
INTRODUCTION
Lung cancer is a malignant tumor with high morbidity and mortality. Lung cancer as the leading cause of cancer-related deaths caused occurred in 1.8 million people and resulted in 1.6 million deaths worldwide in 2012 [1]. It was reported that lung cancer is the highest incidence of malignant tumors in China with the annual new cases about 705,000 [2]. The cigarette smoking, environmental pollution, radon exposure, occupational exposure, and pre-existing lung disease were found to be important contributors to lung cancer risk [3, 4]. However, not all people with these risk factors develop lung cancer. About 8% of lung cancer is due to inherited factors [5]. Previous studies have identified multiple susceptibility polymorphisms in some genes that are associated with the risk of lung cancer, such as TERT [6, 7], TP63 [8] CLPTM1L [9, 10], PSMA4 [11], CHRNA3 [12], CHRNA5 [13], CRP and GPC5 [14], and so on.
Telomeres are special functional structure at the end of chromosomes and they have been deemed as a crucial role in maintaining the stability and integrity of the genome [15]. Telomere dysfunction could lead to chromosomal instability and the increase of carcinoma susceptibility so that the cells would get growth advantages, even develop into tumor cells [16, 17]. The TERT (telomerase reverse transcriptase) gene, encoding the catalytic subunit of the telomerase, is primarily regulated at the level of transcriptional initiation. The TERC (telomerase RNA component) gene, encoding the subunit of the enzyme, provides the template for the synthesis of the TTAGGG repeats. The two major telomerase genes are maintained telomere DNA length, chromosomal stability, and cellular immortality.
A number of studies have reported that single nucleotide polymorphisms (SNPs) of TERT and TERC gene are associated with a significantly higher susceptibility to multiple types of cancers [6, 17-20]. But few studies were reported on the association between SNPs in TERC gene and the risk of lung cancer. In the study, we conducted a case-control study consisting of 554 lung cancer patients and 603 healthy controls to investigate whether SNPs in TERT and TERC are associated with lung cancer risk in the Chinese Han population, and further to verify whether the conclusion we drew was in accord with the former studies.
RESULTS
In our study, the basic characteristics were shown in the Table 1. There were 416 males and 138 females in the case group with a mean age of 58.18±10.534, 496 males and 134 females in the control group with a mean age of 48.24±13.053. There was a statistical difference in age between two groups and no significant difference in gender. We adjusted gender and age in later multivariate unconditional logistic regression analysis in order to eliminate those residual confounding effects.
Table 1: Basic characteristics of the cases and controls
Characteristic |
Case (n=554) |
Control (n=603) |
P-value |
|
|---|---|---|---|---|
Gender |
Male |
416 |
469 |
1.16a |
Female |
138 |
134 |
||
Age (year; mean ± SD) |
58.18±10.534 |
48.24±13.053 |
<0.001b |
SD: Standard deviation
aP value was calculated by Welch’s t test;
bP value was calculated by Pearson’s χ 2 test.
The allele distributions and minor allele frequency (MAF) of the SNPs and the results of Hardy-Weinberg equilibrium (HWE) test were shown in the Table 2. Genotype distributions of the five SNPs appeared compatible with HWE in control subjects (all HWE-P > 0.05). In the allele model, the allele “C” of rs10936599 (TERC) and the allele “T” of rs10069690 (TERT) were found to be associated with increased the risk of lung cancer (OR = 1.32, 95% CI: 1.12-1.55, P = 0.001; OR = 1.41, 95% CI: 1.14-1.76, P = 0.002, respectively). At the level of P<0.002 (0.05/(5*5)), rs10936599 in the TERC remained significant after Bonferroni correction.
Table 2: Association between the SNPs of TERC and TERT and the risk of lung cancer
SNP ID |
Position |
Band |
Alleles A/B |
Gene(s) |
MAF |
HWE-P |
OR (95 % CI) |
P |
|
|---|---|---|---|---|---|---|---|---|---|
Case |
Control |
||||||||
rs10936599 |
169492101 |
3q26.2 |
C/T |
TERC |
0.493 |
0.425 |
0.6169 |
1.32 (1.12-1.55) |
0.001 |
rs10069690 |
1279790 |
5p15.33 |
T/C |
TERT |
0.196 |
0.147 |
0.1899 |
1.41 (1.14-1.76) |
0.002 |
rs2242652 |
1280028 |
5p15.33 |
A/G |
TERT |
0.190 |
0.163 |
0.5489 |
1.21 (0.98-1.50) |
0.079 |
rs2853677 |
1287194 |
5p15.33 |
G/A |
TERT |
0.403 |
0.366 |
0.7929 |
1.17 (1.00-1.39) |
0.060 |
rs2853676 |
1288547 |
5p15.33 |
T/C |
TERT |
0.175 |
0.154 |
0.7565 |
1.16 (0.93-1.45) |
0.184 |
SNP: single nucleotide polymorphism; MAF: Minor allele frequency; HWE: Hardy–Weinberg equilibrium; OR: Odds ratio; 95 % CI: 95 % Confidence interval
P values were calculated using two-sided Chi-squared test.
