
INTRODUCTION
Aging is a physiological process associated with progressive structural and functional declines of tissues and organs [1]. The hair follicle is a mini-organ that undergoes repetitive cyclic regeneration under physiological conditions during postnatal life, thus supplying an excellent model for aging-associated disorders. Typical hair follicle aging phenotypes can be observed but not limited to several signs, such as irreversible hair loss, hair thinning and graying [2].
Regenerative hair cycling process in single hair follicle consists of three consecutive phases including growth phase (anagen), regression phase (catagen) and resting phase (telogen) [3-7]. Hair stem cell activation during telogen to anagen transition is mainly controlled by two reciprocal out of phases mechanisms. These include Wnt/β-catenin signaling pathway, which shows crucially roles in hair regeneration [8]. The other one is Bmp signaling pathway, which is decreased in competent telogen phase compared to the refractory telogen phase, leading to hair regeneration [9]. Melanocyte stem cells share the same niche with hair follicle stem cells. Progress has been making in unveiling regenerative behaviors and differentiation of melanocytes [10, 11]. Melanocyte stem cells are activated coordinately with hair follicle stem cells during hair regeneration. They migrate out from the bulge niche to the hair matrix region, and differentiate into melanocytes which generate melanin to pigment hairs.
There is increasing evidence showing that many morphogenetic pathways play key roles in regulating melanocytes behaviors. Of these, Wnt signaling functions as an important pathway controlling the patterning of melanocytes and influencing the decisions of melanocyte stem cells differentiation to pigment the hairs [12, 13]. Wnt3a induces melanocyte stem cell differentiation in vivo and in vitro [14]. Exogenous Wnt recruits β-catenin and Lef1 to bind the promoter of microphthalmia-associated transcription factor (MITF), which functions as a key gene that governs fates of melanocyte lineage cells [15]. Previous study shows that one of the visible signs of hair follicle aging is hair loss, which is due to decreased expression of Collagen XVII, causing the hair follicle stem cells differentiate into the epidermal lineages [2]. However, the mechanism of hair graying as the other obvious sign of hair follicle aging remains further investigation. Whether Wnt signaling acts as a positive or negative regulator in hair follicle aging is unclear.
Therefore, in this study, we first compared hair graying phenotype in young and adult mice. Since the important role of Wnt signaling in aging of other tissues, we examined periodic expression of β-catenin which is the effector of Wnt signaling pathway, in melanocyte lineage cells during hair cycling in Dct-Lac-Z CD1 transgenic mice. We also compared β-catenin expression at telogen phase and anagen phase skin in young mice and 34-month-old aged mice. To explore the function of Wnt signaling on melanocyte differentiation, we over expressed Wnt10b through adenovirus-mediated expression in vivo or in vitro, through intracutaneous injection of adenovirus into the young adult skin, or by adding them into iMC23 melanocyte stem cells, respectively. Our study indicates that Wnt signaling promotes differentiation of melanocyte stem cell, exhaustion of which leads to hair graying during aging.
RESULTS
Aging mice exhibit increased gray hairs
To study the influence of aging on hair pigmentation in mice, we first counted and compared the number of black and gray hairs in dorsal back skin of C57Bl/6 mice aged at postnatal day 23 (23d, first telogen), 35d (second anagen) and 34 months (34M) (Figure 1A-1C). We observed that many hairs turn gray in mice aged at 34M, when compared to the young mice aged at 23d (Figure 1A). While the hairs in 23d young mice are black, the hair fiber completely turns gray from the distal part to the proximal part in 34M old mice (Figure 1B). Statistical analysis reveals that there are significantly more gray hairs in 34M old mice when compared to the 23d and 35d old mice (Figure 1C).
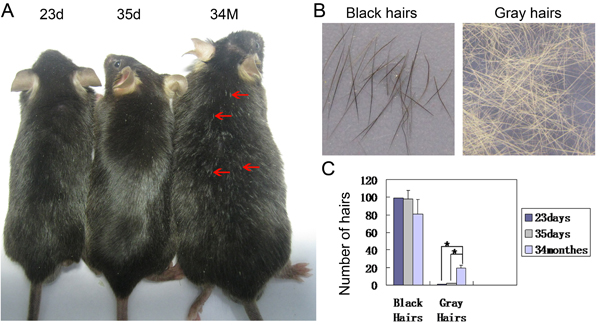
Figure 1: Hairs turn gray during aging in mice. A. Partial hairs in the dorsal back skin turn gray in aged mice. B. Hairs plucked from young mice are black and old mice are gray. C. Statistical analysis reveals that significantly increased gray hairs in dorsal back skin in aged mice. 23d, postnatal 23 day; 34M, postnatal 34 months. N = 24.
