
INTRODUCTION
Osteoarthritis (OA), a common form of arthritis, is characterized by degradation and destruction of cartilage matrix and inflammatory responses in chondrocytes [1]. The pathophysiology of OA is complex. However, accumulating evidence suggested that inflammation played a critical role in the development of OA [2, 3]. IL-1β is an important inflammatory cytokine that involved in the pathologic process of OA [4]. IL-1β could induce the activation of NF-κB signaling pathway, which leads to the release of inflammatory mediators [5]. These inflammatory mediators, such as PGE2, NO, and MMP, lead to articular cartilage damage [6]. In recent years, non-steroidal anti-inflammatory drugs were often used in the treatment of OA [7]. However, due to the numerous side effects, the development of effective and safe drugs to treat OA is urgently needed.
Saikosaponin a (SSa), a bioactive ingredient isolated from Bupleurum falcatum, has been known to exhibit anti-inflammatory effect [8]. Previous study showed that SSa could inhibit LPS-induced inflammatory cytokines production in RAW264.7 cells [9]. SSa also inhibited carrageenan-induced paw edema in rats and acetic acid-induced vascular permeability in mice [8]. Furthermore, SSa has been reported to inhibit oxidative stress and inflammation in LPS-stimulated human umbilical vein endothelial cells [10]. SSa has been reported to inhibit inflammation in hypertrophied 3T3-L1 adipocytes [11]. However, the effects of SSa on IL-1β-stimulated chondrocytes has not been reported. In the present study, we aimed to investigate the anti-inflammatory effect of SSa on IL-1β-stimulated human osteoarthritis chondrocytes and clarify the underling mechanism.
RESULTS
SSa did not affect the viability of human osteoarthritis chondrocytes
In this study, we firstly detected the effects of SSa on the viability of human osteoarthritis chondrocytes. As shown in Figure 1, compared with the control group, SSa at concentration of 5, 10, 15μM did not affect the viability of human osteoarthritis chondrocytes (Figure 1). Therefore, the concentration of 5, 10, 15μM were used in the subsequent study.
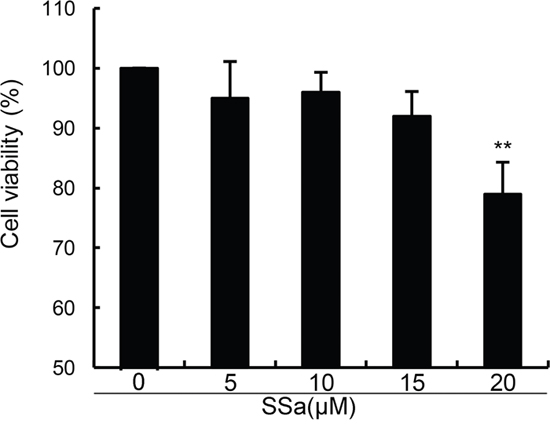
Figure 1: Effects of SSa on the cell viability of chondrocytes. The values presented are the means ± S.E.M. of three independent experiments. *P < 0.05, **P < 0.01 vs. control group.
SSa inhibits IL-1β-induced NO and PGE2 production in human osteoarthritis chondrocytes
We investigated the effects of SSa on IL-1β-induced NO and PGE2 production to test whether SSa exhibited anti-inflammatory effects in human osteoarthritis chondrocytes. As shown in Figure 2, SSa alone did affect the production of NO and PGE2. Compared with the control group, the levels of NO and PGE2 increased significantly in IL-1β-stimulated human osteoarthritis chondrocytes. However, IL-1β-induced NO and PGE2 production were significantly inhibited by treatment of SSa.