P < 0.05 indicates statistical significance.
We further assessed the association between the five SNPs and lung cancer risk under five genetic models including codominant, dominant, recessive, overdominant model and log-additive models (Table 3). In the codominant model, the genotype of “CC” of rs10936599 was associated with increased lung cancer risk compared to the genotype “TT” (OR = 1.72, 95% CI: 1.24-2.38, P = 0.004; adjusted OR = 1.72, 95% CI: 1.21-2.45, P = 0.010, respectively). In addition, rs10936599 was found to be associated with increased lung cancer risk under the dominant model (OR = 1.35, 95% CI: 1.05-1.73, P = 0.020; adjusted OR = 1.36, 95% CI: 1.04-1.79, P = 0.026), recessive model (OR = 1.53, 95% CI: 1.16-2.03, P = 0.003; adjusted OR = 1.52, 95% CI: 1.12-2.06, P = 0.007) and log-additive model (OR = 1.30, 95% CI: 1.11-1.53, P = 0.001; adjusted OR = 1.30, 95% CI: 1.09-1.55, P = 0.003).
Table 3: Genetic model analyses of the association between the SNPs and lung cancer risk
SNP-ID |
Model |
Genotype |
Case (%) |
Control (%) |
Without adjustment |
With adjustment |
||
|---|---|---|---|---|---|---|---|---|
OR (95% CI) |
P |
OR (95% CI) |
P |
|||||
rs10936599 |
Codominant |
T/T |
151 (27.4%) |
203 (33.7%) |
1 |
0.004 |
1 |
0.010 |
C/T |
258 (46.7%) |
288 (47.8%) |
1.20 (0.92-1.58) |
1.22 (0.91-1.64) |
||||
C/C |
143 (25.9%) |
112 (18.6%) |
1.72 (1.24-2.38) |
1.72 (1.21-2.45) |
||||
Dominant |
T/T |
151 (27.4%) |
203 (33.7%) |
1 |
0.020 |
1 |
0.026 |
|
C/T-C/C |
401 (72.6%) |
400 (66.3%) |
1.35 (1.05-1.73) |
1.36 (1.04-1.79) |
||||
Recessive |
T/T-C/T |
409 (74.1%) |
491 (81.4%) |
1 |
0.003 |
1 |
0.007 |
|
C/C |
143 (25.9%) |
112 (18.6%) |
1.53 (1.16-2.03) |
1.52 (1.12-2.06) |
||||
Overdominant |
T/T-C/C |
294 (53.3%) |
315 (52.2%) |
1 |
0.730 |
1 |
0.850 |
|
C/T |
258 (46.7%) |
288 (47.8%) |
0.96 (0.76-1.21) |
0.98 (0.76-1.25) |
||||
Log-additive |
--- |
--- |
--- |
1.30 (1.11-1.53) |
0.001 |
1.30 (1.09-1.55) |
0.003 |
|
rs10069690 |
Codominant |
C/C |
354 (64%) |
436 (73.4%) |
1 |
0.002 |
1 |
0.003 |
C/T |
181 (32.7%) |
141 (23.7%) |
1.58 (1.22-2.05) |
1.62 (1.22-2.15) |
||||
T/T |
18 (3.2%) |
17 (2.9%) |
1.30 (0.66-2.57) |
1.38 (0.66-2.88) |
||||
Dominant |
C/C |
354 (64%) |
436 (73.4%) |
1 |
0.001 |
1 |
0.001 |
|
C/T-T/T |
199 (36%) |
158 (26.6%) |
1.55 (1.21-1.99) |
1.59 (1.21-2.09) |
||||
Recessive |
C/C-C/T |
535 (96.8%) |
577 (97.1%) |
1 |
0.700 |
1 |
0.630 |
|
T/T |
18 (3.2%) |
17 (2.9%) |
1.14 (0.58-2.24) |
1.20 (0.58-2.50) |
||||
Overdominant |
C/C-T/T |
372 (67.3%) |
453 (76.3%) |
1 |
0.001 |
1 |
0.001 |
|
C/T |
181 (32.7%) |
141 (23.7%) |
