
INTRODUCTION
Breast cancer (BC) is one of the leading causes of death among women worldwide [1]. The increasing incidence of BC is due to various genetic and environmental changes that lead to the disruption of the cellular signaling network [2–5]. In 2012, there were 14.1 million new cancer cases globally, and BC accounted for 11.8% of them [6]. The introduction of trastuzumab, a humanized monoclonal antibody targeting the extracellular domain of HER2, revolutionized the treatment of HER2-positive BC [7–9]. The current gold standard in clinical practice is 1 year of adjuvant trastuzumab administration. Several trials have been conducted, looking to further refine the adjuvant treatment of patients with early-stage HER2-positive BC.
Catenin beta-1, also known as β-catenin, is a protein that is encoded by the CTNNB1 gene. β-Catenin is an oncogene that plays a key role in the signaling output of the canonical Wnt cascade [10]. Wnt signaling results in β-catenin accumulation and transcriptional activation of specific target genes that regulate a remarkable variety of cellular processes, such as cell proliferation, cell survival and migration [11]. Mutations and overexpression of β-catenin are associated with many cancers, including hepatocellular carcinoma, colorectal carcinoma, lung cancer, breast tumors, ovarian cancer and endometrial cancer [12]. Although mutation of CTNNB1 is rare in BC [13], mounting evidence has revealed that the mutations in CTNNB1 are often associated with an upregulation of β-catenin and the pathogenesis of endometrial cancer and ovarian cancer [14]. Several variants of CTNNB1 were found to be associated with BC risk [15, 16], but the mechanism of CTNNB1 in BC is still unknown.
Glycogen synthase kinase 3 beta, also known as GSK3B, is an enzyme that in humans is encoded by the GSK3B gene. GSK3B, a substrate of PI3K/Akt signaling, plays an important role in fundamental functions, such as the cell cycle, cytoskeletal integrity, apoptosis, transcription factor expression and formation of neurofibrillary tangles through PI3K/Akt signaling [17, 18]. GSK3B has also been shown to interact with CTNNB1 [19], as the phosphorylation of GSK3B leads to β-catenin nuclear accumulation [20].
There are no reliable biomarker profiles available to identify key genes and pathways in BC with trastuzumab treatment. Furthermore, numerous clinical studies have been performed with data (GSE22358) from the Gene Expression Omnibus (GEO, www.ncbi.nlm.nih.gov/geo/) [21–25]. The Glück et al. [26] dataset (GSE22358) includes 154 stage II–III samples from patients with operable early-stage breast cancer prior to neoadjuvant chemotherapy of capecitabine plus docetaxel, with (34) or without (120) trastuzumab. The data include histologic grade, molecular subtype, ER-status, PR-status, HER2-status, p53 status and response to treatment. Therefore, we chose GSE22358 to identify a molecular predictor of trastuzumab benefit in BC.
In this study, we chose GSE22358 from GEO and used the GEO2R online tool to detect the differentially expressed genes (DEGs). Subsequently, the DEGs were screened using Gene-E software and hub genes with a high degree of connectivity were selected. Next, we established a PPI network of the DEGs gene ontology (GO) and pathway enrichment analysis. Moreover, analyses of biological process (BP), molecular function (MF), cellular component (CC) and KEGG pathways of the DEGs and three modules were performed. Overall survival (OS) analysis of these hub genes was performed using the Kaplan-Meier plotter online database (http://kmplot.com/analysis/). By analyzing the biological functions and pathways, we may gain further insight into BC treatment at a molecular level and explore the potential candidate biomarkers for diagnosis, prognosis, and drug targets.
RESULTS
Identification of DEGs
A total of 154 operable early-stage BC samples receiving neoadjuvant capecitabine plus docetaxel, with (34) or without trastuzumab (120) were analyzed. The series from each chip was analyzed separately using GENE-E software, which identified the DEG lists. Based on the GENE-E analysis, using P < 0.05 criteria, a total of 2284 genes were identified after the analyses of GSE22358 of which 1231 were up-regulated and 1053 were down-regulated. The DEG expression heat map (top 50 up-regulated and down-regulated genes) is shown in Figure 1. We uploaded all DEGs to the online software DAVID to identify overrepresented GO categories and KEGG pathways. GO analysis results showed that up-regulated DEGs were significantly enriched in biological processes (BP), including DNA replication, protein N-linked glycosylation via asparagine, response to toxic substance, mRNA 3′-end processing, and anaphase-promoting complex-dependent catabolic process (Table 1); the down-regulated DEGs were significantly enriched in biological processes, including axon guidance, protein localization to plasma membrane, protein stabilization, protein glycosylation, and regulation of phosphatidylinositol 3-kinase signaling (Table 1). For molecular function (MF), the upregulated DEGs were enriched in DNA polymerase binding, ATPase activity, protein binding, microtubule binding, and poly(A) RNA binding, and the down-regulated DEGs were enriched in solute:proton symporter activity, metallocarboxypeptidase activity, ErbB-3 class receptor binding, calcium-dependent protein binding, and protein tyrosine phosphatase activity (Table 1). In addition, GO cell component (CC) analysis showed that the up-regulated DEGs were significantly enriched in the nucleoplasm, membrane, melanosome, nucleus, and intracellular membrane-bound organelles, and down-regulated DEGs enriched in the Golgi membrane, endoplasmic reticulum membrane, membrane, Golgi apparatus, and actin cytoskeleton (Table 1).
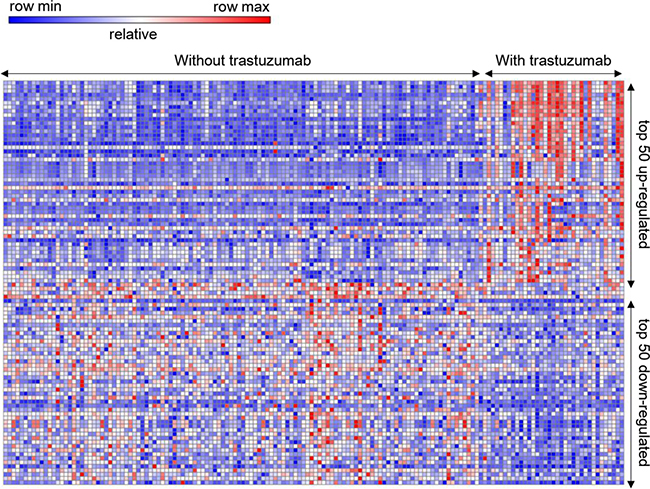
Figure 1: Heat map of the top 100 differentially expressed genes. Hierarchical clustering analysis to categorize the data into two groups that had similar expression patterns with or without trastuzumab Red: up-regulation, N = 50; Purple: down-regulation, N = 50.
