
INTRODUCTION
Gliomas, the most common neoplasms found in primary brain tumors, are graded according to histologic subtype [1]. WHO (World Health Organization) defines glioblastomas (GBMs) as grade IV gliomas, which are the most common primary brain tumor in adults [2]. The prognosis of patients with GBMs is very poor. The mean survival is only from 10 to 14 months [3, 4], indicating variable clinical behavior and response to therapy.
GBMs are difficult to treat for several reasons. First, a large number of drugs cannot get into tumor sites directly due to the existence of the blood-brain barrier (BBB). Second, since GBMs can infiltrate the surrounding tissues, the standard therapeutic regimens, including surgery, radiation, and chemotherapy, cannot remove them completely. Finally, chemotherapy drugs, which are specific to one tumor cell type, cannot kill all tumor cells due to their heterogeneity.
Temozolomide (TMZ), an alkylating agent prodrug, is an orally administered chemotherapy with a good penetration of the BBB and limited side effects. It is the only US FDA-approved drug for treating refractory anaplastic astrocytomas in adult patients since 1999 [5], and newly diagnosed GBMs in adult patients since 2005 [6]. Its active form can methylate DNA at the sites of O6-guanine, N7-guanine, or N3-adenine. The DNA damage is primarily mediated by the O6-methylguanine (O6-MeG), which induces double-strand breaks and base mispairing, and thereby causes apoptosis and cell death [7, 8]. Currently, radiotherapy with concomitant and adjuvant TMZ is the gold standard for patients with newly diagnosed GBMs [9]. Although the combined use of TMZ and radiation have improved patients’ survival more than radiation alone (median survival from 12.1 to 14.6 months in normal population [10], and median survival from 7.6 to 9.3 months in elder population [11]), almost all patients experience tumor progression or recurrence. Local tumor progression is the predominant pattern of treatment failure [12]. In general, tumor recurrence is associated with poor survival because treatment options are limited [13].
Therapeutic resistance is a critical factor affecting the survival rate of cancer patients. TMZ and radiation resistance are two major issues in the management of GBMs. The existing mechanisms of DNA repair in GBM cells limit the cytotoxic effect of TMZ in treatment of GBMs [14, 15]. Moreover, the results obtained from studies of intrinsic and acquired TMZ resistance in GBM cells support the idea that TMZ resistance is not mediated by only a single molecular event, but by multiple ones. Therefore, exploring the possible mechanisms of therapeutic resistance within GBM cells is an important mission of neuro-oncologists, and identification of biomarkers that are associated with therapeutic resistance in GBMs might provide a feasible way for pursuing this goal.
Recent advances in the development of molecular markers by genome-wide studies of the CNS tumors have improved our understanding of the biology in these tumors. To date, however, only a few molecular markers really have clinical relevance in the therapeutic decision-making of GBM patients. MGMT promoter methylation in high-grade astrocytomas and co-deletion of 1p/19q in oligodendrogliomas are proven prognostic and predictive markers that play a role in standard practice, and mutations of IDH1 or IDH2 are of strong prognostic value in lower grade gliomas (LGG), which are the most widely validated biomarkers in neuro-oncology currently [16, 17].
Although the promoter methylation status of MGMT is shown to be a useful prognostic or predicting biomarker in the elderly patients with newly diagnosed GBM [18, 19], the role of MGMT in clinical decision-making remains limited and the routine analysis of the MGMT promoter methylation status is restricted to only a few clinical scenarios. Although various testing methods, including methylation-specific polymerase chain reaction (MS-PCR), pyrosequencing, methylation-specific multiplex ligation-dependent probe amplification, and immunohistochemistry (IHC), are currently being used, however, there is still no uniform methodology for the MGMT testing. Therefore, standardized procedures should be created to allow inter-laboratory reproducibility, especially if future treatment decisions will be based on these results [20]. In fact, the IHC analysis of MGMT protein lacks a significant correlation with the MGMT promoter methylation status, and is not routinely used for diagnostic purposes due to the inter-observer variability [21]. Thus, there is an urgent need to identify a specific and sensitive biomarker for prognosis and predicting the response to therapy, which may provide better therapeutic guide in the management of glioma patients [22–24].
In this study, we identified BICD1 expression as a potential biomarker from cell-based microarray data, and validated its prognostic value in clinical datasets of the TCGA GBM and GBMLGG cohorts, and the CCGA glioma cohort. BICD Cargo Adaptor 1 (BICD1), is a human homologue of the Drosophila Bicaudal-D gene [25]. This is the first report to investigate the association of BICD1 expression with the prognosis and therapeutic outcome of GBM patients. Our results confirmed our hypothesis that BICD1 expression is a potential biomarker for prognosis and predicting the response to therapy in patients with GBMs.
RESULTS
An overview of the overall survival of patients in the TCGA GBMLGG (glioblastoma and lower grade glioma) cohort
From the TCGA GBMLGG cohort data of patients with gliomas (http://www.xenabrowser.net/), the lower grade gliomas (grades II and III gliomas, LGGs) were the less malignant phenotype. The median survival time was 7.29±0.86 years and 5-year survival rate was 61.5% in patients with LGGs. The grade IV gliomas (Glioblastomas, GBMs) were the most malignant phenotype in all gliomas. The median survival time was only 1.13±0.07 years and 2-year survival rate was 20.9% in patients with GBMs. The median survival time was 4.09±0.44 years and 5-year survival rate was 45.7% in all glioma patients (Table 1).
Table 1: The overall survival of patients with different histological grade in the TCGA GBMLGG cohort
Histological grade |
Patient numbers |
Death event number |
Median value |
5-year survival rate |
2-year survival rate |
|||
|---|---|---|---|---|---|---|---|---|
Survival time (years) |
S.D. |
Lower limit (95% CI) |
Upper limit (95% CI) |
|||||
LGG (Grades II+III) |
524 |
133 |
7.29 |
0.86 |
5.6 |
8.98 |
61.5% |
85.6% |
GBM (Grade IV) |
165 |
127 |
1.13 |
0.07 |
1.00 |
1.27 |
0% |
20.9% |
All (Grades II+III+IV) |
689 |
260 |
4.09 |
0.44 |
3.22 |
4.96 |
45.7% |
69.8% |
Discovery of candidate markers by analyzing the TMZ-related genomic alterations in GBM cell lines and validating their prognostic values in the TCGA GBM database
Biomarkers have been developed through several ways, including cell line models [26]. U87 and T98G are two well-known GBM cell lines [27, 28]. U87 is sensitive to TMZ, but T98G is resistant to it. The EC50 of TMZ in U87 and T98G have been well studied by a large number of research groups [29–32]. According to our result of MTT assay, the EC50 of TMZ was 400μM in T98G, and 20μM in U87 (Figure 1A), which were compatible with other study groups’ data. In this study, U87 and T98G cells were treated with their EC50 of TMZ for 6 hours and the genomic alterations in each cell line were observed using gene expression microarrays. A total of 13 probes, which were up-regulated in T98G and down-regulated in U87 after TMZ treatment, were identified by the method of hierarchical clustering analysis (Figure 1B). The expression changes of these probes in GBM cells under different conditions (control vs. EC50 of TMZ) were analyzed using the differential expression analysis of our microarray data. A heat map was constructed by ranking these probes according to the extent of expression change in T98G and U87 (with or without TMZ treatment), and there were 8 genes represented by these 13 probes (Figure 1C). The prognostic values of the 8 genes were then verified by the Kaplan-Meier survival analysis of the clinical dataset in the TCGA GBM cohort. The unadjusted hazard ratio (HR), upper and lower 95% confidence interval (CI), and P value, which were determined by the expression status of the 8 genes, were calculated and compared. And the 8 candidate genes were ranked according to their HR (Figure 1D). Notably, FUBP1 expression was highly increased in T98G after TMZ treatment and was the top-ranked marker out of the 13 identified probes (Figure 1C). However, when examining its prognostic value, FUBP1 expression was shown to have no significance in predicting the overall survival of GBM patients (HR=1.071; 95% CI=0.887-1.294; P=0.475122) (Figure 1D). This was inconsistent with the past research whereby FUBP1 was shown to be associated with poor prognosis in glioma patients [33]. Therefore, we chose BICD1, the top-ranked gene in impacting the overall survival of GBM patients (HR=1.577; 95% CI=1.299-1.914; P=0.000004) (Figure 1D), as a candidate marker because of its high potential in developing a biomarker of GBMs, and its novelty in the study of GBMs. The differential expression of BICD1 mRNA in U87 and T98G (with or without TMZ treatment) was further confirmed by RT-PCR (Figure 1E). Additionally, the gene expression status of MGMT in U87 and T98G (with or without TMZ treatment) and its prognostic value were analyzed and presented as reference (Figure 1C, 1D).
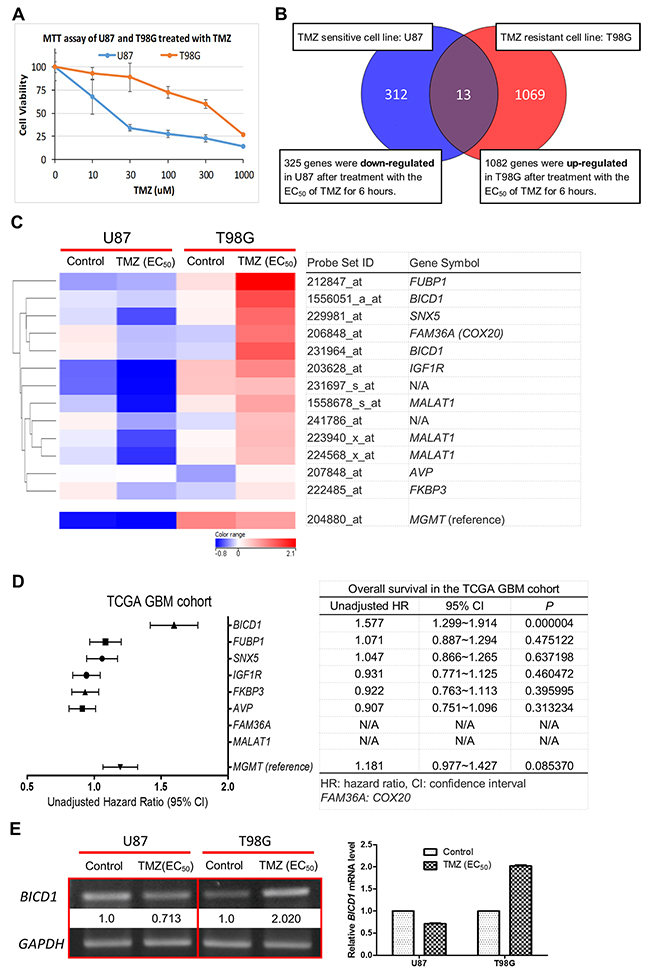
Figure 1: Identification of BICD1 expression as a potential biomarker of GBMs. (A) Results of MTT assay showed a higher cell viability in T98G than in U87 when both cell lines were treated with the alkylating agent TMZ. The EC50 of TMZ was 400μM in T98G and the EC50 of TMZ was 20μM in U87. (B) Hierarchical clustering analysis of the genomic alterations in T98G (TMZ resistant cell line) and in U87 (TMZ sensitive cell line) after treatment with their EC50 of TMZ for 6 hours. A total of 13 probes which were up-regulated in T98G and down-regulated in U87 after TMZ treatment were identified. (C) A heat map for identification of potential candidate genes was constructed according to the extent of gene expression change after TMZ treatment in T98G and U87. A total of 8 candidate genes were identified (2 probes lacked gene symbol; BICD1 was represented by 2 probes; MAMAT1 was represented by 3 probes). (D) The prognostic values of these candidate genes were verified and compared using the Kaplan-Meier survival analysis of the clinical dataset in the TCGA GBM cohort. The unadjusted hazard ratio (HR), upper and lower 95% confidence interval (CI), and P value, which were determined by the expression status of each candidate gene, were ranked and listed according to their HR. (E) RT-PCR was performed to confirm the gene expression change of BICD1 in GBM cell line after TMZ treatment in our microarray data. Results of RT-PCR showed increased BICD1 mRNA expression in T98G cells after TMZ treatment but decreased BICD1 mRNA expression in U87 cells after TMZ treatment (The percentage of brightness and contrast had been adjusted to increase the BICD1 mRNA signal. The percentage of 65% in brightness and 80% in contrast were applied in the presentation of BICD1 mRNA expression). The gene expression status of MGMT and its prognostic value are shown in (C) and (D) as reference.
BICD1 expression was significantly correlated with the WHO grade, patient age, and KPS in the TCGA GBMLGG cohort, and highly associated with the molecular subclassification of GBMs
GBMs (grade IV gliomas) had apparently higher BICD1 expression than LGGs (grades II and III gliomas) in the TCGA GBMLGG cohort (n=689) (Figure 2A). GBM patients had significantly poorer overall survival than LGG patients (P<0.000001) (Figure 2B). The expression levels of BICD1 were significantly higher in GBMs than in LGGs (***) (Figure 2C). GBMs had a significantly higher percentage of high BICD1 expression than LGGs (GBMs: 134/165 vs. LGGs: 210/524, P<0.00001) (Figure 2D) (Table 2).
