
INTRODUCTION
Good health in aging is compromised by a progressive decline in immune function (immunosenescence) which results in more severe consequences of viral and bacterial infections and decreased response to vaccines [1, 2]. Numerous studies in older adults have revealed a profound impact on the phenotype and functions of immune cells in aging, which in the innate immune system, lead to dysregulation of responses with inappropriate elevations and decreases in immune responses [3–5]. This multifactorial immune deterioration--termed “inflamm-aging”--is comprised of a state of low-grade, chronic inflammation characterized by broad dysregulation of cellular stress pathways, oxidative stress, mitochondrial dynamics and autophagy, chronic viral infections, obesity, DNA damage response, and impaired clearance of damaged cells [6–8].
Innate immune cells sense diverse microorganisms through the evolutionarily conserved signaling pattern recognition receptors, Toll-like receptors (TLRs), that recognize highly conserved components of microbial pathogens and mediate the earliest host response to infection [9, 10]. Additional recognition mechanisms include the cytoplasmic retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs), cytosolic DNA sensors, and nucleotide-binding oligomerization domain (NOD)-like receptors (NLRs) that trigger inflammasome activation [11, 12]. With age, expression and function of TLRs, RIG-I, and NLRs declines in several cell lineages including monocytes, dendritic cells (DCs) and polymorphonuclear leukocytes (PMN) [4, 13–20]. Age-associated decreases in pattern recognition receptor expression and downstream signaling cascades result in reduced cytokine production, particularly those related to interferon-dependent signaling, and are associated with lower vaccine responsiveness [1, 4, 21–24].
The powerful TLR activation pathways are regulated to limit inflammation. Indeed, successful immune responses represent a balance of pro- and anti-inflammatory processes. Recent studies have identified the TAM family (Tyro 3, Axl, and Mer) of receptor tyrosine kinases as negative regulators of TLR-mediated immune responses that broadly inhibit both TLR and TLR-induced cytokine receptor cascades to limit inflammation [25–29]. The importance of TAMs in immune activation is illustrated by the observation that reduced levels of TAMs in humans and in a TAM-deficient mouse model are associated with susceptibility to autoimmune disease and higher or chronic inflammation [27, 30, 31]. Further, TAMs play a key role in mediating autophagy [26, 27, 32], and the reduced efficiency of autophagy in aging has been shown to contribute to accumulation of damaged proteins in cells [33].
In previous studies of aging, we have shown dysregulation of immune responses with a paradoxical increase in levels of pro-inflammatory cytokines such as IL-6 and TNF-α and a decrease in anti-viral interferon responses [1, 3, 5, 34]. In examining the role of negative regulation in age-related immune deficiencies, we showed higher expression of Axl in monocyte-derived DCs from older individuals after infection with West Nile virus [21]. These data suggested that enhanced negative signaling in aging may contribute to a more severe outcome in viral infection. The current studies aim towards a fuller molecular understanding of the effects of aging on expression and function of TAMs and may provide insights to immune mechanisms involved in chronic inflammation that contribute to multiple immune dysfunctions in aging.
RESULTS
TAM receptor expression is elevated in aging
To identify effects of aging on TAM receptors, we recruited younger (age 21–30 years, N = 40) and older (age 65–89, N = 50) adults and assessed TAM expression and function. Participants were healthy at the time of sampling but varied significantly by age group as expected for number chronic conditions, and prescription and over the counter medications (Table 1). We quantified surface expression of TAM receptors as a measure of potential functional TAM activity between age groups. Flow cytometry of CD14+, CD11c+ monocytes revealed highly variable expression among healthy donors of both age groups, particularly for Tyro3 and Axl, with some donors expressing almost no Axl or Tyro3 at all (Figure 1 and Supplementary Figure 1). In contrast, Mer was expressed in all donors and was less variable. The surface expression (% positive) of all three TAM receptors was elevated in monocytes of older donors (Tyro3: Young 26.8 + 7.7 vs Old 54.5 + 7.8, p = 0.02; Axl: Young 16.3 + 6.5 vs Old 40.4 + 6.7, p = 0.02; Mer: Young 18.6 + 2.4 vs Old 27.6 + 2.4; p = 0.03). The mean fluorescence intensity (MFI) was not different between age groups.