Table 1: Gene ontology analysis of differentially expressed genes associated with trastuzumab
Expression |
Category |
Term |
Count |
% |
P-Value |
|---|---|---|---|---|---|
Up-regulated |
GOTERM_BP_DIRECT |
DNA replication |
25 |
2 |
8.80E-05 |
GOTERM_BP_DIRECT |
protein N-linked glycosylation via asparagine |
11 |
0.9 |
1.80E-04 |
|
GOTERM_BP_DIRECT |
response to toxic substance |
16 |
1.3 |
1.10E-03 |
|
GOTERM_BP_DIRECT |
mRNA 3’-end processing |
11 |
0.9 |
1.20E-03 |
|
GOTERM_BP_DIRECT |
anaphase-promoting complex-dependent catabolic process |
14 |
1.1 |
1.70E-03 |
|
GOTERM_BP_DIRECT |
DNA damage response, signal transduction by p53 class mediator resulting in cell cycle arrest |
12 |
1 |
2.00E-03 |
|
GOTERM_BP_DIRECT |
cell division |
38 |
3.1 |
2.00E-03 |
|
GOTERM_BP_DIRECT |
mRNA splicing, via spliceosome |
27 |
2.2 |
2.70E-03 |
|
GOTERM_BP_DIRECT |
regulation of signal transduction by p53 class mediator |
18 |
1.5 |
2.80E-03 |
|
GOTERM_BP_DIRECT |
DNA repair |
28 |
2.3 |
2.90E-03 |
|
GOTERM_MF_DIRECT |
DNA polymerase binding |
6 |
0.5 |
4.00E-04 |
|
GOTERM_MF_DIRECT |
ATPase activity |
25 |
2 |
9.80E-04 |
|
GOTERM_MF_DIRECT |
protein binding |
449 |
36.5 |
2.50E-03 |
|
GOTERM_MF_DIRECT |
microtubule binding |
25 |
2 |
4.70E-03 |
|
GOTERM_MF_DIRECT |
poly(A) RNA binding |
96 |
7.8 |
6.50E-03 |
|
GOTERM_MF_DIRECT |
damaged DNA binding |
11 |
0.9 |
6.70E-03 |
|
GOTERM_MF_DIRECT |
histone deacetylase binding |
15 |
1.2 |
8.60E-03 |
|
GOTERM_MF_DIRECT |
helicase activity |
13 |
1.1 |
1.00E-02 |
|
GOTERM_MF_DIRECT |
sulfuric ester hydrolase activity |
5 |
0.4 |
1.40E-02 |
|
GOTERM_MF_DIRECT |
ATP binding |
120 |
9.8 |
1.50E-02 |
|
GOTERM_CC_DIRECT |
nucleoplasm |
238 |
19.3 |
2.70E-07 |
|
GOTERM_CC_DIRECT |
membrane |
181 |
14.7 |
1.00E-04 |
|
GOTERM_CC_DIRECT |
melanosome |
18 |
1.5 |
1.80E-04 |
|
GOTERM_CC_DIRECT |
nucleus |
395 |
32.1 |
3.00E-04 |
|
GOTERM_CC_DIRECT |
intracellular membrane-bounded organelle |
56 |
4.6 |
5.50E-04 |
|
GOTERM_CC_DIRECT |
cytosol |
256 |
20.8 |
8.50E-04 |
|
GOTERM_CC_DIRECT |
kinetochore |
14 |
1.1 |
1.20E-03 |
|
GOTERM_CC_DIRECT |
nucleolus |
76 |
6.2 |
2.70E-03 |
|
GOTERM_CC_DIRECT |
proteasome complex |
11 |
0.9 |
4.50E-03 |
|
GOTERM_CC_DIRECT |
condensed chromosome kinetochore |
13 |
1.1 |
7.40E-03 |
|
Down-regulation |
GOTERM_BP_DIRECT |
axon guidance |
20 |
1.9 |
1.40E-03 |
GOTERM_BP_DIRECT |
protein localization to plasma membrane |
11 |
1 |
1.70E-03 |
|
GOTERM_BP_DIRECT |
protein stabilization |
17 |
1.6 |
3.00E-03 |
|
GOTERM_BP_DIRECT |
protein glycosylation |
15 |
1.4 |
3.00E-03 |
|
GOTERM_BP_DIRECT |
regulation of phosphatidylinositol 3-kinase signaling |
12 |
1.1 |
3.40E-03 |
|
GOTERM_BP_DIRECT |
response to cocaine |
8 |
0.8 |
3.60E-03 |
|
GOTERM_BP_DIRECT |
ERBB2 signaling pathway |
8 |
0.8 |
4.20E-03 |
|
GOTERM_BP_DIRECT |
substrate adhesion-dependent cell spreading |
8 |
0.8 |
4.20E-03 |
|
GOTERM_BP_DIRECT |
centrosome localization |
5 |
0.5 |
4.30E-03 |
|
GOTERM_BP_DIRECT |
O-glycan processing |
10 |
0.9 |
5.20E-03 |
|
GOTERM_MF_DIRECT |
solute:proton symporter activity |
3 |
0.3 |
8.40E-03 |
|
GOTERM_MF_DIRECT |
metallocarboxypeptidase activity |
6 |
0.6 |
1.40E-02 |
|
GOTERM_MF_DIRECT |
ErbB-3 class receptor binding |
3 |
0.3 |
1.60E-02 |
|
GOTERM_MF_DIRECT |
calcium-dependent protein binding |
9 |
0.9 |
1.60E-02 |
|
GOTERM_MF_DIRECT |
protein tyrosine phosphatase activity |
12 |
1.1 |
1.90E-02 |
|
GOTERM_MF_DIRECT |
receptor signaling protein activity |
7 |
0.7 |
2.00E-02 |
|
GOTERM_MF_DIRECT |
protein phosphatase binding |
9 |
0.9 |
2.00E-02 |
|
GOTERM_MF_DIRECT |
protein kinase binding |
31 |
2.9 |
2.20E-02 |
|
GOTERM_MF_DIRECT |
zinc ion binding |
81 |
7.7 |
2.50E-02 |
|
GOTERM_MF_DIRECT |
1-phosphatidylinositol-3-kinase activity |
7 |
0.7 |
2.70E-02 |
|
GOTERM_CC_DIRECT |
Golgi membrane |
58 |
5.5 |
1.70E-05 |
|
GOTERM_CC_DIRECT |
endoplasmic reticulum membrane |
77 |
7.3 |
2.50E-05 |
|
GOTERM_CC_DIRECT |
membrane |
152 |
14.4 |
1.10E-03 |
|
GOTERM_CC_DIRECT |
Golgi apparatus |
67 |
6.4 |
3.30E-03 |
|
GOTERM_CC_DIRECT |
actin cytoskeleton |
22 |
2.1 |
8.50E-03 |
|
GOTERM_CC_DIRECT |
cytoplasm |
317 |
30.1 |
1.10E-02 |
|
GOTERM_CC_DIRECT |
mitochondrial membrane |
12 |
1.1 |
1.20E-02 |
|
GOTERM_CC_DIRECT |
axon |
22 |
2.1 |
1.70E-02 |
|
GOTERM_CC_DIRECT |
cis-Golgi network |
7 |
0.7 |
1.80E-02 |
|
GOTERM_CC_DIRECT |
uropod |
4 |
0.4 |
1.90E-02 |
Abbreviations: GO: Gene Ontology.