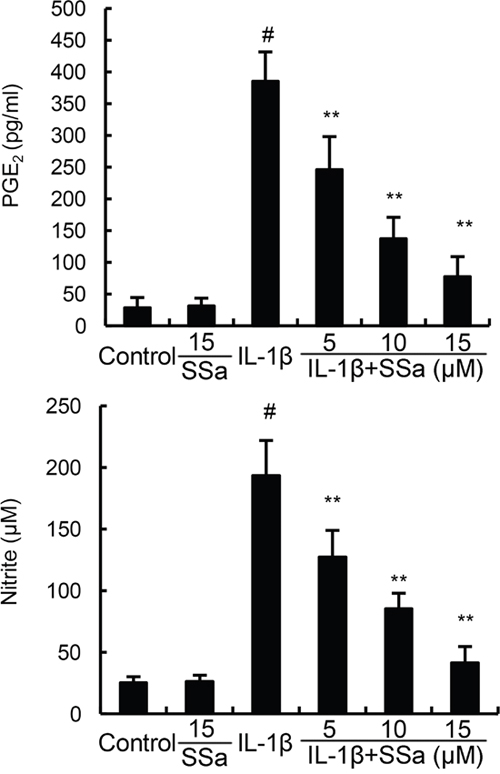
Figure 2: SSa inhibits IL-1β-induced NO and PGE2 production. The data presented are the means ± S.E.M. of three independent experiments. #P < 0.05 vs. control group; *P < 0.05, **P < 0.01 vs. IL-1β group.
Effects of SSa on IL-1β-induced MMP1, MMP3, and MMP13 expression
We investigated the effects of SSa on IL-1β-induced MMP1, MMP3, and MMP13 production by ELISA and qRT-PCR. As shown in Figure 3, SSa alone did affect the expression of MMP1, MMP3, and MMP13. Compared with the control group, the mRNA and protein levels of MMP1, MMP3, and MMP13 increased significantly in IL-1β-stimulated human osteoarthritis chondrocytes. However, IL-1β-induced MMP1, MMP3, and MMP13 production were significantly inhibited by treatment of SSa (Figure 3).
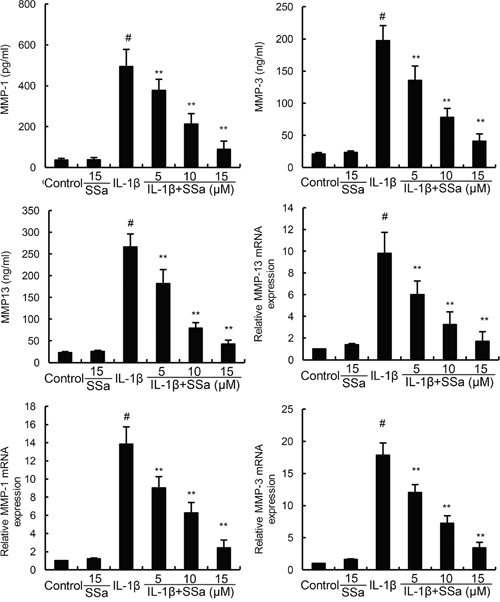
Figure 3: SSa inhibits IL-1β-induced MMP1, MMP3, and MMP13 production. The data presented are the means ± S.E.M. of three independent experiments. #P < 0.05 vs. control group; *P < 0.05, **P < 0.01 vs. IL-1β group.
Effects of SSa on IL-1β-induced NF-κB activation in human osteoarthritis chondrocytes
We investigated the effects of SSa on IL-1β-induced NF-κB activation to test the anti-inflammatory mechanism of SSa in human osteoarthritis chondrocytes. As shown in Figure 4, compared with the control group, the levels of phosphorylation of NF-κB p65 and IκBα increased significantly in IL-1β-stimulated human osteoarthritis chondrocytes. However, IL-1β-induced NF-κB activation were significantly inhibited by treatment of SSa.
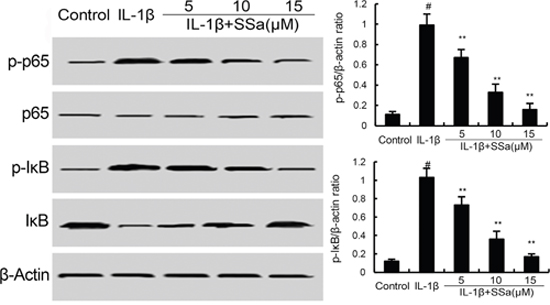
Figure 4: SSa inhibits IL-1β-induced NF-κB activation and IκBα degradation. The values presented are the means ± S.E.M. of three independent experiments. #P < 0.05 vs. control group; *P < 0.05, **P < 0.01 vs. IL-1β group.