1.56 (1.21-2.03) |
1.59 (1.20-2.11) |
||||
Log-additive |
--- |
--- |
--- |
1.41 (1.13-1.76) |
0.002 |
1.45 (1.14-1.83) |
0.002 |
|
rs2242652 |
Codominant |
G/G |
358 (64.9%) |
425 (70.5%) |
1 |
0.100 |
1 |
0.150 |
G/A |
178 (32.2%) |
160 (26.5%) |
1.32 (1.02-1.71) |
1.32 (1.00-1.74) |
||||
A/A |
16 (2.9%) |
18 (3%) |
1.06 (0.53-2.10) |
1.06 (0.50-2.23) |
||||
Dominant |
G/G |
358 (64.9%) |
425 (70.5%) |
1 |
0.041 |
1 |
0.061 |
|
G/A-A/A |
194 (35.1%) |
178 (29.5%) |
1.29 (1.01-1.66) |
1.29 (0.99-1.69) |
||||
Recessive |
G/G-G/A |
536 (97.1%) |
585 (97%) |
1 |
0.930 |
1 |
0.940 |
|
A/A |
16 (2.9%) |
18 (3%) |
0.97 (0.49-1.92) |
0.97 (0.46-2.04) |
||||
Overdominant |
G/G-A/A |
374 (67.8%) |
443 (73.5%) |
1 |
0.033 |
1 |
0.051 |
|
G/A |
178 (32.2%) |
160 (26.5%) |
1.32 (1.02-1.70) |
1.32 (1.00-1.73) |
||||
Log-additive |
--- |
--- |
--- |
1.21 (0.98-1.51) |
0.079 |
1.21 (0.96-1.53) |
0.110 |
|
rs2853677 |
Codominant |
A/A |
191 (34.5%) |
244 (40.5%) |
1 |
0.110 |
1 |
0.031 |
A/G |
279 (50.4%) |
277 (45.9%) |
1.29 (1.00-1.66) |
1.32 (1.00-1.73) |
||||
G/G |
84 (15.2%) |
82 (13.6%) |
1.31 (0.91-1.87) |
1.60 (1.08-2.37) |
||||
Dominant |
A/A |
191 (34.5%) |
244 (40.5%) |
1 |
0.036 |
1 |
0.015 |
|
A/G-G/G |
363 (65.5%) |
359 (59.5%) |
1.29 (1.02-1.64) |
1.38 (1.06-1.79) |
||||
Recessive |
A/A-A/G |
470 (84.8%) |
521 (86.4%) |
1 |
0.450 |
1 |
0.082 |
|
G/G |
84 (15.2%) |
82 (13.6%) |
1.14 (0.82-1.58) |
1.37 (0.96-1.97) |
||||
Overdominant |
A/A-G/G |
275 (49.6%) |
326 (54.1%) |
1 |
0.130 |
1 |
0.250 |
|
A/G |
279 (50.4%) |
277 (45.9%) |
1.19 (0.95-1.50) |
1.16 (0.90-1.49) |
||||
Log-additive |
--- |
--- |
--- |
1.18 (0.99-1.39) |
0.060 |
1.28 (1.06-1.54) |
0.009 |
|
rs2853676 |
Codominant |
C/C |
380 (68.6%) |
429 (71.3%) |
1 |
0.270 |
1 |
0.150 |
C/T |
154 (27.8%) |
160 (26.6%) |
1.09 (0.84-1.41) |
1.14 (0.86-1.52) |
||||
T/T |
20 (3.6%) |
13 (2.2%) |
1.74 (0.85-3.54) |
2.01 (0.93-4.31) |
||||
Dominant |
C/C |
380 (68.6%) |
429 (71.3%) |
1 |
0.320 |
1 |
0.180 |
|
C/T-T/T |
174 (31.4%) |
173 (28.7%) |
1.14 (0.88-1.46) |
1.21 (0.92-1.59) |
||||
Recessive |
C/C-C/T |
534 (96.4%) |
589 (97.8%) |
1 |
0.140 |
1 |
0.085 |
|
T/T |
20 (3.6%) |
13 (2.2%) |
1.70 (0.84-3.44) |
1.93 (0.90-4.13) |
||||
Overdominant |
C/C-T/T |
400 (72.2%) |
442 (73.4%) |
1 |
0.640 |
1 |
0.460 |
|
C/T |
154 (27.8%) |
160 (26.6%) |
1.06 (0.82-1.38) |
1.11 (0.84-1.47) |
||||
Log-additive |
--- |
--- |
--- |
1.16 (0.93-1.44) |
0.180 |
1.23 (0.97-1.56) |
0.085 |
|
SNP: single nucleotide polymorphism; OR: Odds ratio; 95% CI: 95% Confidence interval
P < 0.05 indicates statistical significance.