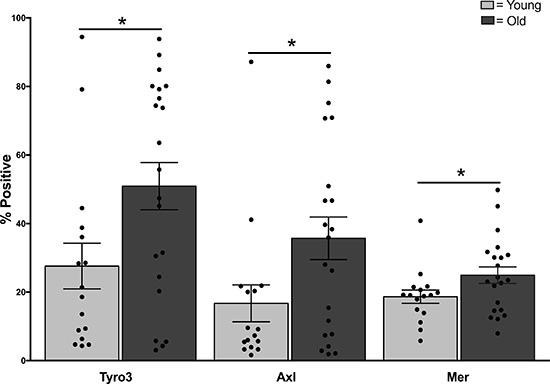
Figure 1: Elevated expression of TAM receptors in monocytes of older adults. PBMCs from younger and older adults were labeled for TAM receptors Tyro3, Axl, and Mer and expression was quantified by LSR-II flow cytometry as described [39]. Data shown is percent expression of TAM receptor in CD14+, CD11c+ monocytes, n = 16 young, N = 21 old, * indicates p < 0.02 using multivariate Generalized Linear Models.
Table 1: Human participants
Patient Characteristics |
Young |
Older |
P-value |
|---|---|---|---|
(N = 90) |
(N = 40) |
(N = 50) |
|
Age (years) range (22–89), mean (SD) |
25.8 (2.2) |
73.6 (6.4) |
< 0.001 |
Female, n (%) |
27 (67.5) |
23 (46.0) |
0.055 |
White Race, n (%) |
25 (62.5) |
46 (92.0) |
0.001 |
Number of Comorbid Conditions, mean (SD) |
0.9 (0.9) |
2.8 (1.8) |
< 0.001 |
Myocardial Infarction, n (%) |
0 (0.0) |
3 (6.0) |
0.249 |
Congestive Heart Failure, n (%) |
0 (0.0) |
1 (2.0) |
1.000 |
Coronary Artery Disease, n (%) |
0 (0.0) |
9 ( 18.0) |
0.004 |
Cardiac Arrhythmia, n (%) |
0 (0.0) |
4 ( 8.0) |
0.124 |
Hypertension, n (%) |
2 (5.0) |
31 ( 62.0) |
< 0.001 |
Peripheral Vascular Disease, n (%) |
0 (0.0) |
2 ( 4.0) |
0.498 |
Chronic Obstructive Pulmonary Disease, n (%) |
0 (0.0) |
1 (2.0) |
1.000 |
Peptic Ulcer Disease, n (%) |
1 (2.5) |
2 (4.0) |
1.000 |
Number of Prescription Medications, Age mean (SD) |
0.7 (0.8) |
3.5 ( 2.7) |
< 0.001 |
Statin Medications, n (%) |
0 (0.0) |
28 ( 56.0) |
< 0.001 |
Number of OTC Medications, mean (SD) |
1.7 (1.2) |
2.9 ( 1.3) |
< 0.001 |
Aspirin, n (%) |
0 ( 0.0) |
32 (64.0) |
< 0.001 |
aProbability for Student t-test for continuous measures, Fisher Exact test for categorical measures.
Effects of aging on activation of Mer
Upon activation, TAM receptors initiate engulfment of apoptotic cells through Ca+2-dependent, phosphatidyl serine (PtdSer) mediated binding of gamma-carboxyglutamic (Gla)-domains of ligands protein S, a plasma glycoprotein usually associated with a role in the coagulation cascade, or the structurally related protein growth arrest-specific gene 6 (Gas6) [26, 35]. The three TAM receptors Tyro3, Axl, and Mer show distinct features and ligands [35, 36]. To assess the functional consequence of the age-related elevation in TAM expression, we focused on Mer, which is expressed by all donors, and highly expressed in macrophages suggesting a key role in tissue particularly relevant for apoptotic cell clearance [37–39]. Adherence-purified monocytes were incubated overnight in the absence of serum to decrease the contribution of Mer ligand Protein S from serum in culture medium. Cells were treated with PtdSer liposomes containing Protein S to engage Mer receptors [35, 36, 40]. We detected a transient increase in phosphorylated forms of Mer at 10 min which declined by 30 min and was not different between age groups (Supplementary Figure 2A). This is consistent with a Protein S-dependent phosphorylation of Mer following treatment in vitro with gas6 which has been shown previously to be maximal at 5 min [41]. Treatment with liposomes or Protein S alone did not lead to stimulation of Mer pathways (data not shown).
Notably, treatment with PtdSer-Protein S liposomes led to increased phosphorylation of the serine/threonine kinase AKT in monocytes from younger adults at 30 min (Figure 2, p = 0.005), but levels of phospho-AKT in older adults were not increased (Figure 2, p = 0.47). This age-dependent difference in signaling mediator AKT is significant for both the higher levels in younger adults and also the change over time following stimulation (Figure 2, p = 0.003). In the same samples, reduction in levels of signaling mediators pSTAT1 and p38 was detected at 10 and 30 min following treatment with PtdSer-Protein S liposomes, however the decreases noted were not different between age groups (Supplementary Figure 2B, 2C).