Effects of SSa on LXRα expression
LXRα, the members of the nuclear hormone receptor superfamily, has been reported to have the ability to regulate the activation of NF-κB [12]. Therefore, in the present study, the effects of SSa on LXRα expression were detected by western blot analysis. As shown in Figure 5, SSa up-regulated the expression of LXRα in a dose-dependent manner.
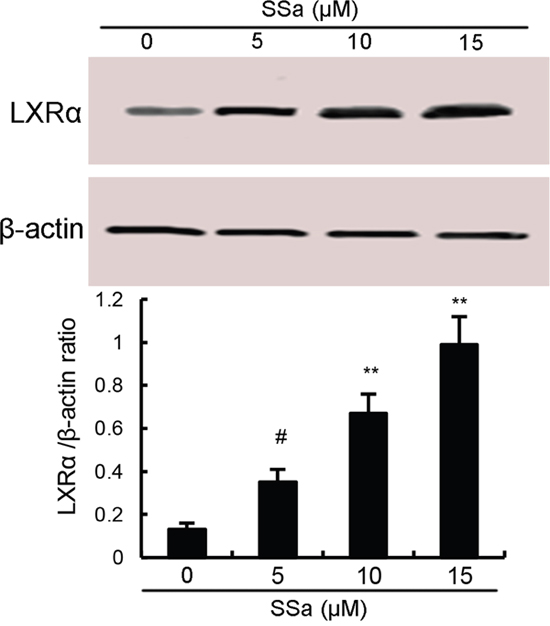
Figure 5: Effects of sesamin on LXRα expression. The values presented are the means ± S.E.M. of three independent experiments. #P < 0.05 vs. control group.
The anti-inflammatory effects of SSa is regulated by LXRα
To further investigate the anti-inflammatory mechanism of SSa, LXRα was blocked by its inhibitor GGPP. As shown in Figure 6, the inhibition of SSa on NO, PGE2, MMP1, MMP3, and MMP13 production, as well as NF-κB activation were reversed by GGPP. The results indicated that SSa inhibited IL-1β-induced inflammation in human osteoarthritis chondrocytes by activating LXRα (Figure 6). Furthermore, the inhibition of SSa on NO, PGE2, MMP1, MMP3, and MMP13 production were reversed when LXRα was knockdown (Figure 7B).
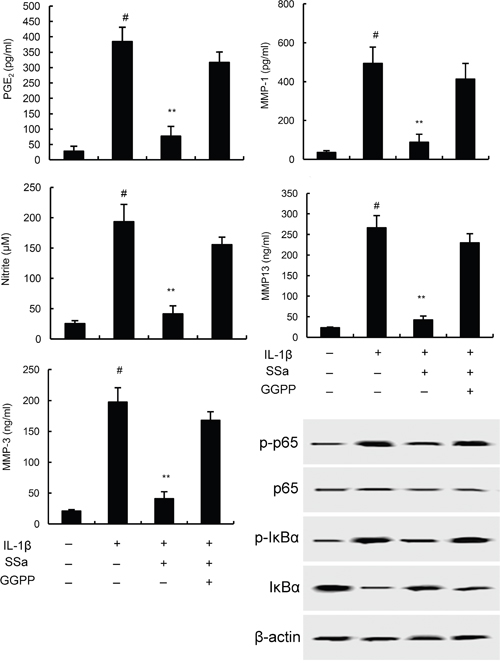
Figure 6: Effects of LXRα inhibitor GGPP (20 μM) on the anti-inflammatory effects of SSa. The values presented are the means ± SEM of three independent experiments. #p < 0.05 vs. control group; *p < 0.05, **p < 0.01 vs. LPS group.