In the codominant model, the genotype “CT” of rs10069690 was also associated with increased lung cancer risk compared to the genotype “CC” (OR = 1.58, 95% CI: 1.22-2.05, P = 0.002; adjusted OR = 1.62, 95% CI: 1.22-2.15, P = 0.003). We also found that rs10069690 was associated with an increased the risk of lung cancer under the dominant model (OR = 1.55, 95% CI: 1.21-1.99, P = 0.001; adjusted OR = 1.59, 95% CI: 1.21-2.09, P = 0.001), overdominant model (OR = 1.56, 95% CI: 1.21-2.03, P = 0.001; adjusted OR = 1.59, 95% CI: 1.20-2.11, P = 0.001), and log-additive model (OR = 1.41, 95% CI: 1.13-1.76, P = 0.002; adjusted OR = 1.45, 95% CI: 1.14-1.83, P = 0.002).
In the codominant model, the genotype “GG and “AG” of rs2853677 were associated with increased lung cancer risk compared to the genotype “AA” (adjusted OR = 1.32, 95% CI: 1.00-1.73, P = 0.031; adjusted OR = 1.60, 95% CI: 1.08-2.37, P = 0.031) after adjustment for gender and age. The SNP rs2853677 was also associated with increased the risk of lung cancer under dominant model (adjusted OR = 1.38, 95% CI: 1.06-1.79, P = 0.015) and log-additive model (adjusted OR = 1.28, 95% CI: 1.06-1.54, P = 0.009) after adjustment for gender and age.
However, the SNP rs2242652 was found to be associated with an increased risk of lung cancer under the dominant model (OR = 1.29, 95% CI: 1.01-1.66, P = 0.041) and overdominant model (OR = 1.32, 95% CI: 1.02-1.70, P = 0.033) without adjustment. At the level of P<0.002 (0.05/(5*5)), only the SNP rs10069690 in the TERT remained significant after Bonferroni correction.
Finally, there was a strong linkage between the candidate SNPs in the TERT gene (rs10069690 and rs2242652), as shown in the Figure 1. Furthermore, we found that the haplotype “TA” of TERT was associated with a 1.32-fold increased the risk of lung cancer (OR = 1.32, 95% CI: 1.06 - 1.65, P = 0.019) (Table 4).
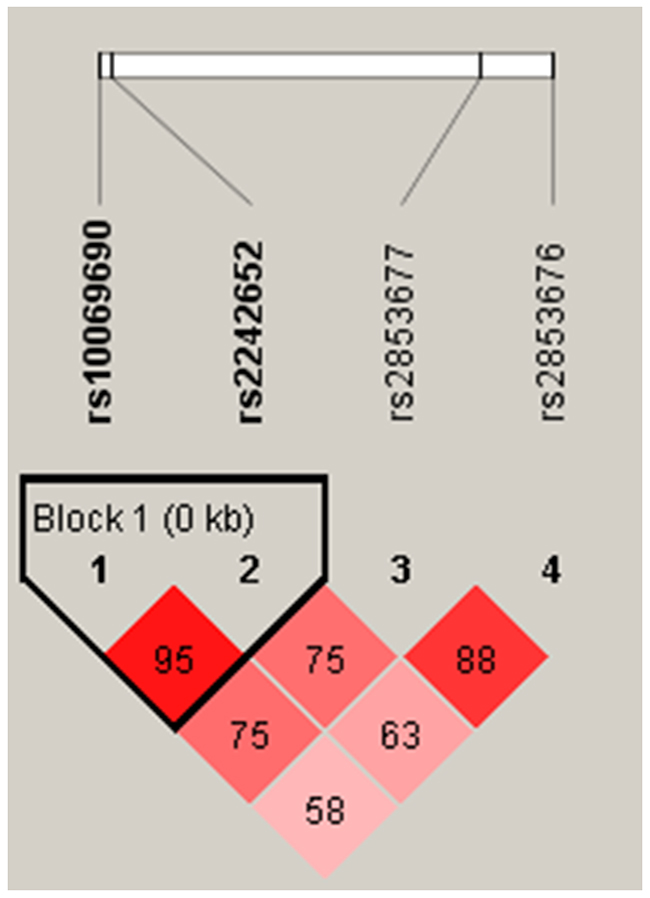
Figure 1: Haplotype block map for SNPs of the TERT gene. LD is displayed by standard color schemes with bright red for very strong LD (LOD > 2, D’ = 1), pink red (LOD > 2, D’< 1) and blue (LOD < 2, D’= 1) for partial linkage, and white (LOD < 2, D’< 1) for complete recombination.
