
INTRODUCTION
Colorectal cancer (CRC) is one of the leading causes of cancer-related human morbidity and mortality worldwide [1]. Despite recent advances in the diagnosis and therapy of CRC, the general survival rate of CRC patients has not improved [2]. Metastasis disposes patients to poor prognosis and is the main cause of mortality [3]. The molecular mechanisms underlying CRC metastasis are not quite clear to date.
Serotonin (5-hydroxytryptamine, 5-HT) is a neurotransmitter that mediates a wide variety of physiological events including central and peripheral action through the binding of multiple receptor subtypes [4]. Although the mitogenic events of serotonin receptors, especially 5-HT1A receptor, 5-HT1B receptor, 5-HT1D receptor and 5-HT2A receptor, in breast cancer, prostate cancer and bladder cancer have been investigated [5–7], there is a lack of evidence regarding whether these receptors play a significant role in CRC.
It has been found that the activation of Wnt signaling in colorectal cancer correlates with more invasive tumor growth, a higher susceptibility of disease recurrence after surgery, and a lower survival rate [8]. However, the underlying molecular mechanism of Wnt signaling is not clear. Recently, a report has shown that 5-HT1B receptors on osteoblasts inhibit cell proliferation by activating PKA and CREB in Wnt signaling [9]. Although there is a great contribution of 5-HT1 group receptors in signaling pathway including Wnt signaling [10], there is a lack of evidence of Wnt signaling on the impact of 5-HT receptors in colorectal cancers.
In this study, we first clarified the molecular mechanism by which 5-HT1D receptor (5-HT1DR) inhibits Wnt signaling, then we disclosed the direct effect of 5-HT1DR on Axin1. We demonstrated that inhibition of 5-HT1DR has potent anti-metastatic effect via Wnt signaling pathway. Our results provided the Wnt targeted mechanism to better understand CRC metastasis.
RESULTS
5-HT1DR is overexpressed in CRC tumors
To study the expression pattern of HTR in colorectal cancer, the protein level of 5-HT1R, 5-HT2R, 5-HT3R and 5-HT7R were quantified by immunohistochemical staining in 90 pairs of colorectal cancers and matched adjacent normal colon tissue samples. Upregulation of 5-HT1DR, 5-HT3CR and 5-HT4R protein level was observed in colorectal cancer samples compared with the paired normal tissues (Figure 1A). Notably, the expression of 5-HT1DR (68 out of 90, 76%) was observed being the highest among all 5-HT receptors. Then we divided the patients into two groups: high level of 5-HT1DR (68) and low level of 5-HT1DR (22). Statistical analysis indicated that patients with high level of 5-HT1DR had significantly worse overall survival (OS) and disease-free survival (DFS) compared with patients with low level of 5-HT1DR (Figure 1B). This was not associated with sex, age, tumor grade, carcino-embryonic antigen (CFA) and fetoprotein (AFP) level (Supplementary Table S1). The 5-year OS was 48.8% for high level of 5-HT1DR and 64.7% for low level of 5-HT1DR patients (p = 0.0023). The 5-year PFS was 39.9% for high level of 5-HT1DR and 56.2% for low level of 5-HT1DR patients (p = 0.0038).
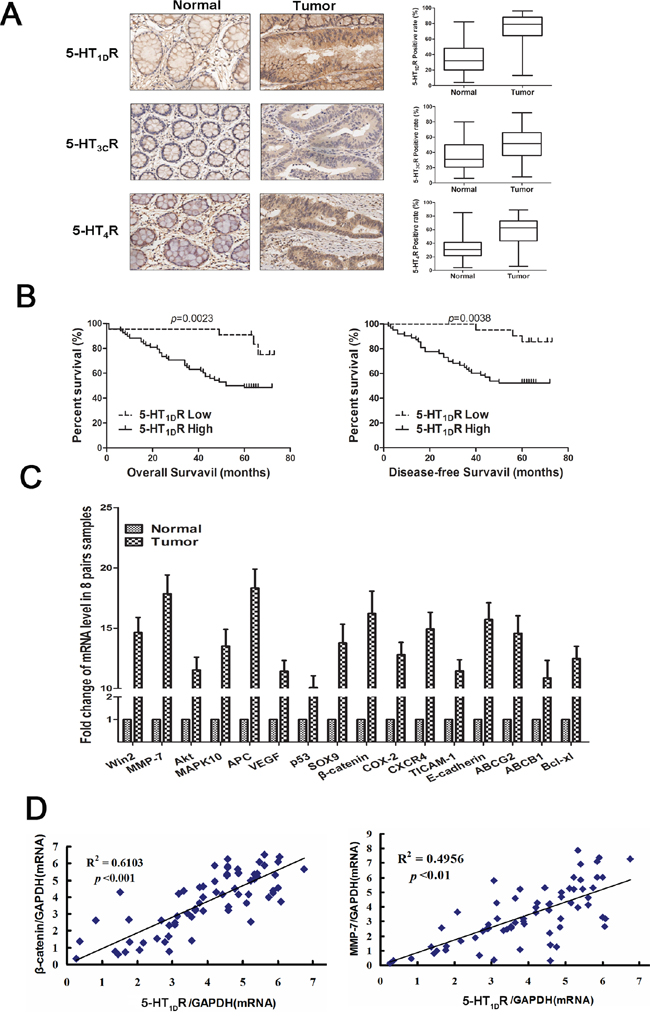
Figure 1: The expression of 5-HT1DR positively correlated with β-catenin and MMP-7 expression in human colorectal cancer tissues. A. Left: The expression of 5-HT1DR, 5-HT3CR and 5-HT4R protein was evaluated by immunohistochemistry. Right: 5-HT1DR, 5-HT3CR and 5-HT4R protein expression quantification exemplified in 90 primary CRC tissue samples and matched with normal tissue. B. OS and DFS curves for all studied patients with high or low 5-HT1DR expression (n = 90). C. Full cDNA microarray analysis is performed to detect the different expression of cancer genes in 8 pairs of patient-matched normal tissues with a positive 5-HT1DR expression. D. Left: Correlation analysis between the relative mRNAs of 5-HT1DR and β-catenin in 5-HT1DR positive human colorectal tumors (n = 68). Right: Correlation analysis between the relative mRNAs of 5-HT1DR and MMP-7in 5-HT1DR positive human colorectal tumors (n = 68). (Spearman correlation test)
To further characterize the role of 5-HT1DR overexpression in colorectal cancer, we performed a full cDNA microarray to screen for different expression of cancer genes in 7 pairs of patient-matched normal tissues (data not shown). As viewed in Figure 1C and Supplementary Figure S1, a total of 16 genes were differentially upregulated (by more than 10 times), and involved seven signal pathways. Interestingly, components of the Wnt signaling pathway, including MMP-7, β-catenin, and APC, were significantly upregulated in high 5-HT1D R tumor tissue, implying a possible link between 5-HT1D R and Wnt signaling pathway in CRC patients.