β-catenin is expressed in melanocyte cell lineages
Wnt/β-catenin signaling is important for proliferation and differentiation of melanocytes. Previous study shows a cyclic expression pattern of β-catenin during hair cycling in C57BL/6 mouse strain [4]. Consistent with those studies, RT-PCR, western blot and their statistical analysis show that β-catenin is highly expressed in early and mid-anagen hair follicles (4d, 8d and 29d), but is decreased in catagen (18d) and telogen (23d) skin in Dct-LacZ transgenic CD1 mice (Figure 2A-2D).
To evaluate the expression of β-catenin in melanocyte cell lineages, we immunostained β-catenin expression in the skin of Dct-LacZ mouse, in which all melanocyte cell lineages are labeled by β-gal. This allows us to visualize LacZ+ melanocytes lineage cells, including melanoblasts, melanocyte stem cells, differentiating melanocytes, and mature melanocytes in the hair follicle. Translocation of β-catenin from cytoplasm to the nucleus indicates the activation of Wnt/β-catenin signaling pathway. We observed that nuclear β-catenin was only present in the early and mid-anagen phase of hair follicle. Close-up view reveals that nuclear β-catenin is expressed in the melanocytes located at hair matrix and secondary hair germ (Shg) region, and a few in the outer root sheath (Figure 2E). Combined with previous studies, our results further suggest that β-catenin regulates behaviors of melanocyte cell lineage during hair regeneration and growth.
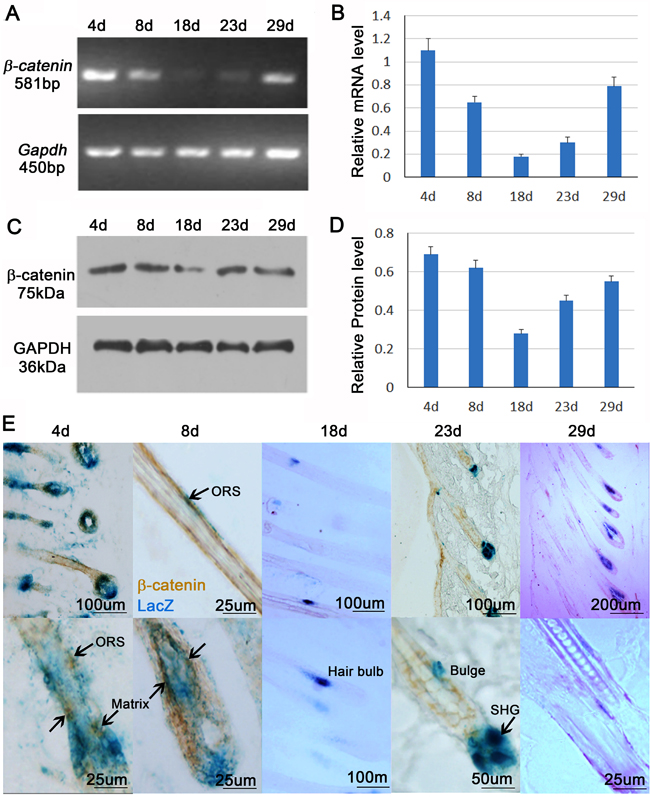
Figure 2: β-catenin expression in the melanocytes. A. RT-PCR shows mRNA expression of β-catenin during hair cycling. B. Statistical analysis reveals that mRNA expression β-catenin is sig nificantly increased in anagen phase when compared to catagen and telogen phase. C. Western Blot shows protein expression of β-catenin during hair cycling. D. Statistical analysis reveals that protein expression of β-catenin is significantly increased in anagen phase when compared to catagen and telogen phase. E. β-catenin immunostaining and LacZ counterstaining show that nuclear β-catenin expression in melanocytes. ORS, outer root sheath; SHG, secondary hair germ. N = 12.