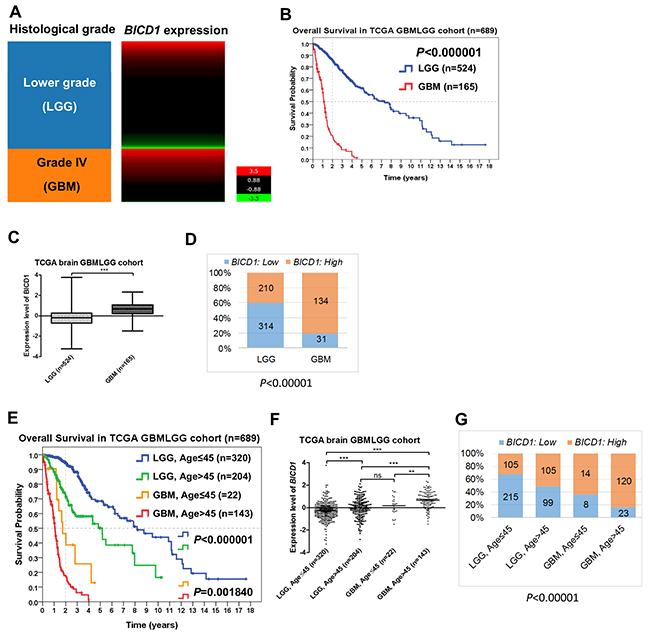
Figure 2: The expression status of BICD1 in different histological grade and age group obtained by the gene expression RNAseq (polyA+ IlluminaHiSeq) in the TCGA GBMLGG cohort (n=689). (A) Grade IV gliomas (GBMs) appeared to have higher BICD1 expression than lower grade gliomas (LGGs). (B) GBM patients had significantly poorer overall survival than LGG patients (P<0.000001). (C) The expression levels of BICD1 were significantly higher in GBMs than in LGGs (***). (D) GBMs had a significantly higher percentage of high BICD1 expression than LGGs (GBMs: 134/165 vs. LGGs: 210/524, P<0.00001). (E) LGG and GBM patients in the TCGA GBMLGG cohort were further separated by age (≤45 vs. >45). Patients with higher age (>45) had significantly poorer overall survival in both LGG and GBM patient groups. The LGG patients with age>45 had significantly poorer overall survival than those with age≤45 (P<0.000001). The GBM patients with age>45 also had significantly poorer overall survival than those with age≤45 (P=0.001840). (F) The expression levels of BICD1 were significantly higher in GBM patients with age>45, than in those with age≤45 (**). The expression levels of BICD1 were also significantly higher in LGG patients with age>45 than in those with age≤45 (***). (G) The GBM patient group with age>45 had the highest percentage of high BICD1 expression (120/143), but the LGG patient group with age≤45 had the lowest one (105/320). The percentage of high BICD1 expression was significantly correlated with the poor prognosis of glioma patients when they were grouped according to the histological grade and patient age in the TCGA GBMLGG cohort (P<0.00001).
Table 2: Correlation of BICD1 expression with the clinicopathological features of patients in the TCGA GBMLGG cohort
Clinicopathological features |
n |
BICD1 expression, n (%) |
P |
|
|---|---|---|---|---|
689 |
Low, n=345 (50.1) |
High, n=344 (49.9) |
||
Age |
<0.000001 |
|||
≤45 |
342 |
223 (65.2) |
119 (34.8) |
|
>45 |
347 |
122 (35.2) |
225 (64.8) |
|
Gender |
0.724838 |
|||
Female |
295 |
150 (50.8) |
145 (49.2) |
|
Male |
394 |
195 (49.5) |
199 (50.5) |
|
WHO Grade |
<0.000001 |
|||
LGG (Grades II+III) |
524 |
314 (59.9) |
210 (40.1) |
|
GBM (Grade IV) |
165 |
31 (18.8) |
134 (81.2) |
|
BICD1 expression was highly and significantly correlated with the WHO histological grade (P<0.000001) and patient age (P<0.000001) in the TCGA GBMLGG cohort (Table 2). LGG and GBM patients in the TCGA GBMLGG cohort were further separated by age (≤45 vs. >45). Undoubtedly, patients with higher age had significantly poorer overall survival in both LGG and GBM patient groups in the TCGA GBMLGG cohort (Figure 2E). The LGG patients with age>45 had significantly poorer overall survival than those with age≤45 (P<0.000001). The GBM patients with age>45 also had significantly poorer overall survival than those with age≤45 (P=0.001840). The expression levels of BICD1 were significantly higher in GBM patients with age>45 than in those with age≤45 (**). The expression levels of BICD1 were also significantly higher in LGG patients with age>45 than in those with age≤45 (***) (Figure 2F). The GBM patients with age>45 had the highest percentage of high BICD1 expression (120/143), and the LGG patients with age≤45 had the lowest percentage of high BICD1 expression (105/320). The percentage of high BICD1 expression was significantly correlated with poor prognosis in glioma patients when they are grouped according to the WHO grade and patient age in the TCGA GBMLGG cohort (P<0.00001) (Figure 2G).
BICD1 expression was significantly but negatively correlated with patients’ clinical performance (Karnofsky performance score, KPS) in the TCGA GBMLGG cohort (≥90 vs. <90, P<0.000001) (Table 3). However, it was not significantly correlated with the KPS (≥90 vs. <90, P=0.609816) (Supplementary Table 1), and the clinicopathological features, including patient age (<65 vs. ≥65, P=0.089722), gender (Female vs. Male, P=0.136863), MGMT expression (Low vs. High, P=0.204737), and overall survival indicator (censor vs. death, P=0.712211), in the TCGA GBM cohort (Table 4).
Table 3: Correlation of BICD1 expression with the KPS of patients in the TCGA GBMLGG cohort
Clinicopathological feature |
n |
BICD1 expression, n (%) |
P |
|
|---|---|---|---|---|
439 |
Low, n=220 (50.1) |
High, n=219 (49.9) |
||
KPS |
<0.000001 |
|||
≥90 |
226 |
147 (65) |
79 (35) |
|
<90 |
213 |
73 (34.3) |
140 (65.7) |
|
KPS: Karnofsky performance score.
Table 4: Correlation of BICD1 expression with the clinicopathological features of patients in the TCGA GBM cohort
Clinicopathological features |
n |
BICD1 expression, n (%) |
P |
|
|---|---|---|---|---|
523 |
Low, n=262 (50.1) |
High, n=261 (49.9) |
||
Age |
0.089722 |
|||
<65 |
347 |
183 (52.7) |
164 (47.3) |
|
≥65 |
176 |
79 (44.9) |
97 (55.1) |
|
Gender |
0.136863 |
|||
Female |
205 |
111 (54.1) |
94 (45.9) |
|
Male |
318 |
151 (47.5) |
167 (52.5) |
|
MGMT expression |
0.204737 |
|||
Low |
262 |
124 (47.3) |
138 (52.7) |
|
High |
261 |
138 (52.9) |
123 (47.1) |
|
Overall survival indicator |
0.712211 |
|||
0 (censor) |
89 |
43 (48.3) |
46 (51.7) |
|
1 (death) |
434 |
219 (50.5) |
215 (49.5) |
|
Molecular Subclassification |
||||
Neural |
87 |
70 (80.5) |
17 (19.5) |
<0.00001 |
Proneural |
138 |
91 (65.9) |
47 (34.1) |
|
Mesenchymal |
156 |
71 (45.5) |
85 (54.5) |
|
Classical |
142 |
30 (21.1) |
112 (78.9) |
|
The molecular classification of GBMs was defined by TCGA. They used the 840 differentially expressed gene signature to classify GBMs into 4 clinically relevant subtypes (proneural, neural, classical, and mesenchymal) [34]. The proneural subtype has better prognosis than other subtypes of GBMs (Supplementary Figure 1A) (Supplementary Table 2). The expression levels of BICD1 varied with the molecular subclassification of GBMs (Figure 3A). These subtypes of GBMs were ranked according to their adjusted HR, and mesenchymal and classical subtypes were more malignant than neural and proneural subtypes (Figure 3B). By dividing GBMs into two subgroups according to the molecular classification (proneural and neural vs. classical and mesenchymal), the difference in overall survival was not significant (P=0.078687) (Figure 3C). However, there was still a distinction in the survival curves between the more malignant subtypes (classical and mesenchymal) and the less malignant (proneural and neural) subtypes. The expression levels of BICD1 were significantly higher in classical and mesenchymal subtypes than in neural and proneural subtypes (Figure 3D). The percentage of high BICD1 expression was significantly greater in classical and mesenchymal subtypes than in neural and proneural subtypes (classical: 112/142, mesenchymal: 85/156, proneural: 47/138, neural: 17/87) (P<0.00001) (Figure 3E) (Table 4).
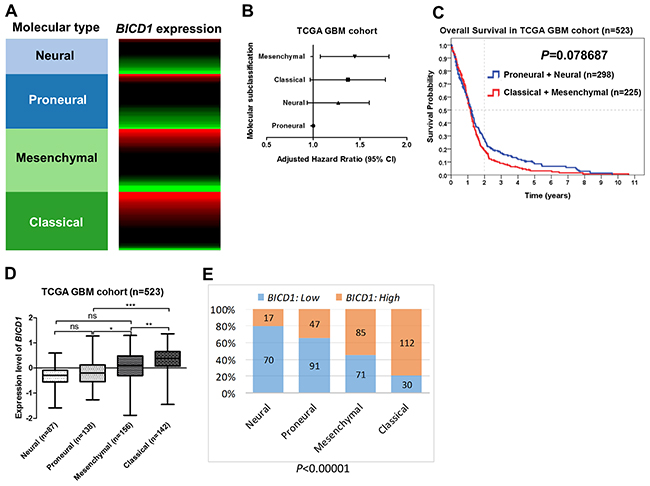
Figure 3: The expression status of BICD1 in different molecular subclassification of GBMs obtained by the gene expression array (AffyU133a) in the TCGA GBM cohort (n=523). The molecular classification of GBMs was defined by TCGA. (A) BICD1 expression varied with the molecular subclassification of GBMs, and appeared to be up-regulated in classical and mesenchymal subtypes. (B) The molecular subtypes of GBMs were ranked according to their adjusted HR, and mesenchymal and classical subtypes were shown to be more malignant than neural and proneural subtypes. (C) By dividing GBMs into two subgroups according to the molecular subtypes (proneural and neural vs. classical and mesenchymal), there was no significant difference in overall survival (P=0.078687). However, there was still a distinction in the survival curves between the more malignant subtypes (classical and mesenchymal) and the less malignant subtypes (proneural and neural). (D) The expression levels of BICD1 were significantly higher in classical and mesenchymal subtypes than in neural and proneural subtypes (***). (E) The classical and mesenchymal subtypes had a significantly greater proportion of high BICD1 expression than the neural and proneural subtypes (classical: 112/143, mesenchymal: 85/156, proneural: 47/138, neural: 17/87) (P<0.00001).
Comparisons of BICD1 and MGMT expression in predicting the overall survival of patients in various glioma datasets
In comparison of BICD1 with MGMT expression in predicting the overall survival of glioma patients by the Kaplan-Meier survival analysis, high BICD1 expression showed more significant impact (P<0.000001) than high MGMT expression (P=0.00003) on worse overall survival in the TCGA GBMLGG cohort (Figure 4A). High BICD1 expression showed significant impact (P=0.000003), while high MGMT expression did not show significance (P=0.084689) on worse overall survival in the TCGA GBM cohort (Figure 4B). High BICD1 expression also showed more significant impact (P=0.009932) than high MGMT expression (P=0.028420) on worse overall survival in the CGGA (Chinese Glioma Genome Atlas) cohort (Figure 4C).
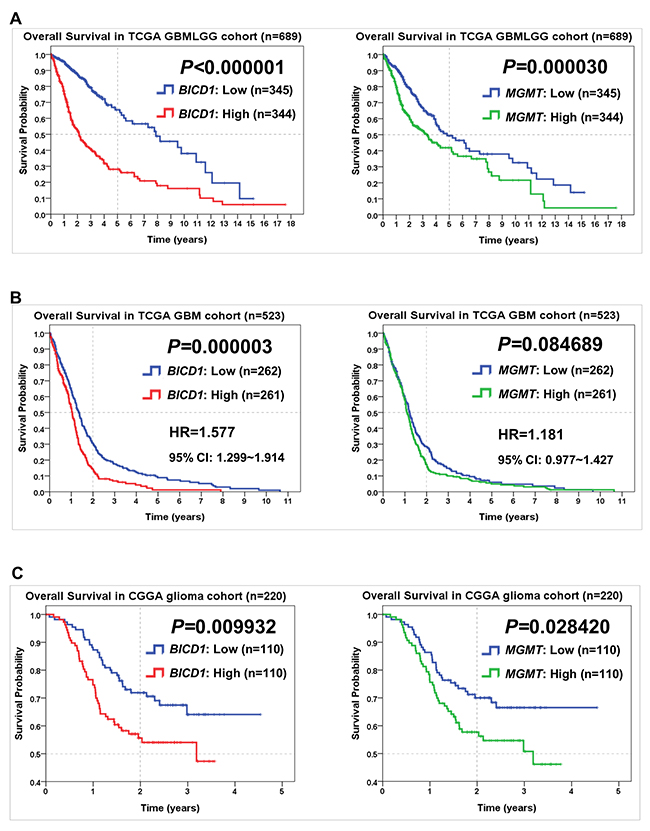
Figure 4: Comparisons of BICD1 with MGMT expression in predicting the overall survival of patients in various glioma cohorts by the Kaplan-Meier survival analysis. (A) High BICD1 expression showed more significant impact on poor overall survival (P<0.000001) than high MGMT expression (P=0.000030) in the TCGA GBMLGG cohort. (B) High BICD1 expression showed highly significant impact on poor overall survival (HR=1.577, P=0.000003), while high MGMT expression did not show significance (HR=1.181, P=0.084689) in the TCGA GBM cohort. (C) High BICD1 expression showed more significant impact on poor overall survival (P=0.009932) than high MGMT expression (P=0.028420) in the CGGA glioma cohort.