Importantly, in 68 CRC patients with high level of 5-HT1DR, a positive and significant association between 5-HT1DR and c-myc or MMP-7 gene was observed (Figure 1D), whereas no such correlation was seen in the other 14 genes. MMP-7 is a known downstream target of Wnt signaling pathway and protein MMP-7 can be upregulated when the Wnt signaling pathway are activated. Collectively, our results suggest a possible link between 5-HT1DR upregulation and Wnt/MMP-7 signaling pathway in CRC progression.
5-HT1DR competitively bound to Axin1 and released Axin1 from the destruction complex
Western blotting analysis revealed that the family members of 5-HTR protein were differentially expressed in 4 human CRC cells (LoVo, HCT-116, HT-29 and SW403) (Figure 2A). As we know, LoVo cells are a well differentiated cells. Interestingly, 5-HT1DR, 5-HT1BR and 5-HT1FR were more overexpressed in LoVo cells than that in HCT-116 cells, which are poor differentiated cells. Since previous evidence indicated that targeting 5-HT1D receptor subsequently targets Wnt pathway, we investigated which factor is involved in 5-HT1DR regulating Wnt pathway. First, using sumatriptan to increase the level of 5-HT1DR, we found the expression of 5-HT1DR in HCT-116 was upregulated, whereas 5-HT1DR protein was downregulated in LoVo cells in a dose-dependent manner after treatment with GR127935 (Figure 2B). Second, to verify the effect of 5-HT1DR on canonical Wnt/β-catenin signaling pathway, other components of the “destruction complex” including APC, Axin1, GSK3β and CKIα were tested. As shown in Figure 2C, when sumatriptan was applied, the Axin1 level was markedly decreased in HCT-116 cells, but increased in LoVo cells (Figure 2C and right-upper). Consistent with our findings in Axin1 expression, β-catenin protein showed a decreased expression in the cytoplasm of HCT-116 cells, but an increased expression in the HCT-116 nucleus in a dose-dependent manner (Figure 2C). These findings suggest that 5-HT1DR is a gene involved in the response to nuclear accumulation of β-catenin in activation of the Wnt/β-catenin pathway. As protein interaction is the first step for epigenetic regulation, we then checked whether 5-HT1DR is associated with Axin1 by ChIP analysis. Among the three members APC, Axin1, GSK3β and CKIα of the destruction complex family, only Axin1 co-precipitated with 5-HT1DR in 5-HT1DR overexpression LoVo colorectal cancer cells (Figure 2D).
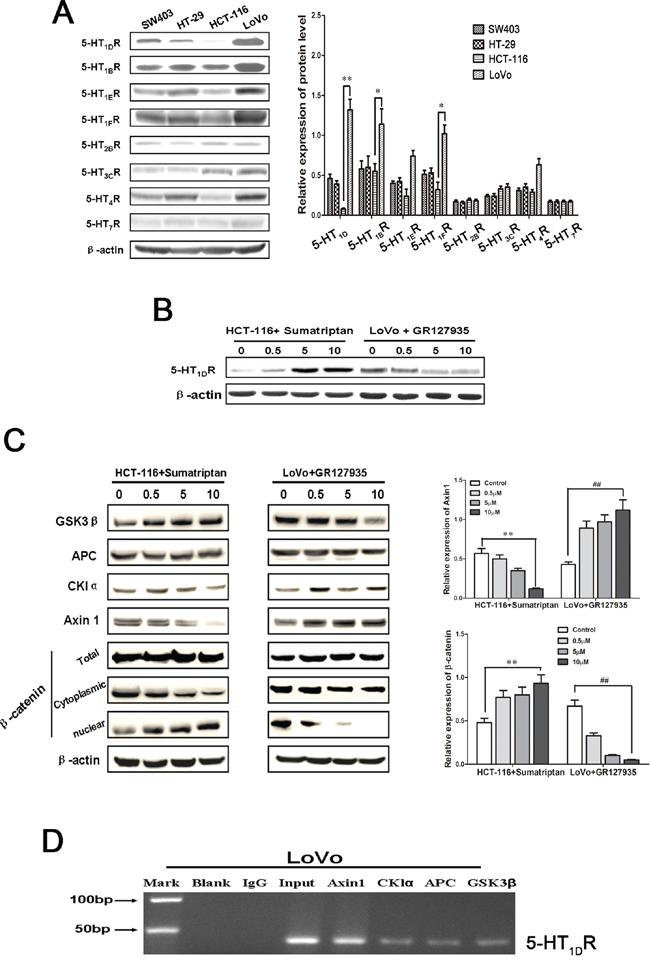
Figure 2: 5-HT1DR competitively bound to Axin1 and regulated pivotal signaling transduction pathways. A. Left: The expression of 5-HTR family members in 4 human CRC cells (LoVo, HCT-116, HT-29 and SW403). Right: Quantification of the relative protein expression. Values are mean ± SEM. (significant differences are indicated by *P < 0.05, **P < 0.01). B. Western blot analysis of 5-HT1DR expression levels in HCT-116 and LoVo cells treated with 5-HT1DR agonist (sumatriptan) and antagonist (GR127935) for 48 h. C. Left: Western blot analysis of APC, Axin1, GSK3β, CKIα and β-catenin expression after HCT-116 and LoVo cells were treated with sumatriptan and GR127935 for 48 h. Right: Quantification of the relative Axin1 and β-catenin expression. Values are mean ± SEM. (significant differences are indicated by **P < 0.01, ##P < 0.01). D. ChIP analysis between 5-HT1DR protein and APC, Axin1, GSK3β, CKIα gene. As a control, mouse monoclonal IgG was used. The blank group was the PCR results with no cDNA, and the input group was the cDNA from cell lysates without RIP procedure. The mean and standard errors from triplicate experiments are indicated.
5-HT1DR regulated Axin1/β-catenin/LEF1/TCF4/MMP-7 signaling
Since LEF1/TCF4 is one of the major downstream of Wnt/β-catenin pathway, we hypothesized that 5-HT1DR may utilize Axin1/β-catenin/LEF/TCF4 to influence its metastatic effects. We evaluated the level of LEF/TCF4 in LoVo cells with or without sumatriptan treatment after transfected with Axin1-overexpressing vector. As expected, the level of LEF/TCF4 was significantly upregulated in Axin1-overexpressing LoVo cells compared to that of LoVo cells transfected with empty vector (Figure 3A). Importantly, the combination of sumatriptan and Axin1-overexpressing vector offset the inhibitory effect of Axin1, suggesting that 5-HT1DR regulates the expression of Wnt/β-catenin/LEF/TCF4 specifically through Axin1 separated from the destruction complex. To further clarify the molecular mechanisms of the 5-HT1DR activation of the Axin1/β-catenin/LEF/TCF4 pathway, we compared the expression profiles of Wnt-related molecules in LoVo cells including MMP-7, cyclin D1, c-Myc, MMP-2 and MMP-9. Western blot result showed that only expression of MMP-7 changed with a similar trend as the level of LEF1/TCF4 (Figure 3B). The findings from this study revealed an inherent role of 5-HT1DR which binds Axin1 and dissociates β-catenin from the complex, moving β-catenin into the nucleus to activate the β-catenin/LEF1/TCF4/MMP-7 pathway in colorectal cancer metastasis (Figure 3C).
