
INTRODUCTION
Men with localized prostate cancer (PCa) are categorized in risk groups that reflect increasing treatment failure rates and prostate cancer specific mortality (e.g. low, intermediate or high) on the basis of pathologic Gleason score (GS), pre-treatment serum prostate specific antigen (PSA, measured in ng/mL) and clinical T-category (TNM staging system) [1-2]. Despite clinical classification, intermediate and high-risk PCa are characterized by significant heterogeneity with respect to clinical outcome after image-guided radiotherapy (IGRT) or radical prostatectomy, with biochemical relapse-free rates (bRFR) ranging from 40 to 80% [3]. This suggests that other pathologic, genomic or molecular factors could further triage patients into responders and non-responders by developing novel prognostic (treatment-independent) or predictive (treatment-dependent) biomarkers.
Malignant progression to the metastatic state is associated with increasing oncogene activation, tumor suppressor gene inactivation and increased genomic instability [4]. These genetic alterations may accumulate due to abnormal DNA damage responses (DDR) in the sensing and repair of DNA damage [5-6]. An abnormality in the DDR pathway could lead to cancer progression and metastases and/or affect the relative sensitivity of a tumour to radiotherapy, in which daily doses of photon radiation lead to DNA double-strand breaks (DSBs), and other DNA damages. Indeed, in vitro experiments support the hypothesis that differential prostate cancer cell radiosensitivity and genetic instability are correlated to differential DDR integrity [7-8].
The earliest and fundamental step in the process of the DDR is the recruitment of sensing and repair machinery to the sites of DNA damage [9]. The initial protein effectors of the DDR are the MRN complex (MRE11, RAD50 and NBS-1/Nibrin encoded by the MRE11A, RAD50 and NBN genes, respectively) and the DNA damage dependent, PI3-like kinases: ATM, ATR and DNA-PKcs (encoded by the ATM, ATR and PRKDC genes, respectively). ATM and NBN mutations have also been suggested to confer increased risk of prostate cancer [10-11] and disease aggression [12-13]. To date, a systematic analysis of copy number alterations (CNAs) within the initial DDR sensor genes (MRE11A, RAD50, NBN, ATM, ATR, PRKDC) as potential prognostic biomarkers has not been explored.
Herein, we collated the alterations in DDR sensors within localized prostate cancer, and show that copy number gains in the NBN locus is a novel adverse and independent factor for outcome following radiotherapy, but not following radical prostatectomy, in localized PCa. We discuss the use of NBN status to intensify therapy in the context of personalized PCa medicine.
RESULTS
CNAs of DDR genes are common in localized PCa
We initially determined whether DDR-sensing genes (MRE11A, RAD50, NBN, ATM, ATR and PRKDC) exhibited allelic gains or losses in our cohort. Upon evaluation of 139 pre-IGRT biopsies by aCGH, allelic changes were observed predominantly in NBN (15.1%), ATR (8.6%), PRKDC (7.9%) and ATM (5.8%). The surgical cohort showed a similar proportion of allelic changes in the NBN (11.6%) and ATM (6.5%) loci but higher in PRKDC (14.9%) and lower in ATR (1.3%) loci. RAD50 and MRE11A showed less than 5% of aberrations within each cohort (see Figures 1A/B). CNA distributions in these six DDR genes were not statistically different across any of the clinical prognostic variables of pretreatment PSA, T-category and GS (data not shown).
To determine how hits within the DDR sensor genes relate to each other, we explored the co-segregation and mutual exclusivity patterns of allelic changes in these six loci. To increase statistical power, we combined both treatment cohorts (pooled cohort) for this analysis. Five gene-pairs were co-altered in more patients than expected by chance (Fisher’s exact test, p < 0.001, Bonferroni correction): ATM-ATR, ATM-MRE11A, NBN-ATM, NBN-ATR and NBN-PRKDC (Supplemental Figure 1). No aberrations in DDR-sensing genes were mutually exclusive. We conclude that ATM and NBN alterations can associate with the other DDR alterations in PCa.
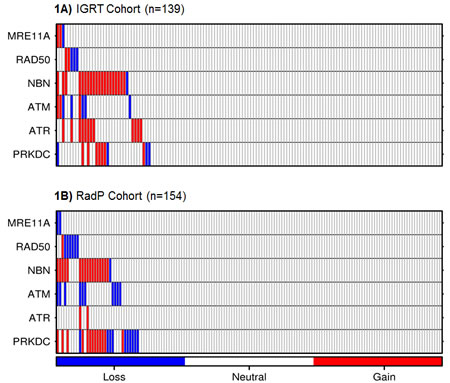
Figure 1: Heatmap of CNAs in DDR genes in tumour biopsies from the IGRT (1A) or RadP (1B) cohorts. Each bar represents a single patient. Gains and losses are represented by red and blue bars, respectively.
