
Thyroid carcinomas of follicular cell origin represent the most frequent endocrine malignancy and include papillary (PTC), follicular (FTC), and anaplastic (ATC) types. PTC with its incidence increasing by almost 5% each year, currently ranks as the eighth most common malignancy diagnosed in women [1]. Surgical treatment, thyroid suppressive therapy and radioiodine treatment in patients at high risk for recurrence is standard [2]. Some clinical factors have been used to predict which patients with thyroid cancer might have worse prognoses. Factors such as older age (>45 years old), male gender, certain histologic subtypes (i.e. tall cell, columnar cell and diffuse sclerosing variants of PTC), tumor size greater than 4 cm and presence of extrathyroidal extension are associated with neck recurrences and distant metastases [3].
Activation of the RET/RAS/BRAF/MAPK (mitogen-activated extra-cellular signal regulated kinase) signaling pathway is the most common oncogenic event in PTC tumorigenesis. More recently certain genetic alterations in PTCs have been analyzed and some of these have also been linked to poorer outcomes. Most PTCs have either a RET/PTC translocation (10–50% of PTCs) [4,5], Ras mutations (about 12% of PTCs) [6], or the BRAFV600E mutation (29–83% of PTCs) [7-9]. The V600E hotspot mutation affects the exon 15 of BRAF, a member of the RAF family of kinases, has been recently identified in PTC. BRAFV600E can not only initiate tumorigenesis in normal thyroid follicular cells but is also required to maintain and promote the progression of PTC to aggressive PTC [10,11]. Of relevance, BRAFV600E oncoprotein elicits elevated kinase activity that leads to activation of the MAPK (i.e. MEK1/2 and ERK1/2) signaling pathway [12-14]. Some clinical correlative studies have pointed to the increased incidence of worse histopathologic parameters such as lymphovascular invasion, extrathyroidal extension, neck lymph nodal disease, and mortality in those with BRAFV600E mutation [10,15, 16]. Animal studies also point to an important role for this mutation in the aggressive behavior of PTCs; thyroid tumors from BRAFV600E transgenic mice display extrathyroidal extension and foci of poorly/undifferentiated thyroid carcinoma [17]. In addition, we have recently shown that an orthotopic mouse model of ATC cells harboring BRAFV600E mutation and engineered to express GFP (Green Fluorescent Protein) displays tumor aggressiveness and lymph node and lung metastases that recapitulates an advanced human thyroid cancer [18].
BRAFV600E is crucially required to maintain and promote thyroid cancer progression [19]. Little information is known in PTC and ATC about which molecular targets of the BRAFV600E mutation and its consequent downstream phospho-MEK1/2 and phospho-ERK1/2 kinases deregulation leads to this worse prognosis. While it is not clearly understood how BRAFV600E mutation leads to more aggressive PTCs, altered genes and pathways that have been studies include: aberrant methylation of important tumor suppressor genes (i.e. TIMP-3), up-regulation of metallo-proteases-2, -3, -9 and -13 (MMP-2, -3, -9 and -13) expression [20,16], up-regulation of micro-RNAs (i.e. miR-146b) [10,21], lower expression of thyroperoxidase (TPO) and sodium iodide symporter (NIS) genes (genes important for iodine metabolism and iodine cell uptake) [22], and chromosomal instability described in rat thyroid cells [23].
We have recently investigated how BRAFV600E positive PTCs differ in the expression of gene sets and found up-regulation of Extracellular Matrix (ECM) microenvironment genes in BRAF mutated PTC compared with wild type (wt) BRAF PTC or normal thyroid tissue [19]. ECM is a three-dimensional structure surrounding human cells [24]. ECM is formed of cellular and non-cellular components. The cellular component includes: endothelial and lymphatic cells, fibroblasts, immune cells (i.e. macrophages, lymphocytes), adipocytes. The non-cellular components include soluble factors such as growth factors and cytokines, and soluble (e.g. Vascular Endothelial Growth Factor (VEGF)) and insoluble proteins (e.g. Thrombospondin-1 (TSP-1), Fibronectin (FN), collagens, laminins, etc.). The normal ECM had been considered for many years as architectural scaffolding but many studies over the last decade have shown important links between genetic changes in tumor cells and post-translational modifications (glycosylation, collagens and laminins cross-linking, mechanical events such as visco-elasticity or stiffness, which have been implicated in tumorigenesis and tumor invasion) of the ECM proteins. These modifications then significantly alter tumor and stromal cellular behavior and influence gene expression. ECM remodeling in the tumor microenvironment represents a complex process able to promote tumor growth, adhesion, motility, invasion, enhance cancer cell survival, enables metastatic dissemination, and facilitate establishment of tumor cells at distant sites [24]. In addition, tumor intracellular signal transduction pathways cross-talk with the ECM cellular and non-cellular components, as well as endothelial cells themselves and may lead to angiogenesis, which is another crucial process important for tumor progression [25].