Comparisons of BICD1 and MGMT expression in predicting the time to experience a new tumor event, the time to experience tumor progression, and the time to experience tumor recurrence in the TCGA GBM dataset
Comparisons of BICD1 with MGMT expression in predicting other survival events of patients in the TCGA GBM cohort were also made by the Kaplan-Meier survival analysis. The time to experience a new tumor event was significantly shorter in patients with high BICD1 expression (P=0.000127) than in those with high MGMT expression (P=0.008955) (Figure 5A). The time to experience tumor progression was also significantly shorter in patients with high BICD1 expression (P=0.000321), while it was not significantly shorter in patients with high MGMT expression (P=0.469433) (Figure 5B). And the time to experience tumor recurrence was significantly shorter in patients with high BICD1 expression (P=0.000117) than in those with high MGMT expression (P=0.005083) (Figure 5C).
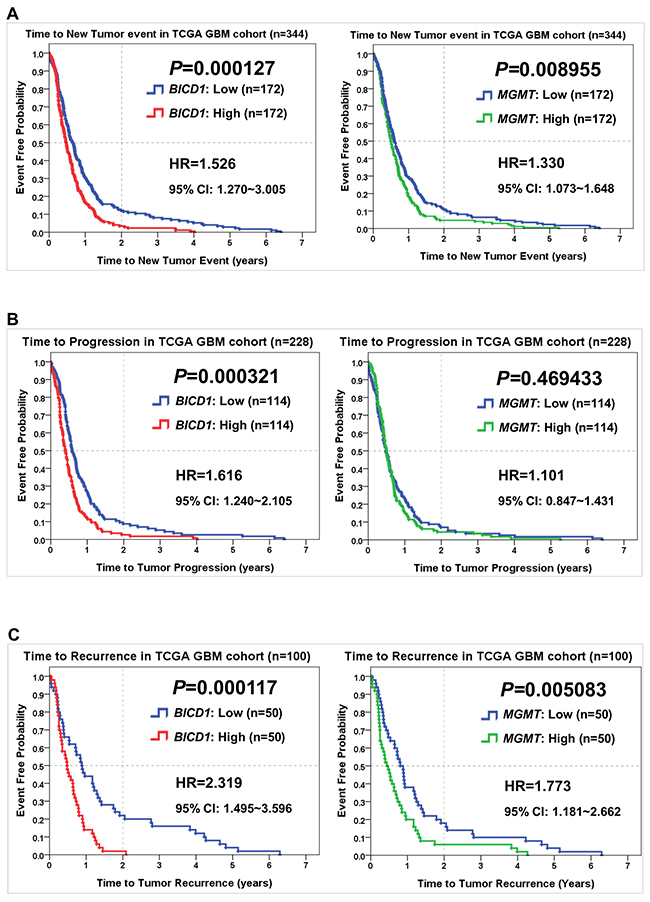
Figure 5: Comparisons of BICD1 with MGMT expression in predicting other survival events of patients in the TCGA GBM cohort by the Kaplan-Meier survival analysis. (A) GBM patients with high BICD1 expression spent a significantly shorter time to experience a new tumor event (HR=1.526, P=0.000127) than those with high MGMT expression (HR=1.330, P=0.008955). (B) GBM patients with high BICD1 expression spent a significantly shorter time to experience tumor progression (HR=1.616, P=0.000321), while those with high MGMT expression did not show significance (HR=1.101, P=0.469433). (C) GBM patients with high BICD1 expression spent a significantly shorter time to experience tumor recurrence (HR=2.319, P=0.000117) than those with high MGMT expression (HR=1.773, P=0.005083).
BICD1 expression, as well as MGMT, were both powerful predictors of the response to TMZ treatment in GBM patients. In patients who received TMZ treatment, high MGMT expression showed significantly stronger impact (P=0.002388) on poor overall survival than high BICD1 expression (P=0.005515) (Figure 6A). In patients without TMZ treatment, high BICD1 expression still showed significant impact (P=0.009929) on poor overall survival, while high MGMT expression did not show significance (P=0.341587) (Figure 6B), which was compatible with the current knowledge that MGMT is a specific biomarker for predicting the response to TMZ treatment in glioma patients [35].
Comparisons of BICD1 and MGMT expression in predicting the response to various therapies in the TCGA GBM cohort
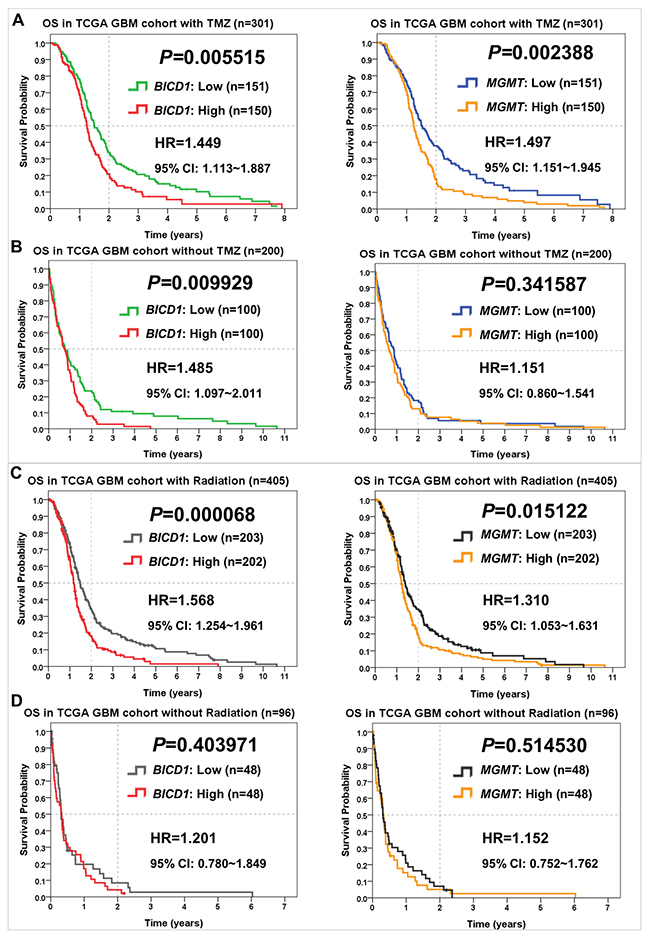
Figure 6: Comparisons of BICD1 with MGMT expression in predicting the therapeutic outcomes of TMZ and radiation therapies in the TCGA GBM cohort by the Kaplan-Meier survival analysis (OS: overall survival). (A) In GBM patients who received TMZ chemotherapy, high MGMT expression showed more significant impact on poor overall survival (HR=1.497, P=0.002388) than high BICD1 expression (HR=1.449, P=0.005515). (B) In GBM patients who did not receive TMZ chemotherapy, high BICD1 expression still showed significant impact on poor overall survival (HR=1.485, P=0.009929), while high MGMT expression did not show significance (HR=1.151, P=0.341587). (C) In GBM patients who received radiation therapy, high BICD1 expression showed more significant impact on poor overall survival (HR=1.568, P=0.000068) than high MGMT expression (HR=1.310, P=0.015122). (D) In GBM patients who did not receive radiation therapy, both BICD1 and MGMT expression did not show significant impact on overall survival, but BICD1 expression showed higher impact (HR=1.201, P=0.403971) than MGMT expression (HR=1.152, P=0.514530).
BICD1 expression was a more powerful predictor of the response to radiation therapy in GBM patients than MGMT expression. In patients who received radiation therapy, high BICD1 expression showed more significant impact (P=0.000068) on poor overall survival than high MGMT expression (P=0.015122) (Figure 6C). In patients without radiation therapy, both BICD1 and MGMT expression did not show significant impact on the overall survival of GBM patients, but BICD1 expression showed higher impact (P=0.403971) than MGMT expression (P=0.514530) (Figure 6D).
Multivariate Cox regression analyses confirmed BICD1 expression as an independent factor affecting the prognosis and response to therapies, including TMZ and radiation therapies in GBM patients
BICD1 expression was an independent factor affecting the overall survival of GBM patients (adjusted HR=1.557, P=0.000009). Its impact power was more significant than MGMT expression (adjusted HR=1.291, P=0.008976), but less significant than age (adjusted HR=2.143, P<0.000001) (Table 5).
Table 5: Univariate and multivariate Cox regression analyses of BICD1 expression and the clinicopathological factors in the overall survival of patients in the TCGA GBM cohort
Variables |
Univariate |
Multivariate |
|||||
|---|---|---|---|---|---|---|---|
HR |
95% CI |
P |
HR |
95% CI |
P |
||
BICD1 |
Low vs. High |
1.577 |
1.299-1.914 |
0.000004 |
1.557 |
1.281-1.894 |
0.000009 |
Age |
<65 vs. ≥65 |
2.139 |
1.741-2.627 |
<0.000001 |
2.143 |
1.742-2.638 |
<0.000001 |
Gender |
Female vs. Male |
1.175 |
0.967-1.429 |
0.104814 |
1.113 |
0.915-1.353 |
0.284136 |
MGMT |
Low vs. High |
1.181 |
0.977-1.427 |
0.085370 |
1.291 |
1.066-1.563 |
0.008976 |
HR: hazard ratio, CI: confidence interval.
BICD1 expression was an independent factor affecting the response to TMZ treatment in GBM patients (adjusted HR=1.576, P=0.000974). However, age (adjusted HR=1.777, P=0.000211) and MGMT expression (adjusted HR=1.647, P=0.000270) showed more significance than BICD1 expression in affecting the response to TMZ treatment (Table 6).
Table 6: Univariate and multivariate Cox regression analyses of BICD1 expression and the clinicopathological factors in the overall survival of patients with TMZ treatment in the TCGA GBM cohort
Variables |
Univariate |
Multivariate |
|||||
|---|---|---|---|---|---|---|---|
HR |
95% CI |
P |
HR |
95% CI |
P |
||
BICD1 |
Low vs. High |
1.449 |
1.113-1.887 |
0.005826 |
1.576 |
1.203-2.065 |
0.000974 |
Age |
<65 vs. ≥65 |
1.746 |
1.290-2.364 |
0.000309 |
1.777 |
1.311-2.408 |
0.000211 |
Gender |
Female vs. Male |
1.247 |
0.949-1.639 |
0.113591 |
1.235 |
0.938-1.626 |
0.133230 |
MGMT |
Low vs. High |
1.497 |
1.151-1.945 |
0.002575 |
1.647 |
1.259-2.154 |
0.000270 |
HR: hazard ratio, CI: confidence interval.
BICD1 expression was also an independent factor affecting the overall survival of GBM patients who did not receive TMZ treatment (adjusted HR=1.416, P=0.027711). Its impact power was more significant than MGMT expression (adjusted HR=1.110, P=0.487235), but less significant than age (adjusted HR=2.250, P<0.000001) (Table 7).
Table 7: Univariate and multivariate Cox regression analyses of BICD1 expression and the clinicopathological factors in the overall survival of patients without TMZ treatment in the TCGA GBM cohort
Variables |
Univariate |
Multivariate |
|||||
|---|---|---|---|---|---|---|---|
HR |
95% CI |
P |
HR |
95% CI |
P |
||
BICD1 |
Low vs. High |
1.485 |
1.097-2.011 |
0.010485 |
1.416 |
1.039-1.929 |
0.027711 |
Age |
<65 vs. ≥65 |
2.286 |
1.682-3.107 |
<0.000001 |
2.250 |
1.655-3.059 |
<0.000001 |
Gender |
Female vs. Male |
1.181 |
0.878-1.589 |
0.271452 |
1.101 |
0.815-1.487 |
0.532466 |
MGMT |
Low vs. High |
1.151 |
0.860-1.541 |
0.342714 |
1.110 |
0.827-1.488 |
0.487235 |
HR: hazard ratio, CI: confidence interval.
BICD1 expression was an independent factor affecting the response to radiation therapy in GBM patients (adjusted HR=1.601, P=0.000044). Age (adjusted HR=1.676, P=0.000072) and MGMT expression (adjusted HR=1.414, P=0.002191) were also significant factors, but not as significant as BICD1 expression (Table 8).
Table 8: Univariate and multivariate Cox regression analyses of BICD1 expression and the clinicopathological factors in the overall survival of patients with radiation therapy in the TCGA GBM cohort
Variables |
Univariate |
Multivariate |
|||||
|---|---|---|---|---|---|---|---|
HR |
95% CI |
P |
HR |
95% CI |
P |
||
BICD1 |
Low vs. High |
1.568 |
1.254-1.961 |
0.000078 |
1.601 |
1.277-2.007 |
0.000044 |
Age |
<65 vs. ≥65 |
1.657 |
1.287-2.134 |
0.000091 |
1.676 |
1.299-2.162 |
0.000072 |
Gender |
Female vs. Male |
1.215 |
0.968-1.524 |
0.093019 |
1.138 |
0.906-1.430 |
0.265744 |
MGMT |
Low vs. High |
1.310 |
1.053-1.631 |
0.015538 |
1.414 |
1.133-1.765 |
0.002191 |
HR: hazard ratio, CI: confidence interval.