Patients with NBN CNAs show increased genomic instability
Considering that DDR genes play a role in genomic integrity, we used the percent genome aberration (PGA) to describe the instability within a patient’s tumour genome.
NBN exhibited the strongest association of a DNA damage response-sensing gene with PGA in both RadP and IGRT cohorts (see Figure 2). Using the pooled cohort, CNAs in all six DDR-sensing genes remained significantly correlated with higher PGA, independent of the clinical prognostic variables (pretreatment PSA, T-category and GS) (Supplemental Table 1).
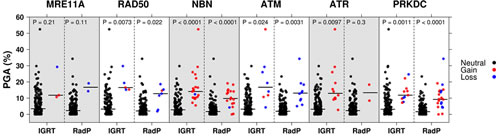
Figure 2: PGA as a function of DDR genes copy number status for IGRT and RadP cohorts. Each dot represents an individual patient with black, red and blue dots representing copy number neutral, loss, or gain respectively. The horizontal black line denotes the median PGA for each group. In both the IGRT and RadP cohorts, DDR genes CNAs associate with increased genomic instability (PGA) compared to patients with copy number neutral DDR gene status (p-values are shown; Mann-Whitney-U-test).
Patients with CNAs in DDR genes are more likely to relapse after IGRT
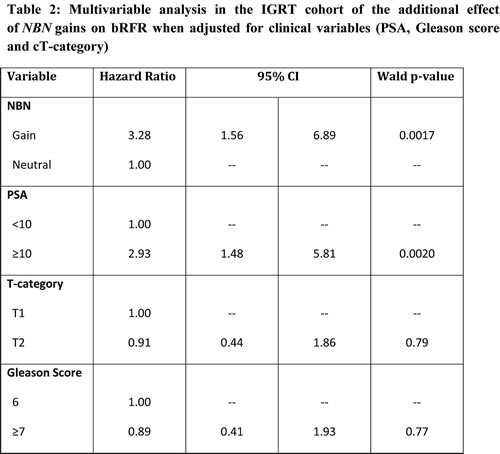
We next examined the impact of allelic changes in the DDR loci on clinical outcome based on biochemical recurrence (BCR). The clinical characteristics the IGRT cohort are shown in Table 1. At five years, 34 patients (24.5%) experienced biochemical failure as per the Phoenix definition. In this predominantly intermediate-risk group cohort, only PSA remained as a significant clinical prognostic factor for BCR based on univariate analysis (p-value = 0.0035) (see Supplemental Figure 2A). We tested DDR genes CNAs (gains or losses) for prognostic capability. Only NBN was prognostic for bRFR at 5 years on univariate analysis (bRFR 46% vs. 77%, log-rank p-value = 0.00067; HR= 3.35; 95% CI: 1.6 – 7.01; see Figure 3A). On multivariate analysis, NBN gains remained a significant independent predictor of bRFR after adjusting for PSA, T-category and GS (HR = 3.28, 95% CI 1.56 – 6.89, Wald p-value = 0.0017; see Table 2). NBN gains remained a significant predictor of bRFR even within low-risk patients (log rank p-value = 0.0037; Supplemental Figure 3); although the number of low-risk patients with NBN gains is low (4 patients).
Table 1: Clinical characteristics of IGRT and RadP cohorts
IGRT cohort |
RadP cohort |
How was used in the analysis |
|
N=139 (%) |
N=154 (%) |
||
T-category T1 T2 T3 |
50 (36%) 89 (64%) - |
79 (51.3%) 66 (42.9%) 9 (5.8%) |
T1 vs T2-T3 |
Gleason-score 5 6 7 8 9 |
- 34 (24.4%) 98 (70.5%) 7 (5%) - |
2 (1.3%) 82 (53.3%) 53 (34.4%) 11 (7.1%) 6 (3.9%) |
GS 5-6 vs 7-9 |
Pretreatment-PSA (ng/mL) Median Range <10 10-20 >20 |
8.0 0.9 – 33 92 (66.2%) 41 (29.5%) 6 (4.3%) |
6.265 1.15 – 506 115 (74.7%) 24 (15.6%) 15 (9.7%) |
PSA <10 vs > 10 |
NCCN Risk Group (Clinical staging) Low Intermediate High |
19 (13.6%) 107 (77.0%) 13 (9.4%) |
45 (29.2%) 79 (51.3%) 30 (19.5%) |
- |
Hormone therapy Neo-adjuvant |
35 (25.2%) |
7 (4.5%) |
- |
RT dose (Gy/fraction) 60/20 66/22 70/35 75.6/42 78/39 79.8/42 Mean equivalent dose= 75.9Gy |
15 (10.8%) 3 (2.1%) 1 (0.7%) 38 (27.3%) 3 (2.1%) 79 (56.8%) |
NA |
- |
BCR (5yrs) Yes No |
34 (24.5%) 105 (75.5%) |
34 (22.1%) 120 (77.9%) |
- |
Abbreviations: Gy=Gray, IGRT=image guided radiotherapy, BCR=biochemical recurrence, NA=not applicable. Mean RT equivalent dose was calculated using BED formula at 2 Gy daily fractions with an alpha/beta ratio of 2.0 for tumor response.