DNA microarray-based gene expression signatures underlying oncogenic pathways have been used to define cancer subtypes, recurrence of disease, and response to specific therapies. We have recently used a genome wide expression profiling analysis, GSEA (Gene Set Enrichment Analysis), to compare the genomic signature of PTCs with and without BRAFV600E as well as normal thyroid tissue [19]. Our data indicated that out of 539 gene sets analyzed, 17 up-regulated and one down-regulated gene sets exclusively linked to the genomic signature of BRAFV600E positive PTCs [19]. The up-regulated gene sets were mainly enriched in leading edge genes known to be important in ECM remodeling process (i.e. cell adhesion, migration, invasion, and metastasis). These 17 up-regulated gene sets included genes involved in the following biological processes: “oncogenic KRAS signaling”, “cell cycle regulation”, “cell adhesion, migration, and invasion”, “cell signaling”, “inflammatory and immune mechanisms”, “EGF receptor activation”, “cytokines mechanisms”, “estrogen-receptor-dependent signal transduction”, “vesicular intracellular trafficking machinery”, “ubiquitination mechanisms”, “caspases cascade”, and “platelet aggregation” [19]. The only down-regulated gene set included genes involved in the tyrosine metabolism and in the establishment of differentiation and polarity in epithelial cells [19].
We therefore identified new gene targets of BRAFV600E in PTCs, including those involved in the regulation of tumor ECM genes expression (Figure 1). Overall, our results demonstrate that the genes listed below are positively regulated by BRAFV600E and elicit important cellular signaling cross-talks, regulate ECM remodeling, and may help explain how this single gene mutation triggers thyroid tumor cell migration and invasion thus causing a worse clinical picture for patients harboring the mutaton [19].
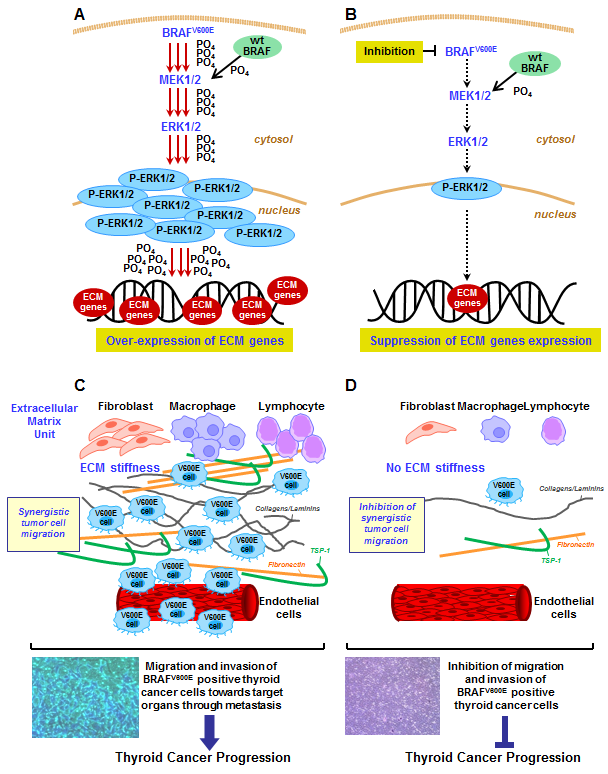
Effects of activated phospho-ERK1/2 signaling pathway by BRAFV600E in human thyroid cancer cells. (A) The BRAFV600E oncoprotein is constitutively active kinase and does not require Ras signaling to phosphorylate MEK1/2 and ERK1/2 kinases. BRAFV600E elicits a strong kinase activity (increased phosphorylation, PO4−) compared with wild type (wt) BRAF and activates MEK1/2 to phospho-MEK1/2 and ERK1/2 to phospho-ERK1/2, which may be involved in the up-regulation of some ECM remodeling genes (i.e. TSP-1, Fibronectin, cathepsins) and ECM receptors (i.e. CD44 and integrin α3, α6, and β1). ECM genes may trigger tumor cell migration and invasion, and cause thyroid cancer progression. (B) BRAFV600E knockdown down-regulates phospho-MEK1/2 and phospho-ERK1/2 protein levels and might result in inhibition of thyroid cancer progression. (C) Thyroid cancer cells harboring BRAFV600E show an increased ECM stiffness (e.g. collagen or laminin cross linking) and disrupts thyroid cell morphogenesis and polarity compared to normal thyroid tissue. This mechanism contributes to thyroid cancer progression by increasing cell adhesion, migration, invasion, and metastasis. (D) BRAFV600E knockdown results in decreased ECM stiffness and inhibition of thyroid cancer cell migration and invasion.