KEGG pathway analysis
Table 2 contains the most significantly enriched pathways of the up-regulated DEGs and down-regulated DEGs analyzed by KEGG analysis. The up-regulated DEGs were enriched in protein processing in the endoplasmic reticulum, base excision repair, pentose and glucuronate interconversions, proteasome, and ascorbate and aldarate metabolism while the down-regulated DEGs were enriched in proteoglycans in cancer, the ErbB signaling pathway, bacterial invasion of epithelial cells, the insulin signaling pathway, and the neurotrophin signaling pathway.
Table 2: KEGG pathway analysis of differentially expressed genes associated with trastuzumab
Category |
KEEG Term |
Count |
% |
P-Value |
|---|---|---|---|---|
Down-regulation |
Proteoglycans in cancer |
23 |
2.2 |
1.40E-03 |
ErbB signaling pathway |
13 |
1.2 |
2.80E-03 |
|
Bacterial invasion of epithelial cells |
12 |
1.1 |
3.40E-03 |
|
Insulin signaling pathway |
17 |
1.6 |
3.70E-03 |
|
Neurotrophin signaling pathway |
15 |
1.4 |
6.00E-03 |
|
Up-regulation |
Protein processing in endoplasmic reticulum |
27 |
2.2 |
1.10E-04 |
Base excision repair |
8 |
0.7 |
6.90E-03 |
|
Pentose and glucuronate interconversions |
8 |
0.7 |
1.10E-02 |
|
Proteasome |
8 |
0.7 |
3.20E-02 |
|
Ascorbate and aldarate metabolism |
6 |
0.5 |
3.70E-02 |
Abbreviations: KEGG: Kyoto Encyclopedia of Genes and Genomes.
Module screening from the PPI network
Based on the information in the STRING database, among these genes, GSK3B showed a 42-node degree. Moreover, a total of 1000 nodes and 2079 edges were analyzed using the plug-in MCODE. The top 3 significant modules were selected, and the functional annotation of the genes involved in the modules were analyzed (Figure 2). Enrichment analysis showed that the genes in module 1 were mainly associated with GSK3B, RAC1, PXN, ERBB2, HSP90AA1, FGF2, PIK3R1, and RAC2.
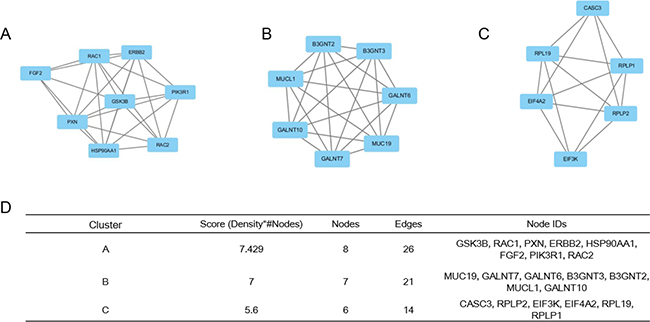
Figure 2: Top 3 modules from the protein–protein interaction network. (A) module 1, (B) module 2, (C) module 3 were analyzed using the plug-in Molecular Complex Detection in Cytoscape based on the information in the STRING database, (D) the enriched pathways and functional annotation of the genes involved in the modules of 1, 2, and 3.
GSK3B is upregulated in breast cancer
Cross-cancer alteration analysis showed that GSK3B has an amplification pattern in most cancer types, especially in breast cancer (Figure 3). To further investigate the function of GSK3B in breast cancer, we performed immunohistochemical staining on breast cancer tissues and adjacent normal tissues. The staining results revealed significantly higher positivity for GSK3B expression in BC tissues than in adjacent normal tissues (Figure 4A). Similarly, GSK3B is also highly expressed in breast cancer patients (Figure 4B, 4C). Beta-catenin encoded by CTNNB1 is a constituent of adherens junctions and acts as an intracellular signal transducer in the Wnt signaling pathway, a pathway that is closely related with the occurrence, development, invasion and metastasis of breast cancer. We also found that β-catenin is upregulated in breast cancer patients (Figure 4B, 4C). GSK3B and CTNNB1 are highly expressed in bladder cancer and breast cancer, and they are notably positively correlated (Figure 4D, 4E). Moreover, overexpression of GSK3B or CTNNB1 increased cell proliferation (Figure 4F). High levels of GSK3B and CTNNB1 were observed in poor survival curves (Figure 5). These findings have revealed that the GSK3B signaling pathway may be a potential target for breast cancer therapy.
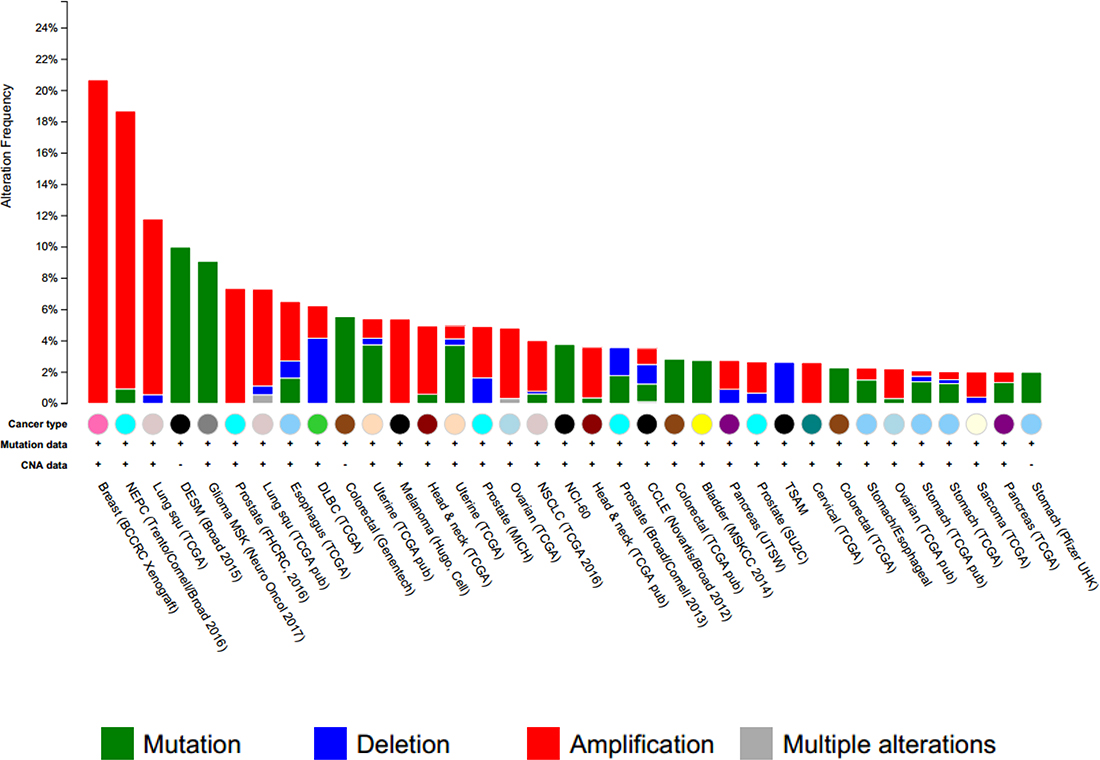
Figure 3: Cross-cancer alteration summary for GSK3B (166 studies/1 gene). GSK3B was analyzed from the cBioPortal for Cancer Genomics (http://www.cbioportal.org). The red column indicates the amplification pattern.