We next interrogated the surgically treated RadP cohort (n=154). At five years after surgery, thirty-four patients (22.1%) experienced biochemical failure. All three known clinical prognostic variables remained significant predictors of biochemical outcome (Supplemental Figure 2B). On univariate analysis, NBN gain did not have significant prognostic value (HR = 1.88, 95% CI 0.78 – 4.54, log-rank p-value = 0.15) (see Figure 3B). To further evaluate the potential prognostic role of NBN in this cohort, we explored the impact of mRNA abundance and its known partner KPNA2 on treatment outcome. In the 108 RadP patients with matched CNA and mRNA information, we found a significant association between NBN copy-number gain and NBN mRNA abundance (Student’s t-test, p-value = 0.0074; Supplemental Figure 4A). However, neither NBN nor KPNA2 mRNA abundance (stratified by median expression) correlated with bRFR (Supplemental Figure 4B/C).
Finally, we evaluated whether NBN gains added clinical value to the standard prognostic variables (PSA, T-category and GS) in our IGRT cohort using concordance-index (c-statistic), net reclassification index (NRI) and integrated discrimination improvement (IDI) (see ‘statistical analysis’ methods). The c-statistic of the clinical model (0.645, 95% CI 0.528-0.761) improved with the addition of NBN gains (0.673, 95% CI 0.562-0.785), indicating that the addition of NBN provides some improvement to the basic clinical model for predicting patient outcome. Similarly, the model including NBN assigned a higher risk to 28.5% of patients who experienced a BCR compared to the model without NBN; while the addition of NBN assigned a higher risk to 10.0% of patients censored at 5 years compared to the model without NBN. The net difference or NRI was therefore 18.5% (95% CI -1.7 – 36.5%, p-value = 0.11). It is also interesting that the average sensitivity of the model with NBN improved 5.0% from the basic clinical model, while the average specificity of the model increased marginally 1.4%. The integrated discrimination improvement (IDI), or net improvement in these quantities showed significant improvement at 6.45% (95% CI: 0.4% - 17.6%, p-value = 0.027). These three methods indicate that knowing the NBN status of a patient can be used to better predict biochemical failure following IGRT (see Supplemental Table 2).
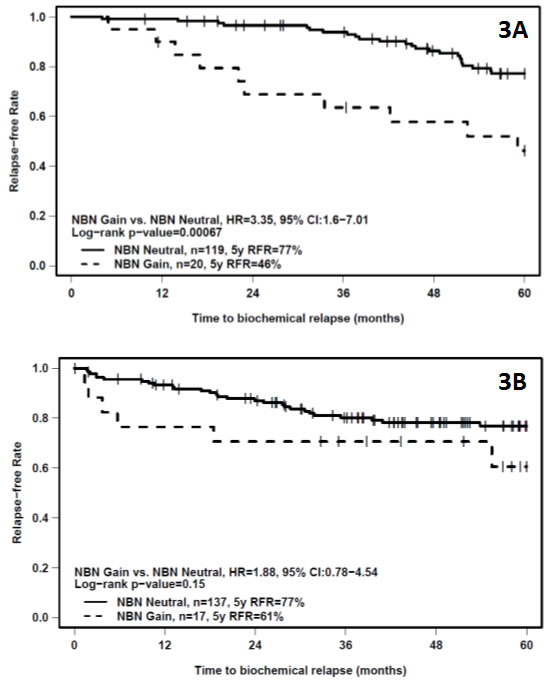
Figure 3: Kaplan-Meier plots of bRFR versus time to recurrence showing the effect of differential NBN CNA status (neutral vs. gain) in the IGRT (3A) or RadP (3B) treatment cohorts.
