
INTRODUCTION
Trichothecenes, a large group of chemically related mycotoxins mainly produced by fungi of genus Fusarium, are among the most commonly occurring contaminants in the food chain. Trichothecenes are divided into four types (A–D) according to their characteristic functional groups. Types A and B are of greater concern to people due to their high toxicity and frequent presence as food contaminants [1, 2]. For both of these reasons, T-2 toxin and deoxynivalenol (DON), which belong to type A and type B, respectively, are considered to be the most important compounds of this class (Figure 1) [3–5]. The most toxic trichothecene, T-2 toxin, causes growth inhibition and sublethal or even lethal toxicosis in humans and farm animals. Alimentary toxic aleukia (ATA), a fatal human disease, is primarily associated with T-2 toxin [6, 7]. In Europe, especially in the Nordic countries, contamination of cereals with T-2 toxin is a serious problem [8–11]. In aerosol form, T-2 toxin can easily penetrate organisms through the lungs. Reactions of the skin or intestinal mucosa are provoked rapidly after direct contact through dermal application or ingestion of the compound. In addition, T-2 toxin is a radiomimetic compound, and thus exacerbates the effect of ionizing radiation [12–14].
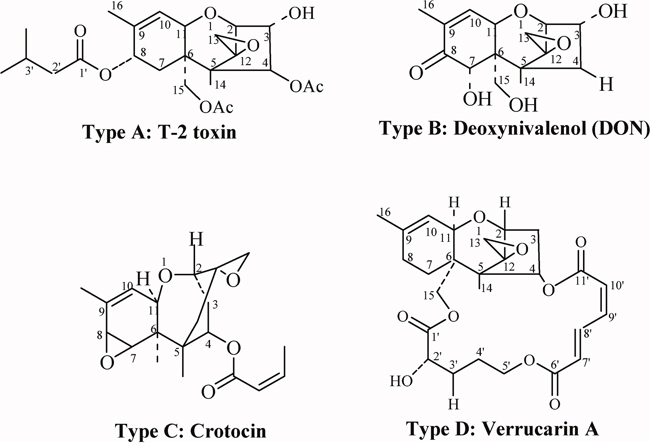
Figure 1: Chemical structure of trichothecenes (Types A-D).
DON is less toxic than T-2 toxin and many other trichothecenes, but it is the most widely distributed trichothecene and is commonly found in barley, corn, wheat, and mixed feed [4, 5, 15]. At the molecular level, DON binds to the ribosome and induces ribotoxic stress, leading to the activation of mitogen-activated protein kinases (MAPK), cell-cycle arrest, and apoptosis [16]. The toxic effects of DON include emesis and anorexia, alteration of intestinal and immune functions, reduced absorption of nutrients, and elevated susceptibility to infection and chronic diseases [17]. Importantly, DON contamination in wheat flour may be involved in the fluctuating but high prevalence of Kashin-Beck disease (KBD) [18]. DON contamination has also been reported in countries including the Czech Republic, Italy, Spain, Croatia, Canada, and Germany [19–24]. The routes of exposure and various toxicities of trichothecenes are illustrated in Figure 2.
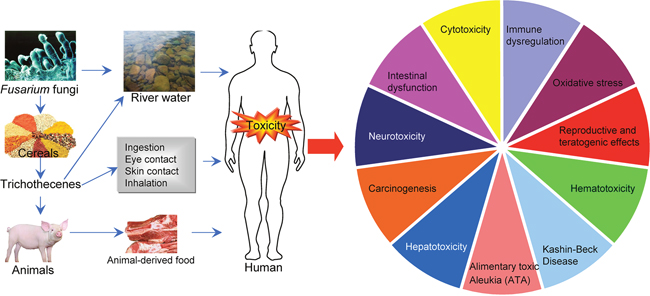
Figure 2: Routes of exposure and various toxicities of trichothecenes.
The evidence accumulated to date shows that oxidative stress is an important toxic mechanism of trichothecenes [25–29], which significantly increase levels of reactive oxygen species (ROS) and deplete intracellular reduced glutathione (GSH). Moreover, these compounds increase lipid peroxidation, leading to single-strand breaks in DNA [30–31]. In addition, they activate signaling pathways including MAPK, JAK/STAT, and NF-κB, which trigger apoptosis [4, 5, 32, 33]. Trichothecenes activate MAPK by a mechanism called the “ribotoxic stress response” (RSR), which drives both cytokine gene expression and apoptosis in macrophages [34]. Over the last decade, mitochondria-related toxic mechanisms became an active area of research in this field. In addition to ribosomes, mitochondria are considered to be targets of trichothecenes [35, 36]. Importantly, mitochondrial dysfunction caused by trichothecenes is largely associated with oxidative stress. Normally, trichothecenes damage the normal function of mitochondria and release free radicals (including ROS), induce lipid peroxidation, and change the antioxidant status of cells, thereby reducing the activity of antioxidant enzymes [4, 5]. Thus, oxidative stress mediated by trichothecenes and their related toxicities pose a great risk to human health.
As trichothecenes cause oxidative stress, researchers are exploring potential antioxidant agents for use against trichothecenes. To date, many natural compounds with antioxidant activity against these compounds have been identified. For example, the traditional antioxidant agents vitamins A, C, and E exhibit beneficial effects against DON-induced protein and lipid peroxidation [37]. Quercetin can reduce apoptosis caused by T-2 toxin [38]. Selenium can prevent erythrocyte membrane damage induced by T-2 toxin [39]. N-Acetyl-cysteine (NAC) protects chicken growth plate chondrocytes from T-2 toxin-induced oxidative stress [40]. In addition, antimicrobial peptides can improve feed efficiency, immune function, and anti-oxidative capacity, as well as alleviate organ damage caused by DON [41]. Moreover, some plant extracts, including polyphenol epigallocatechin 3-gallate (EGCG), leontopodic acid (LA), and quince seed mucilage, have marked antioxidant effects against trichothecenes. Therefore, elucidation of the anti-oxidative activities of various compounds is crucial for the detoxification of trichothecenes. Previously, however, no review article had fully summarized the data regarding such antioxidant agents.
Therefore, in this review, we will first briefly discuss the oxidative stress mechanisms of trichothecenes. Our main goal, however, is to explore our current knowledge regarding potential antioxidant agents against trichothecenes, as well as their antioxidant mechanisms. We focus specifically on the most toxic and widespread trichothecenes, T-2 toxin and DON. Finally, we will discuss some decontamination methods, including bacterial and yeast biotransformation and degradation, as well as mycotoxin-binding agents. We believe that this review provides a comprehensive overview of current knowledge on detoxification of trichothecenes and casts new light on attenuation of oxidative stress.
OXIDATIVE STRESS AND TRICHOTHECENES
Oxidative stress is an important mechanism of trichothecene toxicity. Normally, trichothecenes disrupt the normal function of mitochondria and generate free radicals, including ROS. These oxidative compounds induce lipid peroxidation and change the antioxidant status of the cells, thereby reducing the activity of antioxidant enzymes such as glutathione-S-transferase (GST), superoxide dismutase (SOD), and catalase (CAT) in the body [4]. DNA damage is an early event associated with the generation of ROS and lipid peroxidation. Some signaling pathways, including MAPK, JAK/STAT, and NF-κB, are subsequently induced by oxidative stress, and the caspase-mediated apoptosis pathways are also activated [34].
Oxidative stress is the mechanism by which T-2 toxin causes DNA damage and apoptosis [30, 31, 42]. Indeed, T-2 toxin can induce generation of ROS as early as 30 min after exposure, followed by significant depletion of glutathione levels and elevated lipid peroxidation [30]. Moreover, the ROS-mediated mitochondrial pathway plays an important role in T-2 toxin-induced apoptosis [29, 43]. In granulosa cells and murine embryonic stem cells, T-2 toxin treatment induces ROS accumulation, resulting in reduction of mitochondrial transmembrane potential. In addition, T-2 toxin strongly inhibits the mitochondrial electron transport system (ETS) in rat cardiomyocytes [44]. In murine embryonic stem cells (mESCs), T-2 toxin inhibited mitochondrial biogenesis by increasing ROS levels, leading in turn to inhibition of ESC differentiation [45]. In addition, proteomic changes in chicken primary hepatocytes exposed to T-2 toxin are associated with oxidative stress and mitochondrial enhancement [46]. More recently, our group found that nitric oxide (NO)-mediated mitochondrial damage plays a critical role in T-2 toxin-induced apoptosis and growth hormone deficiency in rat anterior pituitary GH3 cells [7]. We found that T-2 toxin caused significant increases in NO generation, cell apoptosis, iNOS activity, production of inflammatory factors, and caspase pathway activation, while also decreasing growth hormone (GH) production and mitochondrial membrane potential (ΔΨm). These data show that mitochondria are a primary target of T-2 toxin-induced NO, and NO is a key mediator of T-2 toxin-induced apoptosis and GH deficiency via a mitochondria-dependent pathway.
T-2 induces oxidative damage in the liver [46]. Specifically, T-2 toxin exposure causes significant oxidative damage by depleting hepatic glutathione, increasing lipid peroxidation, altering the activity of antioxidant enzymes, and inducing protein oxidation in a time-dependent manner. Moreover, the oxidative stress and apoptosis induced by T-2 toxin are involved in the developmental toxicity of this compound in zebrafish embryos [47]. We also know that T-2 toxin can cross the blood brain barrier [48]. Consistent with this, percutaneously or subcutaneously applied T-2 toxin causes oxidative damage in the brain. Mice treated with T-2 toxin via either exposure route exhibit a time-dependent increase in ROS generation, glutathione depletion, lipid peroxidation, and protein carbonyl content in the brain [27].
The oxidative damage caused by T-2 toxin is involved in the mechanism of KBD [49]. The increase in thiobarbituric acid-reactive substances (TBARS) and decrease in antioxidant levels following T-2 toxin treatment may induce oxidative stress in joint tissues and contribute to the pathological process of cartilage damage in KBD. In addition, in aquatic species, T-2 toxin induces anemia and oxidative stress, as well as altering the immune response [50, 51]. Shrimp exposed to T-2 toxin in the diet showed elevated ROS levels, SOD activity, and histopathological changes in the hepatopancreas [51].