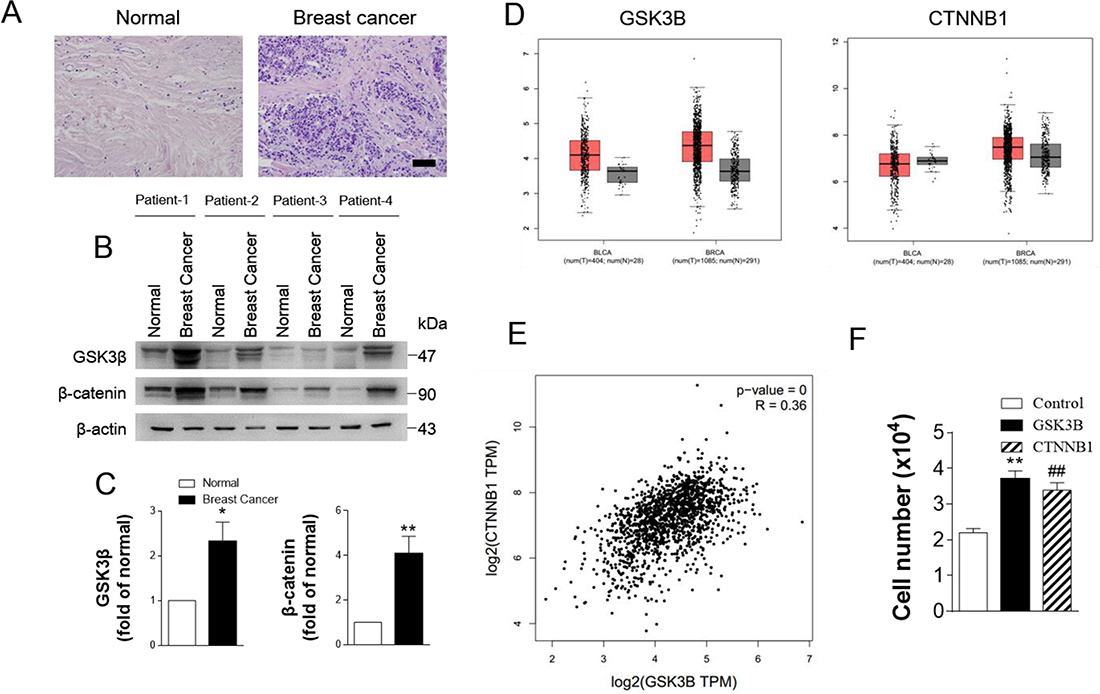
Figure 4: GSK3B is upregulated in breast cancer. (A) Hematoxylin and eosin staining of breast cancer tissue. (B) and (C) Typical western blots and statistical data of GSK3B and β-catenin from normal and breast cancer tissue. N = 5, data are expressed as the mean ± s.e.m.; **P < 0.01. *P < 0.05, normal compared to breast cancer; Student’s t-test. (D) Expression level of GSK3B and CNNTB1 in cancer and normal tissues. BLCA: Bladder Carcinoma; BRCA: Breast Carcinoma. (E) The pair-wise gene expression correlation analysis for GSK3B and CNNTB1 in breast cancer. GSK3B and CNNTB1 are positively correlated. (F) Statistical analysis of cell number in MDA-MB-231 cells infected with Ad-GSK3B (MOI 25) or Ad-CTNNB1 (MOI 25). Results are expressed as mean ± SEM. **p < 0.01, control vs Ad- GSK3B; ##p < 0.01, control vs Ad-CTNNB1 (n = 4).
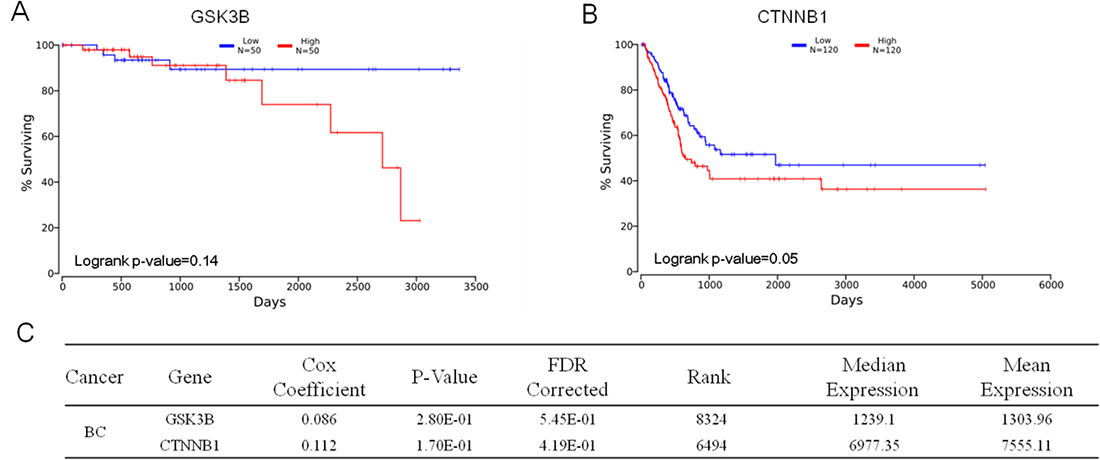
Figure 5: Prognostic value of GSK3B and CTNNB1. (A) and (B) Kaplan plot for GSK3B (A), CTNNB1 (B) in breast cancer. (C) Statistical data of GSK3B (N = 50) and CTNNB1 (N = 120). The data are expressed as the mean ± s.e.m., low expression compared to high; Student’s t-test.
DISCUSSION
As the leading cause of cancer mortality in women, BC is a serious public health problem worldwide, and the age of onset tends to be younger in recent years [1, 5, 27]. In addition, BC is lacking effective methods for early screening and diagnosis. Therefore, sensitive and specific biomarkers for BC are urgently needed. Trastuzumab, an anti-HER2 humanized antibody, has shown great clinical benefits in HER2-positive BC treatment [28]. In recent years, there has been great interest in researching the mechanisms of trastuzumab treatment. Studies have shown that numerous molecules play important roles in how a patient responds to trastuzumab, including ERBB-family SNPs, p53 protein, BAG-1 protein and individual patients’ metabolism [29–32]. Xiong et al. [33] confirmed that CD147 suppression enhances the effects of trastuzumab through MAPK and Akt phosphorylation while HER2 amplification level is not currently a prognostic factor for trastuzumab-based targeted therapy [34]. In addition, studies have shown that AUY922 [35] or taxane [36] plus trastuzumab is an effective regimen for patients with relapsed HER2-positive BC after (neo)adjuvant trastuzumab.