BICD1 (adjusted HR=1.280, P=0.267624) and MGMT expression (adjusted HR=1.115, P=0.620605) were not independent factors affecting the overall survival of GBM patients who did not receive radiation therapy, while age was still a significant factor (adjusted HR=2.126, P=0.001580), which suggested that BICD1 and MGMT expression may be specific biomarkers for predicting the response to radiation therapy (Table 9).
Table 9: Univariate and multivariate Cox regression analyses of BICD1 expression and the clinicopathological factors in the overall survival of patients without radiation therapy in the TCGA GBM cohort
Variables |
Univariate |
Multivariate |
|||||
|---|---|---|---|---|---|---|---|
HR |
95% CI |
P |
HR |
95% CI |
P |
||
BICD1 |
Low vs. High |
1.201 |
0.780-1.849 |
0.405628 |
1.280 |
0.827-1.980 |
0.267624 |
Age |
<65 vs. ≥65 |
2.044 |
1.283-3.256 |
0.001638 |
2.126 |
1.332-3.395 |
0.001580 |
Gender |
Female vs. Male |
1.274 |
0.825-1.969 |
0.274642 |
1.363 |
0.879-2.112 |
0.166421 |
MGMT |
Low vs. High |
1.152 |
0.752-1.762 |
0.515767 |
1.115 |
0.725-1.715 |
0.620605 |
HR: hazard ratio, CI: confidence interval.
Risk stratification of GBM patients according to the combination of age and BICD1 expression may provide more accurate outcome assessment
According to the combination of patient age with BICD1 expression, patients in the TCGA GBM cohort (n=523) were stratified into more distinct risk groups for outcome assessment by the Kaplan-Meier survival analysis. The median survival, 2-year survival rate, and HR of GBM patients in each subgroup were listed. The difference in the overall survival of patients between age<65 and ≥65 was highly significant (P<0.000001). Patients with higher age (≥65) had poorer outcomes (adjusted HR=2.143) than those with lower age (<65) (Figure 7A). The median survival and 2-year survival rate in patients with age<65 were 1.351 years and 28.7%, and in patients with age≥65 were 0.764 years and 10.2%, respectively. In addition to age, BICD1 expression was shown to have a high HR (adjusted HR=1.557) in patients with GBMs (Table 5).
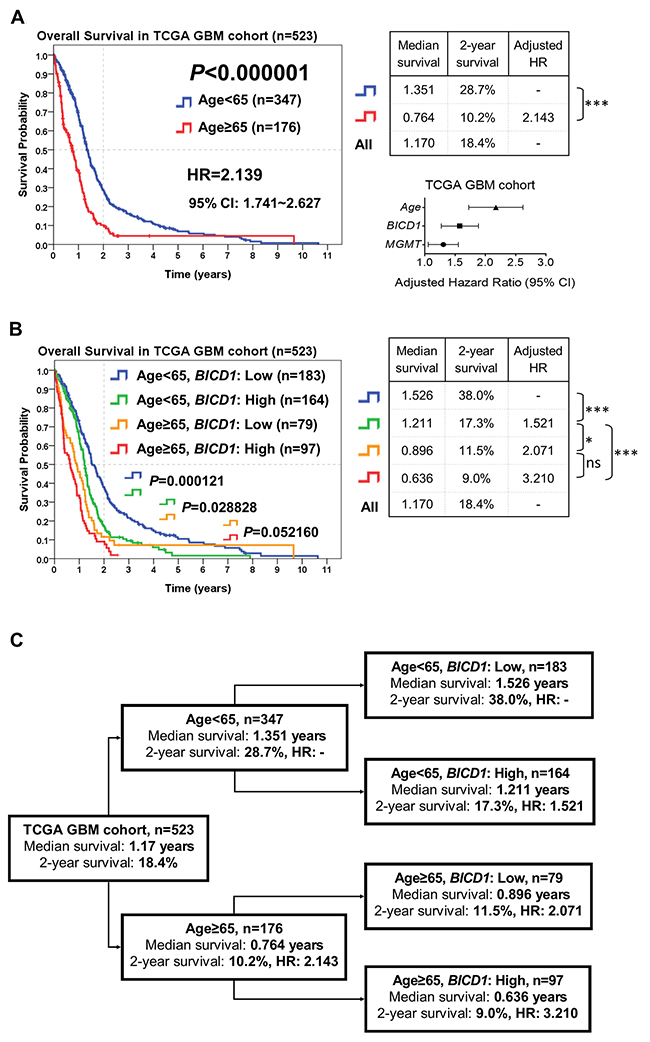
Figure 7: The combined utilization of age and BICD1 expression may provide more accurate outcome assessment of GBM patients by risk stratification of patients in the TCGA GBM cohort (n=523). The median survival, 2-year survival rate and adjusted HR were obtained by the Kaplan-Meier survival analysis. (A) Age was highly significant in determining the outcomes of GBM patients (P<0.000001). Patients with higher age (≥65) had poorer outcomes (adjusted HR=2.143) than those with lower age (<65). The difference in the overall survival of patients between age<65 and ≥65 was highly significant. The median survival and 2-year survival rate in patients with age<65 were 1.351 years and 28.7%, and in patients with age≥65 were 0.764 years and 10.2%, respectively. In addition to age, BICD1 expression was found to have a high adjusted HR (adjusted HR=1.557) in GBM patients (Table 5). (B) By adding BICD1 expression as a cofactor with age, patients with age<65 could be further divided into higher and lower risk groups (median survival: 1.211 vs. 1.526 years, 2-year survival rate: 17.3% vs. 38.0%) (P=0.000121). Patients with age≥65 could also be divided into higher and lower risk groups (median survival: 0.636 vs. 0.896 years, 2-year survival rate: 9.0% vs. 11.5%) (P=0.052160). The patient group with age≥65 and high BICD1 expression had the highest HR and the poorest prognosis (adjusted HR=3.210, median survival: 0.636 years, and 2-year survival rate: 9.0%). (C) A flowchart to represent the application of our results in Figure 7B. This may provide more accurate prediction of patients’ median survival, 2-year survival rate, and adjusted HR when age and BICD1 expression were combinedly utilized for risk stratification of GBM patients.
By adding BICD1 expression as a cofactor with age, patients with age<65 could be further divided into higher and lower risk groups (median survival: 1.211 vs. 1.526 years, 2-year survival rate: 17.3% vs. 38.0%) (P=0.000121). Patients with age≥65 could also be divided into higher and lower risk groups (median survival: 0.636 vs. 0.896 years, 2-year survival rate: 9.0% vs. 11.5%) (P=0.052160) (Figure 7B). The patient group with age≥65 and high BICD1 expression had the highest HR and the poorest prognosis (adjusted HR=3.210, median survival: 0.636 years, and 2-year survival rate: 9.0%). And the patient group with age<65 and low BICD1 expression had the best prognosis (median survival: 1.526 years, and 2-year survival rate: 38.0%).
The flowchart in Figure 7C represents the application of our results in Figure 7B. This may provide more accurate prediction of patients’ prognosis when age and BICD1 expression were combinedly utilized for risk stratification of GBM patient (Figure 7C).
Risk stratification of GBM patients who received TMZ chemotherapy according to the combination of age, MGMT, and BICD1 expression may provide better prediction of the response to TMZ
According to the combination of age, MGMT, and BICD1 expression, patients who received TMZ chemotherapy in the TCGA GBM cohort (n=301) were stratified into distinct risk groups for outcome assessment by the Kaplan-Meier survival analysis. The median survival, 2-year survival rate, and HR of patients in each subgroup were listed. Age was highly significant in determining the outcomes of GBM patients who received TMZ chemotherapy (P=0.000255). Patients with higher age (age≥65) had poorer outcomes (adjusted HR=1.777) than those with lower age (<65) (Figure 8A). The median survival and 2-year survival rate in patients with age<65 were 1.482 years and 31.8%, and in patients with age≥65 were 1.112 years and 17.1%, respectively. In addition to age, MGMT and BICD1 expression were shown to have high HRs (MGMT: adjusted HR=1.647, BICD1: adjusted HR=1.576) in GBM patients with TMZ treatment (Table 6).
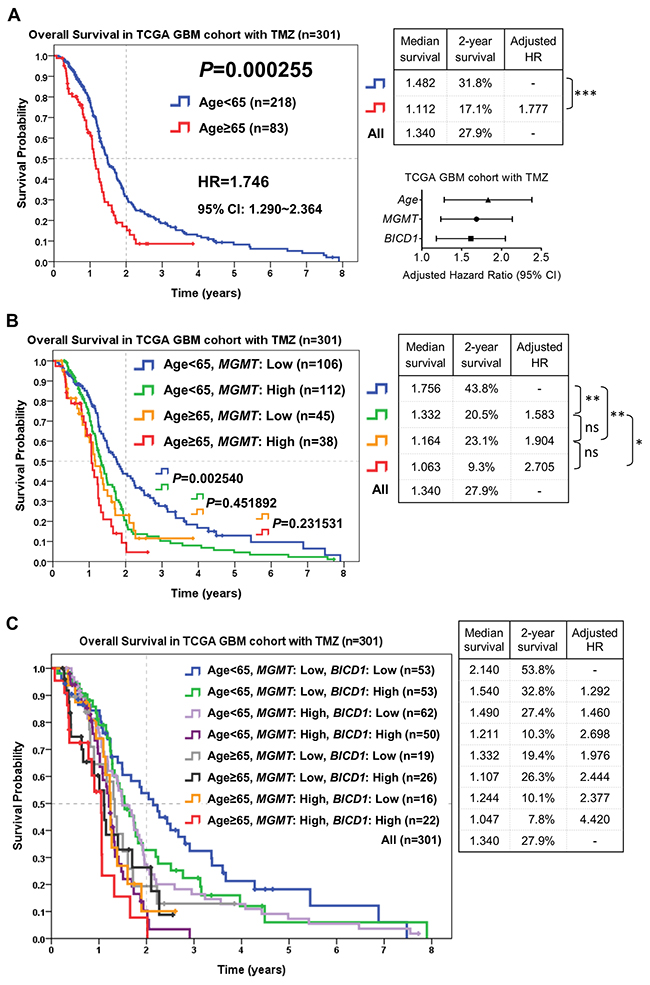
Figure 8: The combined utilization of age, MGMT and BICD1 expression may provide better prediction of patients’ response to TMZ chemotherapy by risk stratification of GBM patients who received TMZ chemotherapy in the TCGA GBM cohort (n=301). The median survival, 2-year survival rate and adjusted HR were obtained by the Kaplan-Meier survival analysis. (A) Age was highly significant in determining the outcomes of patients who received TMZ chemotherapy (P=0.000255). Patients with higher age (≥65) had poorer outcomes (adjusted HR=1.777) than those with lower age (<65). In addition to age, MGMT and BICD1 expression were found to have high adjusted HRs (MGMT: adjusted HR=1.647, BICD1: adjusted HR=1.576) in GBM patients with TMZ treatment (Table 6). (B) The difference in the overall survival of patients with age<65 between low and high MGMT expression was significant (P=0.002540). The patient group with age<65 and low MGMT expression had the best prognosis (median survival: 1.756 years, and 2-year survival rate: 43.8%). The patient group with age≥65 and high MGMT expression had the highest HR and the poorest prognosis (adjusted HR=2.705, median survival: 1.063 years, and 2-year survival rate: 9.3%). (C) By adding BICD1 expression as a cofactor with age and MGMT expression, the patient group with age<65, low MGMT and low BICD1 expression had the best prognosis (median survival: 2.140 years, and 2-year survival rate: 53.8%). The patient group with age≥65, high MGMT and high BICD1 expression had the highest HR and the poorest prognosis (adjusted HR=4.420, median survival: 1.047 years, and 2-year survival rate: 7.8%).
By adding MGMT expression as a cofactor with age, patients with age<65 could be further divided into higher and lower risk groups (median survival: 1.332 vs. 1.756 years, 2-year survival rate: 20.5% vs. 43.8%) (P=0.002540). Patients with age≥65 could also be divided into higher and lower risk groups (median survival: 1.063 vs. 1.164 years, 2-year survival rate: 9.3% vs. 23.1%) (P=0.231531) (Figure 8B). The patient group with age≥65 and high MGMT expression had the highest HR and the poorest prognosis (adjusted HR=2.705, median survival: 1.063 years, and 2-year survival rate: 9.3%). And the patient group with age<65 and low MGMT expression had the best prognosis (median survival: 1.756 years, and 2-year survival rate: 43.8%).
By adding BICD1 expression as a cofactor with age and MGMT expression, the patient group with age<65 and low MGMT expression could be further divided into higher and lower risk groups (median survival: 1.540 vs. 2.140 years, 2-year survival rate: 32.8% vs. 53.8%), the patient group with age<65 and high MGMT expression could be further divided into higher and lower risk groups (median survival: 1.211 vs. 1.490 years, 2-year survival rate: 10.3% vs. 27.4%), the patient group with age≥65 and low MGMT expression could be further divided into higher and lower risk groups (median survival: 1.107 vs. 1.332 years, 2-year survival rate: 26.3% vs. 19.4%), and the patient group with age≥65 and high MGMT expression could be further divided into higher and lower risk groups (median survival: 1.047 vs. 1.244 years, 2-year survival rate: 7.8% vs. 10.1%) (Figure 8C). The patient group with age≥65, high MGMT and high BICD1 expression had the highest HR and the poorest prognosis (adjusted HR=4.420, median survival: 1.047 years, and 2-year survival rate: 7.8%). The patient group with age<65, low MGMT and low BICD1 expression had the best prognosis (median survival: 2.140 years, and 2-year survival rate: 53.8%).