NBN status and expression associates with radioresistance in vitro
Given the potential additional role of NBN gains as a predictive factor for outcome after IGRT, we determined the correlation between absolute DDR gene mRNA expression and intrinsic clonogen radioresistance in a panel of prostate cell lines (see Supplemental Figure 5). Of the six DDR genes, only NBN gene expression showed a positive correlation with increasing clonogen radioresistance (e.g. increasing SF3Gy; r2 = 0.871, p-value of slope = 0.066; see Figure 4). This data suggest a role for NBN gains in intrinsic tumour resistance to radiotherapy, consistent with its poor prognostic impact in IGRT-treated patients.
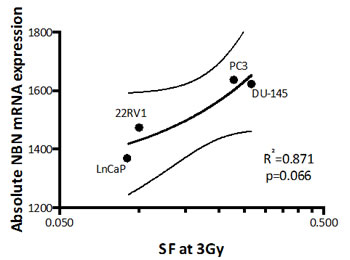
Figure 4: Normalized NBN mRNA abundance plotted against relative radioresistance (clonal surviving fraction after 3 Gy; SF3Gy) . Linear regression (continuous line) and 95% confidence interval (doted lines) are shown. A correlation is observed (R-square = 0.871) with a significant trend in the p-value of the slope (p=0.066). For PrEC cell line, only clonogenic survival fraction after 2Gy (SF2Gy) was available, therefore it was not included in the linear regression calculation.
DISCUSSION
To our knowledge, this is the first study to describe the frequency and importance of alterations in genes that are initial sensors within the DDR pathways within PCa patients. We particularly were interested in the potential effect of abnormalities in genes involved in the initial DNA damage response-sensing for IGRT patients who receive daily doses of DNA DSBs as part of their treatment. Our main finding, that an altered DDR is predictive for IGRT outcome, is important given our results are based on a genomic test using pre-treatment biopsies which are used to direct primary management decisions of localized PCa. This prognostic effect was not observed in surgery patients in whom DDR responses are probably irrelevant.
If our IGRT-based observation with NBN gains is validated in other independent and/or large prospective cohorts using aCGH or FISH (fluorescence in situ hybridization, using a NBN-specific probe), this could become the first predictive test for selecting patients to undergo surgery (rather than IGRT alone) or undergo IGRT treatment intensification (e.g. further radiotherapy dose-escalation or the additional use of systemic agents).
We confirmed our hypothesis that aberrations in DDR genes would associate with unstable PCa genomes. At present, it is not known whether NBN gains, are drivers or passengers within PCa progression. It should be noted that the NBN locus in on chromosome 8q which is one of the most altered areas of the genome in our patient cohort, and therefore NBN gains may strongly relate to other gene loci abnormalities (e.g. c-Myc). We cannot yet conclude that NBN’s effect is gene-specific (e.g. prognostic at the mRNA and/or protein level) or rather a reflection of an important role as a surrogate of genomic instability; results from ongoing multimodal genomic studies (ICGC and TCGA consortia) may help clarify NBN gains linkage to specific CNAs, mutations or gene rearrangements. Furthermore, a recent publication form the TCGA consortia in colorectal cancer has shown that many CNA-driven increases in mRNA levels do not translate into greater abundance of the corresponding protein [14]. Future research is required to characterize NBN gains functional effect in the DDR of PCa tumours.
Based on previous work suggesting that germline NBN mutations predispose to more aggressive phenotypes of disease [13], we speculate that failures in IGRT-treated patients could be related to radioresistance and increased NBN-associated genomic instability that leads to greater capacity for micrometastatic spread. Neither NBN gain nor NBN mRNA expression (or expression of its partner, KPNA2) was prognostic in the surgery cohort; consistent with previous publications on Nibrin expression in surgically treated prostate cancer patients [15-16]. The only current predictive DDR biomarker for radiotherapy response is MRE11 protein expression in bladder cancer [17], which has been independently validated [18]. However, MRE11A alterations were not frequent (nor prognostic) in our PCa cohorts.
We acknowledge a number of limitations to the current study. First, our biomarker and outcome analyses are based on a single TRUS-guided biopsy to the dominant lesion; therefore the known intraprostatic heterogeneity and differential biology of multifocal disease might be underestimated [19]. However, the biopsies were taken from the dominant (index) lesion in each patient, which usually correlates with the majority of local treatment failures [20]. Also, our radiotherapy and surgical cohorts differ slightly in the distribution of clinical prognostic features and duration of follow-up, which could reduce the power to detect significant results in the DNA-based prognosis analyses for the surgical cohort. However, as stated above, protein expression for NBN is not prognostic in other surgical series [15-16].