However, trastuzumab resistance has emerged as a major problem in its clinical application. Many studies have attempted to elucidate the mechanisms underlying trastuzumab resistance. Heregulin, MEOX1 and lncRNA GAS5 confers resistance to the anti-HER2 agent trastuzumab [37–39]. Some studies have demonstrated that inhibition of S100P and depletion of KLK10 results in reversal of trastuzumab-resistance (TzR) [40, 41]. In contrast to trastuzumab that inhibits the ErbB2 homodimer, another therapeutic antibody H2-18 binds to domain I of ErbB2, which induces programmed cell death (PCD) and exhibits greater antitumor efficacy than trastuzumab [42].
In the present study, we extracted data from GSE22358 and identify 1231 up-regulated and 1053 down-regulated DEGs with or without trastuzumab using bioinformatics analysis. To gain a more in-depth understanding of these DEGs, we performed GO function and KEGG pathway analysis. The GO analysis showed that up-regulated DEGs were mainly involved in DNA replication, protein N-linked glycosylation via asparagine, and response to toxic substance, and down-regulated DEGs were involved in axon guidance, protein localization to plasma membrane, protein stabilization, and protein glycosylation. Furthermore, the KEGG pathways of up-regulated DEGs included proteoglycans in cancer, the ErbB signaling pathway, bacterial invasion of epithelial cells, the insulin signaling pathway and the neurotrophin signaling pathway while the down-regulated DEGs were enriched in protein processing in the endoplasmic reticulum, base excision repair, pentose and glucuronate interconversions, and proteasome ascorbate and aldarate metabolism. This finding is consistent with the fact that glycosylation and metabolic processes play an important role in cancer processes.
We also constructed a PPI network with DEGs and listed the top eight hub genes: GSK3B, RAC1, PXN, ERBB2, HSP90AA1, FGF2, PIK3R1 and RAC2. The hub genes play an important role in cancer cell growth, migration, and invasion, especially in breast cancer [43–47]. Furthermore, these genes were involved in significant pathways, including the Fc receptor signaling pathway and regulation of the immune response pathway.
GSK3B was identified as one of the hub genes exhibiting the highest degree of connectivity. Furthermore, our experimental results showed that GSK3B was also highly expressed in breast cancer tissues and associated with poor survival (Figure 4A–4D). We hypothesize that this gene might contribute to the progression of breast cancer with trastuzumab treatment.
GSK3B associates with the destruction complex through a binding site in AXIN1 and phosphorylates β-catenin, which is subsequently targeted for proteosomal degradation [48]. Wang et al. [43] conducted an association study to determine whether common genetic variations in six genes (APC, AXIN1, AXIN2, CSNK1D, CSNK1E, and GSK3B) that encode the destruction complex of the Wnt/β-catenin signaling pathway account in part for the contribution of the pathway to BC risk. Mole et al. [49] also reported that the truncated somatostatin receptor variant sst5TMD4 is associated with increased invasiveness and aggressiveness in BC. sst5TMD4 overexpression increases vimentin, total β-catenin and phosphorylated GSK3B levels.
GSK3B can interact with β-catenin [19], and β-catenin is also highly expressed in BC. β-Catenin is a marker of poor prognosis in human cancer and has been implicated in human breast cancer, via targeting cyclin D1 or vimentin [50–52]. A number of studies have suggested that dysregulation of Wnt/β-catenin signaling occurs in human breast cancer. Blockade of Wnt/β-catenin signaling could suppress breast cancer metastasis [53, 54]. Figure 4E shows the results of the correlation analysis between GSK3B and β-catenin. GSK3B and β-catenin are notably positively correlated. Therefore, we speculated that GSK3B has a similar function as β-catenin in breast cancer, but further verification is still needed.
In conclusion, our data provide a comprehensive bioinformatics analysis of DEGs, which may be involved in the progress of trastuzumab treatment. The study provides a set of useful targets for future investigation into the molecular mechanisms and biomarkers. However, further molecular biological experiments are required to confirm the function of the identified genes.
MATERIALS AND METHODS
Microarray data
The gene expression profiles of GSE22358 were downloaded from the GEO database. GSE22358, which was based on the Agilent GPL5325 platform (Agilent, CA, USA), was submitted by Gluck et al. The GSE22358 dataset contained 154 operable early-stage BC samples receiving neoadjuvant capecitabine plus docetaxel, with (34) or without trastuzumab (120).
Identification of DEGs
The raw data files used for the analysis included TXT files (Agilent platform). The analysis was performed using GENE-E (version 3.0, Broad Institute, USA). We applied hierarchical clustering analysis to categorize the data into two groups that had similar expression patterns with or without trastuzumab. We used a classical t test to identify DEGs with defined a P value cutoff of <0.05 as statistically significant.
Gene ontology and pathway enrichment analysis of DEGs
Gene ontology analysis (GO) is a common useful method for annotating genes and gene products and for identifying characteristic biological attributes of high-throughput genome or transcriptome data [55, 56]. KEGG (http://www.genome.jp/) is a knowledge base for systematic analysis of gene functions, linking genomic information with higher-order functional information [57, 58]. Comprehensively mapping a user’s gene to the relevant biological annotation in the DAVID database (https://david.ncifcrf.gov/) is an essential foundation for the success of any high-throughput functional gene analysis [59]. To analyze the DEGs at the functional level, GO enrichment and KEGG pathway analysis were performed using the DAVID online tool. P < 0.05 was considered statistically significant.
Integration of a protein–protein interaction (PPI) network and module analysis
The Search Tool for the Retrieval of Interacting Genes (STRING) database is an online tool designed to evaluate protein–protein interaction (PPI) information. STRING (version 10.0) covers 9,643,763 proteins from 2031 organisms. To evaluate the interactive relationships among DEGs, we mapped the DEGs to STRING, and only experimentally validated interactions with a combined score >0.4 were selected as significant. Then, PPI networks were constructed using the Cytoscape software. The plug-in Molecular Complex Detection (MCODE) was used to screen the modules of the PPI network in Cytoscape. The criteria were set as follows: MCODE scores >3 and number of nodes >4. Moreover, the function and pathway enrichment analyses were performed for DEGs in the modules. P < 0.05 was considered to have significant differences.
Integrative analysis of complex cancer genomics with cBioPortal
The cBioPortal for Cancer Genomics (http://www.cbioportal.org) provides visualization, analysis and download of large-scale cancer genomics data sets [60, 61]. GSK3B was selected for analysis in different cancers.
Human sample collection
The patients were from Dazhou Central Hospital. (Informed written consent was obtained from all patients. The study protocol conformed to the Ethical Guidelines of the 1975 Declaration of Helsinki and was approved by the Dazhou Central Hospital Human Ethics Committee.)
Cell culture, adenoviral infection
MDA-MB-231 cells (Cell Resource Center, IBMS, CAMS/PUMC, Beijing, China) were cultured at 37° C under 5% CO2 in Dulbecco’s modified Eagle’s medium supplemented with 10% fetal bovine serum (Gibco, Waltham, MA, USA), and 1% penicillin-streptomycin. Approximately 80% confluent MDA-MB-231were infected with adenovirus DNA expressing GSK3B or CTNNB1 at a multiplicity of infection (MOI) of 25, and grown for a further 48 h.