Some examples of genes important in ECM regulation that were discovered to be linked to BRAFV600E mutant PTCs [19] included:
TSP-1
TSP-1 is a crucial component of the ECM and important regulator of angiogenesis. It is a multifunctional matricellular protein (structured with N- and C-terminal domains, and 3 types of sequence repeats) that regulates ECM structure during tissue genesis. TSP-1 binds to a wide variety of integrins (i.e. α3, α6, and β1 involved in cellular proliferation and metastasis pathways) and non-integrin (i.e. proteoglycans) cell surface receptors, indicating its importance in cross-talk between ECM receptors. TSP-1 together with its target TGFβ1 (that was also significantly up-regulated in BRAF mutated PTC) appear to be crucial regulator of epithelial cell growth, motility, polarity and morphogenesis [26].
Integrins (α3, α6, and β1) are the major receptors for cell adhesion to the ECM, are also known to interact with TSP-1 [27] and initiate ‘outside-in’ signal transduction events that modulate gene expression, cell proliferation, migration, and invasion [28].
CD44
CD44 is a transmembrane receptor for hyaluronic acid (HA), which is implicated in various adhesion-dependent cellular processes, including tumor cell migration, invasion, and metastasis [29]. TSP-1 interacts with CD44 ligands or HA binding proteins to influence CD44 expression [30]. Interestingly, CD44 expression is deficient in TSP-1 null mice [30]. CD44 cleavage is mediated by TGFβ1 through the up-regulation of membrane type 1-matrix metalloproteinase and promotes migration of melanoma cells [31].
FN
FN is another crucial ECM component, which interacts with integrins (e.g. α5β1) to regulate cytoskeleton organization, cell shape, and proliferation in human cells [32]. Fibronectin and TSP-1 can form ternary complex and regulate ECM assembly and cell adhesion [33]. There is some evidence that changes in integrin expression can result in the regulation of secretion of cathepsin-B. The 12-lipoxygenase metabolite of arachidonic acid that can up-regulates surface expression of integrins, induces secretion of cathepsin-B from tumor cells [34].
Cathepsin-B (CTS-B) and Cathepsin–S
Cathepsin-B (CTS-B) and Cathepsin–S (CTS-S) are cysteine proteases and have been implicated in ECM remodeling, in particular in processes important for tumor development and progression, including angiogenesis, cell proliferation and invasion [35].
The main level of organization of ECM is the basement membrane that interacts directly with the epithelium and endothelium and consists primarily of collagen IV, laminins, entactin/nidogen and heparin sulphate proteoglycans. The second level of organization is the interstitial matrix, which is characterized of collagen type I and III that along with fibronectin contribute to the mechanical strength of the tissue [24]. Importantly, we found that collagen type I α1 (COL1A1) (5-fold change), collagen type III α1 (COL3A1) (2-fold change), and laminin γ2 (LAMC2) (2.45-fold change) were significantly increased in BRAFV600E positive PTCs [19]. These differentially expressed genes may be related to the carcinogenesis and progression of malignant thyroid growth. These results highlight and reinforce the critical role that BRAFV600E plays to up-regulate the expression of different ECM non-cellular components by altering stoichiometry of interactions between BRAF mutated tumor thyroid cells and ECM molecules, and ultimately promoting thyroid cancer cell migration and invasion (Figure 1).
As future research continues to focus on finding more selective and targeted compounds to truly personalize cancer care, researchers will have to be able to understand the molecular fingerprints and subsequent actions of each one of the mutations in common cancers. Thyroid cancer is clearly one of the most studied human cancers and many different genetic alterations have been studied. Studying how BRAFV600E altered kinase activity in patients with PTC creates alterations in the already complex tumor cell–ECM interactions has been a logical next step for our laboratory. We hope that our work will lead to the development of more selective targeted anticancer therapies and also biomarkers that may include putative effectors (i.e. TSP-1) that may be measurable as secreted-factors in biologic fluids. To assay candidate biomarkers that were emerged from our study will be relevant during clinical trials (e.g. using selective inhibitors of BRAFV600E) in patients with BRAFV600E positive PTC before treatment as well as during treatment as significant prognostic indicators of disease-free and overall survival.