Table 4: Haplotype frequency and their association with lung cancer risk
SNPs |
Haplotype |
Freq% |
Pa |
OR (95%CI) |
Pb |
|
|---|---|---|---|---|---|---|
case |
control |
|||||
rs10069690|rs2242652 |
CG |
0.800 |
0.832 |
- |
1 |
- |
TA |
0.187 |
0.146 |
0.014 |
1.32 (1.06 - 1.65) |
0.019 |
|
OR: Odds ratio; 95% CI: 95% Confidence interval
aP value was calculated by logistic regression crude analysis.
bP value was calculated by logistic regression analysis adjusted by gender and age.
P < 0.05 indicates statistical significance.
DISCUSSION
In the case-control study, we investigated the association between SNPs in TERC, TERT genes and the risk of lung cancer in Chinese Han population. The results showed that rs10936599 in TERC and rs10069690, rs2242652 and rs2853677 in TERT were revealed as risk factors of lung cancer. Besides, the haplotype “TA” of TERT gene was also associated with an increased risk of lung cancer.
Telomeres at the ends of chromosomes play a significant role in maintaining the stability and integrity of the genome [15]. During somatic cell divisions, telomeres progressively shorten. While the telomeres turn into a critical threshold, they may lead to cell senescence or apoptosis [21]. Previous studies reported that telomere length was associated with lung cancer risks, but the conclusions were inconsistent [22-24]. TERT gene at 5p15.33 encodes the catalytic subunit of telomerase reverse transcriptase, which is an important component of telomerase complex and associated with telomere length. TERC gene located at human chromosome 3q26.2 region, which encodes the RNA component of human telomerase. TERC serving as a template for telomere elongation can affect telomere homeostasis.
Recent studies indicated that rs10936599 in TERC was associated with telomere length and with an increased risk of cancers, but the conclusions were inconsistent [20, 25, 26]. To date, there no report on the association between rs10936599 and lung cancer risk. In our study, rs10936599 was found to be associated an increased risk of lung cancer. We suggested that rs10936599 has association with lung cancer risk may by influencing the balancing the telomere length. Further studies need to verify the assumption. Gao et al. revealed that rs10069690 in TERT was associated with an increased risk of lung cancer under dominant model [6]. In our study, there was a strong correlation between rs10069690 and lung cancer risk. Therefore, more powerful evidence in our study was provided to prove rs10069690 act as an increased risk of lung cancer. It has been reported that the rs2242652 allele of TERT influences telomere length, which was associated with risk of several cancers [27-29]. A study found that rs2242652 was associated with an increased lung cancer risk under the dominant model in the Han Chinese population [6]. However, our results showed that rs2242652 increased the risk of lung cancer in dominant model without adjustment. Therefore, it needs to be affirmed by further large-scale samples studies.
Previous studies indicated that the allele of rs2853677 in TERT, disrupted the Snail1 binding site, causing derepression of TERT transcription in response to Snail1 upregulation, was associated with a high risk of lung adenocarcinoma [17, 30, 31]. We found rs2853677 was also associated with an increased lung cancer risk. A meta-analysis study showed that rs2853676 (TERT) was associated with an increased risk of lung adenocarcinoma in Caucasians population [32]. However, in our study, no association was found between rs2853676 and lung cancer risk in the Chinese Han population. The difference in sample size and races may be the result of inconsistencies in the findings.
Finally, there are some potential limitations. First, smoking and other risk factors was not included due to lack of corresponding clinical information. Second, the associations between SNPs and telomere length, telomere length and lung cancer risk were not performed in our study. Moreover, the biological function of each SNP was not explored and these deserved further study.
In conclusion, the study demonstrated that rs10936599 (TERC), rs10069690, rs2242652 and rs2853677 in TERT, and haplotype “TA” of TERT are associated with increased risk of lung cancer in Chinese Han population. Our study not only indicated a new SNP (rs10936599) in TERC which as a risk factor of lung cancer, but supplemented and verified the findings reported before, thus there could be more powerful evidences and they can be used as diagnostic and prognostic markers in clinical studies of lung cancer patients.
MATERIALS AND METHODS
Study participants
A total of 554 patients with lung cancer and 603 healthy controls were recruited in our study. All the patients were treated in the First Affiliated Hospital of Xi’an Jiaotong University between January 2014 and August 2016. All demographic and related clinical data including age, gender were collected through a review of medical records. Inclusion criteria included patients who recently diagnosed with primary lung cancer (confirmed by histopathological examination) and patients who had not yet received any chemotherapy or radiotherapy. Patients with other types of cancers were excluded. The healthy controls were randomly recruited from unrelated subjects living Xi’an and nearby. Moreover, people with chronic disease involving brain, liver, and heart were excluded from our study.
We obtained the informed consent from all of the participants and the study protocols were approved by the Ethics Committee of First Affiliated Hospital of Xi’an Jiaotong University and in accordance with the ethical guidelines of the Declaration of Helsinki.