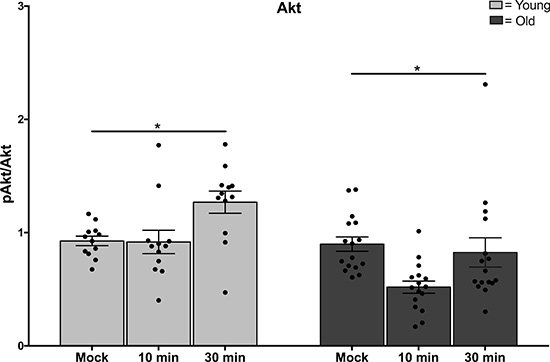
Figure 2: Functional changes in Mer signaling in aging. Monocytes from younger and older adults were incubated overnight in the absence of serum before treatment with 0.5 mM phosphatidylserine liposomes containing 100 nM Protein S. Cells were harvested at the time points indicated and lysates were processed for Immunoblot. Data shown is densitometry for N = 12 young, N = 16 old, * indicates p = 0.003 in multivariate Generalized Linear Models.
Phosphorylation of AKT in monocytes would be expected to lead to elevated transcription of downstream mediators of inflammatory responses. In correlation with the levels of phospho-AKT, treatment of monocytes from younger adults with PtdSer-Protein S liposomes led to increased levels of SOC-1 and IL-8, with a significant change from baseline to 2 hours in younger adults which was not seen in monocytes from older adults (Figure 3A, 3B, p = 0.03). Further, we noted age-related differences in IL-1β responses to PtdSer-Protein S liposomes. Notably, the decrease in levels in younger adults from baseline over 2 hours of treatment and increase in levels in the older adults over the 2 hours of treatment result in a significant difference between younger and older adults (Figure 3C, p = 0.03). In addition, for a subset of subjects with data collected at 8 hours, we noted extension of the same trajectory (data not shown). Age-related differences were not noted in levels of IFNβ and TNF (not shown), suggesting the kinetics of activation of those targets were not captured by our study.
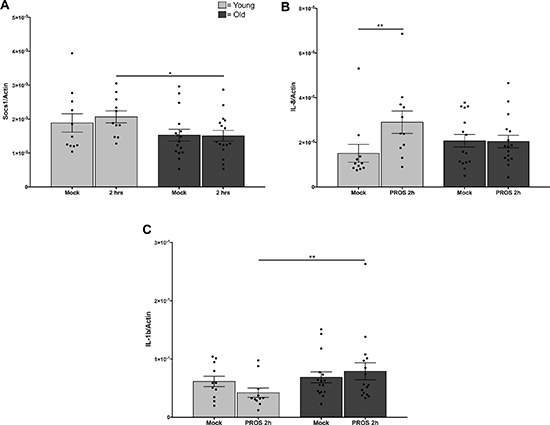
Figure 3: Reduced responses to Mer signaling in monocytes from older adults. Monocytes from younger and older adults were incubated overnight in the absence of serum before treatment with 0.5 mM phosphatidylserine liposomes containing 100 nM Protein S. RNA was extracted for Q-PCR. Data shown is (A) Soc1, (B) IL-8, and (C) Il-1β expressed as a ratio to cellular b-actin from cells untreated or stimulated for 2 hours from N = 12 young, N = 16 old, * indicates p = 0.03 using Type 3 Test of Fixed Effects.
DISCUSSION
The elevated expression of TAM receptors in monocytes from older adults has important implications for dysregulation of immune responses in aging--in particular as the Mer pathway is critical for clearance of apoptotic cells that contribute to inflammation in aging [37, 38]. We have previously used paired samples of monocytes and mature macrophages from healthy adults to show significant upregulation of Mer along the maturation of monocytes to macrophages, suggesting that levels of Mer are highest in tissue, which is relevant for its role in clearance of apoptotic cells [39]. Here we show age-related dysregulated activation of the Mer pathway following binding of Protein S, leading to impaired signaling through AKT in monocytes of older adults. Our findings suggest a possible role for phosphatidylinositol-3 kinase (PI3K) in this age-related deficiency in phosphorylation of AKT. This is consistent both with the established role for PI3K/AKT pathway in Mer activation [42], previous reports of dysregulation of PI3K in aging [43–45], as well as the absence of age-associated changes in Mer stimulated signaling through STAT-1 in the current study (Figure 4).