Acknowledgements
Dr. Carmelo Nucera (M.D., Ph.D.) was a recipient of a doctorate Fellowship in Experimental Endocrinology and Metabolic Diseases (MIUR, Italy). Drs. Sareh Parangi (M.D.), Jack Lawler (PhD), and Richard A. Hodin (M.D.) were funded through the NIH Grants.
References
1. NCCN. Practice guidelines in Oncology. www.nccn.org. 2007.
2. Baudin E, Schlumberger M. New therapeutic approaches for metastatic thyroid carcinoma. Lancet Oncol. 2007; 8: 148-56.
3. Schlumberger M, Pacini F. (2003). Prognostic Factors (In:Thyroid Tumors Paris Edition Nucleon), pp. 111-25.
4. Ciampi R, Nikiforov YE. RET/PTC rearrangements and BRAF mutations in thyroid tumorigenesis. Endocrinology. 2007;148: 936-41.
5. Santoro M, Melillo RM, Carlomagno F, Fusco A, Vecchio G. Molecular mechanisms of RET activation in human cancer. Annals of the New York Academy of Sciences. 2002; 963: 116-21.
6. Giordano TJ, Kuick R, Thomas DG, Misek DE, Vinco M, Sanders D, Zhu Z, Ciampi R, Roh M, Shedden K, Gauger P, Doherty G, Thompson NW, Hanash S, Koenig RJ, Nikiforov YE. Molecular classification of papillary thyroid carcinoma: distinct BRAF, RAS, and RET/PTC mutation-specific gene expression profiles discovered by DNA microarray analysis. Oncogene. 2005; 24: 6646-56.
7. Xing M. BRAF mutation in thyroid cancer. Endocrine-related cancer. 2005; 12: 245-62.
8. Riesco-Eizaguirre G, Santisteban P. New insights in thyroid follicular cell biology and its impact in thyroid cancer therapy. Endocrine-related cancer. 2007; 14: 957-77.
9. Ricarte-Filho JC, Ryder M, Chitale DA, Rivera M, Heguy A, Ladanyi M, Janakiraman M, Solit D, Knauf JA, Tuttle RM, Ghossein RA, Fagin JA. Mutational profile of advanced primary and metastatic radioactive iodine-refractory thyroid cancers reveals distinct pathogenetic roles for BRAF, PIK3CA, and AKT1. Cancer research. 2009; 69: 4885-93.
10. Xing M. BRAF mutation in papillary thyroid cancer: pathogenic role, molecular bases, and clinical implications. Endocr Rev. 2007; 28: 742-62.
11. Nucera C, Goldfarb M, Hodin R, Parangi S. Role of B-Raf(V600E) in differentiated thyroid cancer and preclinical validation of compounds against B-Raf(V600E). Biochimica et biophysica acta. 2009; 1795: 152-61.
12. Davies H, Bignell GR, Cox C, Stephens P, Edkins S, Clegg S, Teague J, Woffendin H, Garnett MJ, Bottomley W, Davis N, Dicks E, Ewing R, Floyd Y, Gray K, Hall S, et al. Mutations of the BRAF gene in human cancer. Nature. 2002; 417: 949-54.
13. Emuss V, Garnett M, Mason C, Marais R. Mutations of C-RAF are rare in human cancer because C-RAF has a low basal kinase activity compared with B-RAF. Cancer research. 2005; 65: 9719-26.
14. Wan PT, Garnett MJ, Roe SM, Lee S, Niculescu-Duvaz D, Good VM, Jones CM, Marshall CJ, Springer CJ, Barford D, Marais R. Mechanism of activation of the RAF-ERK signaling pathway by oncogenic mutations of B-RAF. Cell. 2004; 116: 855-67.
15. Elisei R, Ugolini C, Viola D, Lupi C, Biagini A, Giannini R, Romei C, Miccoli P, Pinchera A, Basolo F. BRAF(V600E) mutation and outcome of patients with papillary thyroid carcinoma: a 15-year median follow-up study. The Journal of clinical endocrinology and metabolism. 2008; 93: 3943-9.
16. Frasca F, Nucera C, Pellegriti G, Gangemi P, Attard M, Stella M, Loda M, Vella V, Giordano C, Trimarchi F, Mazzon E, Belfiore A, Vigneri R. BRAF(V600E) mutation and the biology of papillary thyroid cancer. Endocrine-related cancer. 2008; 15: 191-205.
17. Knauf JA, Ma X, Smith EP, Zhang L, Mitsutake N, Liao XH, Refetoff S, Nikiforov YE, Fagin JA. Targeted expression of BRAFV600E in thyroid cells of transgenic mice results in papillary thyroid cancers that undergo dedifferentiation. Cancer research. 2005; 65: 4238-45.