SNP selection and genotyping
We collected 5mL peripheral blood venous in an EDTA tube from each subject. Genomic DNA was extracted from whole blood samples using the Gold Mag-Mini Whole Blood Genomic DNA Purification Kit according to the manufacturers’ instructions (GoldMag. Co. Ltd., Xi’an, China). The DNA concentration and purity were measured by NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). We reviewed the literatures related to association between TERT and TERC polymorphisms and lung cancer and other cancers then selected five SNPs in TERC (rs10936599) and TERT (rs10069690, rs2242652, rs2853677 and rs2853676) with the minor allele frequencies (MAF) ≥5% in the Chinese Han Beijing (CHB) population by using HapMap database [6, 31-33]. The sequences of primers for amplification and extension were designed by the Sequenom MassARRAY Assay Design 3.0 software (Sequenom, Inc, San Diego, CA, USA), as shown in Table 5. Genotyping was performed using a Sequenom MassARRAY platform (Sequenom, San Diego, CA, USA) in accordance with the manufacturer’s protocol. Sequenom Typer 4.0 software was used to perform data management and analyses.
Table 5: The sequences of primers for SNPs
SNP-ID |
2nd-PCRP |
1st-PCRP |
UEP-SEQ |
|---|---|---|---|
rs10936599 |
ACGTTGGATGCAAGGGTAAAATTCCATTCTG |
ACGTTGGATGTTCCCGCTGTTTGTTCAGTC |
ATGCAGTATTCGCACCA |
rs10069690 |
ACGTTGGATGATGTGTGTTGCACACGGGAT |
ACGTTGGATGCCTGTGGCTGCGGTGGCTG |
GGGATCCTCATGCCA |
rs2242652 |
ACGTTGGATGAGGCTCTGAGGACCACAAGA |
ACGTTGGATGACAGCAGGACACGGATCCAG |
gtcgGAGGACCACAAGAAGCAGC |
rs2853677 |
ACGTTGGATGGCAAGTGGAGAATCAGAGTG |
ACGTTGGATGATCCAGTCTGACAGTCGTTG |
gggtAATCAGAGTGCACCAG |
rs2853676 |
ACGTTGGATGCAAAACTAAGACCCAAGAGG |
ACGTTGGATGTGTCTCCTGCTCTGAGACC |
agatGGAAGTCTGACGAAGGC |
SNP: Single nucleotide polymorphism; PCRP: Polymerase chain reaction primer; UEP: unique base extension primer; SEQ: Single base extension reaction
Sequences are written in the 5’→3’ (left to right) orientation.
Statistical analysis
The SPSS version 20.0 statistical package (SPSS, Chicago, IL, USA) and Microsoft Excel were used for all statistical analyses. We performed Chi-squared/ exact test to compare the expected frequencies of the genotypes in the controls in order to assess the Hardy-Weinberg equilibrium (HWE) of each SNP. All of the minor alleles were deemed as risk alleles for lung cancer susceptibility. The differences in frequency distributions of alleles were compared between cases and controls by Chi-squared test. We performed five genetic models analyses (codominant, dominant, recessive, overdominant and log-additive) to evaluate associations between the SNPs and risk of lung cancer. The odds ratios (ORs) and 95% confidence intervals (CIs) were used for crude logistic regression analysis and logistic regression analysis adjusted by gender and age. We used the Haploview software package (version 4.2) to analyze the linkage disequilibrium (LD) and haplotypes. For LD analysis and haplotype construction, genotype data of control groups were used to estimate the degree of LD by measures D’, and the D’ value > 0.8 demonstrated the related SNPs formed one haplotype block. The P-value < 0.05 was considered statistically significant. Besides, The P-value < 0.002(0.05/(5*5)) indicates statistical significance after Bonferroni correction. All statistical tests were two-sided.
ACKNOWLEDGMENTS
The authors are also grateful to the patients and control individuals for their participation in the study. We thank the clinicians and hospital staff who contributed to sample and data collection for this study.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Organization WH. Global Tuberculosis Report 2014. http://apps.who.int/iris/bitstream/10665/137094/1/9789241564809_eng.pdf.
2. Chen WQ, Zheng RS, Zhang SW. Report of Cancer Incidence and Mortality in China, 2012. China Cancer; 2016.
3. McErlean A, Ginsberg MS. Epidemiology of lung cancer. Semin Roentgenol. 2011; 46:173–77.
4. de Groot P, Munden RF. Lung cancer epidemiology, risk factors, and prevention. Radiol Clin North Am. 2012; 50:863–76.