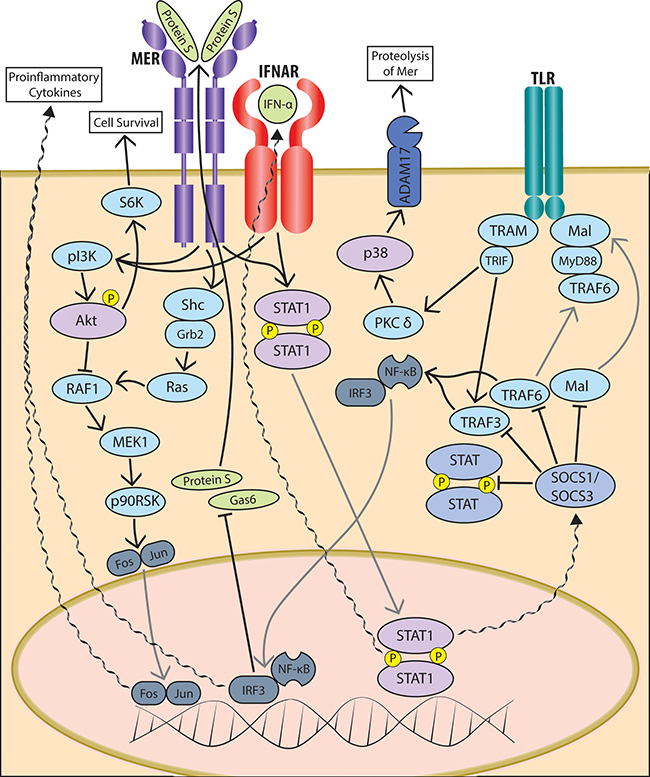
Figure 4: Schematic of TAM pathway signaling in innate immune cells. Schematic representation of some of the most relevant signalling intermediates in the intracellular signaling cascades triggered in response to ligation of TAM family member Mer by its ligand Protein S.
TAM pathways are important in a range of biological functions. Unexpectedly, TAMs have been shown to provide an immune evasion mechanism for certain enveloped viruses through “apoptotic mimicry”, in which they enhance infectivity by bridging virion phosphatidyl serine via gas6 binding to increase uptake through TAM receptors [46–48]. Thus, the elevated levels of TAMs in innate immune cells in aging may contribute to susceptibility to certain viral infections [48]. This may be important in response to West Nile virus, for which aging is a significant risk factor [34], although recent studies with the closely related Zika virus suggest this may be specific to particular viruses [49]. Studies of immune regulation in tumor progression have highlighted a role for TAMs, with inhibition of TAM tyrosine kinase pathways being actively investigated as a novel cancer therapeutic [50–52]. In addition, advances in our understanding of host genetic variation such as polymorphisms in Mer, which are associated with susceptibility to autoimmune disease, or genes related to aging [53–57], may offer insight into mechanisms to regulate TAM pathways. Taken together, improved understanding of TAM pathways may reveal avenues for modulation of chronic tissue inflammation noted in aging and may present targeted therapeutic approaches to address immune deficits in aging.
MATERIALS AND METHODS
Study subjects
Heparinized blood from healthy volunteers (younger 21–30, older > 65) was obtained with written informed consent under an IRB protocol approved annually by the Human Investigations Committee of Yale University. Donors had no acute illness and took no antibiotics or nonsteroidal anti-inflammatory drugs within one month of enrollment. Enrollment of these adults was conducted at flu vaccination clinics in two consecutive years (2013–2014) and demographic characteristics were collected at enrollment (Table 1). Self-reported information comprised demographic data, height, weight, medications, and comorbid conditions; immunocompromised individuals as defined previously were excluded [15]. The proportion of women or race was not statistically different between age groups. As it was not possible to assess each donor for each cell-based assay, samples were randomly chosen for experiments over > 2 years for assays under study at the time of recruitment and sample number is indicated in each figure legend.
Flow cytometry labeling
Human peripheral blood mononuclear cells (PBMCs) were isolated from heparinized blood using Ficoll-Hypaque (GE Healthcare, NJ) as previously described [58]. For measurement of Tyro, Axl, and Mer expression, cells were labeled for 30 min at 4°C protected from light with antibodies for surface lineage markers and TAMs as follows: APC-Cy7 CD16 (BD Biosciences, CA 557758), PerCP CD14 (BD 340585), APC CD11c (BD 559877), PE anti-Axl (R & D FAB154P), anti-Mer (FAB8912P), and anti-Tyro (FAB859P) [39]. The immunostained cells were washed with BD wash buffer and fixed in 1% paraformaldehyde. Data was acquired using an LSR II instrument (BD Biosciences, CA) and analyzed using FlowJo software (Tree Star, OR) as described previously [16]. Gating strategy is shown in Supplementary Figure 1.