Oxidative stress is also an important toxic mechanism of DON, a type B trichothecene. The manifestations of this stress are elevated levels of ROS and malondialdehyde (MDA), and reduced levels of GSH and SOD. In addition, as T-2 toxin, DON can also cause mitochondrial damage by decreasing mitochondrial membrane potential and inducing apoptosis accompanied by upregulation of apoptosis-related factors including caspase-3, caspase-8, and caspase-9 [36, 52]. DON induces apoptosis in splenic lymphocytes and PC12 cells via a ROS-mediated mitochondrial pathway [53, 54]. Yang et al. (2014) found that DON triggered potential genotoxicity in human peripheral blood lymphocytes via oxidative damage [55]. Furthermore, those authors showed that DON exposure increased peroxidation, decreased antioxidant activity, and inhibited DNA repair and expression of heme oxygenase-1 (HO-1).
Thus, it is clear that oxidative stress plays important roles in the toxicity of trichothecenes. Interestingly, some cells can mount their own antioxidant defenses following exposure to these toxins. For example, when HepG2 cells are exposed to DON, ROS levels are significantly elevated, but as time passes, antioxidant enzymes such as SOD and CAT are highly upregulated, presumably reflecting the cells’ effort to detoxify the damage caused by DON [56]. Thus, the antioxidant defense system of HepG2 cells represents an insufficient adaptation aimed at minimizing DON-induced oxidative injury. In addition, JunD, a member of the AP-1 family of transcription factors, plays an important role in defense against oxidative stress induced by T-2 toxin [57]. A similar self-defense capacity has also been observed in in vivo studies. Short-term oral exposure of T-2 toxin initiates lipid oxidation in chicken liver, but the antioxidant defense system eliminates the free radicals and inhibits the oxidative stress [58].
Collectively, these findings show that oxidative stress is an important underlying toxic mechanism of trichothecenes. Trichothecenes generate ROS that induce lipid peroxidation, which in turn leads to changes in membrane integrity, cellular redox signaling, and cellular antioxidant status. DNA damage correlates with, but precedes, the generation of ROS and lipid peroxidation. Because trichothecenes are commonly found in food and feed, the cellular effects of these toxins in relation to oxidative stress, as well as effective measures for combating their toxicity, should be addressed in future studies. The proposed mechanisms underlying oxidative stress induced by trichothecenes are shown in Figure 3.
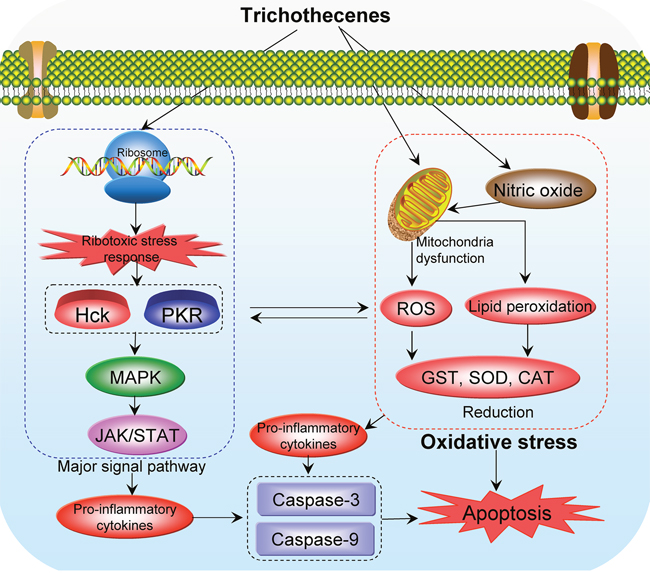
Figure 3: Proposed mechanisms of oxidative stress-mediated toxicity of trichothecenes.
ANTIOXIDANT AGENTS AGAINST TRICHOTHECENES
As discussed above, oxidative stress plays key roles in the toxicity of trichothecenes. Oxidative damage caused by trichothecenes is one of the mechanisms underlying toxin-induced cell injury and DNA damage, which eventually lead to tumorigenesis [4]. Currently, researchers are trying to identify potential agents that effectively prevent trichothecene-induced oxidative stress and the associated immunotoxicity. Interestingly, agents such as vitamins, quercetin, selenium, glucomannan, amino acids, nucleotides, antimicrobial peptides, bacteria, polyunsaturated fatty acids, oligosaccharides, and some plant extracts exert promising anti-oxidative effect against trichothecenes. Accordingly, these compounds have attracted close attention from researchers. The various antioxidant agents with potential for use against trichothecenes are presented in Table 1.
Table 1: Summary of the antioxidant agents against trichothecenes
Entry |
Agents |
Cell or animal model |
Specific effects |
References |
|---|---|---|---|---|
1 |
Vitamins E, A, C |
Murine YAC-1lymphoblastic cell |
Against the DON-induced protein and lipid peroxidation; increased the cell viability and cell proliferation. |
[37] |
2 |
Vitamins C, E |
Rat |
Decrease the lipid peroxides and TBARS values caused by T-2 and DON; decreased mortality. |
[59] |
3 |
Coenzyme Q10, Vitamin E |
Mice |
Decreased DNA damage caused by T-2 toxin; Protection against cell death and glutathione depletion caused by T-2 toxin. |
[61] |
4 |
Lutein |
HT-29 cell |
Decreased DON-induced oxidative stress and apoptosis. |
[26] |
5 |
Quercetin |
Rabbit ovary |
Increased cell proliferation and reduced cell apoptosis caused by T-2 toxin. |
[38] |
6 |
Quercetin |
Porcine ovarian granulosa cell |
Effectived in maintaining and increasing of TAS, activitied SOD and glutathione peroxidase (GPx). |
[62] |
7 |
Selenium |
Mice |
T-2 toxin decreased the number of CD8+, CD3+, CD4+ T-lymphocytes, whereas Se mitigated these decreases. |
[65] |
8 |
Selenium |
Mice |
Prevented erythrocyte membrane damage induced by T-2 toxin. |
[39] |
9 |
Selenium chondroitin sulfate nanoparticles (SeCS) |
KBD patients |
Blocked the T-2 toxin-induced chondrocyte apoptosis by decreasing the expression of ATF2, JNK and p38. |
[69] |
10 |
Modified glucomannan |
Chicken |
Made a 45% reduction of lipid peroxidation in the liver in comparison to the effects of T-2 toxin alone. |
[74] |
11 |
Glucomannan |
Pig |
Restored anti-ovalbumin immunoglobulin G production, which was significantly reduced in pigs expose to T-2 toxin. |
[72] |
12 |
N-acetyl-cysteine (NAC) |
Chicken |
Decreased the levels of ROS and MDA which were increased by T-2 toxin. |
[77] |
13 |
Arginine, Glutamine |
Pig |
Alleviated the impairment induced by DON stress and immune relevant cytokines in growing pigs. |
[78] |
14 |
Glutamic acid |
Pig |
Increased the plasma activities of SOD and GSH-Px and the proliferating cell nuclear antigen (PCNA) labeling indexes for the jejunum and ileum. |
[79] |
15 |
L-carnitine |
Rat |
Reduced toxicity and prevented the hepatocytes from abnormal caspase-3 activity and apoptosis caused by T-2 toxin. |
[81] |
16 |
Nucleotides |
Chicken |
Reduced the extent of DNA damage induced by T-2 toxin in leukocytes. |
[82] |
17 |
Composite antimicrobial peptides (CAP) |
Pig |
DON decreased peripheral lymphocyte proliferation, whereas supplementation with CAP increased it on day 15 and 30. |
[41] |
18 |
Composite antimicrobial peptides (CAP) |
Pig |
Improved intestinal morphology and promoted intestinal epithelial cell proliferation and protein synthesis. |
[84] |
19 |
Epigallocatechin-3-gallate (EGCG) |
HT-29 cell |
Protected against DON mediated-oxidative stress, up regulation of NF-κB, COX-2 and caspase-3 activated apoptosis. |
[85] |
20 |
Leontopodic acid (LA) |
U937 cells |
Protected cells from DON-induced cell damage; enhanced glutathione peroxidase activity. |
[87] |
21 |
Quince seed |
Rabbit |
Showed more and better healing effects on dermal toxicity caused by T-2 toxin. |
[88] |
22 |
Rutin |
Rat |
Reversed the T-2 toxin-mediated increase of lipid peroxidation parameter, SOD and glutathione concentration in liver homogenate. |
[91] |
23 |
lycopene |
Chicken |
Inhibited T-2 toxin induce oxidative stress and protected the cellular level of GSH |
[92] |
24 |
Lactobacillus plantarum JM113 |
Chicken |
Reduced MDA activity in the jejunal mucosa; increased the mRNA levels of Nrf2 and its corresponding downstream HO-1 gene. |
[93] |
25 |
Lactobacillus rhamnosus GG (LGG) |
Mice |
Prevented or treated the unwanted adverse effects of DON/ZEA in mice by regulation of goblet cell mucus secretion, improvement of plasma D-lactate, IL-8/CXCL8 and serum Ig levels. |
[94] |
26 |
PUFA, DHA, EPA |
Mice |
Suppressed DON-induced IgAN. |
[95] |
27 |
Docosahexaenoic acid (DHA) |
Mice |
Suppressed the PKR and CREB kinase pathways thus inhibited the IL-6 transcription |
[97] |
28 |
Galacto-oligosaccharides (GOSs) |
Caco-2 Cell Monolayers; B6C3F1 Mice |
Prevented the DON-induced loss of epithelial barrier function; stabilized the expression and cellular distribution of claudin3 and suppressed; In mice, GOSs prevented the DON-induced mRNA overexpression of claudin3 and CXCL8 homolog keratinocyte hemoattractant. |
[98] |
Vitamins
During the last decade, antioxidant agents such as vitamins and microelements have received a great deal of attention. One earlier study demonstrated that dietary use of vitamins E and C can decrease lipid peroxide and TBARS levels, thereby protecting against acute toxicosis caused by trichothecenes (Entry 1–4, Table 1) [59]. In addition, Strasser et al. (2013) [37] further demonstrated that the antioxidant vitamins E, A, and C had beneficial effects against DON-induced protein and lipid peroxidation. These compounds significantly increased cell viability and cell proliferation following toxin exposure. Lutein, a non-vitamin A carotenoid, has been implicated in maintaining eye health, specifically in regard to age-related macular degeneration [60]. The cytoprotective effect of lutein against DON-induced oxidative stress was tested in HT-29 cells [26]. In that study, pretreatment of cells with 10 μM lutein resulted in 95% cell viability. Moreover, lutein combated DON-induced oxidative stress and downregulated expression of inflammatory genes such as NF-κB and COX-2. Lutein also prevented DON-induced migration of NF-κB into the nucleus. Morphological studies revealed that lutein prevented DON-induced apoptosis. In addition to the vitamins, coenzyme Q10 decreases T-2 toxin-induced DNA damage in mouse liver [61].