Statistical analysis
All data are expressed as the mean ± s.e.m. Statistical analysis was performed with GraphPad PRISM version 5.01 (GraphPad software, Inc.) and SPSS 18.0 software package (SPSS Inc.).
Abbreviations
BC: Breast Cancer; Bcl-xL: B-cell Lymphoma-extra Large; CC: Cell Component; CTNNB1: Catenin Beta 1; DAVID: Database for Annotation, Visualization and Integrated Discovery; DEGs: Differentially Expressed Genes; ERK1/2: Extracellular Regulated Protein Kinases; FGF2: Fibroblast Growth Factor 2; FGFR2: Fibroblast Growth Factor Receptor 2; GO: Gene Ontology; GSK3B: Glycogen Synthase Kinase 3 beta; HER2: Human Epidermal growth factor Receptor 2; HSP90AA1: Heat Shock Protein 90 Alpha Family Class A Member 1; KEGG: Kyoto Encyclopedia of Genes and Genomes pathway; KRT19: Keratin 19; MCODE: Molecular Complex Detection; Mcl-1: Myeloid Cell Leukemia 1; MF: Molecular Function; NF-κB: Nuclear Factor-κB; NUMB: Endocytic Adaptor Protein; PIK3R1: Phosphoinositide-3-Kinase Regulatory Subunit 1; PPI: Protein–Protein Interaction; RAC1: Ras-related C3 botulinum toxin substrate 1; RAC2: Ras-related C3 botulinum toxin substrate 2; RSK2: Ribosomal S6 Kinase 2; STRING: Search Tool for the Retrieval of Interacting Genes.
Author contributions
J.F. and F.H. collected and analyzed the data. Y.T. wrote and revised the manuscript. X.F., F.Z. and Y.C. analyzed the data. F.Z. designed experiments, interpreted the data, and revised the manuscript.
CONFLICTS OF INTEREST
The authors declare that there are no conflicts of interest.
FUNDING
National Natural Science Foundation of China (81471063).
REFERENCES
1. DeSantis C, Ma J, Bryan L, Jemal A. Breast cancer statistics. 2013. CA Cancer J Clin. 2014; 64:52–62. https://doi.org/10.3322/caac.21203.
2. Khalid S, Hanif R, Tareen SH, Siddiqa A, Bibi Z, Ahmad J. Formal modeling and analysis of ER-αassociated Biological Regulatory Network in breast cancer. Peer J. 2016; 4:e2542. https://doi.org/10.7717/peerj.2542.
3. Hotes JL, Ellison LF, Howe HL, Friesen I, Kohler B. Variation in breast cancer counts using SEER and IARC multiple primary coding rules. Cancer Causes Control. 2004; 15:185–91. https://doi.org/10.1023/B:CACO.0000019505.97836.7d.
4. Madigan MP, Ziegler RG, Benichou J, Byrne C, Hoover RN. Proportion of breast cancer cases in the United States explained by well-established risk factors. J Natl Cancer Inst. 1995; 87:1681–85. https://doi.org/10.1093/jnci/87.22.1681.
5. McPherson K, Steel CM, Dixon JM. ABC of breast diseases. Breast cancer-epidemiology, risk factors, and genetics. BMJ. 2000; 321:624–28. https://doi.org/10.1136/bmj.321.7261.624.
6. Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015; 136:E359–86. https://doi.org/10.1002/ijc.29210.
7. Zardavas D, Fouad TM, Piccart M. Optimal adjuvant treatment for patients with HER2-positive breast cancer in 2015. Breast. 2015; 24:S143–48. https://doi.org/10.1016/j.breast.2015.07.034.
8. Zardavas D, Tryfonidis K, Goulioti T, Piccart M. Targeted adjuvant therapy in breast cancer. Expert Rev Anticancer Ther. 2016; 16:1263–75. https://doi.org/10.1080/14737140.2016.1247698.
9. Verma S, Miles D, Gianni L, Krop IE, Welslau M, Baselga J, Pegram M, Oh DY, Diéras V, Guardino E, Fang L, Lu MW, Olsen S, Blackwell K. Trastuzumab Emtansine for HER2-Positive Advanced Breast Cancer. The New England Journal of Medicine. 2012; 367:1783–1791.
10. Clevers H, Nusse R. Wnt/β-catenin signaling and disease. Cell. 2012; 149:1192–205. https://doi.org/10.1016/j.cell.2012.05.012.
11. Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol. 2004; 20:781–810. https://doi.org/10.1146/annurev.cellbio.20.010403.113126.
12. Hoffmeyer K, Raggioli A, Rudloff S, Anton R, Hierholzer A, Del Valle I, Hein K, Vogt R, Kemler R. Wnt/β-catenin signaling regulates telomerase in stem cells and cancer cells. Science. 2012; 336:1549–54. https://doi.org/10.1126/science.1218370.
13. Ueda M, Gemmill RM, West J, Winn R, Sugita M, Tanaka N, Ueki M, Drabkin HA. Mutations of the β- and γ-catenin genes are uncommon in human lung, breast, kidney, cervical and ovarian carcinomas. Br J Cancer. 2001; 85:64–68. https://doi.org/10.1054/bjoc.2001.1863.
14. McConechy MK, Ding J, Senz J, Yang W, Melnyk N, Tone AA, Prentice LM, Wiegand KC, McAlpine JN, Shah SP, Lee CH, Goodfellow PJ, Gilks CB, Huntsman DG. Ovarian and endometrial endometrioid carcinomas have distinct CTNNB1 and PTEN mutation profiles. Mod Pathol. 2014; 27:128–34. https://doi.org/10.1038/modpathol.2013.107.
15. Alanazi MS, Parine NR, Shaik JP, Alabdulkarim HA, Ajaj SA, Khan Z. Association of single nucleotide polymorphisms in Wnt signaling pathway genes with breast cancer in Saudi patients. PLoS One. 2013; 8:e59555. https://doi.org/10.1371/journal.pone.0059555.
16. Lee JY, Park AK, Lee KM, Park SK, Han S, Han W, Noh DY, Yoo KY, Kim H, Chanock SJ, Rothman N, Kang D. Candidate gene approach evaluates association between innate immunity genes and breast cancer risk in Korean women. Carcinogenesis. 2009; 30:1528–31. https://doi.org/10.1093/carcin/bgp084.
17. Porta C, Paglino C, Mosca A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front Oncol. 2014; 4:64. https://doi.org/10.3389/fonc.2014.00064.
18. Lu Y, Lei S, Wang N, Lu P, Li W, Zheng J, Giri PK, Lu H, Chen X, Zuo Z, Liu Y, Zhang P. Protective Effect of Minocycline Against Ketamine-Induced Injury in Neural Stem Cell: Involvement of PI3K/Akt and Gsk-3 Beta Pathway. Front Mol Neurosci. 2016; 9:135. https://doi.org/10.3389/fnmol.2016.00135.
19. Kishida S, Yamamoto H, Hino S, Ikeda S, Kishida M, Kikuchi A. DIX domains of Dvl and axin are necessary for protein interactions and their ability to regulate beta-catenin stability. Mol Cell Biol. 1999; 19:4414–22. https://doi.org/10.1128/MCB.19.6.4414.