The flowchart in Figure 9 represents the application of our results in Figure 8C. This may provide better prediction of GBM patients’ response to TMZ chemotherapy when age, MGMT and BICD1 expression were combinedly utilized for risk stratification of GBM patients who received TMZ chemotherapy. First, GBM patients with TMZ chemotherapy were separated according to whether they had low expression of either BICD1 or MGMT or both, and high expression of both MGMT and BICD1. Second, the patient group with high expression of both MGMT and BICD1 could be further stratified into 2 distinct risk groups according to age. Third, the patient group with low expression of either BICD1 or MGMT or both were also stratified into 2 distinct subgroups according to age, and in each age group (age<65 or age≥65), patients were further stratified into 3 distinct risk groups according to MGMT and BICD1 expression (Figure 9).
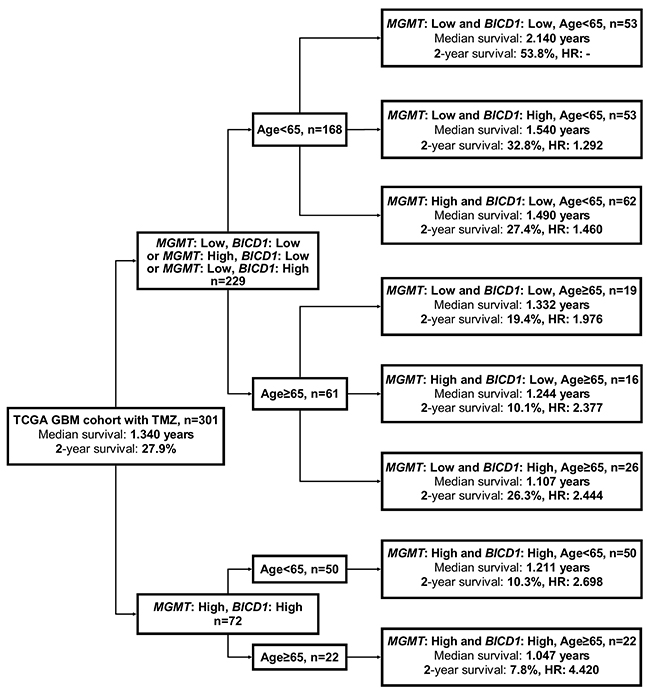
Figure 9: A flowchart to represent the application of our results in Figure 8C. This may provide better prediction of patients’ median survival, 2-year survival rate, and adjusted HR when age, MGMT and BICD1 expression were combinedly utilized for risk stratification of GBM patients who received TMZ chemotherapy. First, GBM patients who received TMZ chemotherapy were separated into 2 subgroups according to whether they had low expression of either BICD1 or MGMT or both, and high expression of both MGMT and BICD1. The patient group with high expression of both genes could be further stratified into 2 distinct risk groups according to age. The patient group with low expression of either BICD1 or MGMT or both, were further stratified into 2 subgroups according to age, and in each age group (age<65 and age≥65), patients were further stratified into 3 distinct risk groups according to their MGMT and BICD1 expression status.
Risk stratification of GBM patients who received radiation therapy according to the combination of age and BICD1 expression may provide better prediction of the response to radiation
According to the combination of age with BICD1 expression, patients who received radiation therapy in the TCGA GBM cohort (n=405) were stratified into distinct risk groups for outcome assessment by the Kaplan-Meier survival analysis. Age was highly significant in determining the outcomes of GBM patients who received radiation therapy (P=0.000076), and patients with higher age (≥65) had poorer outcomes (adjusted HR=1.676) than those with lower age (<65) (Figure 10A). The median survival and 2-year survival rate in patients with age<65 were 1.411 years and 30.6%, and in patients with age≥65 were 1.058 years and 16.5%, respectively. In addition to age, BICD1 expression was found to have a high HR (adjusted HR=1.601) in GBM patients with radiation therapy (Table 8).
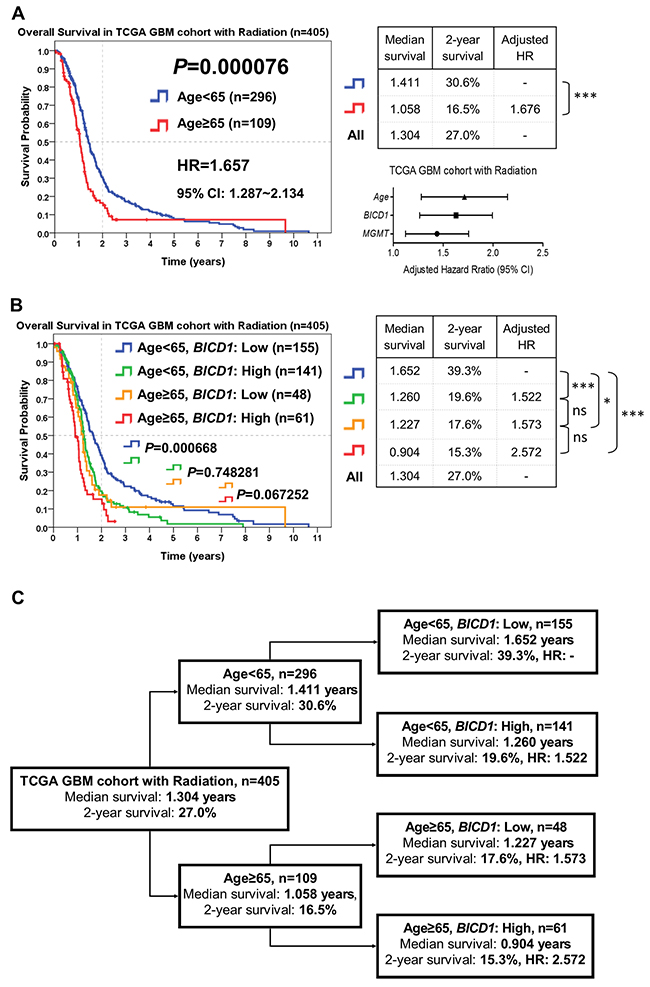
Figure 10: The combined utilization of age and BICD1 expression may provide better prediction of patients’ response to radiation therapy by risk stratification of GBM patients who received radiation therapy in the TCGA GBM cohort (n=405). The median survival, 2-year survival rate, and adjusted HR were obtained by the Kaplan-Meier survival analysis. (A) Age was highly significant in determining the outcomes of patients who received radiation therapy (P=0.000076). Patients with higher age (≥65) had poorer outcomes (adjusted HR=1.676) than those with lower age (<65). In addition to age, BICD1 expression was found to have a high adjusted HR (adjusted HR=1.601) in GBM patients with radiation therapy (Table 8). (B) The difference in the overall survival of patients with age<65 between low and high BICD1 expression was highly significant (P=0.000668). By adding BICD1 expression as a cofactor with age, patients with age<65 could be divided into higher and lower risk groups (median survival: 1.260 vs. 1.652 years, 2-year survival rate: 19.6% vs. 39.3%) (P=0.000668). Patients with age≥65 could also be divided into higher and lower risk groups (median survival: 0.904 vs. 1.227 years, 2-year survival rate: 15.3% to 17.6%) (P=0.067252). The patient group with age≥65 and high BICD1 expression had the highest HR and the poorest prognosis (adjusted HR=2.572, median survival: 0.904 years, and 2-year survival rate: 15.3%). And the patient group with age<65 and low BICD1 expression had the best prognosis (median survival: 1.652 years, and 2-year survival rate: 39.3%). (C) A flowchart to represent the application of our results in Figure 10B. This may provide better prediction of patients’ median survival, 2-year survival rate, and adjusted HR when age and BICD1 expression were combinedly utilized for risk stratification of GBM patients who received radiation therapy.
By adding BICD1 expression as a cofactor with age, patients with age<65 could be divided into higher and lower risk groups (median survival: 1.260 vs. 1.652 years, 2-year survival rate: 19.6% vs. 39.3%) (P=0.000668). Patients with age≥65 could also be divided into higher and lower risk groups (median survival: 0.904 vs. 1.227 years, 2-year survival rate: 15.3% to 17.6%) (P=0.067252) (Figure 10B). The patient group with age≥65 and high BICD1 expression had the highest HR and the poorest prognosis (adjusted HR=2.572, median survival: 0.904 years, and 2-year survival rate: 15.3%). And the patient group with age<65 and low BICD1 expression had the best prognosis (median survival: 1.652 years, and 2-year survival rate: 39.3%).
The flowchart in Figure 10C represents the application of our results in Figure 10B. This may provide better prediction of GBM patients’ response to radiation therapy when age and BICD1 expression were combinedly utilized for risk stratification of GBM patients who received radiation therapy (Figure 10C).
The possible mechanisms of BICD1-associated survival or therapeutic resistance in GBM cells
The possible mechanisms of BICD1-associated survival or therapeutic resistance in GBM cells were proposed according to our observations. BICD1 expression was positively correlated with the EMT spectrum and IDH1 expression, but appeared to be negatively correlated with MGMT expression in the TCGA GBM cohort (n=523) (Figure 11A). The expression levels of BICD1 were significantly and positively correlated with the EMT spectrum in the TCGA GBM cohort (P<0.000001, Pearson’s correlation coefficient=0.503). Patients with high BICD1 expression had a significantly higher percentage of high EMT spectrum than those with low BICD1 expression (High BICD1 expression: 168/261 vs. Low BICD1 expression: 94/262) (P<0.00001) (Figure 11B). The expression levels of BICD1 and IDH1 were also significantly and positively correlated in the TCGA GBM cohort (P<0.000001, Pearson’s correlation coefficient=0.263). Patients with high BICD1 expression had a significantly higher percentage of high IDH1 expression than those with low BICD1 expression (High BICD1 expression: 154/261 vs. Low BICD1 expression: 107/262) (P=0.000033). Patients with high BICD1 expression had a higher percentage of IDH1 wild-type than those with low BICD1 expression, however, which was not statistically significant (High BICD1 expression: 120/124 vs. Low BICD1 expression: 114/124) (P=0.098711) (Figure 11C). The expression levels of BICD1 and MGMT were significantly but negatively correlated in the TCGA GBM cohort (P=0.000002, Pearson’s correlation coefficient=-0.206). Patients with high BICD1 expression had a lower percentage of high MGMT expression than those with low BICD1 expression, however, which was not statistically significant (High BICD1 expression: 123/261 vs. Low BICD1 expression: 138/262) (P=0.204737) (Figure 11D). Our results suggested a significant correlation between BICD1 expression and EMT.
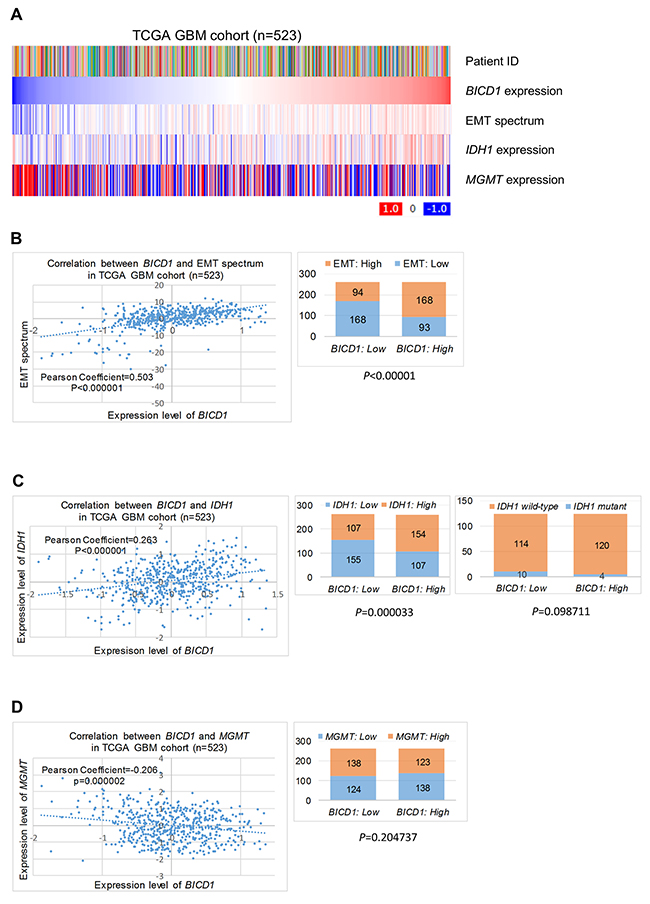
Figure 11: The possible mechanisms of BICD1-associated survival or therapeutic resistance in GBMs. (A) BICD1 expression was positively correlated with the EMT spectrum and IDH1 expression, but appeared to be negatively correlated with MGMT expression in the TCGA GBM cohort (n=523). (B) The expression levels of BICD1 were positively and significantly correlated with the EMT spectrum (P<0.000001, Pearson’s correlation coefficient=0.503). Patients with high BICD1 expression had a significantly greater proportion of high EMT spectrum than those with low BICD1 expression (High BICD1 expression: 168/261 vs. Low BICD1 expression: 94/262) (P<0.00001). (C) The expression levels of BICD1 and IDH1 were positively and significantly correlated in the TCGA GBM cohort (P<0.000001, Pearson’s correlation coefficient=0.263). Patients with high BICD1 expression had a significantly greater proportion of high IDH1 expression than those with low BICD1 expression (High BICD1 expression: 154/261 vs. Low BICD1 expression: 107/262) (P=0.000033). Patients with high BICD1 expression had a greater proportion of IDH1 wild-type than those with low BICD1 expression, which was not statistically significant (High BICD1 expression: 120/124 vs. Low BICD1 expression: 114/124) (P=0.098711). (D) The expression levels of BICD1 and MGMT were significantly and negatively correlated in the TCGA GBM cohort (P=0.000002, Pearson’s correlation coefficient=-0.206). Patients with high BICD1 expression had a smaller proportion of high MGMT expression than those with low BICD1 expression, which was not statistically significant (High BICD1 expression: 123/261 vs. Low BICD1 expression: 138/262) (P=0.204737).