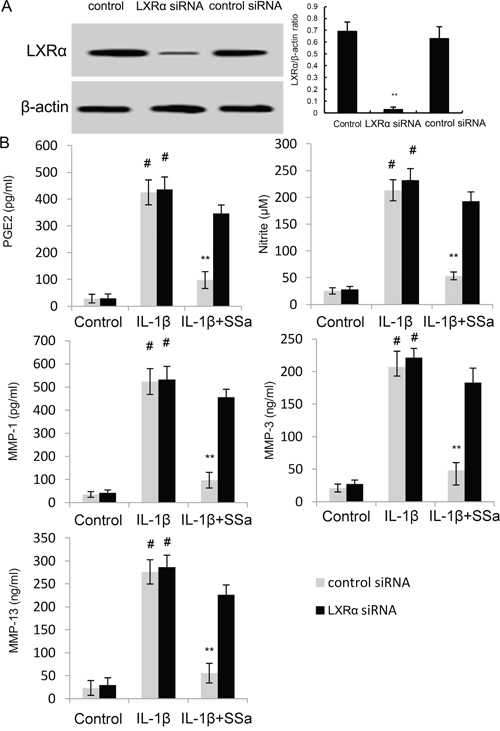
Figure 7: (A) Effects of siRNA on LXRα expression was detected by western blot analysis. (B) Effects of LXRα siRNA on the anti-inflammatory effects of SSa. The values presented are the means ± SEM of three independent experiments. #p < 0.05 vs. control group; *p < 0.05, **p < 0.01 vs. LPS group.
DISCUSSION
SSa has been reported to have anti-inflammatory effect [13]. Previous study showed that SSa could inhibit LPS-induced inflammatory response by inducing LXRα activation [14]. However, whether SSa could inhibit IL-1β-induced inflammatory response remain unclear. The aim of this study was to assess the anti-inflammatory effects of SSa on osteoarthritis. In the present study, we found that SSa significantly inhibited IL-1β-induced inflammatory response in chondrocytes. The anti-inflammatory mechanism of SSa was through activating LXRα, which subsequently inhibited IL-1β-induced NF-κB activation.
Many in vitro and in vivo studies demonstrated that inflammation play a fundamental role in the damage of articular tissues [15, 16]. The release of inflammatory mediators, such as PGE2 and NO result in destruction of the articular joint, leading to development of OA [6, 17]. Furthermore, these inflammatory mediators could induce the production of MMPs, which could induce degradation of ECM in OA articular cartilage [18]. Inhibition of these inflammatory mediators and MMPs could attenuate the development of OA [19, 20]. In the present study, we found that SSa significantly inhibited IL-1β-induced PGE2 and NO, as well as MMP1, MMP3, and MMP13 production. NF-κB is transcription factor that regulates a large body of inflammatory genes expression [10, 21, 22]. Previous study suggested that NF-κB could be used as a potential therapeutic target in OA [23]. Inhibition of NF-κB activation could attenuate pain of OA [24, 25]. In the present study, our results showed that SSa significantly inhibited IL-1β-induced NF-κB activation in chondrocytes. These results indicated that SSa inhibited IL-1β-induced inflammation in chondrocytes by inhibiting NF-κB activation.
LXRα is a nuclear receptor that induces genes controlling cholesterol homeostasis and lipogenesis [26]. It is a ligand-activated transcription factor that recently has been reported to have anti-inflammatory effects [27]. A large body of studies showed that many natural compounds could activate LXRα to exhibit its anti-inflammatory effects [14, 28]. Previous study showed that activating LXRα could attenuate pain in a rat osteoarthritis model [29]. In addition, study showed that LXRα agonist had the ability to inhibit NF-κB activation. To clarify the anti-inflammatory mechanism of SSa, we detected whether SSa could activate LXRα. Our results showed that SSa significantly up-regulated the expression of LXRα. Furthermore, our results showed that blocking LXRα, the anti-inflammatory effects of SSa were inhibited. Taken together, our results showed that SSa exhibited its anti-inflammatory effects by activating LXRα.