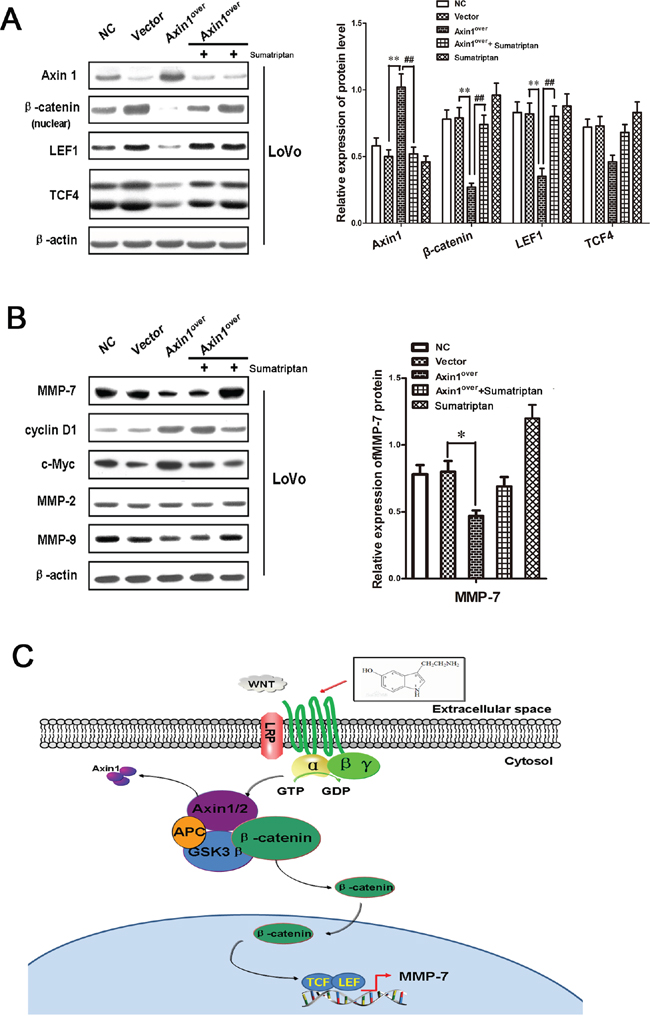
Figure 3: 5-HT1DR regulated Axin1/β-catenin/LEF1/TCF4/MMP-7 signaling pathway. A & B. Western blots showing expressions of Axin1, β-catenin, LEF, TCF4, MMP-7, cyclin D1, c-Myc, MMP-2 and MMP-9 in response to transfection with lentivirus-control (Vector), Axin1 lentivirus (Axin1over), 5-HT1D agonist or co-transfection with sumatriptan and Axin1 lentivirus (Axin1over) for 48 h as described in Materials. Quantification of the relative Axin1, β-catenin, LEF, TCF4, MMP-7 expression. Values are means ± SEM. (significant differences are indicated by *P < 0.05, **P < 0.01). C. Proposed working model of the noted 5-HT1DR lead of Axin1/β-catenin/LEF1/TCF4/MMP-7 signaling pathway mediated invasion and migration in colorectal cancer cells.
5-HT1DR modulates colorectal cancer invasion/migration in vitro
To determine whether the 5-HT1DR may play a role in tumor invasion, GR127935 was applied to LoVo cells and analyzed using a scratch-wound motility assay. The data in Figure 4A showed a time and dose-dependent downregulation of cell migration in response to GR127935 treatment, as shown by as much as 40% delay in wound closure at 48 hours post-treatment with the highest dose of GR127935 (Figure 4A right). Similar results were seen in Matrigel-coated Transwell assays, in which LoVo cells were treated with GR127935 in a dose-dependent manner (Figure 4B). In addition to the offset effect of sumatriptan to Axin1, we also elucidate the role of sumatriptan in Axin1-mediated cell migration. Transfection of Axin1-overexpressed vector and a specific sumatriptan dose were used to inhibit the activity of Axin1 mediated signaling pathway in LoVo cells. As expected, compared with vector cells, Axin1-overexpressing cells showed more migration activity; however the activity was decreased after adding sumatriptan (Figure 4C). Finally, we evaluated cell viability after GR127935 treatment with the trypan blue exclusion assay. As shown in Figure 4D and 4E, the proliferation and migration of LoVo cells remained unchanged, implying that 5-HT1DR plays a key role in the regulatory effect of Axin1-mediated cell migration without the proliferation change.
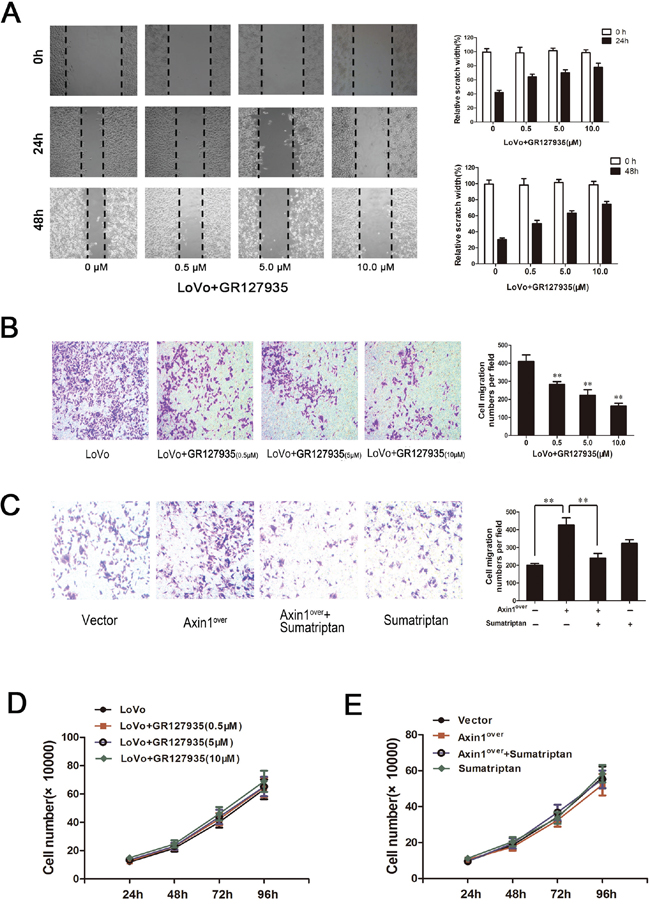
Figure 4: 5-HT1DR modulated invasion and migration in colorectal cancer cell. A & D. Wound-healing assay. Images were taken 0, 24 and 48 hours after wound formation. Data are presented as mean ± SD of triplicate experiments.*P < 0.05, **P < 0.01 ##P < 0.01 in Fig. 4D. B & C. Cell invasion assay result using Matrigel-coated Transwell. (left, representative pictures of invasion chambers; right, average counts from five random microscopic fields). Data are presented as mean ± SD of triplicate experiments. *P < 0.05 vs.Vector group in Fig. 4A; *P < 0.05, **P < 0.01 in Fig. 4B. D. The cell proliferation of GR127935-treated LoVo cells in different concentration at 24 h, 48 h, 72 h, 96 h as described in Materials. E. Cell proliferation was assayed at 24, 48, 72 and 96 hours after LoVo cells were transfected with Axin1 lentivirus (Axin1over) or treated with sumatriptan or treated with both together.