18. Nucera C, Nehs MA, Mekel M, Zhang X, Hodin R, Lawler J, Nose V, Parangi S. A novel orthotopic mouse model of human anaplastic thyroid carcinoma. Thyroid. 2009; 19: 1077-84.
19. Nucera C, Porrello A, Antonello ZA, Mekel M, Nehs MA, Giordano TJ, Gerald D, Benjamin LE, Priolo C, Puxeddu E, Finn S, Jarzab B, Hodin RA, Pontecorvi A, Nose V, Lawler J, et al. B-Raf(V600E) and thrombospondin-1 promote thyroid cancer progression. Proceedings of the National Academy of Sciences of the United States of America. 2010; 107: 10649-54.
20. Mesa C, Jr., Mirza M, Mitsutake N, Sartor M, Medvedovic M, Tomlinson C, Knauf JA, Weber GF, Fagin JA. Conditional activation of RET/PTC3 and BRAFV600E in thyroid cells is associated with gene expression profiles that predict a preferential role of BRAF in extracellular matrix remodeling. Cancer research. 2006; 66: 6521-9.
21. Chou CK, Chen RF, Chou FF, Chang HW, Chen YJ, Lee YF, Yang KD, Cheng JT, Huang CC, Liu RT. miR-146b is highly expressed in adult papillary thyroid carcinomas with high risk features including extrathyroidal invasion and the BRAF(V600E) mutation. Thyroid. 2010; 20: 489-94.
22. Romei C, Ciampi R, Faviana P, Agate L, Molinaro E, Bottici V, Basolo F, Miccoli P, Pacini F, Pinchera A, Elisei R. BRAFV600E mutation, but not RET/PTC rearrangements, is correlated with a lower expression of both thyroperoxidase and sodium iodide symporter genes in papillary thyroid cancer. Endocrine-related cancer. 2008; 15: 511-20.
23. Mitsutake N, Knauf JA, Mitsutake S, Mesa C, Jr., Zhang L, Fagin JA. Conditional BRAFV600E expression induces DNA synthesis, apoptosis, dedifferentiation, and chromosomal instability in thyroid PCCL3 cells. Cancer research. 2005; 65: 2465-73.
24. Erler JT, Weaver VM. Three-dimensional context regulation of metastasis. Clinical & experimental metastasis. 2009; 26: 35-49.
25. Folkman J, Watson K, Ingber D, Hanahan D. Induction of angiogenesis during the transition from hyperplasia to neoplasia. Nature. 1989; 339: 58-61.
26. Adams JC, Lawler J. Cell-type specific adhesive interactions of skeletal myoblasts with thrombospondin-1. Mol Biol Cell. 1994; 5: 423-37.
27. Chen H, Herndon ME, Lawler J. The cell biology of thrombospondin-1. Matrix Biol. 2000; 19: 597-614.
28. Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell. 2002; 110: 673-87.
29. Naor D, Wallach-Dayan SB, Zahalka MA, Sionov RV. Involvement of CD44, a molecule with a thousand faces, in cancer dissemination. Seminars in cancer biology. 2008; 18: 260-7.
30. Kuznetsova SA, Day AJ, Mahoney DJ, Rugg MS, Mosher DF, Roberts DD. The N-terminal module of thrombospondin-1 interacts with the link domain of TSG-6 and enhances its covalent association with the heavy chains of inter-alpha-trypsin inhibitor. The Journal of biological chemistry. 2005; 280: 30899-908.
31. Kuo YC, Su CH, Liu CY, Chen TH, Chen CP, Wang HS. Transforming growth factor-beta induces CD44 cleavage that promotes migration of MDA-MB-435s cells through the up-regulation of membrane type 1-matrix metalloproteinase. International journal of cancer. 2009; 124: 2568-76.
32. Hynes RO. The extracellular matrix: not just pretty fibrils. Science. 2009; 326: 1216-9.
33. Rodrigues RG, Guo N, Zhou L, Sipes JM, Williams SB, Templeton NS, Gralnick HR, Roberts DD. Conformational regulation of the fibronectin binding and alpha 3 beta 1 integrin-mediated adhesive activities of thrombospondin-1. The Journal of biological chemistry. 2001; 276: 27913-22.
34. Koblinski JE, Ahram M, Sloane BF. Unraveling the role of proteases in cancer. Clin Chim Acta. 2000; 291: 113-35.
35. Gocheva V, Zeng W, Ke D, Klimstra D, Reinheckel T, Peters C, Hanahan D, Joyce JA. Distinct roles for cysteine cathepsin genes in multistage tumorigenesis. Genes Dev. 2006; 20: 543-56.