Protein S signaling assays
PBMCs were plated at 2.5 × 106/ml in a 12 well plate in RPMI/20% human serum/1% pen/strep, washed at 2 hr, and incubated overnight in RPMI in the absence of serum as described previously [59]. Adherent monocytes were pre-treated with medium alone or with 0.5 mM phosphatidyl serine liposomes (Avanti Polar Lipids, Inc., Alabaster, AL) complexed for 30 min under N2 gas with 100 nM Protein S (Haematologic Technologies, Inc., Essex Junction, VT) as described [36]. Cells were harvested at 10, 30 min for immunoblot or at 4 hr for Q-PCR.
Quantitative PCR (qPCR) analysis
Total RNA was harvested by RNeasy mini kit (Qiagen, CA) and cDNA was synthesized using AffinityScript Multi Temperature cDNA Synthesis Kit (Stratagene, TX). Primers and probes for qPCRs were from Applied Biosystems: Tyro3, Axl, Mer, SOCS1, IL-6, IL-8, TNF, IL-1β. Amplification in duplicate was on batched samples in an iCycler (Bio-Rad, CA); values were normalized to β-actin [58].
Immunoblot analysis
Cells were harvested using CelLytic M Cell Lysis buffer (Sigma, MO) containing protease inhibitor cocktail as described previously [59]. Immunoblots were probed with antibodies to total- and phospho-Mer, STAT-1, AKT, p38 MAPK, and β-actin, developed using Amersham ECL Reagents (GE Healthcare), and scanned using Image J software.
Statistical analysis
Demographic characteristics of participants were compared between the age groups with Fisher exact tests for categorical factors and with a t-test for continuous factors. Age was sampled to differ, thus ranges are provided. We used multivariable General Linear Models to estimate the effect of age group on the cell expression of TAMs, and signaling studies were adjusted for within person correlation of test conducted at baseline, 10 and 30 minutes. Statistical tests used SAS version 9.2 (SAS Institute, Cary, NC) and were 2-tailed with p < 0.05 considered significant. Difference for qPCR data was determined using Mann-Whitney test with Bonferroni correction or unpaired t-test using Graphpad Prism (GraphPad Software, Inc.).
Abbreviations
RPMI, PBMC, TAM.
Author contributions
XW, AM, and FQ performed the experiments; RRM conceived the idea and drafted the manuscript; HGA and CR conducted statistical analysis; all the authors reviewed the manuscript.
ACKNOWLEDGMENTS
The authors are grateful to Barbara Siconolfi and Lida Yuan for valuable assistance, and the Yale IMAGIN team for insightful discussions.
CONFLICTS OF INTEREST
The authors declare no competing financial interests.
FUNDING
This work was supported in part by the National Institutes of Health (HHS N272201100019C) and by the Yale Claude D. Pepper Older Americans Independence Center (P30 AG021342).
REFERENCES
1. Shaw AC, Goldstein DR, Montgomery RR. Age-dependent dysregulation of innate immunity. Nat Rev Immunol. 2013; 13:875–887.
2. Carr EJ, Dooley J, Garcia-Perez JE, Lagou V, Lee JC, Wouters C, Meyts I, Goris A, Boeckxstaens G, Linterman MA, Liston A. The cellular composition of the human immune system is shaped by age and cohabitation. Nat Immunol. 2016; 17:461–468.
3. Montgomery RR, Shaw AC. Paradoxical changes in innate immunity in aging: Recent progress and new directions. Journal of Leukocyte Biology. 2015; 98:937–943.
4. Pillai PS, Molony RD, Martinod K, Dong H, Pang IK, Tal MC, Solis AG, Bielecki P, Mohanty S, Trentalange M, Homer RJ, Flavell RA, Wagner DD, et al. Mx1 Reveals Innate Pathways to Antiviral Resistance and Lethal Influenza Disease. Science. 2016; 352:463–466.
5. Molony RD, Malawista A, Montgomery RR. Reduced dynamic range of antiviral innate immune responses in aging. Exp Gerontol. 2017 Aug 16. [Epub ahead of print].
6. Franceschi C, Bonafe M, Valensin S, Olivieri F, De Luca M, Ottaviani E, De Benedictis G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann N Y Acad Sci. 2000; 908:244–254.
7. Bettigole SE, Glimcher LH. Endoplasmic reticulum stress in immunity. Annu Rev Immunol. 2015; 33:107–138.
8. Kapetanovic R, Bokil NJ, Sweet MJ. Innate immune perturbations, accumulating DAMPs and inflammasome dysregulation: A ticking time bomb in ageing. Ageing Res Rev. 2015; 24:40–53.
9. Medzhitov R. Toll-like receptors and innate immunity. Nature Rev Immunol. 2001; 1:135–145.
10. Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol. 2003; 21:335–376.