5. Yang IA, Holloway JW, Fong KM. Genetic susceptibility to lung cancer and co-morbidities. J Thorac Dis. 2013; 5:S454–62.
6. Gao L, Thakur A, Liang Y, Zhang S, Wang T, Chen T, Meng J, Wang L, Wu F, Jin T, Li X, Liu JJ, Chen C, Chen M. Polymorphisms in the TERT gene are associated with lung cancer risk in the Chinese Han population. Eur J Cancer Prev. 2014; 23:497–501.
7. Ding Y, Du S, Sun P, Niu H, Duan M, Jin T, Yao H. Association between the TERT rs2736100 polymorphism and lung cancer risk: evidence from a case-control study and a meta-analysis. Int J Clin Exp Pathol. 2016; 9:5818-5827.
8. Hu QY, Jin TB, Wang L, Zhang L, Geng T, Liang G, Kang LL. Genetic variation in the TP63 gene is associated with lung cancer risk in the Han population. Tumour Biol. 2014; 35:1863–66.
9. Xun X, Wang H, Yang H, Wang H, Wang B, Kang L, Jin T, Chen C. CLPTM1L genetic polymorphisms and interaction with smoking and alcohol drinking in lung cancer risk: a case-control study in the Han population from northwest China. Medicine (Baltimore). 2014; 93:e289.
10. Liang Y, Thakur A, Gao L, Wang T, Zhang S, Ren H, Meng J, Geng T, Jin T, Chen M. Correlation of CLPTM1L polymorphisms with lung cancer susceptibility and response to cisplatin-based chemotherapy in a Chinese Han population. Tumour Biol. 2014; 35:12075–82.
11. Wang T, Chen T, Thakur A, Liang Y, Gao L, Zhang S, Tian Y, Jin T, Liu JJ, Chen M. Association of PSMA4 polymorphisms with lung cancer susceptibility and response to cisplatin-based chemotherapy in a Chinese Han population. Clin Transl Oncol. 2014; 35:12075–82.
12. Zhou W, Geng T, Wang H, Xun X, Feng T, Zou H, Kang L, Jin T, Chen C. CHRNA3 genetic polymorphism and the risk of lung cancer in the Chinese Han smoking population. Tumour Biol. 2015; 36:4987–92.
13. Huang CY, Xun XJ, Wang AJ, Gao Y, Ma JY, Chen YT, Jin TB, Hou P, Gu SZ. CHRNA5 polymorphisms and risk of lung cancer in Chinese Han smokers. Am J Cancer Res. 2015; 5:3241–48.
14. Zhang S, Thakur A, Liang Y, Wang T, Gao L, Yang T, Li Y, Geng T, Jin T, Chen T, Liu JJ, Chen M. Polymorphisms in C-reactive protein and Glypican-5 are associated with lung cancer risk and Gartrokine-1 influences Cisplatin-based chemotherapy response in a Chinese Han population. Dis Markers. 2015; 2015:824304.
15. Blackburn EH. Telomeres and telomerase: the means to the end (Nobel lecture). Angew Chem Int Ed Engl. 2010; 49:7405–21.
16. Lin TT, Letsolo BT, Jones RE, Rowson J, Pratt G, Hewamana S, Fegan C, Pepper C, Baird DM. Telomere dysfunction and fusion during the progression of chronic lymphocytic leukemia: evidence for a telomere crisis. Blood. 2010; 116:1899–907.
17. Li X, Xu X, Fang J, Wang L, Mu Y, Zhang P, Yao Z, Ma Z, Liu Z. Rs2853677 modulates Snail1 binding to the TERT enhancer and affects lung adenocarcinoma susceptibility. Oncotarget. 2016; 7:37825–38. https://doi.org/10.18632/oncotarget.9339.
18. Li G, Jin TB, Wei XB, He SM, Liang HJ, Yang HX, Cui Y, Chen C, Cai LB, Gao GD. Selected polymorphisms of GSTP1 and TERT were associated with glioma risk in Han Chinese. Cancer Epidemiol. 2012; 36:525–27.
19. Jin TB, Zhang JY, Li G, Du SL, Geng TT, Gao J, Liu QP, Gao GD, Kang LL, Chen C, Li SQ. RTEL1 and TERT polymorphisms are associated with astrocytoma risk in the Chinese Han population. Tumour Biol. 2013; 34:3659–66.
20. Jones AM, Beggs AD, Carvajal-Carmona L, Farrington S, Tenesa A, Walker M, Howarth K, Ballereau S, Hodgson SV, Zauber A, Bertagnolli M, Midgley R, Campbell H, et al. TERC polymorphisms are associated both with susceptibility to colorectal cancer and with longer telomeres. Gut. 2012; 61:248–54.