The correlations between expression of BICD1 and EMT markers in the TCGA GBM cohort (n=523) were further analyzed. BICD1 expression was positively correlated with the EMT spectrum and expression of the mesenchymal markers of CDH2, ZEB2, ZEB1, VIM, FN1, TWIST1 and SNAI2, but negatively correlated with expression of the epithelial markers of CDH1, MUC1, TJP3, CLDN7 and CLDN4 (Figure 12A). The expression levels of BICD1 were significantly and positively correlated with CDH2 expression (P<0.000001, Pearson’s correlation coefficient=0.711) (Figure 12B), VIM expression (P<0.000001, Pearson’s correlation coefficient=0.523) (Figure 12C), ZEB1 expression (P<0.000001, Pearson’s correlation coefficient=0.442) (Figure 12D), and ZEB2 expression (P<0.000001, Pearson’s correlation coefficient=0.344) (Figure 12E), but negatively correlated with CLDN4 expression (P<0.000001, Pearson’s correlation coefficient=-0.431) (Figure 12F), CLDN7 expression (P<0.000001, Pearson’s correlation coefficient=-0.391) (Figure 12G), and CDH1 expression (P=0.000001, Pearson’s correlation coefficient=-0.208) (Figure 12H). These results further proved the strong correlation between BICD1 expression and EMT.
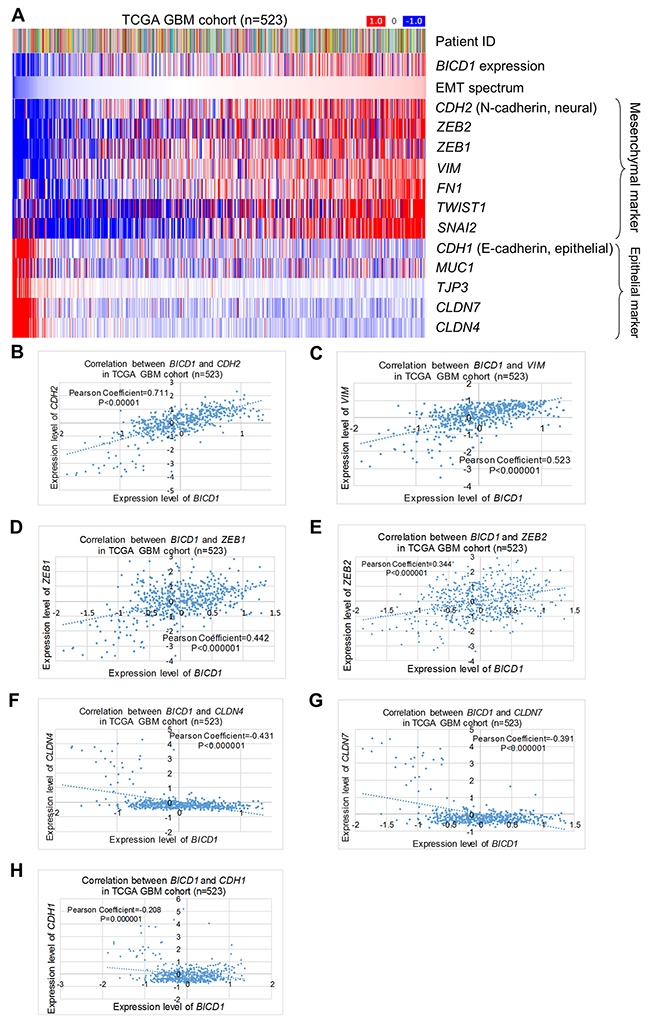
Figure 12: Correlations of BICD1 with EMT-related markers in the TCGA GBM cohort (n=523). (A) BICD1 expression was positively correlated with the EMT spectrum and expression of the mesenchymal markers of CDH2, ZEB2, ZEB1, VIM, FN1, TWIST1, and SNAI2, but negatively correlated with expression of the epithelial markers of CDH1, MUC1, TJP3, CLDN7 and CLDN4. (B) The expression levels of BICD1 were significantly and positively correlated with CDH2 expression (P<0.000001, Pearson’s correlation coefficient=0.711). (C) The expression levels of BICD1 were significantly and positively correlated with VIM expression (P<0.000001, Pearson’s correlation coefficient=0.523). (D) The expression levels of BICD1 were significantly and positively correlated with ZEB2 expression (P<0.000001, Pearson’s correlation coefficient=0.344). (E) The expression levels of BICD1 were significantly and positively correlated with ZEB1 expression (P<0.000001, Pearson’s correlation coefficient=0.442). (F) The expression levels of BICD1 were significantly and negatively correlated with CLDN7 expression (P<0.000001, Pearson’s correlation coefficient=-0.391). (G) The expression levels of BICD1 were significantly and negatively correlated with CLDN4 expression (P<0.000001, Pearson’s correlation coefficient=-0.431). (H) The expression levels of BICD1 were significantly and negatively correlated with CDH1 expression (P=0.000001, Pearson’s correlation coefficient=-0.208).
The diagram in Figure 13 illustrated a proposed mechanism of BICD1-associated survival or therapeutic resistance in GBM cells when treated with DNA-damaging agents (e.g., TMZ and radiation). On the left, the GBM cells with intrinsically high BICD1 expression may have primary resistance to DNA damage through EMT. On the right, the GBM cells with originally low BICD1 expression, after exposure to a DNA-damaging agent, may experience up-regulation of BICD1 and thus, acquire adaptive resistance to DNA damage via EMT.
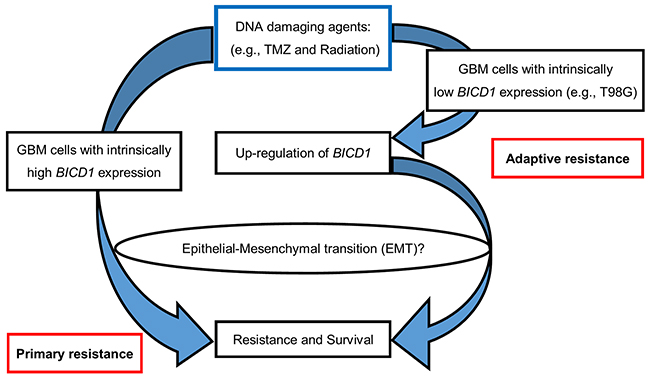
Figure 13: A proposed mechanism of BICD1-associated survival or therapeutic resistance in response to DNA-damaging agents (e.g., TMZ and radiation) in GBM cells. On the left, the GBM cells with intrinsically high BICD1 expression may have primary resistance to DNA damage through EMT. On the right, the GBM cells with originally low BICD1 expression, after exposure to a DNA-damaging agent, may experience up-regulation of BICD1, and thus, acquire adaptive resistance to DNA damage via EMT.
DISCUSSION
This is the first study to identify TMZ-related genomic markers using a cell-based microarray model. In our study, we identified BICD1 gene expression as a TMZ-related genomic marker, which also showed high significance in prognosis and the response to therapies, including TMZ and radiation therapies, in GBM patients. In our results, high BICD1 expression was significantly correlated with poor overall survival in GBM patients. Patients with high BICD1 expression spent a significantly shorter time to experience a new tumor event, tumor progress, and tumor recurrence than those with low BICD1 expression. The most impressive finding in this study is that BICD1, as well as MGMT expression, both showed high significance in predicting the outcomes of GBM patients with TMZ or radiation therapies. Finally, we confirmed BICD1 expression as an independent factor affecting the prognosis and therapeutic response of TMZ and radiation in GBM patients. According to the combination of patient age, MGMT, and BICD1 expression, GBM patients who received TMZ treatment could be further separated into more distinct risk groups, which may provide more predictive information of patients’ outcomes, and help neuro-oncologists make better individualized therapeutic decisions, and develop personalized therapy or precision medicine for GBM patients in the future.
Notably, in our microarray result, the most differentially expressed gene in T98G after TMZ treatment was the Far Upstream Element-Binding Protein 1 (FUBP1), which suggested that FUBP1 may play a certain role in response to TMZ treatment in GBM cells. FUBP1 has been reported as a biomarker for nasopharyngeal carcinoma gliomas [36] and clear cell renal cell carcinoma [37], and previously described as a transcriptional regulator of the important proto-oncogene c-Myc[38], as well as a potential therapeutic target. In addition, FUBP1 has been shown to be associated with poor prognosis in glioma patients [33], and mutations in CIC and FUBP1 have been reported to contribute to human oligodendroglioma [39]. Although FUBP1 had clinical importance in various cancers, and was shown to be the top-ranked TMZ-related gene in our microarray data. When examining its prognostic value in the TCGA GBM cohort, FUBP1 expression was found to have no significance in predicting the overall survival of GBM patients, which suggested FUBP1 may not be an ideal biomarker for the prognosis of GBM patients.
IDH1 mutation is a key molecular marker in WHO grades II and III gliomas [17]. It encodes the enzymes involved in cancer metabolism [40], and was initially described in diffuse glioma in 2008 [41]. IDH1 mutations were present commonly in 70% of lower grade gliomas and secondary GBMs. However, they occurred at relatively lower frequency (5%) in primary GBMs [42] and in other malignancy [43, 44]. This finding was similar to our observation that there was only a small percentage of IDH1 mutant in proneural subtype (13/60), neural subtype (1/43), and in all GBMs (14/248) (Supplementary Figure 1B). Therefore, this suggested that the IDH1 mutant status may not be an ideal biomarker for predicting the survival of GBM patients due to the already existing high percentage of IDH1 wild-type within primary GBMs, although the IDH1 mutant status has been the most well-known prognostic and predicting biomarker for patients with lower grade gliomas. In our result, we also found BICD1 expression was significantly correlated with IDH1 expression, instead of the IDH1 mutant status (Figure 11C), which suggested an emerging role of BICD1 in IDH1-mediated cancer metabolism. In addition, the expression levels and expression status of IDH1 were significantly higher in the classical and mesenchymal subtypes than in the proneural and neural subtypes of GBMs (Supplementary Figure 1C, 1D). This trend was similar with that in BICD1 (Figure 3D, 3E).
MGMT is well-known for its repairing role in DNA damage induced by the alkylating agent TMZ, and has been identified as a powerful predicting biomarker for TMZ resistance. MGMT expression instead of MGMT promoter methylation was selected as a marker in this study because a straightforward correlation could be established between two different gene expressions. Another reason was a standard protocol to quantify the promoter methylation of MGMT is still lacking. Our analyses confirmed the well-known characteristic of MGMT expression in predicting patients’ response to TMZ treatment. Interestingly, we also found MGMT expression could significantly predict the response to radiation therapy, but not the overall survival of GBM patients.
Molecular classification of tumors is essential for developing personalized therapies. Since TMZ resistance is not mediated by only a single molecular event, but by multiple ones, using a multi-gene signature to predict the survival and therapeutic outcomes in cancer patients has become increasingly common recently. In this study, we stratified GBM patients into further distinct risk groups according to the combination of patient age, MGMT and BICD1 expression to provide better prediction of patient’s prognosis and response to therapies, including TMZ and radiation. Although MGMT is a well-known biomarker for TMZ resistance in GBMs, combining BICD1 with MGMT expression for risk stratification of GBM patients who received TMZ chemotherapy, may offer more therapeutic information for outcome assessment than MGMT alone.
Many cellular responses have been proposed to cause chemotherapeutic resistance in cancer cells, including apoptosis inhibition, DNA damage repair, drug target alteration, the epithelial-mesenchymal transition (EMT), drug efflux, and drug inactivation [45]. EMT is a process that controls the progressive loss of epithelial characteristics and the gain of mesenchymal features, the maintenance of cancer stemness, and the acquisition of chemoresistance [46]. The EMT spectrum, derived from cancer-specific transcriptomic EMT signatures for various cancers, is a method for universal scoring of EMT [47]. Our results showed that the expression levels of BICD1 were significantly correlated with the EMT spectrum scoring (Figure 11A, 11B) and the expression levels of EMT markers in GBMs (Figure 12). This may suggest a possible connection between BICD1 expression and EMT processing, which was associated with therapeutic resistance in cancers [48, 49].