In conclusion, our study suggests that gain of NBN is a potentially novel predictive factor of IGRT failure, and patients should consider surgery if their tumour harbours this CNA. If validated, this would be the first outcome-associated predictive biomarker for local treatment in localized PCa. IGRT patients whose tumours harbour NBN gains should be assessed for additional systemic therapies to improve outcome [21].
MATERIALS AND METHODS
Study Patient Cohorts
DNA samples for array comparative genomic hybridization (aCGH) analysis were derived from pre-treatment frozen biopsies in patients undergoing radical image-guided radiotherapy (REB#00-0443-C; the Princess Margaret Cancer Centre), as previously described [22]. The clinical target volume (CTV) encompassed the prostate gland alone. The planning target volume (PTV) was defined by a 10 mm expansion around the CTV except posteriorly in which the margin was 7 mm. All patients were treated with 6-field conformal or intensity modulated image-guided radiotherapy. The radiotherapy dose was escalated over the period of accrual in a series of separate Phase I-II studies (see Table 1). Some patients received short-term, neoadjuvant and concurrent anti-androgen treatment (bicalutamide 150 mg po od). None of the patients received LHRH agonists/antagonists nor adjuvant androgen deprivation therapy. Staging bone scans and computed tomography (CT) were not routinely carried out on low- and intermediate-risk disease patients. Patients were followed at 4-6 monthly intervals after completing treatment with clinical examination and PSA measurement. Additional tests and the management of patients with recurrent disease were at the discretion of the treating physician.
Sufficient tumour for aCGH analysis was identified in 139 patients who also had long term follow-up information pertaining to biochemical outcome (see Table 1). The median follow-up of surviving patients was 7.9 years following the start of treatment, with 62 (44.6%) patients experiencing biochemical failure during follow-up.
We interrogated the publicly-available Memorial Sloan Kettering Cancer Center Prostate Cancer database [23] for aCGH data pertaining to 154 PCa tumours of patients treated by radical prostatectomy (RadP). Gene-level CNA calls were collapsed to losses or gains. Patients were placed into low-, intermediate-, and high-risk NCCN categories [2] based on pre-surgery clinical staging. (see Table 1). The median follow-up for this cohort was 4.8 years, with 38 patients (24.7%) experiencing biochemical failure during the entire follow-up. For selected RadP cohort patients (108 out of 154), we also used this database to compare mRNA abundance to allelic gain or loss in DDR genes.
Array Comparative Genomic Hybridization (aCGH) for CNA Calls
For the IGRT cohort, DNA extraction, labeling and hybridization onto arrays containing 26,819 bacterial artificial chromosome (BAC)-derived amplified fragment pools was carried out as described previously [24]. The resulting data set was normalized using a stepwise normalization procedure [25]. This data is available in NCBI’s Gene Expression Omnibus with accession number GSE41120. Areas of aberrant copy number were identified using a robust Hidden Markov Model [26] and classified as either loss, neutral or gain for all clones. The liftOver tool from UCSC was used to map the copy number segments to the hg19 human genome build. Fragments overlapping centromeres, telomeres, or other gaps in the hg18 build were trimmed conservatively (regions were shortened rather than elongated). To generate contiguous CNA regions, probe-based CNA calls were collapsed with neighbouring probes within the same chromosome with the same copy number. CNA regions with only one supporting probe were removed. In addition, any CNAs found entirely within centromeres or telomeres, as defined by the UCSC ‘gap’ table, were removed. CNA regions were intersected with a merged and collapsed version of the RefSeq gene annotation (GRCh37/hg19) to generate gene-based CNA calls. This gene list was further filtered to match the published gene list from the radical prostatectomy (RadP) cohort (n = 17,603).
For both IGRT and RadP cohorts, percent genome aberration (PGA) was used as a measure of genomic instability and defined as the cumulative size of the genetic alterations found in each patient DNA sample divided by the total size of the human genome as previously described [27]. To account for the fact that a gene may be found in a large aberration (which can skew PGA values), we removed chromosomal effects from PGA-association calculations for each gene. For example, for MRE11A (which is on chromosome 11), we calculated PGA using chromosomes 1-10 and 12-22 only. Thus, if MRE11A is truly associated with PGA, the effect should be observed throughout the genome and unaffected by this adjustment. The raw PGA and the adjusted-PGA are highly correlated (Supplemental Figure 6) and as such, we show adjusted-PGA values throughout the manuscript.
mRNA Expression and In Vitro Radiosensitivity Assays
To corroborate the findings at the CNA level in the surgery cohort, mRNA abundance data for the RadP cohort was also considered for candidate genes. We focused on the 108 patients with primary disease that were previously studied at the mRNA and CNA level [23]. Raw CEL files were downloaded from GEO (accession GSE21034) and normalized with RMA [28], as previously described [29].