β-catenin expression is increased is skin of aged mice
Several lines of evidence show that aging of some tissues and organs is related to the activation of Wnt/β-catenin signal pathway [16-18]. To explore the changes of β-catenin expression in skin during aging, we harvested dorsal back skin from both young and old Dct-LacZ CD1 mice (Figure 3A). We also compared skin samples at anagen or telogen phase of hair cycle by RT-PCR and western blot (Figure 3B-3C). Statistical analysis reveals that β-catenin expression was significantly increased both at 34 month telogen (34MT) skin and 34 month anagen (34MA) skin in aged mice, when compared to the young mice aged at 23d (telogen skin) and 35d (anagen skin), respectively (Figure 3D). Immunostaining further confirms that β-catenin expression is increased in both 34MT and 34MA skin when compared to the young mice skin aged at 23d and 35d, respectively. We observed that β-catenin expression is not only increased hair follicles of aged mice, but also increased at the dermal microenvironment.
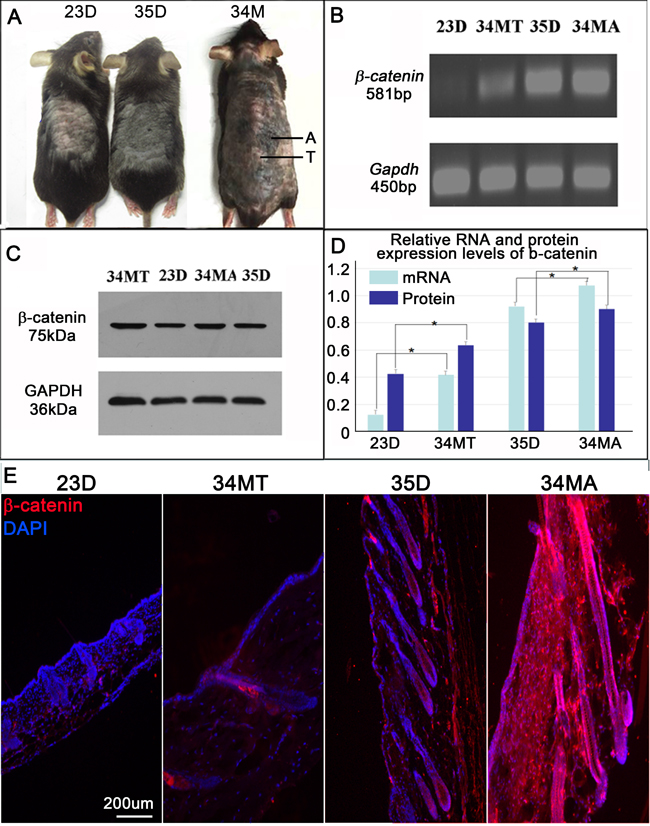
Figure 3: β-catenin expression is increased skin of aged mice. A. Mice at different age. A, anagen; T, telogen. B. RT-PCR shows mRNA expression of β-catenin in mouse skin at different age. C. Western blot shows mRNA expression of β-catenin in mouse skin at different age. D. Statistical analysis reveals that mRNA and protein expressions of β-catenin are significantly increased in skin of aged mice, when compared to skin of young mice. E. Immunostaining shows β-catenin expression is highly expressed in anagen skin of aged mice. D, day; MT, month telogen; MA; month anagen. N = 12.
Overexpression of Wnt10b promotes anagen reentry with increased number of melanocytes in young mice in vivo
We previously showed that Wnt10b can induce hair regeneration [19]. Overexpression of Wnt10b also promotes more pigmentation of hair follicles [20]. To explore if Wnt signaling promotes melanocyte differentiation in hair follicles, we overexpressed Wnt10b by intradermal injection of adenovirus-mediated overexpression of Wnt10b (AdWnt10b) or control (AdGFP) into C57BL/6 mice at postnatal day 56 (P56), when the hair follicles are all at telogen phase. Consistent with previous studies done in C57BL/6 normal mouse [19], our result shows that hair follicles enter anagen phase represented by the black region where was injected with AdWnt10b, when compared to the control group which was injected with AdGFP and shows pink color that represents the telogen phase (Figure 4A). We also did this assay in Dct-LacZ mice, which allow us to better visualize melanocyte cell behavior after adenovirus treatment. Overexpression of Wnt10b leads to increased expression of β-catenin in skin epidermis and hair follicles, when compared to the AdGFP-treated control group, which shows little β-catenin expression (Figure 4B). β-gal staining reveals that the number of melanocyte lineage cells was largely increased in AdWnt10b-treated group, particularly in the hair follicle and dermis (Figure 4B). We further used β-gal staining counterstained with eosin to show melanocytes behavior during AdWnt10b-induced hair regeneration. The result shows that the number of melanocytes is increased at the secondary hair germ region of AdWnt10b-treated group, in which hair follicle starts to enter anagen phase (Figure 4C). The hair follicles progress to full anagen at 12 day and telogen at 21 days after AdWnt10b treatment. Whereas the hair follicles in AdGFP-treated group remain at telogen phase at corresponding time points paralleling to the AdWnt10b-treated group. When entering anagen phase, hair follicles treated with AdWnt10b show stronger and some nuclear β-catenin expression in melanocytes in the secondary hair germ and hair matrix region, when compared to the control group that remains at telogen phase with low expression of β-catenin (Figure 4D).