Quercetin
Quercetin, a member of the large family of flavonoid compounds that exert manifold biological effects, has anti-inflammatory, antioxidant, free radical-scavenging, and metal-chelating properties. Quercetin increases rabbit ovary cell proliferation and diminishes the apoptosis caused by T-2 toxin [38]. Recent work showed that quercetin has no effect in the elimination of ROS generation induced by T-2 toxin, but is still effective in maintaining and increasing total antioxidant status (TAS) and the activities of SOD and glutathione peroxidase (GPx) in porcine granulosa cells in vitro [62]. Thus, based on the in vitro studies, quercetin has the potential to inhibit T-2 toxin-induced oxidative stress and apoptosis; however, additional in vivo studies are needed to verify this conclusion (Entry 5-6, Table 1).
Selenium
Selenium is an essential micronutrient that exerts multiple and complex effects on human health. This element is important for human well-being largely due to its potent antioxidant, anti-inflammatory, and antiviral properties (Entry 7-9, Table 1) [63]. Selenium participates in protection of cells against excess H2O2, heavy metal detoxification, and regulation of the immune and reproductive systems [64]. In the context of mycotoxins, selenium can prevent erythrocyte membrane damage induced by T-2 toxin [39]. The protective effect of selenium may be due to its membrane-stabilizing properties. Salimian et al. (2014) [65] also found that selenium could exert a marked effect against the immunotoxic effects of T-2 toxin in T lymphocytes. KBD is an endemic osteochondropathy manifested by chondrocyte necrosis and apoptosis, cartilage degeneration, and matrix degradation [66], and T-2 toxin is one of the important etiological factors for this disease [67]. In addition, selenium deficiency has also been proposed as a risk factor for KBD. In this view, compromised selenoprotein function leads to oxidative stress and apoptosis, which ultimately manifests as KBD [68]. Recently, selenium chondroitin sulfate nanoparticles (SeCS) with a size range of 30–200 nm were obtained by the research group of Guo et al., who investigated the inhibitory effects of SeCS on T-2 toxin-induced apoptosis of chondrocyte from KBD patients [69]. The results revealed that SeCS partly blocked T-2 toxin-induced chondrocyte apoptosis by decreasing the activity of ATF2, JNK, and p38. Thus, SeCS could be used for prevention and treatment of KBD, as well as other Se-deficiency diseases.
Glucomannan
Glucomannan is a soluble, fermentable, and highly viscous dietary fiber derived from the root of the elephant yam or konjac plant, which is native to Asia [70]. Because ingestion of this compound promotes human health, over the last two decades it has become more frequently used in western countries [71]. During that time, researchers also studied the antioxidant role of glucomannan against trichothecenes (Entry 10-11, Table 1). In pigs [72], glucomannan dietary supplementation exerted no deleterious effects but protected against T-2 toxin immunotoxicity during a vaccine protocol. In another study [73], a modified glucomannan mycotoxin-adsorbing agent was shown to counteract reduced weight gain in T-2 toxin-exposed pigs. This modified glucomannan also reduced the number of Salmonella typhimurium bacteria in the cecum and cecal contents of T-2 toxin-exposed pigs. In addition, the antioxidant effects of organic selenium (Sel-Plex™) and modified glucomannan (Mycosorb™) against T-2 toxin have been assessed [74]. Inclusion of modified glucomannans into a T-2 toxin-contaminated diet provides partial protection against the detrimental effects of the toxin on antioxidant defenses in chicken liver [61], and the combination of modified glucomannans with organic selenium confers further protection against toxin-induced antioxidant depletion and lipid peroxidation in that tissue.
Amino acids
Amino acids play important roles in digestion and absorption of dietary nutrients, metabolism of glucose and lipids, acid–base balance, anti-oxidative responses, detoxification of xenobiotics, and immunity [75]. Dietary antioxidants and their roles in preventing mycotoxin toxicity have attracted increasing attention in recent years in studies aimed at evaluating the potential benefits of amino acids in the diet (Entry 12-15, Table 1). NAC is an effective source of sulfhydryl groups in cells and a scavenger of free radicals that may interact with ROS such as OH• and H2O2 [76]. Recently, NAC was shown to protect chicken growth plate chondrocytes from T-2 toxin-induced oxidative stress [77]; specifically, NAC significantly decreased ROS and MDA levels induced by toxin exposure. Moreover, a greater increase in CAT and SOD activity was observed. Thus, NAC may confer a therapeutic benefit against metaphyseal chondrodysplasia by improving the antioxidant capacity of growth plate chondrocytes. Similarly, supplementation with glutamic acid in piglets exerts a strong anti-oxidative effect against DON. A series of studies [78, 79] showed that DON significantly induces oxidative stress in piglets, but this stress is remarkably reduced by glutamic acid supplementation, as reflected by changes in oxidative parameters in blood and tissues. Meanwhile, DON causes obvious intestinal injury, as determined by microscopic observations and the use of permeability indicators, but this damage is alleviated by glutamic acid supplementation. Moreover, the inhibitory effect of DON on the Akt/mTOR/4EBP1 signaling pathway is reduced by glutamic acid supplementation [79]. Addition of glutamic acid to DON-treated cells increases plasma activities of SOD and glutathione peroxidase (GSH-Px) and the proliferating cell nuclear antigen (PCNA) labeling indexes in jejunum and ileum. These findings indicate that glutamic acid has the potential to repair injuries associated with oxidative stress, intestinal injury, and signaling inhibition.
Arginine and glutamine also play important roles in pig nutrition. The effects of dietary supplementation with arginine and glutamine on both the impairments induced by DON stress and immune cytokines in growing pigs have been studied [78]. The results of that work revealed that levels of IGF1, GH, and SOD in the amino acid-treated groups were significantly higher than those in the toxin control. Moreover, the IL-2 and TNF-α values in the amino acid-treated groups were similar to those in the non-toxin control, and significantly lower than those in the toxin control. Thus, dietary supplementation with arginine and glutamine can alleviate the impairments induced by DON stress and immune cytokines in growing pigs.
L-carnitine not only diminishes oxidative stress but also protects mitochondria against fatty acid stress and apoptosis by inhibiting mitochondrial swelling and Cyt-c release [80]. Pretreatment of rats with L-carnitine prevents hepatocytes from expressing abnormal caspase-3 activity and undergoing apoptosis in response to T-2 toxin [81]. Thus, due to its mitochondrial protective effects, L-carnitine supplementation represents a promising method for diminishing or preventing the toxicity induced by T-2 toxin.
Nucleotides
Nucleotides are “semi-” or “conditionally” essential nutrients, but they may become essential in pathological conditions that demand intense nucleic acid and protein synthesis. Evidence has shown that dietary nucleotides have the potential to reduce the extent of DNA damage induced by T-2 toxin in immune cells [82]. In a study by Frankic et al. (2006) [82], T-2 toxin induced DNA fragmentation in chicken spleen leukocytes, but supplementation with nucleotides reduced the amount of damage (Entry 16, Table 1). This finding highlights the possible beneficial effect of dietary nucleotides on the immune system in cases of mycotoxin intoxication.
Antimicrobial peptides
Antimicrobial peptides are small cationic molecules that are part of the nonspecific defense system; these compounds kill bacteria, modulate bacterial infections, and coordinate host responses to infection [83]. Antimicrobial peptides have the capacity to improve feed efficiency, immune function, and anti-oxidant capacity, and alleviate organ damage caused by DON, and thus have a protective effect in piglets challenged with this toxin [41]. In addition, composite antimicrobial peptides (CAPs) can repair intestinal injury induced by DON; specifically, CAPs improve intestinal morphology and promote intestinal epithelial cell proliferation and protein synthesis (Entry 17-18, Table 1) [84].
Plant extracts
Many natural antioxidants, which provide multiple health benefits, are present in tea, a common beverage (Entry 19–23, Table 1). In a recent study of the cytoprotective effect of green tea on DON-induced toxicity in HT-29 cells [85], EGCG prevented DON-induced cytotoxicity to HT-29 cells in a dose-dependent manner. Even the lowest concentration (5 μM) of EGCG protected against the highest concentration of DON tested, and pretreatment with 20 μM EGCG yielded 99% cell viability. EGCG also protected against oxidative stress, upregulation of NF-κB, COX-2, and caspase-3 activated apoptosis. These results suggest that EGCG acts as cytoprotective agent against DON-induced toxicity. Therefore, the use of EGCG represents an attractive strategy for cytoprotection of cells against the actions of trichothecenes.
LA is a fully substituted hexaric acid derivative obtained from Leontopodium alpinum Cass., commonly known as edelweiss [86]. This molecule exerts a strong antioxidant capacity, and in particular was shown to protect U937 cells from DON-induced oxidative damage and increase GPx activity in these cells [87]. The increase in the activity of detoxifying enzymes is probably the main mechanism underlying antioxidant-mediated chemoprevention.