21. Young NS. Telomere biology and telomere diseases: implications for practice and research. Hematology (Am Soc Hematol Educ Program). 2010; 2010:30–35.
22. Karimi B, Yunesian M, Nabizadeh R, Mehdipour P, Aghaie A. Is Leukocyte Telomere Length Related with Lung Cancer Risk?: A Meta-Analysis. Iran Biomed J. 2016.
23. Lan Q, Cawthon R, Gao Y, Hu W, Hosgood HD 3rd, Barone-Adesi F, Ji BT, Bassig B, Chow WH, Shu X, Cai Q, Xiang Y, Berndt S, et al. Longer telomere length in peripheral white blood cells is associated with risk of lung cancer and the rs2736100 (CLPTM1L-TERT) polymorphism in a prospective cohort study among women in China. PLoS One. 2013; 8:e59230.
24. Jang JS, Choi YY, Lee WK, Choi JE, Cha SI, Kim YJ, Kim CH, Kam S, Jung TH, Park JY. Telomere length and the risk of lung cancer. Cancer Sci. 2008; 99:1385–89.
25. Weng Q, Du J, Yu F, Huang T, Chen M, Lv H, Ma H, Hu Z, Jin G, Hu Y, Shen H. The known genetic loci for telomere length may be involved in the modification of telomeres length after birth. Sci Rep. 2016; 6:38729.
26. Wang M, Chu H, Lv Q, Wang L, Yuan L, Fu G, Tong N, Qin C, Yin C, Zhang Z, Xu J. Cumulative effect of genome-wide association study-identified genetic variants for bladder cancer. Int J Cancer. 2014; 135:2653–60.
27. Wu Y, Yan M, Li J, Li J, Chen Z, Chen P, Li B, Chen F, Jin T, Chen C. Genetic polymorphisms in TERT are associated with increased risk of esophageal cancer. Oncotarget. 2017; 8:10523–30. https://doi.org/10.18632/oncotarget.14451.
28. Bojesen SE, Pooley KA, Johnatty SE, Beesley J, Michailidou K, Tyrer JP, Edwards SL, Pickett HA, Shen HC, Smart CE, Hillman KM, Mai PL, Lawrenson K, et al, and Australian Cancer Study, and Australian Ovarian Cancer Study, and Kathleen Cuningham Foundation Consortium for Research into Familial Breast Cancer (kConFab), and Gene Environment Interaction and Breast Cancer (GENICA), and Swedish Breast Cancer Study (SWE-BRCA), and Hereditary Breast and Ovarian Cancer Research Group Netherlands (HEBON), and Epidemiological study of BRCA1 & BRCA2 Mutation Carriers (EMBRACE), and Genetic Modifiers of Cancer Risk in BRCA1/2 Mutation Carriers (GEMO). Multiple independent variants at the TERT locus are associated with telomere length and risks of breast and ovarian cancer. Nat Genet. 2013; 45:371–84, e1–2.
29. Pellatt AJ, Wolff RK, Torres-Mejia G, John EM, Herrick JS, Lundgreen A, Baumgartner KB, Giuliano AR, Hines LM, Fejerman L, Cawthon R, Slattery ML. Telomere length, telomere-related genes, and breast cancer risk: the breast cancer health disparities study. Genes Chromosomes Cancer. 2013; 52:595–609.
30. Zanetti KA, Wang Z, Aldrich M, Amos CI, Blot WJ, Bowman ED, Burdette L, Cai Q, Caporaso N, Chung CC, Gillanders EM, Haiman CA, Hansen HM, et al. Genome-wide association study confirms lung cancer susceptibility loci on chromosomes 5p15 and 15q25 in an African-American population. Lung Cancer. 2016; 98:33–42.
31. Shiraishi K, Kunitoh H, Daigo Y, Takahashi A, Goto K, Sakamoto H, Ohnami S, Shimada Y, Ashikawa K, Saito A, Watanabe S, Tsuta K, Kamatani N, et al. A genome-wide association study identifies two new susceptibility loci for lung adenocarcinoma in the Japanese population. Nat Genet. 2012; 44:900–03.
32. Cao JL, Yuan P, Abuduwufuer A, Lv W, Yang YH, Hu J. Association between the TERT genetic polymorphism rs2853676 and cancer risk: meta-analysis of 76 108 cases and 134 215 controls. PLoS One. 2015; 10:e0128829.
33. Liu M, Li B, Guo W, Zhang X, Chen Z, Li J, Yan M, Chen C, Jin T. Association between single nucleotide polymorphisms in the TSPYL6 gene and breast cancer susceptibility in the Han Chinese population. Oncotarget. 2016; 7:54771–81. https://doi.org/10.18632/oncotarget.10754.