5-HT1DR modulates colorectal cancer invasion/migration in vivo
To further test our hypothesis of 5-HT1DR’s role in migration and invasion in vivo, we first established a subcutaneous xenograft tumor model of colorectal cancer cell line. Compared with normal saline group, the tumor growths were not apparent after treatment with GR127935 (Figure 5A). The toxicity of GR127935 was evaluated by observing the body weight, and no significant weight loss was observed (Supplementary Figure S2). However, immunohistochemistry analyses of Axin1 displayed a downregulation in the tumor of mice treated with GR127935 compared with that of the control group. Similar to the result in vitro, the levels of β-catenin, LEF1, TCF4 and MMP-7 increased after treatment with GR127935 compared with those of the control group (Figure 5B). Next, we generated a xenograft tumor model using fluorescence-labeled Axin1-overexpressed colorectal cancer cell. When we treated these tumor-bearing mice with sumatriptan, the fluorescence activity of Axin1 decreased in a dose-dependent manner (Figure 5C). We also used fluorescently-labeled Axin1-overexpressed cells in a colonic orthotopic xenograft. After sumatriptan treatment for 28 days, the fluorescent activity of Axin1 was significantly attenuated compared with that of the control group (Figure 5D). Furthermore, we examined the invasion and migration by hematoxylin and eosin (H&E) staining in main organ tissue slices. We found more colonic metastases in liver and lung tissue in Axin1-overexpressed group, which were decreased in Axin1-overespressed group treated with sumatriptan (Figure 5E). Corresponding survival curves showed a significantly increase in median survival of sumatriptan-treated group. We also saw that the luc-vector group had significantly prolonged overall survival than the Axin1-overexpressed group with or without sumatriptan (Figure 5F).
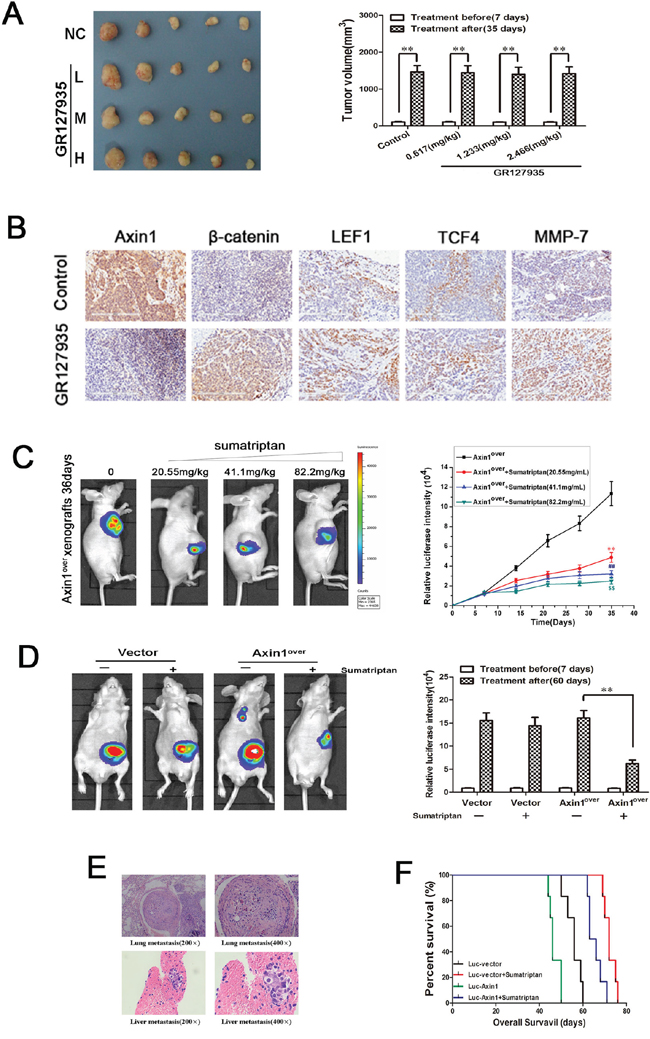
Figure 5: 5-HT1DR modulated colorectal cancer invasion/migration in vivo. A. Excised tumors on day 28 of treatment with GR127935 (35 days after inoculation). Tumors were weighed (right) and photographed (left). Data are means ± SEM. Significant differences are indicated by **P < 0.01 (representative one in each group has been shown). B. The subcutaneous transplanted tumor tissue in Fig. 5A were subjected to immunohistochemical analysis using Axin1, β-catenin, LEF1, TCF4 and MMP-7 antibody. Images were taken at 200x magnification. Brown staining indicated positive cells. C. Left: Luciferase imaging of the mice with Axin1 overexpression and sumatriptan xenografts on day 36 after tumor cell implantation. Right: the quantification of luciferase intensities in tumors of the four groups. Data are presented as mean ± SD of triplicate experiments. **P < 0.01 sumatriptan (0.617 mg/kg) vs. Control. ##P < 0.01 sumatriptan (1.233 mg/kg) vs. Control. $$P < 0.01 sumatriptan (2.466 mg/kg) vs.Control. D. Luciferase imaging revealed primary tumor growth and its distant metastasis. Right: the quantification of luciferase intensities of the mice treated with sumatriptan for 60 days. Data are means ± SD. significant differences are indicated by **P < 0.01. E. H&E analyses of lung and liver sections in Axin1 overexpressing orthotopic implanted animals by day 60, which was the only group with distant metastasis. F. overall survival of orthotopic model is shown in Fig. 5D.