In conclusion, in the present study, we showed that SSa significantly inhibited IL-1β-induced inflammatory response by inhibiting NF-κB activation. Furthermore, SSa has been found to up-regulate the expression of LXRα, which subsequently inhibited the activation of NF-κB. SSa may be a potential therapeutic agent for osteoarthritis.
MATERIALS AND METHODS
Chemicals and reagents
SSa (purity>98%) was purchased from the National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China). Geranyl geranyl pyrophosphate (GGPP) was purchased from Sigma Chem. (St. Louis, USA). Recombinant human IL-1β, MMP1, MMP3, MMP13, and PGE2 ELISA kits were purchased from R&D systems (Minneapolis, MN, USA). Antibodies for LXRα, IκBα, and NF-κB were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). NO detection kit was purchased from Nanjing Jiancheng Bioengineering Institute. (Nanjing, China).
Cell culture and treatment
The experiments were done in accordance with the guidelines established by the Jilin University Animal Care and Use Committee. The protocols were reviewed and approved by the committee. OA human cartilage tissues were obtained from 12 OA patients. Primary human osteoarthritis chondrocytes were isolated as described previously [30]. Then, the cells were cultured in DMEM containing 10% fetal bovine serum. The cells were pretreated with SSa (5, 10, 15μM) 12 h before IL-1β treatment. For LXRα inhibitory experiment, the cells were pretreated with or without 20 μM GGPP for 2 h, then the cells were pretreated with SSa (5, 10, 15μM) 12 h before IL-1β treatment. Cells between passages 1 to 3 were used in this study.
Cell viability
The effects of SSa on the viability of human osteoarthritis chondrocytes were detected by MTT assay. In brief, human osteoarthritis chondrocytes were cultured in 96-well plate (5×103cells/well) for 12 h. Then, the cells were treated with different concentrations of SSa. 24 h later, the cells were incubated with MTT for 4 h and the insoluble formazan product was dissolved in DMSO. Finally, the optical density was read using a microplate reader.
Inflammatory mediator assay
The level of NO in the culture medium was detected by Griess reagent (Nanjing Jiancheng Bioengineering Institute, Jiangsu, China) according to the manufacturer’s instructions. The levels of PGE2, MMP1, MMP3, and MMP13 in the culture medium were detected by ELISA kits (R&D systems, Minneapolis, MN, USA) according to the manufacturer’s instructions.
Western blot analysis
The cells were pretreated with SSa (5, 10, 15μM) 12 h before IL-1β (10 ng/ml) treatment. 1 h later, the cells were incubated with 200 μl cell lysis buffer (Beyotime, China) for 5min and subjected to centrifugation with 12000 g/min for 10min. Protein concentration were detected by BCA method. Equal amount of proteins were separated on 12% SDS-PAGE and transferred to PVDF membranes. Then, the membranes were blocked with 3% BSA and incubated with primary antibodies and secondary antibodies. Finally, the membranes were visualized by ECL detection reagents and imaged with the Pxigel imaging system.
RNA extraction and qRT-PCR
Total RNA was extracted using the RNA simple total RNA kit (Tiangen, China). The cDNA was synthesized using a primescript RT reagent kit (Takara, Japan). Primers were obtained from Sangon Biotech Co. Ltd (Shanghai, China). Synthesized cDNA used for RT-PCR was performed with a 7500 real-time PCR system (Applied Biosystems, Carlsbad, CA, USA) as following: 50 °C for 2 min and 95°C for 10min followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min.
Cells transfection with siRNA
Chondrocytes were transfected with siRNA-LXRα (100 nM) or control siRNA (100 nM) with Lipofectamine 2000 transfection reagent (Thermo, USA) for 24 h according to the manufacturer’s instructions. Then, the cells were incubated with SSa and stimulated with IL-1β. The inhibition of siRNA on LXRα expression was detected by western blot analysis.
Statistical analysis
All the values were presented as the mean ± S.E.M. The differences between groups were analyzed using one-way ANOVA followed by post hoc analyses using the Tukey test. P <0.05 were considered to indicate statistical significance.