Quince seed mucilage exerts healing effects on dermal toxicity caused by T-2 toxin [88]. Materials obtained from Cydonia oblonga species (including quince seed mucilage) are an excellent natural source of phenolic acids and flavonoids, which are considered potent antioxidants [89, 90]. Quince seed mucilage may diminish the dermal toxicity of T-2 toxin via its antioxidant properties. Similarly, rutin exhibits strong anti-oxidative capacity against T-2 toxin in rat liver, and decreases T-2 toxin-induced lipid peroxidation [91]. In addition, lycopene from fresh tomatoes prevents T-2 toxin-induced oxidative stress and helps to maintain the cellular level of GSH [92].
Bacteria
Bacteria exhibit promising antioxidant activities against trichothecenes (Entry 24-25, Table 1). A very recent study investigated the antioxidant capacity of Lactobacillus plantarum JM113 isolated from healthy intestinal contents of Tibetan chicken and its protective effect on broiler chickens challenged with DON [93]. The results revealed that dietary supplementation with DON decreased SOD activity in serum and increased MDA in the jejunal mucosa. However, supplementation with L. plantarum JM113 to a DON-contaminated diet caused a significant reduction in MDA activity in the jejunal mucosa. Moreover, DON decreased the expression of nuclear factor erythroid 2-related factor 2 (Nrf2), whereas Nrf2 mRNA levels and expression of the downstream gene HO-1 increased following L. plantarum JM113 treatment. Thus, L. plantarum JM113 has high antioxidant activity, and supplementation of this compound in feed protected the integrity of the intestinal barrier in broilers challenged with DON, suggesting that it could be used to alleviate the negative effects of DON in poultry. Another study assessed the ability of L. rhamnosus GG (LGG) to improve intestinal barrier functions and ameliorate inflammation in Balb/c mice fed with DON-containing diets [94]. The results showed that LGG partially prevented or reversed the adverse effects of DON in mice by regulating goblet cell mucus secretion and normalizing plasma D-lactate, IL-8/CXCL8, and serum Ig levels. Thus, LGG offers a potential dietary intervention strategy against intestinal exposure to mycotoxins.
Polyunsaturated fatty acid
Consumption of DON induces IL-6–dependent IgA nephropathy (IgAN) in mice [95]. Consumption of the n-3 polyunsaturated fatty acid (PUFA), docosahexaenoic acid (DHA), or eicosapentaenoic acid (EPA) suppresses DON-induced IgAN in mice, in agreement with the proposed anti-inflammatory action of these fatty acids [95]. Moreover, these results are consistent with randomized clinical trials demonstrating that fish oil consumption retards loss of renal function in IgAN patients [96]. In a subsequent study [97], DHA was shown to suppress the RNA-activated protein kinase R (PKR) and cAMP response element-binding protein (CREB) kinase pathways, thereby inhibiting the IL-6 transcription. Therefore, suppression of IL-6 expression by DHA might have general importance in human health, specifically in regard to the prevention and treatment of inflammatory and autoimmune diseases mediated by this pro-inflammatory cytokine (Entry 26–27, Table 1).
Oligosaccharide
Galacto-oligosaccharides (GOS) are beneficial food additives capable of protecting vulnerable segments of the human population against the adverse effects of DON and its derivatives (Entry 28, Table 1) [98, 99]. In human Caco-2 cell monolayers and mice, GOS protect the intestinal barrier by maintaining the tight junction network and modulating inflammatory responses after a challenge with DON [98]. Furthermore, GOS stabilize the expression and cellular distribution of claudin 3 and suppress DON-induced synthesis and release of IL-8 [IL8/chemokine CXC motif ligand (CXCL8)] [99]. In mice, GOS prevent DON-induced mRNA overexpression of claudin 3 and CXCL8 homolog keratinocyte hemoattractant (Kc) (Cxcl1), as well as DON-induced morphologic defects [98]. Considering that the DON-induced alterations in the intestinal tract of mice resemble those in humans with chronic inflammatory diseases or regular exposure to DON, further studies should assess in greater detail the potential beneficial effects of GOS to support therapy aimed at prevention of toxin-induced inflammatory bowel diseases and related syndromes.
In summary, many natural compounds exhibit anti-oxidative capacity against trichothecenes. As we described above, NAC and green tea EGCG inhibit ROS generation; coenzyme Q10 and nucleotides reduce DNA damage; and vitamins A, E, and C, glucomannnan, and rutin can reduce lipid peroxidation. On the other hand, other compounds target the antioxidant enzymes: quercetin, LA, and L. plantarum JM113 increase the activity of antioxidant enzymes such as SOD and CAT, thereby reducing oxidative stress and apoptosis. GOS prevent DON-induced overexpression of mRNAs encoding claudin 3 and CXCL8. L-carnitine not only diminishes oxidative stress, but also protects mitochondria against fatty acid stress and apoptosis by inhibiting mitochondrial swelling and Cyt-c release. In addition, L-carnitine and green tea EGCG inhibit caspase activity, thereby directly inhibiting cell apoptosis. Lutein combats DON-induced oxidative stress and downregulates expression of the inflammatory factors NF-κB and COX-2. SeCS can partly block trichothecene-induced chondrocyte apoptosis by decreasing MAPK signaling activity. A schematic illustration of the preventive effect of various antioxidants on trichothecene-induced oxidative stress is shown in Figure 4.
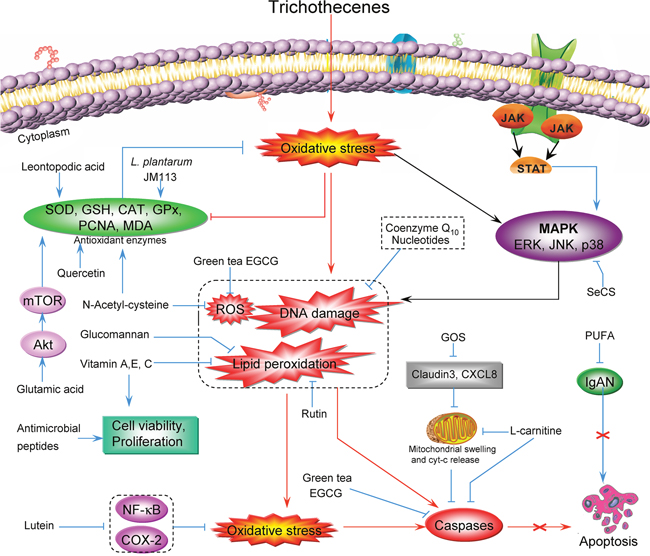
Figure 4: Schematic illustration of the preventive effects of various antioxidants on trichothecene-induced oxidative stress.
OTHER DECONTAMINATION STRATEGIES
In addition to the antioxidant and detoxification agents described above, some decontamination methods are available. These approaches include bacterial and yeast biotransformation and degradation, as well as mycotoxin-binding agents. Because biological reactions offer a specific, irreversible, efficient, and environmentally friendly means of detoxification that leaves neither toxic residues nor any undesired by-products, microbial biotransformation of mycotoxins into non-toxic or low-toxin metabolites has been acknowledged as a promising approach [100]. The isolation and characterization of microorganisms that can biotransform mycotoxins is crucial for practical applications of biotechnology [101]. Based on the ability of lactic acid bacteria (LAB) to inhibit fungal development and remove multiple mycotoxins, including DON and aflatoxins, along with their Generally Recognized As Safe (GRAS) status and probiotic potential, these organisms are candidates for the biological control of fungi such as F. graminearum in crops or even during food production and processing [102, 103]. Increasing evidence shows that LABs could effectively remove trichothecenes T-2 and DON from cereals. For example, Franco et al. (2011) [103] evaluated the ability of different LAB strains to remove DON in vitro. All of the isolated strains and commercial cultures tested in that study inhibited fungal growth and decreased DON levels by 16.41–55.30%. Although the mechanism remains unclear, the detoxification capacity of LAB could be related to metabolic degradation or adsorption of trichothecenes by the bacterial cell wall [104]. In another study, five strains of LAB were tested for their ability to remove DON and T-2 toxin [105]. The capacity of Lactobacillus plantarum strain 102 (LP102) was the strongest after incubation at 37 °C for 72 h. Recently, Zhao et al. (2016) [106] identified a microbial community, Devosia sp. ANSB714, which could degrade 97.34% DON after 24 h. In mice, a diet containing DON reduces growth performance and affects liver, kidney, and immune function. Addition of Devosia sp. ANSB714 to DON-contaminated diets eliminates these effects of DON. Therefore, detoxification with Devosia sp. ANSB714 could have an enormous impact in the livestock industry.
In addition to LAB, the yeast Geotrichum candidum also exhibited a strong ability to reduce T-2 toxin levels in cereals. For example, an interaction between G. candidum and F. langsethiae led to a drastic (93%) reduction in T-2 toxin production [107]. The mechanism remains unclear, but we suspect that the yeast might transform T-2 toxin to another compound or inhibit its production by the fungi. Indeed, McCormick et al. (2012) [108] showed that several yeast species can convert T-2 toxin to 3-acetyl-T-2 toxin, T-2-3-glucoside, and neosolaniol (NEO), which are much less toxic than the parent compound [1]. In addition, a mixture of inorganic (activated carbon) and organic (yeast cell wall) adsorbents contributed to in vitro removal of DON. The highest percentage of adsorption occurred when 2.0% activated carbon and yeast cell wall were present at a 30:70 ratio (>95.6%) and incubation continued for 30, 60, or 90 min [109]. Another study confirmed that yeast-based feed additives could reduce DON-induced oxidative stress in pigs [110].
Microbial detoxification represents an alternative approach to physical and chemical methods for detoxification of DON-contaminated grain. A recent study showed that a novel bacterium, Devosia mutans 17-2-E-8, was capable of transforming DON to a non-toxic stereoisomer, 3-epi-DON, under aerobic conditions, mild temperature, and neutral pH [40]. Moreover, the process was enzymatic in nature and had a high detoxification capacity. These observations suggest the possibility of using the isolated bacterium as a feed treatment to address DON contamination under empirical field conditions. Another trial was conducted to evaluate a feed additive containing epoxidase activity from a bacterium (Mycofix-S) as potential protection against the adverse effects of T-2 toxin in growing male broiler chickens [111]. The results showed that T-2 toxin at 2.5 ppm significantly decreased body weight gain and cumulative feed intake. However, the feed additive counteracted these adverse effects. Moreover, a significant decrease in amylase activity was observed in chickens receiving T-2 toxin, indicating that the additive also protected against duodenal lesions caused by T-2 toxin.