20. Liu L, Zhou XM, Yang FF, Miao Y, Yin Y, Hu XJ, Hou G, Wang QY, Kang J. TRIM22 confers poor prognosis and promotes epithelial-mesenchymal transition through regulation of AKT/GSK3β/β-catenin signaling in non-small cell lung cancer. Oncotarget. 2017; 8:62069–80.
21. Triulzi T, De Cecco L, Sandri M, Prat A, Giussani M, Paolini B, Carcangiu ML, Canevari S, Bottini A, Balsari A, Menard S, Generali D, Campiglio M, et al. Whole-transcriptome analysis links trastuzumab sensitivity of breast tumors to both HER2 dependence and immune cell infiltration. Oncotarget. 2015; 6:28173–82. https://doi.org/10.18632/oncotarget.4405.
22. Mannelqvist M, Wik E, Stefansson IM, Akslen LA. An 18-gene signature for vascular invasion is associated with aggressive features and reduced survival in breast cancer. PLoS One. 2014; 9:e98787. https://doi.org/10.1371/journal.pone.0098787.
23. Lehmann BD, Jovanović B, Chen X, Estrada MV, Johnson KN, Shyr Y, Moses HL, Sanders ME, Pietenpol JA. Refinement of Triple-Negative Breast Cancer Molecular Subtypes: Implications for Neoadjuvant Chemotherapy Selection. PLoS One. 2016; 11:e0157368. https://doi.org/10.1371/journal.pone.0157368.
24. Dennison JB, Molina JR, Mitra S, González-Angulo AM, Balko JM, Kuba MG, Sanders ME, Pinto JA, Gómez HL, Arteaga CL, Brown RE, Mills GB. Lactate dehydrogenase B: a metabolic marker of response to neoadjuvant chemotherapy in breast cancer. Clin Cancer Res. 2013; 19:3703–13. https://doi.org/10.1158/1078-0432.CCR-13-0623.
25. Kwok HF, Zhang SD, McCrudden CM, Yuen HF, Ting KP, Wen Q, Khoo US, Chan KY. Prognostic significance of minichromosome maintenance proteins in breast cancer. Am J Cancer Res. 2014; 5:52–71.
26. Glück S, Ross JS, Royce M, McKenna EF Jr, Perou CM, Avisar E, Wu L. TP53 genomics predict higher clinical and pathologic tumor response in operable early-stage breast cancer treated with docetaxel-capecitabine ± trastuzumab. Breast Cancer Res Treat. 2012; 132:781–91. https://doi.org/10.1007/s10549-011-1412-7.
27. Jemal A, Siegel R, Ward E, Murray T, Xu J, Thun MJ. Cancer statistics, 2007. CA Cancer J Clin. 2007; 57:43–66. https://doi.org/10.3322/canjclin.57.1.43.
28. Piccart-Gebhart MJ, Procter M, Leyland-Jones B, Goldhirsch A, Untch M, Smith I, Gianni L, Baselga J, Bell R, Jackisch C, Cameron D, Dowsett M, Barrios CH, et al; Herceptin Adjuvant (HERA) Trial Study Team. Trastuzumab after adjuvant chemotherapy in HER2-positive breast cancer. N Engl J Med. 2005; 353:1659–72.
29. Toomey S, Madden SF, Furney SJ, Fan Y, McCormack M, Stapleton C, Cremona M, Cavalleri GL, Milewska M, Elster N, Carr A, Fay J, Kay EW, et al. The impact of ERBB-family germline single nucleotide polymorphisms on survival response to adjuvant trastuzumab treatment in HER2-positive breast cancer. Oncotarget. 2016; 7:75518–25. https://doi.org/10.18632/oncotarget.12782.
30. Fountzilas G, Giannoulatou E, Alexopoulou Z, Zagouri F, Timotheadou E, Papadopoulou K, Lakis S, Bobos M, Poulios C, Sotiropoulou M, Lyberopoulou A, Gogas H, Pentheroudakis G, et al. TP53 mutations and protein immunopositivity may predict for poor outcome but also for trastuzumab benefit in patients with early breast cancer treated in the adjuvant setting. Oncotarget. 2016; 7:32731–53. https://doi.org/10.18632/oncotarget.9022.
31. Papadakis E, Robson N, Yeomans A, Bailey S, Laversin S, Beers S, Sayan AE, Ashton-Key M, Schwaiger S, Stuppner H, Troppmair J, Packham G, Cutress R. A combination of trastuzumab and BAG-1 inhibition synergistically targets HER2 positive breast cancer cells. Oncotarget. 2016; 7:18851–64. https://doi.org/10.18632/oncotarget.7944.
32. Miolo G, Muraro E, Caruso D, Crivellari D, Ash A, Scalone S, Lombardi D, Rizzolio F, Giordano A, Corona G. Pharmacometabolomics study identifies circulating spermidine and tryptophan as potential biomarkers associated with the complete pathological response to trastuzumab-paclitaxel neoadjuvant therapy in HER-2 positive breast cancer. Oncotarget. 2016; 7:39809–22. https://doi.org/10.18632/oncotarget.9489.
33. Xiong L, Ding L, Ning H, Wu C, Fu K, Wang Y, Zhang Y, Liu Y, Zhou L. CD147 knockdown improves the antitumor efficacy of trastuzumab in HER2-positive breast cancer cells. Oncotarget. 2016; 7:57737–51. https://doi.org/10.18632/oncotarget.10252.
34. Xu QQ, Pan B, Wang CJ, Zhou YD, Mao F, Lin Y, Guan JH, Shen SJ, Zhang XH, Xu YL, Zhong Y, Wang XJ, Zhang YN, Sun Q. HER2 amplification level is not a prognostic factor for HER2-positive breast cancer with trastuzumab-based adjuvant treatment: a systematic review and meta-analysis. Oncotarget. 2016; 7:63571–82. https://doi.org/10.18632/oncotarget.11541.
35. Kong A, Rea D, Ahmed S, Beck JT, López López R, Biganzoli L, Armstrong AC, Aglietta M, Alba E, Campone M, Hsu Schmitz SF, Lefebvre C, Akimov M, Lee SC. Phase 1B/2 study of the HSP90 inhibitor AUY922 plus trastuzumab in metastatic HER2-positive breast cancer patients who have progressed on trastuzumab-based regimen. Oncotarget. 2016; 7:37680–92. https://doi.org/10.18632/oncotarget.8974.
36. Xu B, Hu X, Zheng H, Wang X, Zhang Q, Cui S, Liu D, Liao N, Luo R, Sun Q, Yu S. Outcomes of re-treatment with first-line trastuzumab plus a taxane in HER2 positive metastatic breast cancer patients after (neo)adjuvant trastuzumab: A prospective multicenter study. Oncotarget. 2016; 7:50643–55.
37. Nonagase Y, Yonesaka K, Kawakami H, Watanabe S, Haratani K, Takahama T, Takegawa N, Ueda H, Tanizaki J, Hayashi H, Yoshida T, Takeda M, Chiba Y, et al. Heregulin-expressing HER2-positive breast and gastric cancer exhibited heterogeneous susceptibility to the anti-HER2 agents lapatinib, trastuzumab and T-DM1. Oncotarget. 2016; 7:84860–71. https://doi.org/10.18632/oncotarget.12743.