CONFLICTS OF INTEREST
All authors declare that they have no conflicts of interest.
REFERENCES
1. Hedbom E, Hauselmann HJ. Molecular aspects of pathogenesis in osteoarthritis: the role of inflammation. Cell Mol Life Sci. 2002; 59:45-53.
2. Kapoor M, Martel-Pelletier J, Lajeunesse D, Pelletier JP, Fahmi H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat Rev Rheumatol. 2011; 7:33-42.
3. Pelletier JP, Martel-Pelletier J, Abramson SB. Osteoarthritis, an inflammatory disease: potential implication for the selection of new therapeutic targets. Arthritis Rheum. 2001; 44:1237-1247.
4. Richette P, Francois M, Vicaut E, Fitting C, Bardin T, Corvol M, Savouret JF, Rannou F. A high interleukin 1 receptor antagonist/IL-1beta ratio occurs naturally in knee osteoarthritis. J Rheumatol. 2008; 35:1650-1654.
5. Csaki C, Mobasheri A, Shakibaei M. Synergistic chondroprotective effects of curcumin and resveratrol in human articular chondrocytes: inhibition of IL-1beta-induced NF-kappaB-mediated inflammation and apoptosis. Arthritis Res Ther. 2009; 11:R165.
6. Abramson SB, Attur M, Amin AR, Clancy R. Nitric oxide and inflammatory mediators in the perpetuation of osteoarthritis. Curr Rheumatol Rep. 2001; 3:535-541.
7. Silverstein FE, Faich G, Goldstein JL, Simon LS, Pincus T, Whelton A, Makuch R, Eisen G, Agrawal NM, Stenson WF, Burr AM, Zhao WW, Kent JD, et al. Gastrointestinal toxicity with celecoxib vs nonsteroidal anti-inflammatory drugs for osteoarthritis and rheumatoid arthritis: the CLASS study: a randomized controlled trial. Celecoxib Long-term Arthritis Safety Study. JAMA. 2000; 284:1247-1255.
8. Lu CN, Yuan ZG, Zhang XL, Yan R, Zhao YQ, Liao M, Chen JX. Saikosaponin a and its epimer saikosaponin d exhibit anti-inflammatory activity by suppressing activation of NF-kappaB signaling pathway. Int Immunopharmacol. 2012; 14:121-126.
9. Zhu J, Luo C, Wang P, He Q, Zhou J, Peng H. Saikosaponin A mediates the inflammatory response by inhibiting the MAPK and NF-kappaB pathways in LPS-stimulated RAW 264.7 cells. Exp Ther Med. 2013; 5:1345-1350.
10. Fu Y, Hu X, Cao Y, Zhang Z, Zhang N. Saikosaponin a inhibits lipopolysaccharide-oxidative stress and inflammation in Human umbilical vein endothelial cells via preventing TLR4 translocation into lipid rafts. Free Radic Biol Med. 2015; 89:777-785.
11. Kim SO, Park JY, Jeon SY, Yang CH, Kim MR. Saikosaponin a, an active compound of Radix Bupleuri, attenuates inflammation in hypertrophied 3T3-L1 adipocytes via ERK/NF-kappaB signaling pathways. Int J Mol Med. 2015; 35:1126-1132.
12. Bi X, Song J, Gao J, Zhao J, Wang M, Scipione CA, Koschinsky ML, Wang ZV, Xu S, Fu G. Activation of liver X receptor attenuates lysophosphatidylcholine-induced IL-8 expression in endothelial cells via the NF-kappaB pathway and SUMOylation. J Cell Mol Med. 2016; 20:2249-2258.
13. Wu SJ, Tam KW, Tsai YH, Chang CC, Chao JC. Curcumin and saikosaponin a inhibit chemical-induced liver inflammation and fibrosis in rats. Am J Chin Med. 2010; 38:99-111.
14. Wei Z, Wang J, Shi M, Liu W, Yang Z, Fu Y. Saikosaponin a inhibits LPS-induced inflammatory response by inducing liver X receptor alpha activation in primary mouse macrophages. Oncotarget. 2016; 7:48995-49007. https://doi.org/10.18632/oncotarget.9863.