Several commercially available feed additives have the potential to reduce the toxicity of trichothecenes [112–114]. Some of these materials, such as clays, are being used as anti-caking agents to improve flow during feed processing, but they may also serve as mycotoxin-binding agents [110–115]. For example, in a study by Weaver et al. (2013) [110], the feed additives clay and dried yeast were tested to improve the growth and health of pigs chronically challenged with DON. Dietary DON tended to decrease average daily gain and altered the immune system by increasing the levels of monocytes and immunoglobulins. DON also caused tissue damage in the form of liver bile ductal hyperplasia and karyomegaly. However, the additives reduced the effects of DON on the immune system and liver, and also improved growth. Ozonation is another effective and rapid way to degrade DON in wheat, especially in wheat flour [116]. On the other hand, water-soluble DON poses a major threat as a potential organic pollutant to water environmental quality. In wet processing, soluble-toxin molecules can be extracted from the contaminated grain by DON, posing a potentially serious threat to the water environment. In a very recent study [117], ZnO/graphene monolayer hybrid with high photocatalytic performance was shown to effectively degrade DON in water (99% reduction). This work provided important inspiration for the development of graphene-based photocatalysts for use in applications related to trichothecene detoxification and environmental remediation.
These detoxifiers can be broadly categorized into two different classes, mycotoxin binders and mycotoxin modifiers/biotransforming agents. The latter class includes microbes such as bacteria, fungi, yeast, and enzymes that biologically transform the toxins into non-toxic metabolites. LAB, a promising candidate for the biological control of trichothecenes, may biotransform the toxins to less toxic products. In addition, some bacteria including Devosia mutans 17-2-E-8 and Mycofix-S can also biotransform DON into non-toxic compounds, thereby protecting against duodenal lesions caused by T-2 toxin. However, although some decontamination has already been reported, very few commercially available feed additives can reduce the toxicity of mycotoxins such as trichothecenes. Moreover, the biotransforming agents tested to date, such as LAB and yeast, are still limited in the laboratory. Future work should seek to develop methods that allow these agents to be used as real feed additives in the commercial market.
CONCLUSIONS
Trichothecenes inhibit eukaryotic protein synthesis, and oxidative stress is one of their most important underlying toxic mechanisms. They are able to generate free radicals, including ROS, which induce lipid peroxidation that leads to changes in membrane integrity, cellular redox signaling, and cellular antioxidant status. DNA damage correlates with, but precedes, ROS generation and lipid peroxidation. The MAPK signaling pathway is induced by oxidative stress, which also induces caspase-mediated cellular apoptosis pathways. Thus, it is clear that oxidative stress plays important roles in the toxicity of trichothecenes. Notably, however, cells have their own antioxidant defenses against these toxins. The cellular effects of trichothecenes in relation to oxidative stress, as well as effective measures for combating their toxicity, should be addressed further in future studies.
Because oxidative stress plays critical roles in the toxicity of trichothecenes, researchers are trying to identify agents that effectively prevent trichothecene-induced oxidative stress and the associated immunotoxicity. Multiple agents, including vitamins, quercetin, selenium, glucomannan, amino acids, nucleotides, antimicrobial peptides, bacteria, polyunsaturated fatty acids, oligosaccharides, and some plant extracts, exert promising anti-oxidative effects against trichothecenes. Thus, these antioxidant agents should be considered as new toxin control strategies for trichothecenes. We conclude that these compounds inhibit oxidative stress via various mechanisms, e.g., diminishing ROS generation (NAC, EGCG), decreasing DNA damage (nucleotides and coenzyme Q10), reducing lipid peroxidation (vitamins A, C, and E, glucomannan, and rutin), increasing the activity of the antioxidant enzymes (LA, L. plantarum JM113, NAC, and quercetin), blocking the induction of MAPK signaling (SeCS), downregulating the expression of inflammatory genes NF-κB and COX-2 (lutein), inhibiting caspase activity (EGCG and L-carnitine), preventing mRNA overexpression of claudin 3 and CXCL8 (GOS), or inhibiting mitochondrial swelling and cyt-c release (L-carnitine).
Some of these compounds are also employed in the detoxification of other mycotxoins. For example, ochratoxin A (OTA), a widely spread mycotoxin [118], inhibits protein synthesis and energy production, and induces oxidative stress and DNA adduct formation, as well as apoptosis/necrosis and cell cycle arrest [119]. Similar to previous observations in trichothecenes, vitamins A, C, E, EGCG, and quercetin increase kidney cell proliferation, and decrease OTA mediated-ROS generation and DNA fragmentation [120–122]. Understanding these mechanisms of antioxidant agents will aid in the development of methods for detoxification of mycotoxins, as well as new toxin control strategies. In future studies, additional in vivo studies should be carried out to characterize their anti-oxidative activities in animals and humans.
As we discussed in the above chapters, the antioxidant defences induced by dietary supplementation would seem to provide a more reasonable and practical approach for the reduction of oxidative stress levels [123]. In addition, previous studies [123–125] have shown that a variety of proteins function as scavengers of superoxide and hydrogen peroxide, including catalase, glutathione peroxidase, thioredoxin, and the peroxiredoxin protein family. Together with these, a whole host of non-protein scavengers, including, but not limited to, intracellular ascorbate and glutathione [124] also reduce oxidative stress levels. However, the successful implementation of these approaches will probably require a much greater understanding of the pharmacological properties of many of these agents, including their rates of absorption, tissue distribution, metabolism, and the microenvironment in which they must act [123]. Interestingly, Mycobacterium tuberculosis (Mtb), a virulent and deadly bacterial species, has evolved strategies to counter ROS-mediated host defences by expressing enzymes that detoxify a variety of free radical species [126]. It is noteworthy that a novel multicomponent protein complex with both peroxidase and peroxynitrite reductase activity has been identified [127]. A future study should address the possibility of using such multicomponent protein complexes as antioxidant therapies against trichothecenes.
In addition to the antioxidant and detoxification agents described above, several decontamination methods exist, including bacterial and yeast biotransformation and degradation, as well as mycotoxin-binding agents. Because biological reactions offer a specific, irreversible, efficient, and environmentally friendly means of detoxification that leaves neither toxic residues nor any undesired by-products, microbial biotransformation of mycotoxins into non-toxic or less toxic metabolites has been acknowledged as a promising approach. Finally, we should not forget that various food processing methods including baking, boiling, frying, steaming, and extrusion cooking have some positive effects in the controlling of the contamination in foods [128, 129].
Abbreviations
ATA: alimentary toxic aleukia; CAPs: composite antimicrobial peptides; CAT: catalase; mESCs: embryonic stem cells; CREB: cAMP response element-binding protein; CXCL8: IL8/chemokine CXC motif ligand; DHA: docosahexaenoic acid; DON: deoxynivalenol; EGCG: epigallocatechin 3-gallate; EPA: eicosapentaenoic acid; GH: growth hormone; GOS: galacto-oligosaccharides; GPx: glutathione peroxidase; GSH: glutathione; GSH-Px: glutathione peroxidase; GST: glutathione-S-transferase; HO-1: heme oxygenase-1; IgAN: IgA nephropathy; KBD: Kashin-Beck disease; LA: leontopodic acid; LAB: lactic acid bacteria; LGG: L. rhamnosus GG; MAPK: mitogen-activated protein kinases; MDA; malondialdehyde; NAC: N-Acetyl-cysteine; NEO: neosolaniol; NO: nitric oxide; PCNA: proliferating cell nuclear antigen; PKR: RNA-activated protein kinase R; PUFA: polyunsaturated fatty acid; ROS: reactive oxygen species; RSR: ribotoxic stress response; SeCS: selenium chondroitin sulfate; SOD: superoxidae dismutase; TAS: total antioxidant status; TBARS: thiobarbituric acid reactive substances; ΔΨm: mitochondrial membrane potential.
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
FUNDING
This work was supported by the National Natural Science Foundation of China (grant no. 31602114 and 31572575), the Yangtze Fund for Youth Teams of Science and Technology Innovation (2016cqt02), as well as the project of Long-term development plan UHK.
REFERENCES
1. Wu Q, Dohnal V, Kuca K, Yuan Z. Trichothecenes: structure-toxic activity relationships. Curr Drug Metab. 2013; 14:641-660.
2. Wei S, van der Lee T, Verstappen E, van Gent M, Waalwijk C. Targeting trichothecene biosynthetic genes. Methods Mol Biol. 2017; 1542:173-189.
3. Wu Q, Vlastimil D, Huang L, Kuca K, Yuan Z. Metabolic pathways of trichothecenes. Drug Metab Rev. 2010; 42:250-267.
4. Wu QH, Wang X, Yang W, Nüssler AK, Xiong LY, Kuča K, Dohnal V, Zhang XJ, Yuan ZH. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: an update. Arch Toxicol. 2014; 88:1309-1326.
5. Wu Q, Wang X, Wan D, Li J, Yuan Z. Crosstalk of JNK1-STAT3 is critical for RAW264.7 cell survival. Cell Signal. 2014; 26:2951-2960.
6. Wu Q, Huang L, Liu Z, Yao M, Wang Y, Dai M, Yuan Z. A comparison of hepatic in vitro metabolism of T-2 toxin in rats, pigs, chickens and carp. Xenobiotica. 2011; 41:863-873.
7. Liu X, Guo P, Liu A, Wu Q, Xue X, Dai M, Hao H, Qu W, Xie S, Wang X, Yuan Z. Nitric oxide (NO)-mediated mitochondrial damage plays a critical role in T-2 toxin-induced apoptosis and growth hormone deficiency in rat anterior pituitary GH3 cells. Food Chem Toxicol. 2017; 102:11-23.
8. Beyer M, Ferse I, Mulac D, Wurthwein EU, Humpf HU. Structural elucidation of T-2 toxin thermal degradation products and investigations toward their occurrence in retail food. J Agric Food Chem. 2009; 57:1867-1875.