Clonogenic assay data on radiation surviving fraction at 3Gy (SF3Gy) for PCa cells (PrEC, LNCaP, 22RV1, PC-3, and DU-145) were determined as previously published[22]. All cell lines were obtained from American Type Culture Collection and authenticated by short tandem repeat DNA profiling. Nuclear and cytoplasmic RNA was extracted and assayed with the NanoString nCounter platform. Gene-specific mRNA abundance was determined for MRE11A, RAD50, NBN, ATM, ATR and PRKDC genes, housekeeping genes (β2-microglobulin, β-actin, and GAPDH). As recommended, positive control probes (6), geometric mean of housekeeping gene expression and negative control probes (8) were used to normalize the data.
Statistical Analyses
Patients with copy number gain and/or loss in each of the six DDR genes (MRE11A, RAD50, NBN, ATM, ATR and PRKDC) were compared to patients with copy neutral status for distribution of clinical variables of interest and treatment outcome. Chi-squared tests were used to compare the proportion of patients between CNA groups for clinical variables (PSA, T-category and GS). Association of CNA status of DDR-sensing genes with PGA was assessed by Student’s t-tests, and a multiple linear regression model was used to evaluate the effect of DDR genes on PGA adjusting for clinical variables (PSA, T-category and GS). Fisher’s exact test was used to assess mutual exclusivity and co-segregation of pairs of DDR-sensing genes. Student’s t-tests were calculated to determine the association between copy-number and mRNA abundance. Linear regression was used to describe the relationship between DDR genes mRNA abundance and clonogen radioresistance in vitro.
The primary clinical outcome variable for both cohorts was time to biochemical recurrence, censored at five years. Cox proportional hazards regression models were used for univariate and multivariate models to assess the prognostic ability of DDR genes on biochemical relapse-free rate adjusting for known clinical prognostic factors. (PSA <10 vs. >=10; T1 vs. T2-T3; Gleason score 5-6 vs. 7 or higher). The proportional hazards assumption was checked by investigating the Schoenfeld residuals. Five-year relapse-free rates were estimated using the Kaplan-Meier method. The additional utility of any copy number alteration relative to clinical parameters was described by the use of c-statistic, net reclassification index (NRI), and integrated discrimination improvement (IDI) [30]. The c-statistic presents the probability that a classification model gives a higher score to a patient that experiences a biochemical relapse by 5 years compared to a patient censored at five years. Thus, in the clinical setting, adding one biomarker to a validated clinical model may not change the classification of an event case to a non-event case. NRI represents another informative measure of added value, which considers whether adding NBN to the basic clinical model increases the model’s risk scores for those patients that experienced a biochemical recurrence or decreases the risk scores for patients that were censored at 5 years, respectively. In turn, IDI calculates the improvement in sensitivity without changing specificity by calculating the difference in survival probability for each patient from the clinical model to the model with NBN gain incorporated.
All tests are two-sided and statistical significance was assessed using p<0.05. Multiple testing correction was applied where appropriate with the Bonferroni method, as specified in the text. All statistical analyses were performed using the open-source software R (v3.0-1). The survival package (v2.37-4) was used to plot Kaplan-Meier curves and run Cox proportional-hazards regression models; the lattice (v0.20-15) and latticeExtra (v0.6-24) packages were used for data visualization.
ACKNOWLEDGMENTS
Supported by grants to RGB from Prostate Cancer Canada (CPC-GENE projects with monies from the MOVEMBER Foundation), the Ontario Institute for Cancer Research, the PMH Foundation, and the Canadian Foundation for Innovation grant to the STTARR Innovation Facility. This research was funded in part by the Ontario Ministry of Health and Long Term Care. The views expressed do not necessarily reflect those of the Ontario Ministry of Health and Long Term Care. RGB is a Canadian Cancer Society Research Scientist. This study was conducted with the support of the Ontario Institute for Cancer Research to PCB through funding provided by the Government of Ontario. This work was supported by Prostate Cancer Canada and is proudly funded by the Movember Foundation - Grant #RS2014-01. Dr. Boutros was supported by a Terry Fox Research Institute New Investigator Award.