5-HT1DR modulates colorectal cancer invasion/migration in IECs
Because increased motility and invasion are among the principal effects of upregulated Axin1/β-catenin signaling, those features were tested in 5-HT1DR overexpressing colorectal cancer cells. To test whether 5-HT1DR also regulate motility and invasion via Axin1/β-catenin signaling in intestinal epithelium cells, we use Caco-2 cell as intestinal epithelium cells, and measured the level of 5-HT1DR with or without sumatriptan and GR127935. Western blot results showed a low level of 5-HT1DR in Caco-2 cell, and sumatriptan or GR127935 could increase or decrease the level of 5-HT1DR respectively (Figure 6A). Next, we assessed its toxicity on in Caco-2 cells. As shown in Figure 6B, sumatriptan or GR127935 treatments do not significantly inhibit cell growth or slow the rate of Caco-2 cell cycle (Figure 6C). To investigate the potential mechanism of Caco-2 cells and LoVo cells, we used PCR to examine the expression of mRNA in 19 genes including their difference in the tumor sample previously found and Axin1, LEF1, TCF4. As shown in Figure 6D, Axin1, β-catenin, LEF, TCF4 and MMP-7 were downregulated in Caco-2 cells. Furthermore, we also tested the effect of sumatriptan in the Axin1/β-catenin-related signal pathway, which has been verified in LoVo cells. Figure 6E and 6F showed that 5-HT1DR may utilize Axin1/β-catenin/LEF/TCF4 to influence its metastatic effects.
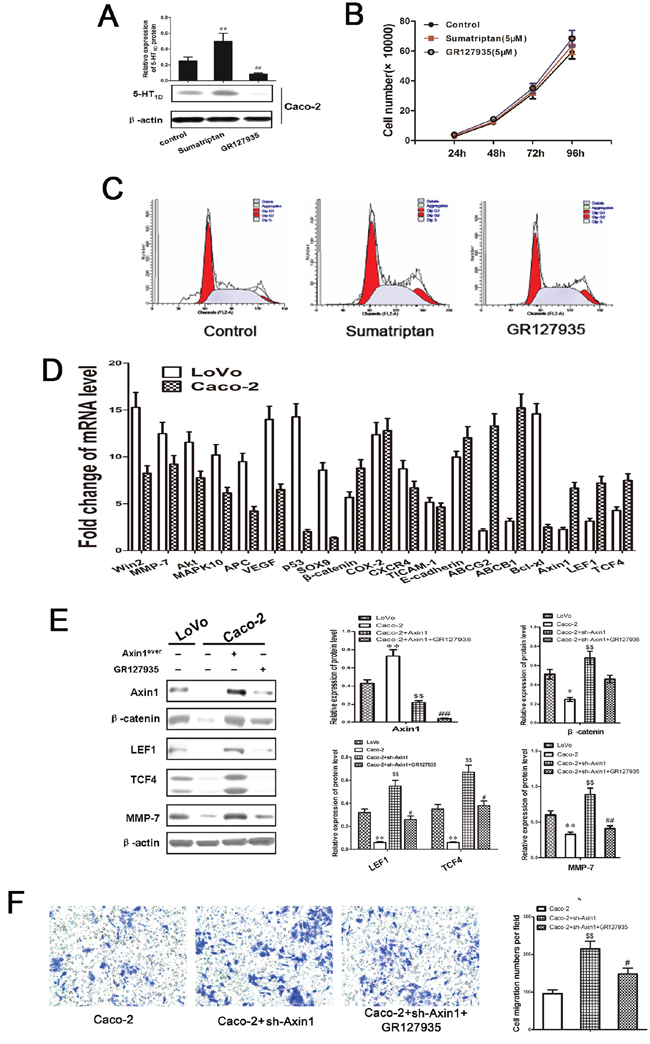
Figure 6: 5-HT1DR modulated colorectal cancer invasion/migration in IECs. A. Relative expression levels of 5-HT1DR were detected by Western blot in Caco-2 cells treated with sumatriptan and GR127935. B. The cell proliferation of sumatriptan and GR127935 treated Caco-2 cells in different concentrations at 24 h, 48 h, 72 h, and 96 h. C. Flow cytometry analysis was used to distinguish cells in different phases of the Caco-2 cell cycle as described in Methods. D. The mRNA levels of 16 genes were quantified by qRT-PCR in LoVo and Caco-2 cell. E. Western blots showing expression levels of Axin1, β-catenin, LEF, TCF4, and MMP-7 in LoVo and Caco-2 cell infected by Axin1 lentivirus (Axin1over), co-treated with GR127935 and Axin1 lentivirus (Axin1over) for 48 h as described in Materials. Data are presented as mean ± SD of triplicate experiments. **P < 0.01 Caco-2 vs. LoVo; $$P < 0.01 Caco-2+Axin1over vs. Caco-2; ##P < 0.01 Caco-2+Axin1over+GR127935 vs. Caco-2+Axin1over. F. Caco-2 cell invasion assay result using Matrigel-coated Transwell. The bold horizontal bar represents mean expression levels; Data are presented as mean ± SD of triplicate experiments. $$P < 0.01 Caco-2+Axin1over vs. Caco-2; #P < 0.05 Caco-2+Axin1over+GR127935 vs. Caco-2+Axin1over
DISCUSSION
Cancer invasion and metastasis are complex processes involving increase in cell motility, adhesion to underlying stroma, digestion of extracellular matrix, vascular invasion, evasion of immune attack, and growth in foreign environments [17–19]. One way to identify the genes involved in these processes is to compare the gene expression profile between normal and tumor tissue. In the present study, 5-HT1DR was shown to be a significantly overexpressed in the human colorectal cancer sample (76%), which was assumed to drive poor outcome in large patient cohorts. Moreover, we found 16 genes upregulated in 7 pairs of 5-HT1DR overexpressing tumor compared to their normal tissues by genome-wide microarray analysis. Similar to studies of hepatocellular carcinoma [20], approximately 30.8% of all patients showed high expression of CTHRC1 and this was associated with a lower 10-year overall and disease-free survival rates. Collectively, our results suggest that 5-HT1DR is a promising target in clinical therapy of patients with advanced CRC. Unfortunately, the molecular mechanisms underlying 5-HT1DR in CRC metastasis are still unclear.
Wnt pathway is known as a “destruction complex” in which Axin and adenomatous polypopsis coli (APC) form a scaffold facilitating β-catenin phosphorylated by CK1a and GSK3β at serine and threonine residues in its amino-terminal region [21, 22]. In a normal steady state, the cells are not exposed to Wnt signaling [23, 24]. However, activation of Wnt/β-catenin signal transduction pathway is thought to lead to a higher susceptibility of disease recurrence after surgery and a lower survival rate [25, 26]. Its mechanism is illustrated as follows: Axin dissociates from the APC/Axin/GSK/β-catenin-complex, then β-catenin translocates to the nucleus where it binds to T-cell factors (Tcf) and activates the transcription of specific target genes, including c-Myc, cyclin D1 and MMP7 [27, 28].
