zborowska E, Willson JK, Brattain MG. Reorganization of ErbB family and cell survival signaling after Knock-down of ErbB2 in colon cancer cells. J Biol Chemistry 2005;280(29):27383-92.
67. Buck E, Eyzaguirre A, Haley JD, Gibson NW, Cagnoni P, Iwata KK. Inactivation of Akt by the epidermal growth factor receptor inhibitor erlotinib is mediated by HER-3 in pancreatic and colorectal tumor cell lines and contributes to erlotinib sensitivity. Mol Cancer Ther. 2006;5(8):2051-9.
68. Cho HJ, Kim WK, Kim EJ, Jung KC, Park S, Lee HS, Tyner AL, Park JH. Conjugated linoleic acid inhibits cell proliferation and ErbB3 signaling in HT-29 human colon cell line. Am J Physiol Gastrointest Liver Physiol 2003;284(6):G996-1005.
69. Ferby I, Reschke M, Kudlacek O, Knyazev P, Pantè G, Amann K, Sommergruber W, Kraut N, Ullrich A, Fässler R, Klein R. Mig6 is a negative regulator of EGF receptor-mediated skin morphogenesis and tumor formation. Nat Med 2006;12(5):568-73.
70. Gyorffy B, Lage H. A Web-based data warehouse on gene expression in human malignant melanoma. J Invest Dermatol 2007;127(2):394-9.
71. Reschke M, Mihic-Probst D, van der Horst EH, Knyazev P, Wild PJ, Hutterer M, Meyer S, Dummer R, Moch H, Ullrich A. HER3 is a determinant for poor prognosis in melanoma. Clin Cancer Res 2008;14(16):5188-97.
72. Rudloff U, Samuels Y. A growing family: adding mutated Erbb4 as a novel cancer target. Cell Cycle. 2010 Apr 15;9(8):1487-503.
73. Schaefer KL, Brachwitz K, Wai DH, Braun Y, Diallo R, Korsching E, Eisenacher M, Voss R, Van Valen F, Baer C, Selle B, Spahn L, Liao SK, Lee KA, Hogendoorn PC, Reifenberger G, et al. Expression profiling of t(12;22) positive clear cell sarcoma of soft tissue cell lines reveals characteristic up-regulation of potential new marker genes including ERBB3. Cancer Res. 2004;64(10):3395-405.
74. Schaefer KL, Brachwitz K, Braun Y, Diallo R, Wai DH, Zahn S, Schneider DT, Kuhnen C, Vollmann A, Brockhoff G, Gabbert HE, Poremba C. Constitutive activation of neuregulin/ERBB3 signaling pathway in clear cell sarcoma of soft tissue. Neoplasia 2006;8(7):613-22.
75. Mandai M, Konishi I, Koshiyama M, Mori T, Arao S, Tashiro H, Okamura H, Nomura H, Hiai H, Fukumoto M. Expression of metastasis-related nm23-H1 and nm23-H2 genes in ovarian carcinomas: correlation with clinicopathology, EGFR, c-erbB-2, and c-erbB-3 genes, and sex steroid receptor expression. Cancer Res. 1994;54(7):1825-30.
76. Rajkumar T, Stamp GW, Hughes CM, Gullick WJ. c-erbB3 protein expression in ovarian cancer. Clin Mol Pathol. 1996;49(4):M199-202.
77. Gilmour LM, Macleod KG, McCaig A, Sewell JM, Gullick WJ, Smyth JF, Langdon SP. Neuregulin expression, function, and signaling in human ovarian cancer cells. Clin Cancer Res. 2002;8(12):3933-42.
78. Tanner B, Hasenclever D, Stern K, Schormann W, Bezler M, Hermes M, Brulport M, Bauer A, Schiffer IB, Gebhard S, Schmidt M, Steiner E, Sehouli J, Edelmann J, Läuter J, Lessig R, et al. ErbB-3 predicts survival in ovarian cancer. J Clin Oncol. 2006;24(26):4317-23.
79. Sheng Q, Liu X, Fleming E, Yuan K, Piao H, Chen J, Moustafa Z, Thomas RK, Greulich H, Schinzel A, Zaghlul S, Batt D, Ettenberg S, Meyerson M, Schoeberl B, Kung AL, et al. An activated ErbB3/NRG1 autocrine loop supports in vivo proliferation in ovarian cancer cells. Cancer Cell. 2010;17(3):298-310.