AB received a Clínica Alemana de Santiago Educational Award, Canadian Urologic Oncology Group (CUOG) Research Award, the Terry Fox Foundation Excellence in Radiation Research for the 21st Century (EIRR21) at CIHR scholarship award, and ASCO-GU Merit Award.
EL is a recipient of a Canadian Institute for Health Research Doctoral Research Award.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Siegel R, Naishadham D, Jemal A. Cancer statistics, 2013. CA Cancer J Clin. 2013; 63: 11–30.
2. Mohler JL, Kantoff PW, Armstrong AJ, Bahnson RR, Cohen M, D’Amico AV, Eastham JA, Enke CA, Farrington TA, Higano CS, Horwitz EM, Kawachi MH, Kuettel M, et al. Prostate cancer, version 1.2014. J Natl Compr Canc Netw. 2013; 11: 1471–9.
3. Nichol AM, Warde P, Bristow RG. Optimal treatment of intermediate-risk prostate carcinoma with radiotherapy: clinical and translational issues. Cancer. 2005; 104: 891–905.
4. Hanahan D, Weinberg RA. Hallmarks of Cancer: The Next Generation. Cell. 2011; 144: 646–74.
5. Haffner MC, Aryee MJ, Toubaji A, Esopi DM, Albadine R, Gurel B, Isaacs WB, Bova GS, Liu W, Xu J, Meeker AK, Netto G, De Marzo AM, et al. Androgen-induced TOP2B-mediated double-strand breaks and prostate cancer gene rearrangements. Nat Genet. 2010; 42: 668–75.
6. Swanson TA, Krueger SA, Galoforo S, Thibodeau BJ, Martinez AA, Wilson GD, Marples B. TMPRSS2/ERG fusion gene expression alters chemo- and radio-responsiveness in cell culture models of androgen independent prostate cancer. Prostate. 2011; 71: 1548-58.
7. El-Awady RA, Dikomey E, Dahm-Daphi J. Radiosensitivity of human tumour cells is correlated with the induction but not with the repair of DNA double-strand breaks. Br J Cancer. 2003; 89: 593–601.
8. Fan R, Kumaravel TS, Jalali F, Marrano P, Squire JA, Bristow RG. Defective DNA strand break repair after DNA damage in prostate cancer cells: implications for genetic instability and prostate cancer progression. Cancer Res. 2004; 64: 8526–33.
9. Ciccia A, Elledge SJ. The DNA damage response: making it safe to play with knives. Mol Cell. 2010; 40: 179–204.
10. Meyer A, Wilhelm B, Dörk T, Bremer M, Baumann R, Karstens JH, Machtens S. ATM missense variant P1054R predisposes to prostate cancer. Radiother Oncol. 2007; 83: 283–8.
11. Cybulski C, Górski B, Debniak T, Gliniewicz B, Mierzejewski M, Masojć B, Jakubowska A, Matyjasik J, Złowocka E, Sikorski A, Narod SA, Lubiński J. NBS1 is a prostate cancer susceptibility gene. Cancer Res. 2004; 64: 1215–9.
12. Silva J, Teixeira AL, Lobo F, Maurício J, Medeiros R. DNA repair system and prostate cancer progression: the role of NBS1 polymorphism (rs1805794). DNA Cell Biol. 2012; 31: 1182–6.
13. Cybulski C, Wokołorczyk D, Kluźniak W, Jakubowska A, Górski B, Gronwald J, Huzarski T, Kashyap A, Byrski T, Dębniak T, Gołąb A, Gliniewicz B, Sikorski A, et al. An inherited NBN mutation is associated with poor prognosis prostate cancer. Br J Cancer. 2013; 108: 461–8.
14. Zhang B, Wang J, Wang X, Zhu J, Liu Q, Shi Z, Chambers MC, Zimmerman LJ, Shaddox KF, Kim S, Davies SR, Wang S, Wang P, et al. Proteogenomic characterization of human colon and rectal cancer. Nature. 2014; Jul 20: [Epub ahead of print].
15. Mortezavi A, Hermanns T, Seifert HH, Baumgartner MK, Provenzano M, Sulser T, Burger M, Montani M, Ikenberg K, Hofstädter F, Hartmann A, Jaggi R, Moch H, et al. KPNA2 expression is an independent adverse predictor of biochemical recurrence after radical prostatectomy. Clin Cancer Res. 2011; 17: 1111–21.
16. Grupp K, Boumesli R, Tsourlakis MC, Koop C, Wilczak W, Adam M, Sauter G, Simon R, Izbicki JR, Graefen M, Huland H, Steurer S, Schlomm T, et al. The prognostic impact of high Nijmegen breakage syndrome (NBS1) gene expression in ERG-negative prostate cancers lacking PTEN deletion is driven by KPNA2 expression. Int J Cancer. 2014; Feb 8: [Epub ahead of print].