9. Morcia C, Tumino G, Ghizzoni R, Badeck FW, Lattanzio VM, Pascale M, Terzi V. Occurrence of Fusarium langsethiae and T-2 and HT-2 Toxins in Italian Malting Barley. Toxins (Basel). 2016.
10. Bolechova M, Benesova K, Belakova S, Caslavsky J, Pospichalova M, Mikulikova R. Determination of seventeen mycotoxins in barley and malt in the Czech Republic. Food Control. 2015; 47:108-113.
11. Aureli G, Amoriello T, Belocchi A, D’Egidio MG, Fornara M, Melloni S, Quaranta F. Preliminary survey on the co-occurrence of DON and T2+HT2 toxins in drum wheat in Italy. Cereal Res Commun. 2015; 43:481-491.
12. Kuca K, Pohanka M. Chemical warfare agents. Mol Clin Environ Toxicol. 2010; 100:543-558.
13. Tucker JB. The “yellow rain” controversy: lessons for arms control compliance. Nonprolif Rev. 2001; 8:25-42.
14. Hendry KM, Cole EC. A review of mycotoxins in indoor air. J Toxicol Environ Health. 1993; 38:183-198.
15. Bennett JW, Klich M. Mycotoxins. Clin Microbiol Rev. 2003; 16:497-516.
16. Wang Z, Wu Q, Kuča K, Dohnal V, Tian Z. Deoxynivalenol: signaling pathways and human exposure risk assessment--an update. Arch Toxicol. 2014; 88:1915-1928.
17. Payros D, Alassane-Kpembi I, Pierron A, Loiseau N, Pinton P, Oswald IP. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch Toxicol. 2016; 90:2931-2957.
18. Lei R, Jiang N, Zhang Q, Hu S, Dennis BS, He S, Guo X. Prevalence of selenium, T-2 toxin, and deoxynivalenol in Kashin-Beck Disease areas in Qinghai province, Northwest China. Biol Trace Elem Res. 2016; 171:34-40.
19. Malachova A, Dzuman Z, Veprikova Z, Vaclavikova M, Zachariasova M, Hajslova J. Deoxynivalenol, deoxynivalenol-3-glucoside, and enniatins: the major mycotoxins found in cereal-based products on the Czech market. J Agric Food Chem. 2011; 59:12990-12997.
20. Camerini M, Amoriello T, Aureli G, Belocchi A, Fornara M, Melloni S, Quaranta F. Deoxynivalenol content in Italian Organic Durum Wheat: results of a six-year survey. Cereal Res Commun. 2016, 44:122-130.
21. Vidal A, Marín S, Ramos AJ, Cano-Sancho G, Sanchis V. Determination of aflatoxins, deoxynivalenol, ochratoxin A and zearalenone in wheat and oat based bran supplements sold in the Spanish market. Food Chem Toxicol. 2013; 53:133-138.
22. Pleadin J, Zadravec M, Perši N, Vulić A, Jaki V, Mitak M. Mould and mycotoxin contamination of pig feed in northwest Croatia. Mycotoxin Res. 2012; 28:157-162.
23. Tran ST, Smith T, Girgis GN. A survey of free and conjugated deoxynivalenol in the 2008 corn crop in Ontario, Canada. J Sci Food Agric. 2012; 92:37-41.
24. Brockmeyer A, Thielert G. Deoxynivalenol (DON) in Hartweizen. Mycotoxin Res. 2004; 20:37-41.
25. El Golli E, Hassen W, Bouslimi A, Bouaziz C, Ladjimi MM, Bacha H. Induction of Hsp 70 in Vero cells in response to mycotoxins cytoprotection by sub-lethal heat shock and by Vitamin E. Toxicol Lett. 2006; 166:122-130.
26. Krishnaswamy R, Devaraj SN, Padma VV. Lutein protects HT-29 cells against deoxynivalenol-induced oxidative stress and apoptosis: prevention of NF-kappaB nuclear localization and down regulation of NF-kappaB and Cyclo-Oxygenase-2 expression. Free Radic Biol Med. 2010; 49:50-60.
27. Chaudhary M, Rao PV. Brain oxidative stress after dermal and subcutaneous exposure of T-2 toxin in mice. Food Chem Toxicol. 2010; 48:3436-3442.
28. Wu J, Jing L, Yuan H, Peng S. T-2 toxin induces apoptosis in ovarian granulose cells of rats through reactive oxygen species mediated mitochrondrial pathway. Toxicol Lett. 2011; 202:168-177.
29. Fang H, Wu Y, Guo J, Rong J, Ma L, Zhao Z, Zuo D, Peng S. T-2 toxin induces apoptosis in differentiated murine embryonic stem cells through reactive oxygen species-mediated mitochondrial pathway. Apoptosis. 2012; 17:895-907.
30. Chaudhari M, Jayaraj R, Bhaskar AS, Lakshmana Rao PV. Oxidative stress induction by T-2 toxin cause DNA damage and triggers apoptosis via caspase pathway in human cervical cancer cells. Toxicology. 2009; 262:153-161.
31. Chaudhari M, Jayaraj R, Santhosh SR, Lakshmana Rao PV. Oxidative damage and gene expression profile of antioxidant enzymes after T-2 toxin exposure in mice. J Biochem Mol Toxicol. 2009; 23:212-221.
32. Sehata S, Kiyosawa N, Atsumi F, Ito K, Yamoto T, Teranishi M, Uetsuka K, Uetsuka K, Nakayama H, Doi K. Microarray analysis of T-2 toxin-induced liver, placenta and fetal liver lesions in pregnant rats. Exp Toxicol Pathol. 2005; 57:15-28.
33. Van De WJ, Romier B, Larondelle Y, Schneider YJ. Influence of deoxynivalenol on NF-κB activation and IL-8 secretion in human intestinal Caco-2 cells. Toxicol lett. 2008; 177:205-214.
34. Zhou HR, Pestka JJ. Deoxynivalenol-induced apoptosis mediated by p38 MAPK-dependent p53 gene induction in RAW264.7 macrophages. Toxicologist. 2003; 72:330.
35. Bin-Umer MA, McLaughlin JE, Butterly MS, McCormick S, Tumer NE. Elimination of damaged mitochondria through mitophagy reduces mitochondrial oxidative stress and increases tolerance to trichothecenes. Proc Natl Acad Sci U S A. 2014; 111:11798-11803.
36. Li D, Ye Y, Lin S, Deng L, Fan X, Zhang Y, Deng X, Li Y, Yan H, Ma Y. Evaluation of deoxynivalenol-induced toxic effects on DF-1 cells in vitro: cell-cycle arrest, oxidative stress, and apoptosis. Environ Toxicol Pharmacol. 2014; 37:141-149.
37. Strasser A, Carra M, Ghareeb K, Awar W, Bohm J. Protective effects of antioxidants on deoxynivalenol-induced damage in murine lymphoma cells. Mycotoxin Res. 2013; 29:203-208.
38. Leśniak-Walentyn A, Kolesarova A, Medvedova M, Maruniakova N, Capcarova M, Kalafova A, Hrabia A, Sirotkin AV. Proliferation and apoptosis in the rabbit ovary after administration of T-2 toxin and quercetin. J Anim Feed Sci. 2013; 22:264-271.
39. Keshavarz SA, Memarbashi A, Balali M. Preventive effect of selenium on T-2 toxin membrane toxicity. Adv Exp Med Biol. 2001; 500:463-466.
40. He JW, Hassan YI, Perilla N, Li XZ, Boland GJ, Zhou T. Bacterial epimerization as a route for deoxynivalenol detoxification: the influence of growth and environmental conditions. Front Microbiol. 2016; 7:572.
41. Xiao H, Wu MM, Tan BE, Yin YL, Li TJ, Xiao DF, Li L. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: I. Growth performance, immune function, and antioxidation capacity. J Anim Sci. 2013; 91:4772-4780.
42. Bócsai A, Pelyhe C, Zándoki E, Ancsin Z, Szabó-Fodor J, Erdélyi M, Mézes M, Balogh K. Short-term effects of T-2 toxin exposure on some lipid peroxide and glutathione redox parameters of broiler chickens. J Anim Physiol Anim Nutr (Berl). 2016; 100:520-525.
43. Wu J, Tu D, Yuan LY, Yuan H, Wen LX. T-2 toxin exposure induces apoptosis in rat ovarian granulosa cells through oxidative stress. Environ Toxicol Pharmacol. 2013; 36:493-500.
44. Ngampongsa S, Hanafusa M, Ando K, Ito K, Kuwahara M, Yamamoto Y, Yamashita M, Tsuru Y, Tsubone H. Toxic effects of T-2 toxin and deoxynivalenol on the mitochondrial electron transport system of cardiomyocytes in rats. J Toxicol Sci. 2013; 38:495-502.
45. Fang H, Cong L, Zhi Y, Xu H, Jia X, Peng S. T-2 toxin inhibits murine ES cells cardiac differentiation and mitochondrial biogenesis by ROS and p-38 MAPK-mediated pathway. Toxicol Lett. 2016; 258:259-266.
46. Mu P, Xu M, Zhang L, Wu K, Wu J, Jiang J, Chen Q, Wang L, Tang X, Deng Y. Proteomic changes in chicken primary hepatocytes exposed to T-2 toxin are associated with oxidative stress and mitochondrial enhancement. Proteomics. 2013; 13:3175-3188.
47. Yuan G, Wang Y, Yuan X, Zhang T, Zhao J, Huang L, Peng S. T-2 toxin induces developmental toxicity and apoptosis in zebrafish embryos. J Environ Sci (China). 2014; 26:917-925.
48. Weidner M, Hüwel S, Ebert F, Schwerdtle T, Galla HJ, Humpf HU. Influence of T-2 and HT-2 toxin on the blood-brain barrier in vitro: new experimental hints for neurotoxic effects. PLoS One. 2013; 8:e60484.