11. Kawai T, Akira S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int Immunol. 2009; 21:317–337.
12. Brubaker SW, Bonham KS, Zanoni I, Kagan JC. Innate immune pattern recognition: a cell biological perspective. Annual review of immunology. 2015; 33:257–290.
13. van Duin D, Mohanty S, Thomas V, Ginter S, Montgomery RR, Fikrig E, Allore HG, Medzhitov R, Shaw AC. Age-associated defect in human TLR-1/2 function. J Immunol. 2007; 178:970–975.
14. Nyugen J, Agrawal S, Gollapudi S, Gupta S. Impaired functions of peripheral blood monocyte subpopulations in aged humans. Journal of clinical immunology. 2010; 30:806–813.
15. Panda A, Qian F, Mohanty S, van Duin D, Newman FK, Zhang L, Chen S, Towle V, Belshe RB, Fikrig E, Allore HG, Montgomery RR, Shaw AC. Age-associated decrease in TLR function in primary human dendritic cells predicts influenza vaccine response. J Immunol. 2010; 184:2518–2527.
16. Qian F, Guo X, Wang X, Yuan X, Chen S, Malawista SE, Bockenstedt LK, Allore HG, Montgomery RR. Reduced bioenergetics and toll-like receptor 1 function in human polymorphonuclear leukocytes in aging. Aging (Albany NY). 2014; 6:131–139. https://doi.org/10.18632/aging.100642.
17. Finak G, Langweiler M, Jaimes M, Malek M, Taghiyar J, Korin Y, Raddassi K, Devine L, Obermoser G, Pekalski ML, Pontikos N, Diaz A, Heck S, et al. Standardizing Flow Cytometry Immunophenotyping Analysis from the Human ImmunoPhenotyping Consortium. Scientific reports. 2016; 6:20686.
18. Stout-Delgado HW, Vaughan SE, Shirali AC, Jaramillo RJ, Harrod KS. Impaired NLRP3 inflammasome function in elderly mice during influenza infection is rescued by treatment with nigericin. J Immunol. 2012; 188:2815–2824.
19. Youm YH, Grant RW, McCabe LR, Albarado DC, Nguyen KY, Ravussin A, Pistell P, Newman S, Carter R, Laque A, Munzberg H, Rosen CJ, Ingram DK, et al. Canonical Nlrp3 inflammasome links systemic low-grade inflammation to functional decline in aging. Cell metabolism. 2013; 18:519–532.
20. Bauernfeind F, Niepmann S, Knolle PA, Hornung V. Aging-Associated TNF Production Primes Inflammasome Activation and NLRP3-Related Metabolic Disturbances. The Journal of Immunology. 2016; 197:2900–2908.
21. Qian F, Wang X, Zhang L, Lin A, Zhao H, Fikrig E, Montgomery RR. Impaired interferon signaling in dendritic cells from older donors infected in vitro with West Nile virus. J Infect Dis. 2011; 203:1415–1424.
22. Metcalf TU, Cubas RA, Ghneim K, Cartwright MJ, Grevenynghe JV, Richner JM, Olagnier DP, Wilkinson PA, Cameron MJ, Park BS, Hiscott JB, Diamond MS, Wertheimer AM, et al. Global analyses revealed age-related alterations in innate immune responses after stimulation of pathogen recognition receptors. Aging Cell. 2015; 14:421–432.
23. Mohanty S, Joshi SR, Ueda I, Wilson J, Blevins TP, Siconolfi B, Meng H, Devine L, Raddassi K, Tsang S, Belshe RB, Hafler DA, Kaech SM, et al. Prolonged pro-inflammatory cytokine production in monocytes modulated by interleukin-10 after influenza vaccination in older adults. J Infect Dis. 2015; 211:1174–1184.
24. Thakar J, Mohanty S, West AP, Joshi SR, Ueda I, Wilson J, Meng H, Blevins TP, Tsang S, Trentalange M, Siconolfi B, Park K, Gill TM, et al. Aging-dependent alterations in gene expression and a mitochondrial signature of responsiveness to human influenza vaccination. Aging (Albany NY). 2015; 7:38–52. https://doi.org/10.18632/aging.100720.
25. Lu Q, Lemke G. Homeostatic regulation of the immune system by receptor tyrosine kinases of the Tyro 3 family. Science. 2001; 293:306–311.
26. Lemke G, Rothlin CV. Immunobiology of the TAM receptors. Nat Rev Immunol. 2008; 8:327–336.
27. Rothlin CV, Carrera-Silva EA, Bosurgi L, Ghosh S. TAM receptor signaling in immune homeostasis. Annu Rev Immunol. 2015; 33:355–391.