38. Sun L, Burnett J, Gasparyan M, Xu F, Jiang H, Lin CC, Myers I, Korkaya H, Liu Y, Connarn J, He H, Zhang N, Wicha MS, Sun D. Novel cancer stem cell targets during epithelial to mesenchymal transition in PTEN-deficient trastuzumab-resistant breast cancer. Oncotarget. 2016; 7:51408–22.
39. Li W, Zhai L, Wang H, Liu C, Zhang J, Chen W, Wei Q. Downregulation of LncRNA GAS5 causes trastuzumab resistance in breast cancer. Oncotarget. 2016; 7:27778–86.
40. Merry CR, McMahon S, Forrest ME, Bartels CF, Saiakhova A, Bartel CA, Scacheri PC, Thompson CL, Jackson MW, Harris LN, Khalil AM. Transcriptome-wide identification of mRNAs and lincRNAs associated with trastuzumab-resistance in HER2-positive breast cancer. Oncotarget. 2016; 7:53230–44. https://doi.org/10.18632/oncotarget.10637.
41. Wang Z, Ruan B, Jin Y, Zhang Y, Li J, Zhu L, Xu W, Feng L, Jin H, Wang X. Identification of KLK10 as a therapeutic target to reverse trastuzumab resistance in breast cancer. Oncotarget. 2016; 7:79494–502.
42. Lu Q, Wang L, Zhang Y, Yu X, Wang C, Wang H, Yang Y, Chong X, Xia T, Meng Y, Wang Y, Lu C, Zhou L, Li B. An anti-ErbB2 fully human antibody circumvents trastuzumab resistance. Oncotarget. 2016; 7:67129–41. https://doi.org/10.18632/oncotarget.11562.
43. Wang X, Goode EL, Fredericksen ZS, Vierkant RA, Pankratz VS, Liu-Mares W, Rider DN, Vachon CM, Cerhan JR, Olson JE, Couch FJ. Association of genetic variation in genes implicated in the beta-catenin destruction complex with risk of breast cancer. Cancer Epidemiol Biomarkers Prev. 2008; 17:2101–08. https://doi.org/10.1158/1055-9965.EPI-08-0134.
44. Zou T, Mao X, Yin J, Li X, Chen J, Zhu T, Li Q, Zhou H, Liu Z. Emerging roles of RAC1 in treating lung cancer patients. Clin Genet. 2017; 91:520–528.
45. Li D, Marchenko ND. ErbB2 inhibition by lapatinib promotes degradation of mutant p53 protein in cancer cells. Oncotarget. 2017; 8:5823–5833. https://doi.org/10.18632/oncotarget.12878.
46. Cizkova M, Vacher S, Meseure D, Trassard M, Susini A, Mlcuchova D, Callens C, Rouleau E, Spyratos F, Lidereau R, Bièche I. PIK3R1 underexpression is an independent prognostic marker in breast cancer. BMC Cancer. 2013; 13:545. https://doi.org/10.1186/1471-2407-13-545.
47. Czaplinska D, Mieczkowski K, Supernat A, Skladanowski AC, Kordek R, Biernat W, Zaczek AJ, Romanska HM, Sadej R. Interactions between FGFR2 and RSK2-implications for breast cancer prognosis. Tumour Biol. 2016; 37:13721–31. https://doi.org/10.1007/s13277-016-5266-9.
48. Liu C, Kato Y, Zhang Z, Do VM, Yankner BA, He X. beta-Trcp couples beta-catenin phosphorylation-degradation and regulates Xenopus axis formation. Proc Natl Acad Sci USA. 1999; 96:6273–78. https://doi.org/10.1073/pnas.96.11.6273.
49. Molè D, Gentilin E, Ibañez-Costa A, Gagliano T, Gahete MD, Tagliati F, Rossi R, Pelizzo MR, Pansini G, Luque RM, Castaño JP, degli Uberti E, Zatelli MC. The expression of the truncated isoform of somatostatin receptor subtype 5 associates with aggressiveness in medullary thyroid carcinoma cells. Endocrine. 2015; 50:442–52. https://doi.org/10.1007/s12020-015-0594-x.
50. Geyer FC, Lacroix-Triki M, Savage K, Arnedos M, Lambros MB, MacKay A, Natrajan R, Reis-Filho JS. β-Catenin pathway activation in breast cancer is associated with triple-negative phenotype but not with CTNNB1 mutation. Mod Pathol. 2011; 24:209–31. https://doi.org/10.1038/modpathol.2010.205.
51. Lin SY, Xia W, Wang JC, Kwong KY, Spohn B, Wen Y, Pestell RG, Hung MC. Beta-catenin, a novel prognostic marker for breast cancer: its roles in cyclin D1 expression and cancer progression. Proc Natl Acad Sci USA. 2000; 97:4262–66. https://doi.org/10.1073/pnas.060025397.
52. Gilles C, Polette M, Mestdagt M, Nawrocki-Raby B, Ruggeri P, Birembaut P, Foidart JM. Transactivation of vimentin by β-catenin in human breast cancer cells. Cancer Res. 2003; 63:2658–64.
53. Zardawi SJ, O’Toole SA, Sutherland RL, Musgrove EA. Dysregulation of Hedgehog, Wnt and Notch signalling pathways in breast cancer. Histol Histopathol. 2009; 24:385–98.
54. Jang GB, Kim JY, Cho SD, Park KS, Jung JY, Lee HY, Hong IS, Nam JS. Blockade of Wnt/β-catenin signaling suppresses breast cancer metastasis by inhibiting CSC-like phenotype. Sci Rep. 2015; 5:12465. https://doi.org/10.1038/srep12465.
55. Gene Ontology Consortium. The Gene Ontology (GO) project in 2006. Nucleic Acids Res. 2006; 34:D322–26. https://doi.org/10.1093/nar/gkj021.
56. Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, et al, and The Gene Ontology Consortium. Gene ontology: tool for the unification of biology. Nat Genet. 2000; 25:25–29. https://doi.org/10.1038/75556.
57. Ogata H, Goto S, Sato K, Fujibuchi W, Bono H, Kanehisa M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 1999; 27:29–34.
58. Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000; 28:27–30. https://doi.org/10.1093/nar/28.1.27.
59. Dennis G Jr, Sherman BT, Hosack DA, Yang J, Gao W, Lane HC, Lempicki RA. DAVID: Database for Annotation, Visualization, and Integrated Discovery. Genome Biol. 2003; 4:3. https://doi.org/10.1186/gb-2003-4-5-p3.
60. Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, Antipin Y, Reva B, Goldberg AP, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012; 2:401–04. https://doi.org/10.1158/2159-8290.CD-12-0095.
61. Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, Sumer SO, Sun Y, Jacobsen A, Sinha R, Larsson E, Cerami E, Sander C, Schultz N. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci Signal. 2013; 6:pl1. https://doi.org/10.1126/scisignal.2004088.