BICD1 was first reported to play a role in drosophila oocyte differentiation [50]. Another study group demonstrated that BICD1 regulated G-protein signaling and internalization [51]. BICD1 was also found to determine RNA binding and translational repression [52], and regulate the intracellular sorting and signaling of neurotrophin receptors [53]. An important study by Matanis et al., published in Nature Cell Biology (2002), demonstrated that BICD1 controls the coat complex coatomer protein I (COPI) independent Golgi-ER transport by recruiting the dynein-dynactin complex [54]. Dyneins are microtubule motors that are involved in many cellular processes, including mitosis and spindle assembly, nuclear migration and cell motility, and the transport of mRNA and a variety of cellular cargoes, including axonal and dendritic vesicles [55]. Another study suggested that the migration and proliferation of glioma cells correlate with high expression of cytoplasmic dynein and its regulatory proteins [56]. Wang et al. also reported that expression of dynein, cytoplasmic 2, heavy chain 1 (DYNC2H1) is associated with TMZ resistance in GBM cells [57]. Our analysis also showed a high correlation between the expression levels of BICD1 and cytoplasmic dyneins (Supplementary Figure 2), which suggested another possible mechanism by which high BICD1 expression may result in TMZ resistance in GBM cells through the dynein-mediated pathway.
In summary, our study indicated that high BICD1 expression is associated with poor prognosis and therapeutic response in GBMs. Further investigation will be needed to explore the definite mechanism of BICD1-associated survival or therapeutic resistance in GBM cells by connecting with IDH1, EMT, or even dynein-mediated pathway, which may provide more understanding of the pathogenesis and therapeutic resistance in GBMs.
MATERIALS AND METHODS
Cell culture
Two human glioma cell lines (U87 and T98G) were maintained in DMEM (GIBCO, Grand Island, NY, USA). Mediums were all supplemented with 10% fetal bovine serum, penicillin (100unit/ml), and streptomycin (100μg/ml). Cells were incubated in 95% air, 5% CO2 humidified atmosphere at 37°C.
MTT assay for cell viability analysis
U87MG and T98G cell viability were determined using MTT assays (Sigma Aldrich®). Cells were seeded in 96 well plates and add 20μl of 5mg/ml MTT at the end of the exposure time. The cells were incubated at 37°C for 4 hours, the medium was carefully removed, and 100μl of DMSO were added to each well. Absorbance was read at 570nm using an ELISA reader (Epoch, BioTek).
Gene expression profiling from mRNA expression microarray
Total RNA extracted from cells with the A260/280 ratio greater than 1.9 was used in the Affymetrix microarray analysis. In the analysis, hybridization was performed by using Affymetrix human U133 2.0 plus arrays and the chips were scanned by Affymetrix GeneChip scanner 3000. Then, Affymetrix DAT files were processed by Affymetrix Gene Chip Operating System (GCOS) to generate CEL files. The raw intensities in CEL files were normalized by robust multi-chip analysis, and fold-change analysis was performed using GeneSpring GX11 (Agilent Technologies).
RT-PCR
Equal amounts of total RNA were reverse transcribed into single-strand cDNA using the iScriptTM cDNA Synthesis Kit (Bio-RadTM). The mRNA was amplified using gene specific primers designed on the basis of available BICD1 mRNAs conserved in the NCBI GenBank database. Each primer was check using the NCBI primer design tool (http://www.ncbi.nlm.nih.gov/tools/primer-blast/). The expression level of BICD1 mRNAs were quantified relative to the expression level of the housekeeping gene, Glyceraldehyde-3-Phosphate-Dehydrogenase (GAPDH). The following primers were used:
BICD1: forward 5’-TGTTGAAAGCCAACA AGCAG-3’
and reverse 5’-TTGCAAACATTGCTCTCAGG-3’
(25 cycles, temperature of melting (Tm) is 50°C),
GAPDH: forward 5’-GAAGGTGAAGGTCGG AGT-3’
and reverse 5’-GAAGATGGTGATGGGATTTC-3’
(25 cycles, Tm is 60°C).
Clinical data of glioma patients from the TCGA and CGGA websites
TCGA provides two distinct cohort databases of gliomas, including glioblastoma multiforme (GBM) cohort (grade IV glioma, n=523), and glioblastoma multiforme and lower grade glioma (GBMLGG) cohort (grades II, III and IV gliomas, n=689). All clinical data was downloaded from the TCGA Portal (http://www.xenabrowser.net/). In order to verify the consistency of results, we also collected clinical data in the Chinese Glioma Genome Atlas (CGGA) cohort from the CGGA Data Portal (http://cgga.org.cn/).
Patients’ clinical information, including gender, age, Karnofsky performance score (KPS), molecular subclassification, overall survival time, time to a new tumor event, time to tumor progression, time to tumor recurrence, and therapeutic type, were collected from the aforementioned website. Patients were split for survival analysis according to age and gender. For the age factor, a cut-off of 45 years was chosen in the TCGA GBMLGG cohort for approximately separating patients into two equal groups (age≤45, n=342 vs. age>45, n=347). And the age of 65 was determined in the TCGA GBM cohort because the definition for elderly is usually over 65 as in NEJM 2017 by Perry JR et al.[11] In each patient group, the overall survival, median survival time, 2 year-survival rate and HR, were obtained and compared using the Kaplan-Meier survival analysis.
Genomic data of glioma patients from the TCGA and CGGA websites
The expression levels of candidate genes in the TCGA GBM cohort database (n=523) obtained by gene expression array (AffyU133a) and in the TCGA GBMLGG cohort database (n=689) obtained by gene expression RNAseq (polyA+ IlluminaHiSeq) were downloaded from the above TCGA Portal. For verifying the consistency of results, the gene expression profiling in the CGGA glioma cohort (n=220) obtained by mRNA expression microarray was also downloaded from the above CGGA Data Portal. Patients were split equally for survival analysis according to the expression status of BICD1 and MGMT. Patients were also grouped according to the molecular subclassification of GBMs defined by TCGA [34] and the IDH1 mutation status. In each patient group, the overall survival, median survival time, 2 year-survival rate and HR, were obtained and compared using the Kaplan-Meier survival analysis.
Statistical analysis
All statistical analyses were performed using SPSS version 20.0 software (SPSS, Chicago, Illinois, USA). The unpaired t-test (the Student’s t-test) was used for analysis of the differences in gene expression between different subgroups of patients in the TCGA GBM and GBMLGG cohorts. Associations between BICD1 expression and clinicopathological categorical variables were analyzed by Pearson’s Chi-square test. Estimates of the 2-year survival rate and survival curves were calculated using the Kaplan-Meier method, and differences in survival were compared by the log-rank test. To identify the factors that might have a significant influence on survival, univariate and multivariate analyses were performed using Cox’s proportional hazards regression modeling with and without adjustment for the expression status of candidate markers, age, gender, and molecular subclassifications, which were potentially related to survival. The scatter plot and box pictures were draw by using Prism 5 software (GraphPad software Inc.). For all analyses, a P value of <0.05 was considered statistically significant (ns: not significant; *: P<0.05; **: P<0.01; ***: P<0.001).
CONCLUSION
There is an urgent need to identify biomarkers of GBMs to indicate patients’ prognosis and response to therapy. BICD1 expression, a novel TMZ-related marker identified from GBM cell lines, may be a potential biomarker for prognosis and predicting the response to therapies, including TMZ and radiation therapies. However, the underlying mechanisms involved in BICD1-associated survival or therapeutic resistance in GBM cells, need further investigation.
Author contributions
Conception and design: S. P. H., Y. F. L. and M. H.
Development of methodology: S. P. H., Y. C. C., Y. F. L. and M. H.
Acquisition of data (provided animals, acquired and managed patients, provided facilities, etc.): S. P. H., Y. C. C., A. W., C. L. C. and Y. F. L.
Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational analysis): S. P. H., Q. H. L., C. L. C., Y. F. L., and M. H.
Drafting and revising of the manuscript: S. P. H., Q. H. L., C. L. C., Y. F. L., and M. H.
Study supervision: M.H.
Critical revision of the manuscript for important intellectual content: all authors.
ACKNOWLEDGMENTS
The study was supported by the Ministry of Science and Technology (MOST104-2320-B-038-010- MY2 and MOST 104-2320-B-038-061-MY3) to Yuan-Feng Lin. This study was also supported by Academia Sinica and Ministry of Science and Technology [MOST 104-0210-01-09-02, MOST 105-0210-01-13-01, MOST 106-0210-01-15-02] to Michael Hsiao.
CONFLICTS OF INTEREST
None declared.
REFERENCES
1. Louis DN, Ohgaki H, Wiestler OD, Cavenee WK, Burger PC, Jouvet A, Scheithauer BW, Kleihues P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007; 114: 97-109. https://doi.org/10.1007/s00401-007-0243-4.
2. Ostrom QT, Gittleman H, Fulop J, Liu M, Blanda R, Kromer C, Wolinsky Y, Kruchko C, Barnholtz-Sloan JS. CBTRUS statistical report: primary brain and central nervous system tumors diagnosed in the United States in 2008-2012. Neuro Oncol. 2015; 17: iv1-62. https://doi.org/10.1093/neuonc/nov189.
3. Reddy SP, Britto R, Vinnakota K, Aparna H, Sreepathi HK, Thota B, Kumari A, Shilpa BM, Vrinda M, Umesh S, Samuel C, Shetty M, Tandon A, et al. Novel glioblastoma markers with diagnostic and prognostic value identified through transcriptome analysis. Clin Cancer Res. 2008; 14: 2978-87. https://doi.org/10.1158/1078-0432.CCR-07-4821.
4. Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, Janzer RC, Ludwin SK, Allgeier A, Fisher B, Belanger K, Hau P, Brandes AA, Gijtenbeek J, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009; 10: 459-66. https://doi.org/10.1016/S1470-2045(09)70025-7.
5. Yung WK, Prados MD, Yaya-Tur R, Rosenfeld SS, Brada M, Friedman HS, Albright R, Olson J, Chang SM, O’Neill AM, Friedman AH, Bruner J, Yue N, et al. Multicenter phase II trial of temozolomide in patients with anaplastic astrocytoma or anaplastic oligoastrocytoma at first relapse. Temodal Brain Tumor Group. J Clin Oncol. 1999; 17: 2762-71. https://doi.org/10.1200/JCO.1999.17.9.2762.
6. Cohen MH, Johnson JR, Pazdur R. Food and Drug Administration Drug approval summary: temozolomide plus radiation therapy for the treatment of newly diagnosed glioblastoma multiforme. Clin Cancer Res. 2005; 11: 6767-71. https://doi.org/10.1158/1078-0432.CCR-05-0722.
7. Thomas RP, Recht L, Nagpal S. Advances in the management of glioblastoma: the role of temozolomide and MGMT testing. Clin Pharmacol. 2013; 5: 1-9. https://doi.org/10.2147/CPAA.S26586.
8. Zhang J, Stevens MF, Bradshaw TD. Temozolomide: mechanisms of action, repair and resistance. Curr Mol Pharmacol. 2012; 5: 102-14.
9. Preusser M, de Ribaupierre S, Wohrer A, Erridge SC, Hegi M, Weller M, Stupp R. Current concepts and management of glioblastoma. Ann Neurol. 2011; 70: 9-21. https://doi.org/10.1002/ana.22425.
10. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, Curschmann J, Janzer RC, Ludwin SK, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005; 352: 987-96. https://doi.org/10.1056/NEJMoa043330.
11. Perry JR, Laperriere N, O’Callaghan CJ, Brandes AA, Menten J, Phillips C, Fay M, Nishikawa R, Cairncross JG, Roa W, Osoba D, Rossiter JP, Sahgal A, et al. Short-course radiation plus temozolomide in elderly patients with glioblastoma. N Engl J Med. 2017; 376: 1027-37. https://doi.org/10.1056/NEJMoa1611977.
12. Wallner KE, Galicich JH, Krol G, Arbit E, Malkin MG. Patterns of failure following treatment for glioblastoma multiforme and anaplastic astrocytoma. Int J Radiat Oncol Biol Phys. 1989; 16: 1405-9.
13. Niyazi M, Siefert A, Schwarz SB, Ganswindt U, Kreth FW, Tonn JC, Belka C. Therapeutic options for recurrent malignant glioma. Radiother Oncol. 2011; 98: 1-14. https://doi.org/10.1016/j.radonc.2010.11.006.
14. Roos WP, Batista LF, Naumann SC, Wick W, Weller M, Menck CF, Kaina B. Apoptosis in malignant glioma cells triggered by the temozolomide-induced DNA lesion O6-methylguanine. Oncogene. 2007; 26: 186-97. https://doi.org/10.1038/sj.onc.1209785.
15. Hegi ME, Diserens AC, Gorlia T, Hamou MF, de Tribolet N, Weller M, Kros JM, Hainfellner JA, Mason W, Mariani L, Bromberg JE, Hau P, Mirimanoff RO, et al. MGMT gene silencing and benefit from temozolomide in glioblastoma. N Engl J Med. 2005; 352: 997-1003. https://doi.org/10.1056/NEJMoa043331.
16. Haynes HR, Camelo-Piragua S, Kurian KM. Prognostic and predictive biomarkers in adult and pediatric gliomas: toward personalized treatment. Front Oncol. 2014; 4: 47. https://doi.org/10.3389/fonc.2014.00047.