28. Choi JY, Seo JY, Yoon YS, Lee YJ, Kim HS, Kang JL. Mer signaling increases the abundance of the transcription factor LXR to promote the resolution of acute sterile inflammation. Science signaling. 2015; 8:ra21.
29. Chan PY, Carrera Silva EA, De Kouchkovsky D, Joannas LD, Hao L, Hu D, Huntsman S, Eng C, Licona-Limon P, Weinstein JS, Herbert DR, Craft JE, Flavell RA, et al. The TAM family receptor tyrosine kinase TYRO3 is a negative regulator of type 2 immunity. Science. 2016; 352:99–103.
30. Qin B, Wang J, Ma N, Yang M, Fu H, Liang Y, Huang F, Yang Z, Zhong R. The association of Tyro3/Axl/Mer signaling with inflammatory response, disease activity in patients with primary Sjogren’s syndrome. Joint Bone Spine. 2015; 82:258–263.
31. Zheng S, Hedl M, Abraham C. TAM receptor-dependent regulation of SOCS3 and MAPKs contributes to proinflammatory cytokine downregulation following chronic NOD2 stimulation of human macrophages. J Immunol. 2015; 194:1928–1937.
32. Sen P, Wallet MA, Yi Z, Huang Y, Henderson M, Mathews CE, Earp HS, Matsushima G, Baldwin AS Jr, Tisch RM. Apoptotic cells induce Mer tyrosine kinase-dependent blockade of NF-kappaB activation in dendritic cells. Blood. 2007; 109:653–660.
33. Cuervo AM, Wong E. Chaperone-mediated autophagy: roles in disease and aging. Cell Res. 2014; 24:92–104.
34. Montgomery RR. Age-related alterations in immune responses to West Nile virus infection. Clin Exp Immunol. 2017; 187:26–34.
35. Lew ED, Oh J, Burrola PG, Lax I, Zagorska A, Traves PG, Schlessinger J, Lemke G. Differential TAM receptor-ligand-phospholipid interactions delimit differential TAM bioactivities. Elife. 2014; 3:03385.
36. Tsou WI, Nguyen KQ, Calarese DA, Garforth SJ, Antes AL, Smirnov SV, Almo SC, Birge RB, Kotenko SV. Receptor Tyrosine Kinases, TYRO3, AXL, and MER, Demonstrate Distinct Patterns and Complex Regulation of Ligand-induced Activation. J Biol Chem. 2014; 289:25750–25763.
37. Scott RS, McMahon EJ, Pop SM, Reap EA, Caricchio R, Cohen PL, Earp HS, Matsushima GK. Phagocytosis and clearance of apoptotic cells is mediated by MER. Nature. 2001; 411:207–211.
38. Seitz HM, Camenisch TD, Lemke G, Earp HS, Matsushima GK. Macrophages and dendritic cells use different Axl/Mertk/Tyro3 receptors in clearance of apoptotic cells. J Immunol. 2007; 178:5635–5642.
39. Malawista A, Wang X, Trentalange M, Allore HG, Montgomery RR. Coordinated expression of tyro3, axl, and mer receptors in macrophage ontogeny. Macrophage. 2016; 3:e1261.
40. Takasugi Y, Kurai F, Kazume I, Otsuka M, Nehishi Y, Tuda R, Aramaki Y. Down regualtion of MyD88 in Macrophages Treated with Liposomes composed of Phospahtidylserine. Pharmacol Pharm. 2013; 4:248–254.
41. Park HJ, Baen JY, Lee YJ, Choi YH, Kang JL. The TAM-family receptor Mer mediates production of HGF through the RhoA-dependent pathway in response to apoptotic cells. Mol Biol Cell. 2012; 23:3254–3265.
42. Eken C, Martin PJ, Sadallah S, Treves S, Schaller M, Schifferli JA. Ectosomes released by polymorphonuclear neutrophils induce a MerTK-dependent anti-inflammatory pathway in macrophages. J Biol Chem. 2010; 285:39914–39921.
43. Agrawal A, Agrawal S, Cao JN, Su H, Osann K, Gupta S. Altered innate immune functioning of dendritic cells in elderly humans: a role of phosphoinositide 3-kinase-signaling pathway. J Immunol. 2007; 178:6912–6922.
44. Bektas A, Zhang Y, Lehmann E, Wood WH 3rd, Becker KG, Madara K, Ferrucci L, Sen R. Age-associated changes in basal NF-kappaB function in human CD4+ T lymphocytes via dysregulation of PI3 kinase. Aging (Albany NY). 2014; 6:957–974. https://doi.org/10.18632/aging.100705.