17. Choudhury A, Nelson LD, Teo MT, Chilka S, Bhattarai S, Johnston CF, Elliott F, Lowery J, Taylor CF, Churchman M, Bentley J, Knowles MA, Harnden P, et al. MRE11 expression is predictive of cause-specific survival following radical radiotherapy for muscle-invasive bladder cancer. Cancer Res. 2010; 70: 7017–26.
18. Laurberg JR, Brems-Eskildsen AS, Nordentoft I, Fristrup N, Schepeler T, Ulhøi BP, Agerbaek M, Hartmann A, Bertz S, Wittlinger M, Fietkau R, Rödel C, Borre M, et al. Expression of TIP60 (tat-interactive protein) and MRE11 (meiotic recombination 11 homolog) predict treatment-specific outcome of localised invasive bladder cancer. BJU Int. 2012; 110: E1228–36.
19. Fraser M, Berlin A, Bristow RG, van der Kwast T. Genomic, pathological, and clinical heterogeneity as drivers of personalized medicine in prostate cancer. Urol Oncol. 2014; Apr 22: [Epub ahead of print].
20. Chopra S, Toi A, Taback N, Evans A, Haider MA, Milosevic M, Bristow RG, Chung P, Bayley A, Morton G, Vesprini D, Warde P, Catton C, et al. Pathological predictors for site of local recurrence after radiotherapy for prostate cancer. Int J Radiat Oncol Biol Phys. 2012; 82: e441–8.
21. Bristow RG, Berlin A, Dal Pra A. An arranged marriage for precision medicine: hypoxia and genomic assays in localized prostate cancer radiotherapy. Br J Radiol. 2014; 87: 20130753.
22. Locke JA, Zafarana G, Ishkanian AS, Milosevic M, Thoms J, Have CL, Malloff CA, Lam WL, Squire JA, Pintilie M, Sykes J, Ramnarine VR, Meng A, et al. NKX3.1 haploinsufficiency is prognostic for prostate cancer relapse following surgery or image-guided radiotherapy. Clin Cancer Res. 2012; 18: 308–16.
23. Taylor BS, Schultz N, Hieronymus H, Gopalan A, Xiao Y, Carver BS, Arora VK, Kaushik P, Cerami E, Reva B, Antipin Y, Mitsiades N, Landers T, et al. Integrative genomic profiling of human prostate cancer. Cancer Cell. 2010; 18: 11–22.
24. Ishkanian AS, Mallof CA, Ho J, Meng A, Albert M, Syed A, van der Kwast T, Milosevic M, Yoshimoto M, Squire JA, Lam WL, Bristow RG. High-resolution array CGH identifies novel regions of genomic alteration in intermediate-risk prostate cancer. Prostate. 2009; 69: 1091–100.
25. Khojasteh M, Lam WL, Ward RK, MacAulay C. A stepwise framework for the normalization of array CGH data. BMC Bioinformatics. 2005; 6: 274.
26. Shah SP, Xuan X, DeLeeuw RJ, Khojasteh M, Lam WL, Ng R, Murphy KP. Integrating copy number polymorphisms into array CGH analysis using a robust HMM. Bioinformatics. 2006; 22: e431–9.
27. Zafarana G, Ishkanian AS, Malloff CA, Locke JA, Sykes J, Thoms J, Lam WL, Squire JA, Yoshimoto M, Ramnarine VR, Meng A, Ahmed O, Jurisca I, et al. Copy number alterations of c-MYC and PTEN are prognostic factors for relapse after prostate cancer radiotherapy. Cancer. 2012; 118: 4053–62.
28. Irizarry RA, Hobbs B, Collin F, Beazer-Barclay YD, Antonellis KJ, Scherf U, Speed TP. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. Biostatistics. 2003; 4: 249–64.
29. Cortese R, Kwan A, Lalonde E, Bryzgunova O, Bondar A, Wu Y, Gordevicius J, Park M, Oh G, Kaminsky Z, Tverkuviene J, Laurinavicius A, Jankevicius F, et al. Epigenetic markers of prostate cancer in plasma circulating DNA. Hum Mol Genet. 2012; 21: 3619–31.
30. Pencina MJ, D’ Agostino RB, D’ Agostino RB, Vasan RS. Evaluating the added predictive ability of a new marker: from area under the ROC curve to reclassification and beyond. Statist Med. 2008; 27: 157–72; discussion 207–12.