17. Brat DJ, Verhaak RG, Aldape KD, Yung WK, Salama SR, Cooper LA, Rheinbay E, Miller CR, Vitucci M, Morozova O, Robertson AG, Noushmehr H, Laird PW, et al; Cancer Genome Atlas Research Network. Comprehensive, integrative genomic analysis of diffuse lower-grade gliomas. N Engl J Med. 2015; 372: 2481-98. https://doi.org/10.1056/NEJMoa1402121.
18. Gerstner ER, Yip S, Wang DL, Louis DN, Iafrate AJ, Batchelor TT. Mgmt methylation is a prognostic biomarker in elderly patients with newly diagnosed glioblastoma. Neurology. 2009; 73: 1509-10. https://doi.org/10.1212/WNL.0b013e3181bf9907.
19. Yin AA, Zhang LH, Cheng JX, Dong Y, Liu BL, Han N, Zhang X. The predictive but not prognostic value of MGMT promoter methylation status in elderly glioblastoma patients: a meta-analysis. PLoS One. 2014; 9: e85102. https://doi.org/10.1371/journal.pone.0085102.
20. Wick W, Weller M, van den Bent M, Sanson M, Weiler M, von Deimling A, Plass C, Hegi M, Platten M, Reifenberger G. MGMT testing--the challenges for biomarker-based glioma treatment. Nat Rev Neurol. 2014; 10: 372-85. https://doi.org/10.1038/nrneurol.2014.100.
21. Preusser M, Charles Janzer R, Felsberg J, Reifenberger G, Hamou MF, Diserens AC, Stupp R, Gorlia T, Marosi C, Heinzl H, Hainfellner JA, Hegi M. Anti-O6-methylguanine-methyltransferase (MGMT) immunohistochemistry in glioblastoma multiforme: observer variability and lack of association with patient survival impede its use as clinical biomarker. Brain Pathol. 2008; 18: 520-32. https://doi.org/10.1111/j.1750-3639.2008.00153.x.
22. Galanis E, Wu W, Sarkaria J, Chang SM, Colman H, Sargent D, Reardon DA. Incorporation of biomarker assessment in novel clinical trial designs: personalizing brain tumor treatments. Curr Oncol Rep. 2011; 13: 42-9. https://doi.org/10.1007/s11912-010-0144-x.
23. Ludwig JA, Weinstein JN. Biomarkers in cancer staging, prognosis and treatment selection. Nat Rev Cancer. 2005; 5: 845-56. https://doi.org/10.1038/nrc1739.
24. Esteller M, Garcia-Foncillas J, Andion E, Goodman SN, Hidalgo OF, Vanaclocha V, Baylin SB, Herman JG. Inactivation of the DNA-repair gene MGMT and the clinical response of gliomas to alkylating agents. N Engl J Med. 2000; 343: 1350-4. https://doi.org/10.1056/NEJM200011093431901.
25. Baens M, Marynen P. A human homologue (BICD1) of the Drosophila bicaudal-D gene. Genomics. 1997; 45: 601-6. https://doi.org/10.1006/geno.1997.4971.
26. Kalinina J, Peng J, Ritchie JC, Van Meir EG. Proteomics of gliomas: initial biomarker discovery and evolution of technology. Neuro Oncol. 2011; 13: 926-42. https://doi.org/10.1093/neuonc/nor078.
27. Stein GH. T98G: an anchorage-independent human tumor cell line that exhibits stationary phase G1 arrest in vitro. J Cell Physiol. 1979; 99: 43-54. https://doi.org/10.1002/jcp.1040990107.
28. Ponten J, Macintyre EH. Long term culture of normal and neoplastic human glia. Acta Pathol Microbiol Scand. 1968; 74: 465-86.
29. Kanzawa T, Germano IM, Kondo Y, Ito H, Kyo S, Kondo S. Inhibition of telomerase activity in malignant glioma cells correlates with their sensitivity to temozolomide. Br J Cancer. 2003; 89: 922-9. https://doi.org/10.1038/sj.bjc.6601193.
30. Alonso MM, Gomez-Manzano C, Bekele BN, Yung WK, Fueyo J. Adenovirus-based strategies overcome temozolomide resistance by silencing the O6-methylguanine-DNA methyltransferase promoter. Cancer Res. 2007; 67: 11499-504. https://doi.org/10.1158/0008-5472.CAN-07-5312.
31. van Nifterik KA, van den Berg J, van der Meide WF, Ameziane N, Wedekind LE, Steenbergen RD, Leenstra S, Lafleur MV, Slotman BJ, Stalpers LJ, Sminia P. Absence of the MGMT protein as well as methylation of the MGMT promoter predict the sensitivity for temozolomide. Br J Cancer. 2010; 103: 29-35. https://doi.org/10.1038/sj.bjc.6605712.
32. Munoz JL, Rodriguez-Cruz V, Greco SJ, Ramkissoon SH, Ligon KL, Rameshwar P. Temozolomide resistance in glioblastoma cells occurs partly through epidermal growth factor receptor-mediated induction of connexin 43. Cell Death Dis. 2014; 5: e1145. https://doi.org/10.1038/cddis.2014.111.
33. Ding Z, Liu X, Liu Y, Zhang J, Huang X, Yang X, Yao L, Cui G, Wang D. Expression of far upstream element (FUSE) binding protein 1 in human glioma is correlated with c-Myc and cell proliferation. Mol Carcinog. 2015; 54: 405-15. https://doi.org/10.1002/mc.22114.
34. Verhaak RG, Hoadley KA, Purdom E, Wang V, Qi Y, Wilkerson MD, Miller CR, Ding L, Golub T, Mesirov JP, Alexe G, Lawrence M, O’Kelly M, et al. Integrated genomic analysis identifies clinically relevant subtypes of glioblastoma characterized by abnormalities in PDGFRA, IDH1, EGFR, and NF1. Cancer Cell. 2010; 17: 98-110. https://doi.org/10.1016/j.ccr.2009.12.020.
35. Wick W, Meisner C, Hentschel B, Platten M, Schilling A, Wiestler B, Sabel MC, Koeppen S, Ketter R, Weiler M, Tabatabai G, von Deimling A, Gramatzki D, et al. Prognostic or predictive value of MGMT promoter methylation in gliomas depends on IDH1 mutation. Neurology. 2013; 81: 1515-22. https://doi.org/10.1212/WNL.0b013e3182a95680.
36. Liu ZH, Hu JL, Liang JZ, Zhou AJ, Li MZ, Yan SM, Zhang X, Gao S, Chen L, Zhong Q, Zeng MS. Far upstream element-binding protein 1 is a prognostic biomarker and promotes nasopharyngeal carcinoma progression. Cell Death Dis. 2015; 6: e1920. https://doi.org/10.1038/cddis.2015.258.
37. Duan J, Bao X, Ma X, Zhang Y, Ni D, Wang H, Zhang F, Du Q, Fan Y, Chen J, Wu S, Li X, Gao Y, et al. Upregulation of far upstream element-binding protein 1 (FUBP1) promotes tumor proliferation and tumorigenesis of clear cell renal cell carcinoma. PLoS One. 2017; 12: e0169852. https://doi.org/10.1371/journal.pone.0169852.
38. Duncan R, Bazar L, Michelotti G, Tomonaga T, Krutzsch H, Avigan M, Levens D. A sequence-specific, single-strand binding protein activates the far upstream element of c-myc and defines a new DNA-binding motif. Genes Dev. 1994; 8: 465-80.
39. Bettegowda C, Agrawal N, Jiao Y, Sausen M, Wood LD, Hruban RH, Rodriguez FJ, Cahill DP, McLendon R, Riggins G, Velculescu VE, Oba-Shinjo SM, Marie SK, et al. Mutations in CIC and FUBP1 contribute to human oligodendroglioma. Science. 2011; 333: 1453-5. https://doi.org/10.1126/science.1210557.
40. Ichimura K. Molecular pathogenesis of IDH mutations in gliomas. Brain Tumor Pathol. 2012; 29: 131-9. https://doi.org/10.1007/s10014-012-0090-4.
41. Parsons DW, Jones S, Zhang X, Lin JC, Leary RJ, Angenendt P, Mankoo P, Carter H, Siu IM, Gallia GL, Olivi A, McLendon R, Rasheed BA, et al. An integrated genomic analysis of human glioblastoma multiforme. Science. 2008; 321: 1807-12. https://doi.org/10.1126/science.1164382.
42. Yan H, Parsons DW, Jin G, McLendon R, Rasheed BA, Yuan W, Kos I, Batinic-Haberle I, Jones S, Riggins GJ, Friedman H, Friedman A, Reardon D, et al. IDH1 and IDH2 mutations in gliomas. N Engl J Med. 2009; 360: 765-73. https://doi.org/10.1056/NEJMoa0808710.
43. Kang MR, Kim MS, Oh JE, Kim YR, Song SY, Seo SI, Lee JY, Yoo NJ, Lee SH. Mutational analysis of IDH1 codon 132 in glioblastomas and other common cancers. Int J Cancer. 2009; 125: 353-5. https://doi.org/10.1002/ijc.24379.
44. Amary MF, Bacsi K, Maggiani F, Damato S, Halai D, Berisha F, Pollock R, O’Donnell P, Grigoriadis A, Diss T, Eskandarpour M, Presneau N, Hogendoorn PC, et al. IDH1 and IDH2 mutations are frequent events in central chondrosarcoma and central and periosteal chondromas but not in other mesenchymal tumours. J Pathol. 2011; 224: 334-43. https://doi.org/10.1002/path.2913.
45. Housman G, Byler S, Heerboth S, Lapinska K, Longacre M, Snyder N, Sarkar S. Drug resistance in cancer: an overview. Cancers (Basel). 2014; 6: 1769-92. https://doi.org/10.3390/cancers6031769.
46. Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, Campbell LL, Polyak K, Brisken C, et al. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008; 133: 704-15. https://doi.org/10.1016/j.cell.2008.03.027.
47. Salt MB, Bandyopadhyay S, McCormick F. Epithelial-to-mesenchymal transition rewires the molecular path to PI3K-dependent proliferation. Cancer Discov. 2014; 4: 186-99. https://doi.org/10.1158/2159-8290.CD-13-0520.
48. Zheng X, Carstens JL, Kim J, Scheible M, Kaye J, Sugimoto H, Wu CC, LeBleu VS, Kalluri R. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature. 2015; 527: 525-30. https://doi.org/10.1038/nature16064.
49. Fischer KR, Durrans A, Lee S, Sheng J, Li F, Wong ST, Choi H, El Rayes T, Ryu S, Troeger J, Schwabe RF, Vahdat LT, Altorki NK, et al. Epithelial-to-mesenchymal transition is not required for lung metastasis but contributes to chemoresistance. Nature. 2015; 527: 472-6. https://doi.org/10.1038/nature15748.
50. Mahone M, Saffman EE, Lasko PF. Localized Bicaudal-C RNA encodes a protein containing a KH domain, the RNA binding motif of FMR1. EMBO J. 1995; 14: 2043-55.
51. Swift S, Xu J, Trivedi V, Austin KM, Tressel SL, Zhang L, Covic L, Kuliopulos A. A novel protease-activated receptor-1 interactor, Bicaudal D1, regulates G protein signaling and internalization. J Biol Chem. 2010; 285: 11402-10. https://doi.org/10.1074/jbc.M110.105403.
52. Zhang Y, Park S, Blaser S, Sheets MD. Determinants of RNA binding and translational repression by the Bicaudal-C regulatory protein. J Biol Chem. 2014; 289: 7497-504. https://doi.org/10.1074/jbc.M113.526426.
53. Terenzio M, Golding M, Russell MR, Wicher KB, Rosewell I, Spencer-Dene B, Ish-Horowicz D, Schiavo G. Bicaudal-D1 regulates the intracellular sorting and signalling of neurotrophin receptors. EMBO J. 2014; 33: 1582-98. https://doi.org/10.15252/embj.201387579.
54. Matanis T, Akhmanova A, Wulf P, Del Nery E, Weide T, Stepanova T, Galjart N, Grosveld F, Goud B, De Zeeuw CI, Barnekow A, Hoogenraad CC. Bicaudal-D regulates COPI-independent Golgi-ER transport by recruiting the dynein-dynactin motor complex. Nat Cell Biol. 2002; 4: 986-92. https://doi.org/10.1038/ncb891.
55. Kong X, Cho MH, Anderson W, Coxson HO, Muller N, Washko G, Hoffman EA, Bakke P, Gulsvik A, Lomas DA, Silverman EK, Pillai SG; ECLIPSE Study NETT Investigators. Genome-wide association study identifies BICD1 as a susceptibility gene for emphysema. Am J Respir Crit Care Med. 2011; 183: 43-9. https://doi.org/10.1164/rccm.201004-0541OC.
56. Suzuki SO, McKenney RJ, Mawatari SY, Mizuguchi M, Mikami A, Iwaki T, Goldman JE, Canoll P, Vallee RB. Expression patterns of LIS1, dynein and their interaction partners dynactin, NudE, NudEL and NudC in human gliomas suggest roles in invasion and proliferation. Acta Neuropathol. 2007; 113: 591-9. https://doi.org/10.1007/s00401-006-0180-7.
57. Wang H, Feng W, Lu Y, Li H, Xiang W, Chen Z, He M, Zhao L, Sun X, Lei B, Qi S, Liu Y. Expression of dynein, cytoplasmic 2, heavy chain 1 (DHC2) associated with glioblastoma cell resistance to temozolomide. Sci Rep. 2016; 6: 28948. https://doi.org/10.1038/srep28948.