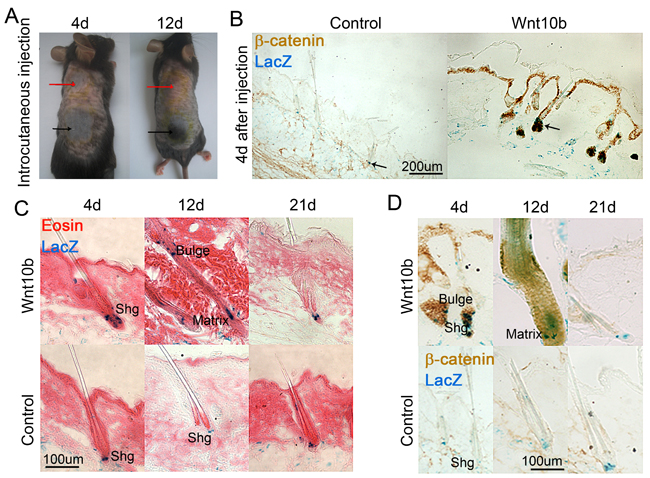
Figure 4: Overexpression of Wnt signaling promotes anagen reentry and melanocyte differentiation in vivo. A. Intracutaneous injection of AdWnt10b results in accelerated hair regeneration in mice. B. Immunostaining and LacZ counterstaining show increased β-catenin expression in epidermis and hair follicles, and melanocytes differentiation in hair follicle and dermis, respectively, in AdWnt10b-overexpressed mice when compared to the AdGFP control group. C. Eosin and LacZ double staining shows that overexpression Wnt10b leads to increased melanocytes in secondary hair germ (Shg) region and hair matrix region. D. Immunostaining and LacZ counterstaining show nuclear β-catenin expression in melanocytes in AdWnt10b-overexpressed mice at secondary hair germ region and hair matrix, whereas the AdGFP control group shows no nuclear β-catenin expression. N = 12.
Overexpression of Wnt10b promotes melanocyte progenitor cell differentiation in vitro
To further confirm the role of Wnt/β-catenin in melanocyte differentiation, we treated melanocyte progenitor cells (iMC23 cells) with AdWnt10b in vitro. To reduce the toxicity of adenovirus on iMC23 cells, we first infected epidermal cells (JB6 cells) with AdWnt10b or AdGFP (control), and harvested the supernatant which was then added to the cultured iMC23 cells (Figure 5A). Robust green fluorescence produced by JB6 cells indicates successful infection of adenovirus to the cells (Figure 5B). RT-PCR and statistical analysis reveal that mRNA expression of β-catenin was significantly increased in iMC23 cells which are cultured by Wnt10b-containing medium for 12 hours, when compared to that of the control group (Figure 5C-5D). We also analyzed melanin production after the cells were cultured by Wnt10b-containing medium for 48 hours. The melanin production can be directly observed through the pigmented cell pellets after the iMC23 cells were digested and centrifugated to the bottom of the tube. The melanin formation was significantly increased in Wnt10b-treated iMC23 cells indicated by the black pellet, when compared to the control group in which showing gentle melanogenesis (Figure 5E). Moreover, the tyrosinase activity assay reflecting melanogenesis was used to test the influence of Wnt signaling on melanocyte differentiation. Tyrosinase activity is significantly increased in Wnt10b-treated group compared to the control group (Figure 5F). These data further suggest that Wnt10b/β-catenin signaling promotes melanocyte differentiation.