49. Chen JH, Xue S, Li S, Wang ZL, Yang H, Wang W, Song D, Zhou X, Chen C. Oxidant damage in Kashin-Beck disease and a rat Kashin-Beck disease model by employing T-2 toxin treatment under selenium deficient conditions. J Orthop Res. 2012; 30:1229-1237.
50. Matejova I, Faldyna M, Modra H, Blahova J, Palikova M, Markova Z, Franc A, Vicenova M, Vojtek L, Bartonkova J, Sehonova P, Hostovsky M, Svobodova Z. Effect of T-2 toxin-contaminated diet on common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 2017; 60:458-465.
51. Deng Y, Wang Y, Zhang X, Sun L, Wu C, Shi Q, Wang R, Sun X, Bi S, Gooneratne R. Effects of T-2 toxin on Pacific white shrimp Litopenaeus vannamei: growth, and antioxidant defenses and capacity and histopathology in the hepatopancreas. J Aquat Anim Health. 2017; 29:15-25.
52. Zhang X, Jiang L, Geng C, Cao J, Zhong L. The role of oxidative stress in deoxynivalenol-induced DNA damage in HepG2 cells. Toxicon. 2009; 54:513-518.
53. Ren Z, Wang Y, Deng H, Deng Y, Deng J, Zuo Z, Wang Y, Peng X, Cui H, Shen L. Deoxynivalenol induces apoptosis in chicken splenic lymphocytes via the reactive oxygen species-mediated mitochondrial pathway. Environ Toxicol Pharmacol. 2015; 39:339-346.
54. Wang X, Xu W, Fan M, Meng T, Chen X, Jiang Y, Zh D, Hu W, Gong J, Feng S, Wu J, Li Y. Deoxynivalenol induces apoptosis in PC12 cells via the mitochondrial pathway. Environ Toxicol Pharmacol. 2016; 43:193-202.
55. Yang W, Yu M, Fu J, Bao W, Wang D, Hao L, Yao P, Nüssler AK, Yan H, Liu L. Deoxynivalenol induced oxidative stress and genotoxicity in human peripheral blood lymphocytes. Food Chem Toxicol. 2014; 64:383-396.
56. Bodea GO, Munteanu MC, Dinu D, Serban AI, Roming FI, Costache M, Dinischiotu A. Influence of deoxynivalenol on the oxidative status of HepG2 cells. Rom Biotechnol Lett. 2009; 14:4349-4359.
57. Zhuang Z, Yang D, Huang Y, Wang S. Study on the apoptosis mechanism induced by T-2 toxin. PLoS One. 2013; 8:e831.
58. Bocsai A, Ancsin Z, Fernye C, Zandoki E, Szabo-Fodor J, Erdelyi M, Mezes M, Balogh K. Dose-dependent short-term effects of T-2 toxin exposure on lipid peroxidation and antioxidant parameters of laying hens. Eur Poult Sci. 2015. https://doi.org/10.1399/eps.2015.115.
59. Rizzo AF, Atroshi F, Ahotupa M, Sankari S, Elovaara E. Protective effect of antioxidants against free radical-mediated lipid peroxidation induced by DON or T-2 toxin. Zentralbl Veterinarmed A. 1994; 41:81-90.
60. Rehak M, Fric E, Wiedemann P. [Lutein and antioxidants in the prevention of age-related macular degeneration]. [Article in German]. Opththalmologe. 2008; 105:37-38.
61. Atroshi F, Rizzo A, Biese I, Veijalainen P, Antila E, Westermarck T. T-2 toxin-induced DNA damage in mouse livers, the effect of pretreatment with coenzyme Q10 and alpha-tocopherol. Mol Aspects Med. 1997; 18:255-258.
62. Capcarova M, Petruska P, Zbynovska K, Kolesarova A, Sirotkin AV. Changes in antioxidant status of porcine ovarian granulosa cells after quercetin and T-2 toxin treatment. J Environ Sci Health B. 2015; 50:201-206.
63. Wrobel JK, Power R, Toborek M. Biological activity of selenium: revisited. IUBMB Life. 2016; 68:97-105.
64. Kieliszek M, Błażejak S. Current knowledge on the importance of selenium in food for living organisms: a review. Molecules. 2016.
65. Salimian J, Arefpour MA, Riazipour M, Poursasan N. Immunomodulatory effects of selenium and vitamin E on alterations in T lymphocyte subsets induced by T-2 toxin. Immunopharmacol Immunotoxicol. 2014; 36:275-281.
66. Yao YF, Pei FX, Kang PD. Selenium, iodine, and the relation with Kashin-Beck disease. Nutrition. 2011; 27:1095-1100.
67. Guan F, Li SY, Wang ZL, Yang HJ, Xue SH, Wang W, Song DQ, Zhou XR, Zhou W, Chen JH, Caterson B, Hughes C. Histopathology of chondronecrosis development in knee articular cartilage in a rat model of Kashin–Beck disease using T-2 toxin and selenium deficiency conditions. Rheumatol Int. 2013; 33:157-166.
68. Tan JA, Zhu WY, Wang WY, Li RB, Zou SF, Wang DC, Yang LS. Selenium in soil and endemic diseases in China. Sci Total Environ. 2002; 284:227-235.
69. Han J, Guo X. Down-regulation of ATF2 in the inhibition of T-2-toxininduced chondrocyte apoptosis by selenium chondroitin sulfate nanoparticles. J Nanopart Res. 2013; 15:2037.
70. Keithley J, Swanson B. Glucomannan and obesity: a critical review. Altern Ther Health Med. 2005; 11:30-34.
71. González Canga A, Fernández Martínez N, Sahagún AM, García Vieitez JJ, Díez Liébana MJ, Calle Pardo AP, Castro Robles LJ, Sierra Vega M. Glucomannan: properties and therapeutic applications. Nutr Hosp. 2004; 19:45-50.
72. Meissonnier GM, Raymond I, Laffitte J, Cossalter AM, Pinton P, Benoit E, Bertin G, Galtier P, Oswald IP. Dietary glucomannan improves the vaccinal response in pigs exposed to aflatoxin B-1 or T-2 toxin. World Mycotoxin J. 2009; 2:161-172.
73. Verbrugghe E, Croubels S, Vandenbroucke V, Goossens J, De Backer P, Eeckhout M, De Saeger S, Boyen F, Leyman B, Van Parys A, Haesebrouck F, Pasmans F. A modified glucomannan mycotoxin-adsorbing agent counteracts the reduced weight gain and diminishes cecal colonization of Salmonella Typhimurium in T-2 toxin exposed pigs. Res Vet Sci. 2012; 93:1139-1141.
74. Dvorska JE, Pappas AC, Karadas F, Speake BK, Surai PF. Protective effect of modified glucomannans and organic selenium against antioxidant depletion in the chicken liver due to T-2 toxin-contaminated feed consumption. Comp Biochem Physiol C Toxicol Pharmacol. 2007; 145:582-587.
75. Hou R, Jiang C, Zheng Q, Wang CF, Xu JR. The AreA transcription factor mediates the regulation of deoxynivalenol (DON) synthesis by ammonium and cyclic adenosine monophosphate (cAMP) signalling in Fusarium graminearum. Mol Plant Pathol. 2015; 16:987-999.
76. Ueno T, Yamada M, Sugita Y, Ogawa T. N-acetylcysteine protects TMJ chondrocytes from oxidative stress. J Dent Res. 2011; 90:353-359.
77. He SJ, Hou JF, Dai YY, Zhou ZL, Deng YF. N-acetyl-cysteine protects chicken growth plate chondrocytes from T-2 toxin-induced oxidative stress. J Appl Toxicol. 2012; 32:980-985.
78. Wu L, Wang W, Yao K, Zhou T, Yin J, Li T, Yang L, He L, Yang X, Zhang H, Wang Q, Huang R, Yin Y. Effects of dietary arginine and glutamine on alleviating the impairment induced by deoxynivalenol stress and immune relevant cytokines in growing pigs. PLoS One. 2013; 8:e69502.
79. Wu M, Xiao H, Ren W, Yin J, Tan B, Liu G, Li L, Nyachoti CM, Xiong X, Wu G. Therapeutic effects of glutamic acid in piglets challenged with deoxynivalenol. PLoS One. 2014; 9:e100591.
80. Oyanagi E, Yano H, Uchida M, Utsumi K, Sasaki J. Protective action of L-carnitine on cardiac mitochondrial function and structure against fatty acid stress. Biochem Biophys Res Commun. 2011; 412:61-67.
81. Moosavi M, Rezaei M, Kalantari H, Behfar A, Varnaseri G. l-carnitine protects rat hepatocytes from oxidative stress induced by T-2 toxin. Drug Chem Toxicol. 2016; 39:445-450.
82. Frankic T, Pajk T, Rezar V, Levart A, Salobir J. The role of dietary nucleotides in reduction of DNA damage induced by T-2 toxin and deoxynivalenol in chicken leukocytes. Food Chem Toxicol. 2006; 44:1838-1844.
83. Brogden KA. Antimicrobial peptides: pore formers or metabolic inhibitors in bacteria? Nat Rev Microbiol. 2005; 3:238-250.
84. Xiao H, Tan BE, Wu MM, Yin YL, Li TJ, Yuan DX, Li L. Effects of composite antimicrobial peptides in weanling piglets challenged with deoxynivalenol: II. Intestinal morphology and function. J Anim Sci. 2013; 91:4750-4756.
85. Kalaiselvi P, Rajashree K, Priya LB, Padma VV. Cytoprotective effect of epigallocatechin-3-gallate against deoxynivalenol-induced toxicity through anti-oxidative and anti-inflammatory mechanisms in HT-29 cells. Food Chem Toxicol. 2013; 56:110-118.
86. Schwaiger S, Cervellati R, Seger C, Ellmerer EP, About N, Renimel I, Godenir C, Andre P, Gafne F, Stuppner H. Leontopodic acid—a novel highly substituted glucaric acid derivative from Edelweiss (Leontopodium alpinum Cass.) and its antioxidative and DNA protecting properties. Tetrahedron. 2005; 61:4621-4630.