80. Lee H, Maihle NJ. Isolation and characterization of four alternate c-erbB3 transcripts expressed in ovarian carcinoma-derived cell lines and normal human tissues. Oncogene. 1998;16(25):3243-52.
81. Schoeberl B, Pace EA, Fitzgerald JB, Harms BD, Xu L, Nie L, Linggi B, Kalra A, Paragas V, Bukhalid R, Grantcharova V, Kohli N, West KA, Leszczyniecka M, Feldhaus MJ, Kudla AJ, et al. Therapeutically targeting ErbB3: a key node in ligand-induced activation of the ErbB receptor-PI3K axis. Sci Signal. 2009;2(77):ra31.
82. Schoeberl B, Faber AC, Li D, Liang MC, Crosby K, Onsum M, Burenkova O, Pace E, Walton Z, Nie L, Fulgham A, Song Y, Nielsen UB, Engelman JA, Wong KK. An ErbB3 antibody, MM-121, is active in cancers with ligand-dependent activation. Cancer Res. 2010;70(6):2485-94.
83. AACR Meeting Abstr 2008: LB-20, LB-21
84. ASCO Meeting Abstr 2011: 3026
85. Aurisicchio L, Marra E, Luberto L, Carlomosti F, De Vitis C, Noto A, Gunes Z, Roscilli G, Mesiti G, Mancini R, Alimandi M, Ciliberto G. Novel anti-ErbB3 monoclonal antibodies show therapeutic efficacy in xenografted and spontaneous mouse tumors. J Cell Physiol 2011. doi: 10.1002/jcp.24037.
86. Belleudi F, Marra E, Mazzetta F, Fattore L, Giovagnoli MR, Mancini R, Aurisicchio L, Torrisi MR, Ciliberto G. Monoclonal antibody-induced ErbB3 receptor internalization and degradation inhibits growth and migration of human melanoma cells. Cell Cycle 2012;11(7):1455-67.
87. Ben-Kasus T, Schechter B, Lavi S, Yarden Y, Sela M. Persistent elimination of ErbB-2/HER2-overexpressing tumors using combinations of monoclonal antibodies: relevance of receptor endocytosis. Proc Natl Acad Sci 2009;106(9):3294-9.
88. AACR Meeting Abstr 2012: 2508
89. AACR Meeting Abstr 2012: 2509
90. Sala G, Traini S, D’Egidio M, Vianale G, Rossi C, Piccolo E, Lattanzio R, Piantelli M, Tinari N, Natali PG, Muraro R, Iacobelli S; Consorzio Interuniversitario Nazionale per la Bio-Oncologia. An ErbB-3 antibody, MP-RM-1, inhibits tumor growth by blocking ligand-dependent and independent activation of ErbB-3/Akt signaling. Oncogene 2012;31(10):1275-86. doi: 10.1038/onc.2011.322.
91. AACR Meeting Abstr 2012: 2733
92. AACR Meeting Abstr 2012: 2718
93. haefer G, Haber L, Crocker LM, Shia S, Shao L, Dowbenko D, Totpal K, Wong A, Lee CV, Stawicki S, Clark R, Fields C, Lewis Phillips GD, Prell RA, Danilenko DM, Franke Y, et al. A two-in-one antibody against HER3 and EGFR has superior inhibitory activity compared with monospecific antibodies. Cancer Cell 2011;20(4):472-86.
94. Kamath AV, Lu D, Gupta P, Jin D, Xiang H, Wong A, Leddy C, Crocker L, Schaefer G, Sliwkowski MX, Damico-Beyer LA. Preclinical pharmacokinetics of MEHD7945A, a novel EGFR/HER3 dual-action antibody, and prediction of its human pharmacokinetics and efficacious clinical dose. Cancer Chemother Pharmacol. 2012;69(4):1063-9.
95. McDonagh CF, Huhalov A, Harms BD, Adams S, Paragas V, Oyama S, Zhang B, Luus L, Overland R, Nguyen S, Gu J, Kohli N, Wallace M, Feldhaus MJ, Kudla AJ, Schoeberl B, et al. Antitumor activity of a novel bispecific antibody that targets the ErbB2/ErbB3 oncogenic unit and inhibits heregulin-induced activation of ErbB3. Mol Cancer Ther. 2012;11(3):582-93.
96. AACR Meeting Abstr 2012: 2719.
97. Faber AC, Wong KK, Engelman JA. Differences underlying EGFR and HER2 oncogene addiction. Cell Cycle. 2010 Mar 1;9(5):851-2.