15. Barrett AJ. The possible role of neutrophil proteinases in damage to articular cartilage. 1978. Agents Actions. 1994; 43:194-200; discussion 200-191.
16. Lee CG, Da Silva CA, Dela Cruz CS, Ahangari F, Ma B, Kang MJ, He CH, Takyar S, Elias JA. Role of chitin and chitinase/chitinase-like proteins in inflammation, tissue remodeling, and injury. Annu Rev Physiol. 2011; 73:479-501.
17. Goldring MB. Osteoarthritis and cartilage: the role of cytokines. Curr Rheumatol Rep. 2000; 2:459-465.
18. Goldring SR, Goldring MB. The role of cytokines in cartilage matrix degeneration in osteoarthritis. Clin Orthop Relat Res. 2004; S27-36.
19. Sandell LJ, Aigner T. Articular cartilage and changes in arthritis. An introduction: cell biology of osteoarthritis. Arthritis Res. 2001; 3:107-113.
20. Goldring MB, Otero M, Plumb DA, Dragomir C, Favero M, El Hachem K, Hashimoto K, Roach HI, Olivotto E, Borzi RM, Marcu KB. Roles of inflammatory and anabolic cytokines in cartilage metabolism: signals and multiple effectors converge upon MMP-13 regulation in osteoarthritis. Eur Cell Mater. 2011; 21:202-220.
21. Pahl HL. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene. 1999; 18:6853-6866.
22. Wang J, Guo C, Wei Z, He X, Kou J, Zhou E, Yang Z, Fu Y. Morin suppresses inflammatory cytokine expression by downregulation of nuclear factor-kappaB and mitogen-activated protein kinase (MAPK) signaling pathways in lipopolysaccharide-stimulated primary bovine mammary epithelial cells. J Dairy Sci. 2016; 99:3016-3022.
23. Roman-Blas JA, Jimenez SA. NF-kappaB as a potential therapeutic target in osteoarthritis and rheumatoid arthritis. Osteoarthritis Cartilage. 2006; 14:839-848.
24. Marcu KB, Otero M, Olivotto E, Borzi RM, Goldring MB. NF-kappaB signaling: multiple angles to target OA. Curr Drug Targets. 2010; 11:599-613.
25. Rigoglou S, Papavassiliou AG. The NF-kappaB signalling pathway in osteoarthritis. Int J Biochem Cell Biol. 2013; 45:2580-2584.
26. Schultz JR, Tu H, Luk A, Repa JJ, Medina JC, Li L, Schwendner S, Wang S, Thoolen M, Mangelsdorf DJ, Lustig KD, Shan B. Role of LXRs in control of lipogenesis. Genes Dev. 2000; 14:2831-2838.
27. Hong C, Tontonoz P. Coordination of inflammation and metabolism by PPAR and LXR nuclear receptors. Curr Opin Genet Dev. 2008; 18:461-467.
28. Fu Y, Wei Z, Zhou E, Zhang N, Yang Z. Cyanidin-3-O-beta-glucoside inhibits lipopolysaccharide-induced inflammatory response in mouse mastitis model. J Lipid Res. 2014; 55:1111-1119.
29. Li N, Rivera-Bermudez MA, Zhang M, Tejada J, Glasson SS, Collins-Racie LA, Lavallie ER, Wang Y, Chang KC, Nagpal S, Morris EA, Flannery CR, Yang Z. LXR modulation blocks prostaglandin E2 production and matrix degradation in cartilage and alleviates pain in a rat osteoarthritis model. Proc Natl Acad Sci U S A. 2010; 107:3734-3739.
30. Cheng AW, Stabler TV, Bolognesi M, Kraus VB. Selenomethionine inhibits IL-1beta inducible nitric oxide synthase (iNOS) and cyclooxygenase 2 (COX2) expression in primary human chondrocytes. Osteoarthritis Cartilage. 2011; 19:118-125.