45. Sapey E, Greenwood H, Walton G, Mann E, Love A, Aaronson N, Insall RH, Stockley RA, Lord JM. Phosphoinositide 3-kinase inhibition restores neutrophil accuracy in the elderly: toward targeted treatments for immunosenescence. Blood. 2014; 123:239–248.
46. Morizono K, Xie Y, Olafsen T, Lee B, Dasgupta A, Wu AM, Chen IS. The soluble serum protein Gas6 bridges virion envelope phosphatidylserine to the TAM receptor tyrosine kinase Axl to mediate viral entry. Cell Host Microbe. 2011; 9:286–298.
47. Bhattacharyya S, Zagorska A, Lew ED, Shrestha B, Rothlin CV, Naughton J, Diamond MS, Lemke G, Young JA. Enveloped viruses disable innate immune responses in dendritic cells by direct activation of TAM receptors. Cell Host Microbe. 2013; 14:136–147.
48. Miner JJ, Daniels BP, Shrestha B, Proenca-Modena JL, Lew ED, Lazear HM, Gorman MJ, Lemke G, Klein RS, Diamond MS. The TAM receptor Mertk protects against neuroinvasive viral infection by maintaining blood-brain barrier integrity. Nat Med. 2015; 21:1464–1472.
49. Hastings AK, Yockey LJ, Jagger BW, Hwang J, Uraki R, Gaitsch HF, Parnell LA, Cao B, Mysorekar IU, Rothlin CV, Fikrig E, Diamond MS, Iwasaki A. TAM Receptors Are Not Required for Zika Virus Infection in Mice. Cell Rep. 2017; 19:558–568.
50. Linger RM, Keating AK, Earp HS, Graham DK. TAM receptor tyrosine kinases: biologic functions, signaling, and potential therapeutic targeting in human cancer. Adv Cancer Res. 2008; 100:35–83.
51. Cummings CT, Linger RM, Cohen RA, Sather S, Kirkpatrick GD, Davies KD, DeRyckere D, Earp HS, Graham DK. Mer590, a novel monoclonal antibody targeting MER receptor tyrosine kinase, decreases colony formation and increases chemosensitivity in non-small cell lung cancer. Oncotarget. 2014; 5:10434–10445. https://doi.org/10.18632/oncotarget.2142.
52. Lee-Sherick AB, Zhang W, Menachof KK, Hill AA, Rinella S, Kirkpatrick G, Page LS, Stashko MA, Jordan CT, Wei Q, Liu J, Zhang D, DeRyckere D, et al. Efficacy of a Mer and Flt3 tyrosine kinase small molecule inhibitor, UNC1666, in acute myeloid leukemia. Oncotarget. 2015; 6:6722–6736. https://doi.org/10.18632/oncotarget.3156.
53. Ruan Q, Qian F, Yu Z. Effects of polymorphisms in immunity-related genes on the immune system and successful aging. Curr Opin Immunol. 2014; 29:49–55.
54. Sebastiani P, Gurinovich A, Bae H, Andersen S, Malovini A, Atzmon G, Villa F, Kraja AT, Ben-Avraham D, Barzilai N, Puca A, Perls TT. Four Genome-Wide Association Studies Identify New Extreme Longevity Variants. J Gerontol A Biol Sci Med Sci. 2017; 72:1453–1464.
55. Mostafavi H, Berisa T, Day FR, Perry JRB, Przeworski M, Pickrell JK. Identifying genetic variants that affect viability in large cohorts. PLoS Biol. 2017; 15:e2002458.
56. Gopalan S, Carja O, Fagny M, Patin E, Myrick JW, McEwen LM, Mah SM, Kobor MS, Froment A, Feldman MW, Quintana-Murci L, Henn BM. Trends in DNA Methylation with Age Replicate Across Diverse Human Populations. Genetics. 2017; 206:1659–1674.
57. McDaid AF, Joshi PK, Porcu E, Komljenovic A, Li H, Sorrentino V, Litovchenko M, Bevers RPJ, Rueger S, Reymond A, Bochud M, Deplancke B, Williams RW, et al. Bayesian association scan reveals loci associated with human lifespan and linked biomarkers. Nature communications. 2017; 8:15842.
58. Kong KF, Delroux K, Wang X, Qian F, Arjona A, Malawista SE, Fikrig E, Montgomery RR. Dysregulation of TLR3 impairs the innate immune response to West Nile virus in the elderly. J Virol. 2008; 82:7613–7623.
59. Qian F, Wang X, Zhang L, Chen S, Piecychna M, Allore H, Bockenstedt LK, Malawista SE, Bucala R, Shaw A, Fikrig E, Montgomery RR. Age-associated elevation in TLR5 leads to increased inflammatory responses in the elderly. Aging Cell. 2012; 11:104–110.