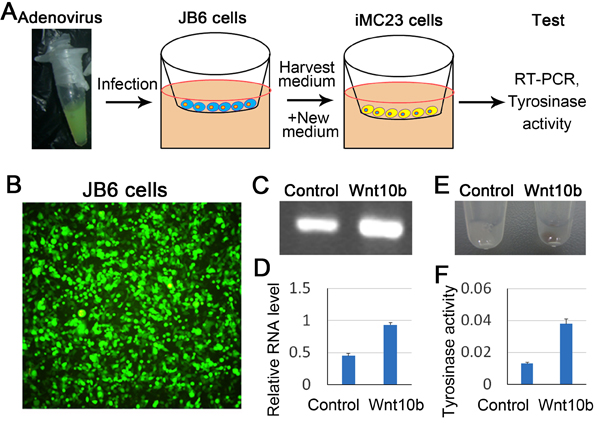
Figure 5: Overexpression of Wnt signaling promotes melanocyte stem cell differentiation in vitro. A. Model of overexpression Wnt10b in iMC23 cells. B. Green fluorescence shows successful infection of adenovirus into JB6 cells. C. RT-PCR shows mRNA expression of β-catenin after Wnt10b treatment. D. Statistical analysis reveals that mRNA expression β-catenin is significantly increased in Wnt10b treated group compared to the control group. E. Melanogenesis after Wnt10b treatment. F. Tyrosinase activity assay of iMC23 cells was performed 2 days after the cells were treated by Wnt10b. N = 12.
DISCUSSION
Tissue aging is characterized by decline of regenerative ability. Previous studies show that loss of function or incomplete maintenance of stem cells lead to compromised tissue regeneration during aging [21]. Hair follicle stands at an idea model to study age-associated tissue regeneration, due to its cyclic behavior in postnatal lifetime [22-25]. Canities is an obvious sign of aging in mammals such as mouse and human, resulting in progressive hair graying during their aging process. Fundamental mechanisms controlling hair graying involve multiple possibilities. One is melanocyte stem cells are exhausted because of high activators that promote melanocyte stem cell differentiation [21]. The other is melanocyte stem cells sharing the same niche with hair follicle stem cells at bulge become silenced due to increased expression of inhibitors during aging process [26]. Though there may be other possibilities such as senescence-caused loss of function or terminal differentiation of stem cells [2].
Here, we found that β-catenin as the effector of Wnt signaling pathway was significantly increased in both telogen and anagen of aged mice skin, when compared to telogen and anagen of young mice skin, respectively. Particularly, β-catenin was expressed at the nucleus of some melanocytes during hair regeneration and growth, indicating the activation of Wnt signaling in these cells. Consistent with previous studies [12, 13], our study provide evidence that Wnt signaling promotes melanocytes differentiation. Moreover, our findings suggest that increased β-catenin expression in skin of adult mice may result in exhaustion of melanocyte stem cells in hair follicles, leading to hair graying during aging process (Figure 6).
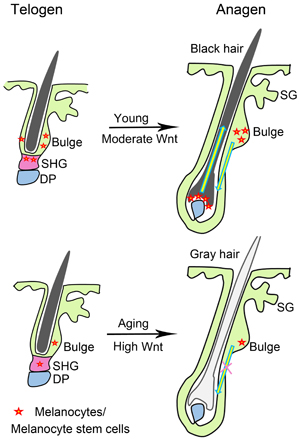
Figure 6: Schematic of Wnt signaling and hair graying. Moderate Wnt signaling allows anagen reentry during hair regeneration, and promotes differentiation of melanocytes in hair follicle of young mice. High Wnt signaling promotes excessive differentiation of melanocytes, leading to decreased melanocyte stem cells which give rise to progressive hair graying during aging. DP, dermal papilla; SG, sebaceous gland; SHG, secondary hair germ.
Previous studies show that aging of tissues and organs is related to the classical Wnt/β-catenin signal pathway. However, it seems that Wnt signaling plays dual roles in contributing to aging among different tissues. Several studies demonstrate that Wnt signaling promotes aging of cells or tissues [16-18]. For example, increased Wnt signaling in klotho mice leads to decreased number of stem cell population and increased number of aged cells in the skin and small intestine, resulting in altered muscle stem cell fate and increased fibrosis [16]. Wnt signaling induces aging of mesenchymal stem cells through regulating p53/p21 pathway [18]. Whereas other studies show that Wnt signaling ameliorates aging of intestinal stem cells [27], neuronal tissue and skeletal muscle [28].