Because of the demonstrated relationship between 5-HT1DR, Wnt/β-catenin and MMP-7 from clinical data analysis, we hypothesized that 5-HT1DR may be involved in Wnt/β-catenin/MMP-7 mediated CRC metastasis. We increased 5-HT1DR expression in HCT-116 cell using an agonist, and revealed a concomitant reduction in Axin1 protein level and a concomitant increase of β-catenin protein level in the nucleus. On the contrary, GR127935 had the opposite effect in LoVo cells. Previous studies have illustrated the separation of “destruction complex” leading to an increased in Axin and [β-catenin/TCF] signaling in the cellular environment [29, 30]. Although APC and GSK3β are reported to be components of the “destruction complex”, we did not find significant changes in the expression levels of APC or GSK3β after the two colorectal cancer cells treatment with sumatriptan or GR127935. Therefore, it seems that highly-differentiated cells have an intrinsic mechanism to target Axin1 and activate β-catenin/TCF signaling pathway [31], which is still maintained when the overall level of 5-HT1DR is elevated in cancer cells. Furthermore, the ChIP assay also confirmed that the 5-HT1DR protein could bind to the Axin1 gene promoter in LoVo cells, but not to APC or GSK3β. Additionally, the results also showed that 5-HT1DR did not affect the other gene, only changed the level of MMP-7. CTHRC1 was also reported to be an activator of the planar cell polarity pathway of Wnt signaling in mouse embryogenesis [32]. In this work, we also provide some insights into the biological effects of overexpression of 5-HT1DR in CRC. Our in vitro data further showed that GR127935 functions as a tumor suppressor via Axin1/β-catenin/MMP-7 pathway in the cell cycle, proliferation, and invasion. This is the first work that we know of to use a mouse model system of Axin1-overexpression on the primary colon tumors to identify the effect of 5-HT1DR in metastasis.
There are indications that some additional genetic or epigenetic alterations are required in normal cells, thereby leading to a gain-of-function phenotype. In agreement with this possibility, our results in the normal primary colon cell, as well as studies by others report, show that the activation of Axin1/β-catenin/MMP-7 pathway is acquired only after 5-HT1DR activation.
We reported in our study that 5-HT1DR activated Axin1/β-catenin/MMP-7-mediated signaling by targeting Axin1 directly induces potent invasion and migration activity both in vitro and in vivo. The correlation between 5-HT1DR expression and Axin1 further provided a mechanism-based rationale to target Axin1. In conclusion, we suggest that Axin1/β-catenin/MMP-7 may be a useful prognostic marker for CRC progression and in determining the sensitivity of 5-HT1DR as a potential target for patients with CRC.
MATERIALS AND METHODS
Cell culture and reagents
The human colorectal cancer LoVo, HCT-116, HT-29 and SW403 cell lines were purchased from the Shanghai Cell Collection (Shanghai, China). Cells were grown in RPMI 1640 medium supplemented with 10% (v/v) heat-inactivated fetal calf serum, 2 mM glutamine, 100 units/ml penicillin, and 100 μg/ml streptomycin (Invitrogen, Carlsbad, CA) at 37°C in a 5% CO2-humidified atmosphere. Monoclonal antibodies against 5-HT family, Axin1, β-catenin, APC, GSK3β, CKIα, LEF1, TCF4, cyclin D1, c-Myc, MMP-2, MMP-9, MMP-7 and β-actin were products of Cell Signaling Technology (Beverly, MA, USA). 5-HT1DR agonist (Sumatriptan) and 5-HT1DR antagonist (GR127935) were from Sigma Chemical Co. (St.Louis, MO).
Clinical samples
From 2008 to 2014, unifocal, primary CRC biopsies surgically resected from 90 patients, who received detailed pathological assessment and regular follow-up at Shuguang Hospital Shanghai University of Traditional Chinese Medicine were collected for this study. The basic clinical characteristics of the 90 patients are presented in Supplementary Table S1. All of the donors or their guardians provided written consent and ethics permission was obtained for the use of all samples. This study was approved by the Medical Ethics and Human Clinical Trial Committee of the affiliated hospitals, Shanghai University of Traditional Chinese Medicine.
Immunohistochemical analysis
The hydrated paraffin section were incubated in a blocking solution (10% donkey serum +5% nonfat dry milk +4% BSA +0.1% Triton X-100) for 10 min, and then incubated at 4°C overnight with anti-5-HT family antibody. After washing with PBS, the sections were incubated with diluted (1:200) biotinylated secondary antibody for 30 min. Subsequently, the sections were washed again in PBS and incubated for 30 min with the preformed avidin-horseradish peroxidase macromolecular complex. Development of peroxidase reaction was achieved by incubation in 0.01% 3, 3-diaminobenzidine tetrahydrochloride (DAB) in PBS containing 0.01% hydrogen peroxide for approximately 5 min at room temperature. Sections were then washed thoroughly in tap water, counterstained in haematoxylin, dehydrated in absolute alcohol, cleared in xylene and mounted in synthetic resin for microscopic examination.
Transfection of lentivirus
Transfection procedures were performed according to manufacturers’ instructions, with Lipofectamin 2000 as transfection reagent (Invitrogen). Axin1-GFP/luciferase lentivirus and control GFP/luciferase Lentivirus were used to transfect LoVo cells. In brief, 2 × 105 cells were plated in each well of a 24-well plate and incubated overnight. Lentiviral supernatants were added to cells according the concentration of lentivirus for 12 h. The GFP positive cells were sorted and the stable clones were isolated.
Analysis of gene expression
Total RNAs were reverse-transcribed to cDNA using the Advantage RT-PCR Kit (Clontech Laboratories, Inc., MountainView, CA, USA). For quantitative real-time PCR (qRT-PCR), target gene expression was normalized to glyceraldehyde-3-phosphate dehydrogenase expression. Primers are described in online Supplementary Table S2.
Western blot analysis
Whole cell lysates for Western blot analysis of 5-HTR family, Axin1, β-catenin, APC, GSK3β, CKIα, LEF1, TCF4, cyclin D1, c-Myc, MMP-2, MMP-9, MMP-7 and β-actin expression were prepared as previously reported [11]. Briefly, the cells were lysed on ice in immunoprecipitation assay buffer for 2 h before being homogenized using a mortar and pestle. The homogenized sample was centrifuged, and the supernatant was collected and stored at −80°C until used. Densitometric analysis was performed using the Scion Imaging software (Scion Corporation), with β-actin as internal reference.
Immunoprecipitation
Immunoprecipitation was performed as described previously [11], except N-ethylmaleimide was added to the cell lysis buffer at a final concentration of 10 mM to preserve poly-ubiquitinated protein conjugates.
Cell viability assay
Cells were stained with 2 mg/ml propidium iodide as described previously [12]. Events in live cell gate were counted by an FACS Calibur. Ratio of live to total events was calculated as percent viability. All experiments were done with 5 replicates per experiment and repeated at least 3 times.