87. Costa S, Schwaiger S, Cervellati R, Stuppner H, Speroni E, Guerra MC. In vitro evaluation of the chemoprotective action mechanisms of leontopodic acid against aflatoxin B1 and deoxynivalenol-induced cell damage. J Appl Toxicol. 2009; 29:7-14.
88. Hemmati AA, Kalantari H, Jalali A, Rezai S, Zadeh HH. Healing effect of quince seed mucilage on T-2 toxin-induced dermal toxicity in rabbit. Exp Toxicol Pathol. 2012; 64:181-186.
89. Silva BM, Andrade PB, Ferreres F, Seabra RM, Oliveira MB, Ferreira MA. Composition of quince (Cydonia oblonga Miller) seeds: phenolics, organic acids and free amino acids. Nat Prod Res. 2005; 19:275-281.
90. Magalhaes AS, Silva BM, Pereira JA, Andrade PB, Valentao P, Carvalho M. Protective effect of quince (Cydonia oblonga Miller) fruit against oxidative hemolysis of human erythrocytes. Food Chem Toxicol. 2009; 47:1372-1377.
91. El-Saw NM, Al-Seeni MN. Assessment of flavonoids as rutin for detoxification of T-2 toxin. J Appl Anim Res. 2009; 35:57-60.
92. Leal M, Shimada A, Ruíz F, González E, Mejía D. Effect of lycopene on lipid peroxidation and glutathione-dependent enzymes induced by T-2 toxin in vivo. Toxicol Lett. 1999; 109:1-10.
93. Yang X, Li L, Duan Y, Yang X. Antioxidant activity of Lactobacillus plantarum JM113 in vitro and its protective effect on broiler chickens challenged with deoxynivalenol. J Anim Sci. 2017; 95:837-846.
94. Wan ML, Turner PC, Allen KJ, El-Nezami H. Lactobacillus rhamnosus GG modulates intestinal mucosal barrier and inflammation in mice following combined dietary exposure to deoxynivalenol and zearalenone. J Funct Foods. 2016; 22:34-43.
95. Shi Y, Pestka JJ. Attenuation of mycotoxin-induced IgA nephropathy by eicosapentaenoic acid in the mouse: dose response and relation to IL-6 expression. J Biochem. 2006; 17:697-706.
96. Donadio JV, Grande JP. The role of fish oil/omega-3 fatty acids in the treatment of IgA nephropathy. Semin Nephrol. 2004; 24:225-243.
97. Shi Y, Pestka JJ. Mechanisms for suppression of interleukin-6 expression in peritoneal macrophages from docosahexaenoic acid-fed mice. J Nutr Biochem. 2009; 20:358-368.
98. Akbari P, Braber S, Alizadeh A, Verheijden KA, Schoterman MH, Kraneveld AD, Garssen J, Fink-Gremmels J. Galacto-oligosaccharides protect the intestinal barrier by maintaining the tight junction network and modulating the inflammatory responses after a challenge with the mycotoxin deoxynivalenol in Human caco-2 cell monolayers and B6C3F1 mice. J Nutr. 2015; 145:1604-1613.
99. Alizadeh A, Braber S, Akbari P, Kraneveld A, Garssen J, Fink-Gremmels J. Deoxynivalenol and its modified forms: are there major differences? Toxins (Basel). 2016.
100. Wu Q, Jezkova A, Yuan Z, Pavlikova L, Dohnal V, Kuca K. Biological degradation of aflatoxins. Drug Metab Rev. 2009; 41:1-7.
101. Awad WA, Ghareeb K, Bohm J, Zentek J. Decontamination and detoxification strategies for the Fusarium mycotoxi deoxynivalenol in animal feed and the effectiveness of microbial biodegradation. Food Addit Contam. 2010; 27:510-520.
102. Leroy F, De Vuyst L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci Technol. 2004; 15:67-78.
103. Franco TS, Garcia S, Hirooka EY, Ono YS, dos Santos JS. Lactic acid bacteria in the inhibition of Fusarium graminearum and deoxynivalenol detoxification. J Appl Microbiol. 2011; 111:739-748.
104. Dalié DKD, Deschamps AM, Richard-Forget F. Lactic acid bacteria – potential for control of mould growth and mycotoxins: a review. Food Control. 2010; 21:370-380.
105. Zou ZY, He ZF, Li HJ, Han PF, Meng X, Zhang Y, Zhou F, Ouyang KP, Chen XY, Tang J. In vitro removal of deoxynivalenol and T-2 toxin by Lactic acid bacteria. Food Sci Biotechnol. 2012; 21:1677-1683.
106. Zhao L, Li X, Ji C, Rong X, Liu S, Zhang J, Ma Q. Protective effect of Devosia sp. ANSB714 on growth performance, serum chemistry, immunity function and residues in kidneys of mice exposed to deoxynivalenol. Food Chem Toxicol. 2016; 92:143-149.
107. Gastélum-Martínez E, Compant S, Taillandier P, Mathieu F. Control of T-2 toxin in Fusarium langsethiae and Geotrichum candidum co-culture. Arh Hig Rada Toksikol. 2012; 63:447-456.
108. McCormick SP, Price NP, Kurtzman CP. Glucosylation and other biotransformations of T-2 toxin by yeasts of the trichomonascus clade. Appl Environ Microbiol. 2012; 78:8694-8702.
109. de Souza AF, Borsato D, Lofrano AD, de Oliyeira AS, Ono MA, Bordini JG, Hirozawa MT, Yabe MJ, Ono EY. In vitro removal of deoxynivalenol by a mixture of organic and inorganic adsorbents. World Mycotoxin J. 2015; 8:113-119.
110. Weaver AC, See MT, Hansen JA, Kim YB, De Souza AL, Middleton TF, Kim SW. The use of feed additives to reduce the effects of aflatoxin and deoxynivalenol on pig growth, organ health and immune status during chronic exposure. Toxins (Basel). 2013; 5:1261-1281.
111. Diaz GJ, Vargas ML, Cortes A. Evaluation of the supplementation of a feed additive as a potential protector against the adverse effects of 2.5 ppm T-2 toxin on growing broiler chickens. Arq Bras Med Vet Zootec. 2016; 68:709-715.
112. Schatzmayr G, Zehner F, Taubel M, Schatzmayr D, Klimitsch A, Loibner AP, Binder EM. Microbiologicals for deactivating mycotoxins. Mol Nutr Food Res. 2006; 50:543-551.
113. Huwig A, Freimund S, Kappeli O, Dutler H. Mycotoxin detoxification of animal feed by different absorbents. Toxicol Lett. 2001; 122:179-188.
114. Ramos AJ, Fink-Gremmels J, Hernandez E. Prevention of toxic effects of mycotoxins by means of nonnutritive adsorbent compounds. J Food Prot. 1996; 59:631-641.
115. Choy JH, Choi SJ, Oh JM, Park T. Clay minerals and layered double hydroxides for novel biological applications. Appl Clay Sci. 2007; 36:122-132.
116. Wang L, Luo Y, Luo X, Wang R, Li Y, Li Y, Shao H, Chen Z. Effect of deoxynivalenol detoxification by ozone treatment in wheat grains. Food Control. 2016; 66:137-144.
117. Bai X, Sun C, Liu D, Luo X, Li D, Wang J, Wang N, Chang X, Zong R, Zhu Y. Photocatalytic degradation of deoxynivalenol using graphene/ZnOhybrids in aqueous suspension. Appl Catal B. 2017; 204:11-20.
118. Wu Q, Dohnal V, Huang L, Kuca K, Wang X, Chen G, Yuan Z. Metabolic pathways of ochratoxin A. Curr Drug Metab. 2010; 12:1-10.
119. Kőszegi T, Poór M. Ochratoxin A: molecular interactions, mechanisms of toxicity and prevention at the molecular level. Toxins (Basel). 2016; 8:111.
120. Costa S, Utan A, Cervellati R, Speroni E, Guerra MC. Catechins: natural free-radical scavengers against Ochratoxin A-induced cell damage in a pig kidney cell line (LLC-PK1). Food Chem Toxicol. 2007; 45:1910-1917.
121. Baldi A, Losio MN, Cheli F, Rebucci R, Sangalli L, Fusi E, Bertasi B, Pavoni E, Carli S, Politis I. Evaluation of the protective effects of alpha-tocopherol and retinol against Ochratoxin A cytotoxicity. Br J Nutr. 2004; 91:507-512.
122. Ramyaa P, Padma VV. Ochratoxin-induced toxicity, oxidative stress and apoptosis ameliorated by quercetin—modulation by Nrf2. Food Chem Toxicol. 2013; 62:205-216.
123. Finkel T, Holbrook NJ. Oxidants, oxidative stress and the biology of ageing. Nature. 2000; 408:239-247.
124. Finkel T. Oxidant signals and oxidative stress. Curr Opin Cell Biol. 2003; 15:247-254.
125. Ozben T. Oxidative stress and apoptosis: impact on cancer therapy. J Pharm Sci. 2007; 96:2181-2196.
126. Köster S Upadhyay S, Chandra P, Papavinasasundaram K, Yang G, Hassan A, Grigsby SJ, Mittal E, Park HS, Jones V, Hsu FF, Jackson M, Sassetti CM, Philips JA. Mycobacterium tuberculosis is protected from NADPH oxidase and LC3-associated phagocytosis by the LCP protein CpsA. Proc Natl Acad Sci U S A. 2017. https://doi.org/10.1073/pnas.1707792114.
127. Bryk R, Lima CD, Erdjument-Bromage H, Tempst P, Nathan C. Metabolic enzymes of mycobacteria linked to antioxidant defense by a thioredoxin-like protein. Science. 2002; 295:1073-1077.
128. Wu Q, Kuča K, Humpf HU, Klímová B, Cramer B. Fate of deoxynivalenol and deoxynivalenol-3-glucoside during cereal-based thermal food processing: a review study. Mycotoxin Res. 2017; 33:79-91.
129. Schmidt H, Becker S, Cramer B, Humpf HU. Impact of mechanical and thermal energies on the degradation of T 2 and HT 2 toxins during extrusion cooking of oat flour. J Agr Food Chem. 2017; 65:4177-4183.