Cell cycle analysis
Cells were treated with 100 nM Dasatinib or DMSO for 24–72 h. Cell cycle was determined using the FITC BrdU Flow kit (BD Biosciences, San Diego, CA, USA) following the manufacturer’s instructions [13]. Subsequently, cells were washed once with PBS and analyzed by FACS. The percentage of cells in various phases of the cell cycle, namely, G1, G2, and S, were analyzed using ModFit software.
Cell invasion and wound healing assays
The Matrigel invasion assay was done using the BD Biocoat Matrigel Invasion Chamber (pore size: 8 mm, 24-well; BD Biosciences, USA) following the manufacturer’s protocol [14]. From five randomly selected fields, the invading cells were counted under a light microscope. For wound-healing assay, cell monolayers were scratched with a clean pipette tip and cell migration was observed for up to 24 h.
Animals and xenograft models
Male athymic nude mice (NCr-nu), 8–12 weeks old, were purchased from Sino-British SIPPR/BK lab Animal Co., Ltd (Shanghai, China, license No. SCXK 2008-0016), and maintained under specific-pathogen-free conditions. All animal protocols were approved by the Institutional Animal Use and Care Committee. All the experiments and animal care were approved by Shanghai Medical Experimental Animal Care Commission and in accordance with the Provision and General Recommendation of Chinese Experimental Animals Administration Legislation
For experiment 1, 24 mice were subcutaneously injected with 1.0 × 106 LoVo cells per animal. When the tumors reach an average size of 100 mm3, the mice were randomized into 4 groups (n = 6 per group) and received intragastric administration of 5-HT1DR antagonist (GR127935) at the doses of 0 mg/kg, 0.617 mg/kg, 1.233 mg/kg and 2.466 mg/kg every day as previously described [15]
For experiment 2, 24 mice were subcutaneously injected with Axin1-overexpressed luciferase LoVo cells per animal. When the tumors reach an average size of 100 mm3, the mice were randomized into 4 groups (n = 12 per group) and received intragastric administration of sumatriptan at the doses of 20.55 mg/kg, 41.1 mg/kg and 82.2 mg/kg. In clinical practice, 5-HT1DR agonist (Sumatriptan) is usually prescribed at a maximum daily dose of 200 mg. When this human dose is converted into an animal dose (a person of 60 kg, and a conversion factor of 12.33 between human and mouse), it was equivalent to the middle dose (41.1 mg/kg) used in this study. Six mice were sacrificed in each group on the 28th day after treatment (35 day after transplantation), the other 6 mice in the same group were observed longer for survival time
For experiment 3, 24 mice underwent colon orthotopic transplantation with Axin1-overexpressed luciferase or vector luciferase LoVo cells per animal. The treatment was similar to experiment 2, except that the time of intragastric administration of sumatriptan started from 30 days after transplantation due to a prior observation that metastasis of colorectal cancer needs at least one month to take place [16]. Six mice were sacrificed in each group on the 60th day after transplantation, the other 6 mice in the same group were observed longer for survival time. The body weight of the animals and the two perpendicular diameters (A and B) were recorded every 3 days and tumor volume (V) was estimated according to the following formula [16] :V= π/6 × [(A+B)/2]3. The survival time for each group and overall significance was plotted on a Kaplan-Meier survival curve using GraphPad Prism
Bioluminescence imaging
Bioluminescence imaging and data acquisition were performed using D-luciferin potassium salt and the IVIS 100 imaging system coupled to the Living Image software (Xenogen) as previously reported [2].
H & E staining
The colon tissues containing primary tumors, liver and lungs harvested from CRC xenograft tumor group 5 and 6 mice were explanted, imaged, and immediately fixed in 10% neutral buffered formalin for 24 h. The tissues were then processed, embedded in paraffin, and sectioned for hematoxylin and eosin (H & E) staining.
ACKNOWLEDGMENTS AND GRANT SUPPORT
This work was supported by the National Natural Science Foundation of China (No.81202812, 81373862, 81473478), Program of Shanghai Municipal Education Commission (No. 12YZ058), and Shanghai Municipal Health Bureau (No. 2011ZJ030, 2010QL050B).
CONFLICTS OF INTEREST
All authors declare no conflicts of interest.
REFERENCES
1. Kang YJ, Park HJ, Chung HJ, Min HY, Park EJ, Lee MA, Shin Y, Lee SK. Wnt/β-catenin signaling mediates the antitumor activity of magnolol in colorectal cancer cells. MolPharmaco. 2012; 82:168–177.
2. Sui H, Cai GX, Pan SF, Deng WL, Wang YW, Chen ZS, Cai SJ, Zhu HR, Li Q. miR-200c attenuates P-gp mediated MDR and metastasis by targeting JNK2/c-Jun signaling pathway in colorectal cancer. Mol Cancer Ther. 2014; 13:3137–3151.
3. Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011; 331:1559–1564.
4. Jensen JB, du Jardin KG, Song D, Budac D, Smagin G, Sanchez C, Pehrson AL. Vortioxetine, but not escitalopram or duloxetine, reverses memory impairment induced by central 5-HT depletion in rats: evidence for direct 5-HT receptor modulation. EurNeuropsychopharmacol. 2014; 24:148–159.
5. Dizeyi N, Bjartell A, Nilsson E, Hansson J, Gadaleanu V, Cross N, Abrahamsson PA. Expression of serotonin receptors and role of serotonin in human prostate cancer tissue and cell lines. Prostate. 2004; 59:328–336.
6. Siddiqui EJ, Shabbir MA, Mikhailidis DP, Mumtaz FH, Thompson CS. The effect of serotonin and serotonin antagonists on bladder cancer cell proliferation. BJU Int. 2006; 97:634–639.
7. Sonier B, Arseneault M, Lavigne C, Ouellette RJ, Vaillancourt C. The 5-HT2A serotoninergic receptor is expressed in the MCF-7 human breast cancer cell line and reveals a mitogenic effect of serotonin. BiochemBiophys Res Commun. 2006; 343:1053–1059.
8. Coskun M, Olsen AK, Bzorek M, Holck S, Engel UH, Nielsen OH, Troelsen JT. Involvement of CDX2 in the cross talk between TNF-α and Wnt signaling pathway in the colon cancer cell line Caco-2. Carcinogenesis. 2014; 35:1185–1192.
9. Galli C, Macaluso G, Passeri G. a novel bone mass controller may have implications for alveolar bone. J Negat Results Biomed. 2013; 12:12.
10. Boudin E, Jennes K, de Freitas F, Tegay D, Mortier G, Van Hul W. No mutations in the serotonin related TPH1 and HTR1B genes in patients with monogenic sclerosing bone disorders. Bone. 2013; 55:52–56.
11. Sui H, Pan SF, Feng Y, Jin BH, Liu X, Zhou LH, Hou FG, Wang WH, Fu XL, Han ZF Ren JL, Shi XL, Zhu HR, Li Q. Zuo Jin Wan reverses P-gp-mediated drug-resistance by inhibiting activation of the PI3K/Akt/NF-κB pathway. BMC Complement Altern Med. 2014; 14:279.
12. Kawakami Y, Nagai N, Ohama K, Zeki K, Yoshida Y, Kuroda E, Yamashita U. Macrophage-colony stimulating factor inhibits the growth of human ovarian cancer cells in vitro. Eur J Cancer. 2000; 36:1991–1997.
13. Sui H, Liu X, Jin BH, Pan SF, Zhou LH, Yu NA, Wu J, Cai JF, Fan ZZ, Zhu HR, Li Q.Zuo Jin Wan, a Traditional Chinese Herbal Formula, Reverses P-gp-Mediated MDR In Vitro and In Vivo. Evid Based Complement Alternat Med. 2013; 2013:957078. doi: 10.1155/2013/957078.
14. Ji Q, Zhang L, Liu X, Zhou L, Wang W, Han Z, Sui H, Tang Y, Wang Y, Liu N, Ren J, Hou F, Li Q. Long non-coding RNA MALAT1 promotes tumour growth and metastasis in colorectal cancer through binding to SFPQ and releasing oncogene PTBP2 from SFPQ/PTBP2 complex. Br J Cancer. 2014; 111:736–748.
15. Keegan A, Morecroft I, Smillie D, Hicks MN, MacLean MR. Contribution of the 5-HT(1B) receptor to hypoxia-induced pulmonary hypertension: converging evidence using 5-HT(1B)-receptor knockout mice and the 5-HT(1B/1D)-receptor antagonist GR127935. Circ Res. 2001; 89:1231–1239.
16. Kruse J, von Bernstorff W, Evert K, Albers N, Hadlich S, Hagemann S, Günther C, van Rooijen N, Heidecke CD, Partecke LI. Macrophages promote tumour growth and liver metastasis in an orthotopic syngeneic mouse model of colon cancer. Int J Colorectal Dis. 2013; 28:1337–1349.
17. Wang H, Shi J, Luo Y, Liao Q, Niu Y, Zhang F, Shao Z, Ding Y, Zhao L. LIM and SH3 Protein 1 Induces TGFβ-Mediated Epithelial-Mesenchymal Transition in Human Colorectal Cancer by Regulating S100A4 Expression. Clin Cancer Res. 2014; 20:5835–5847.
18. Shi F, Shang L, Pan BQ, Wang XM, Jiang YY, Hao JJ, Zhang Y, Cai Y, Xu X, Zhan QM, Wang MR. Calreticulin promotes migration and invasion of esophageal cancer cells by up-regulating neuropilin-1 expression via STAT5A. Clin Cancer Res. 2014; 20:6153–662.
19. Yu J, Wu WK, Li X, He J, Li XX, Ng SS, Yu C, Gao Z, Yang J, Li M, Wang Q, Liang Q, Pan Y, et al. Novel recurrently mutated genes and a prognostic mutation signature in colorectal cancer. Gut. 2015; 64:636–645.
20. Chen YL, Wang TH, Hsu HC, Yuan RH, Jeng YM. Overexpression of CTHRC1 in hepatocellular carcinoma promotes tumor invasion and predicts poor prognosis. PLoS One. 2013; 8:e70324.
21. Amit S, Hatzubai A, Birman Y, Andersen JS, Ben-Shushan E, Mann M, Ben-Neriah Y, Alkalay I. Axin-mediated CKI phosphorylation of beta-catenin at Ser 45: a molecular switch for the Wnt pathway. Genes Dev. 2002; 16:1066–1076.
22. Behrens J, Jerchow BA, Würtele M, Grimm J, Asbrand C, Wirtz R, Kühl M, Wedlich D, Birchmeier W. Functional interaction of an axin homolog, conductin, with beta-catenin, APC, and GSK3beta. Science. 1998; 280:596–599.
23. Kolligs FT, Bommer G, Göke B. Wnt/beta-catenin/tcf signaling: a critical pathway in gastrointestinal tumorigenesis. Digestion. 2002; 66:131–144.
24. Malanchi I, Santamaria-Martínez A, Susanto E, Peng H, Lehr HA, Delaloye JF, Huelsken J. Interactions between cancer stem cells and their niche govern metastatic colonization. Nature. 2011; 481:85–89.
25. Fiorillo M, Verre AF, Iliut M, Peiris-Pagés M, Ozsvari B, Gandara R, Cappello AR, Sotgia F, Vijayaraghavan A, Lisanti MP. Graphene oxide selectively targets cancer stem cells, across multiple tumor types: Implications for non-toxic cancer treatment, via “differentiation-based nano-therapy”. Oncotarget. 2015; 6:3553–3562.
26. Kongkham PN, Northcott PA, Croul SE, Smith CA, Taylor MD, Rutka JT. The SFRP family of WNT inhibitors function as novel tumor suppressor genes epigenetically silenced in medulloblastoma. Oncogene. 2010; 29:3017–3024.
27. Wang JL, Chen ZF, Chen HM, Wang MY, Kong X, Wang YC, Sun TT, Hong J, Zou W, Xu J, Fang JY. Elf3 drives β-catenin transactivation and associates with poor prognosis in colorectal cancer. Cell Death Dis. 2014; 5:e1263.
28. Kolligs FT, Bommer G, Göke B. Wnt/beta-catenin/tcf signaling: a critical pathway in gastrointestinal tumorigenesis. Digestion. 2002; 66:131–144.
29. Gehrke I, Gandhirajan RK, Kreuzer KA. Targeting the WNT/beta-catenin/TCF/LEF1 axis in solid and haematological cancers: Multiplicity of therapeutic options. Eur J Cancer. 2009; 45:2759–2767.
30. Schmitz Y, Rateitschak K, Wolkenhauer O. Analysing the impact of nucleo-cytoplasmic shuttling of β-catenin and its antagonists APC, Axin and GSK3 on Wnt/β-catenin signalling. Cell Signal. 2013; 25:2210–2221.
31. Ding Y, Su S, Tang W, Zhang X, Chen S, Zhu G, Liang J, Wei W, Guo Y, Liu L, Chen YG, Wu W. The S-G2 phase enriched β-catenin/TCF complex ensures cell survival and cellcycle progression. J Cell Sci. 2014; 127:4833–4845.
32. Yamamoto S, Nishimura O, Misaki K, Nishita M, Minami Y, Yonemura S, Tarui H, Sasaki H. Cthrc1 selectively activates the planar cell polarity pathway of Wnt signaling by stabilizing the Wnt-receptor complex. Dev Cell. 2008; 15:23–36.