Wnt signaling usually maintains at a low level which is under the threshold for hair follicle stem cell activation at telogen [29]. Overexpression of β-catenin in epidermis induces ectopic hair follicle regeneration [30], and even tumorigenesis [31]. In our study, we found β-catenin expression is increased in telogen skin of aged mice compared to those in young mice. Increased expression of β-catenin should shorten the telogen phase, meaning an accelerated telogen to anagen reentry of the hair follicles. However, previous study shows that the length of telogen phase is gradually increased in duration during aging in mice, and this is because of decreased activators such as Follistatin and increased inhibitors such as Bmp2, Dkk1 and Sfrp4 that are periodically expressed by the extra-follicular macro-environment [26]. With increasing inhibitors expressed in skin during aging process, activators can also be induced through the reaction-diffusion (activator-inhibitor) mechanism. Thus, we suppose that the increased level of β-catenin in telogen hair follicle, particularly in the melanocytes stem cells or progenitor cells may contribute to melanocyte stem cell differentiation in the bulge niche, resulting in decreased number of melanocyte stem cells. Whereas the expression level of β-catenin is insufficient to counterbalance the increased inhibitors, leading to the inactivation of hair follicle stem cells and a longer telogen phase in the aged skin. In addition, different from telogen-anagen transition which is a collective behavior in the skin, hair graying in both mice and human tends to be an individual behavior, with gray hairs regeneration in a progressive process. We speculate that there is a differential expression level of β-catenin and its inhibitors in the bulge region of each hair follicle in the aged skin, leading to the progressive hair graying during aging in animal.
Indeed, previous study suggests that Wnt/ β-catenin signaling has distinct roles of in determine melanocyte stem cell fate [12]. Low level of Wnt/ β-catenin signaling is required to maintain the undifferentiated state of melanocyte stem cells, whereas high Wnt/ β-catenin signaling promotes melanocytes differentiation. This is controlled by regulation between β-catenin, Pax3 and MITF [32]. Hence, in anagen hair follicle of aged skin, increased Wnt/β-catenin signaling further promotes melanocytes terminal differentiation, leading to this lineage be exhausted. Our study further supports this hypothesis. Overexpression of Wnt10b not only accelerates hair follicles to enter anagen phase, but also promotes differentiation of melanocytes in vivo. Moreover, overexpression of Wnt10b in iMC23 cells promotes melanocyte stem cell differentiation, as indicated by melanogenesis and tyrosinase activity assays. Nevertheless, the exact β-catenin expression level that balances hair stem cell activation and melanocyte stem cell differentiation remains further investigation. Future study will evaluate if decrease of Wnt signaling can alleviate exhaustion of melanocyte stem cells in aged mice.
Since Wnt/β-catenin is also expressed in other regions of hair follicles where melanocytes were absent. For example, Wnt/β-catenin is expressed in the other regions of hair bulb as well as in the DP where melanocytes are absent. How does Wnt/β-catenin signaling coordinate the cellular behavior of other epithelial cell differentiation and melanocytes differentiation in the hair follicle? First, different regions express different Wnt receptors, which can sense different Wnt ligand to regulate cellular behaviors such as proliferation and differentiation. Second, Wnt signaling is also regulated by other signals which have differential expression in different regions of hair follicle; Third, epithelial-mesenchymal interaction also functions as a critical mechanism in regulating cellular behaviors in the hair follicle [33]. Mesenchymal cells may constitute micro-niches with different epithelial cell lineages, resulting in different epithelial-mesenchymal interactions [34].
In summary, our results demonstrate that β-catenin expression is increased in hair follicles undergoing aging, compared to the hair follicle in young mice at the corresponding cycle stages. Increased Wnt signaling is insufficient to induce hair regeneration but may promote melanocyte stem cell differentiation in hair follicle bulge niche, leading to the exhaustion of melanocyte stem cells during aging process (Figure 6). Our study provides a potential mechanism to modulate expression levels of activator or inhibitor in the skin microenvironment and macroenvironment, aiding in the development of a strategy for the treatment aging-associated degenerative disorders, such as canities.
MATERIALS AND METHODS
Mice and skin samples
Dct-LacZ CD1 transgenic mice were gifted from Professor Lan Jackson at the MRC Laboratory (UK). Animals were maintained in a SPF room at the Experimental Animal Center of the Third Military Medical University. For examining the expression of β-catenin, the dorsal back skins were harvested from the male mice at postnatal 4d (P4, early anagen of the first hair cycle), 8d (P8, mid-anagen of the first hair cycle), 18d (P18, catagen of the first hair cycle), 23d (P23, telogen of the first hair cycle) and 29d (P29, early anagen of the second hair cycle). Intracutaneous injection of adenovirus was performed as previously described [3, 19].
Cell culture and tyrosinase activity assay
JB6 cells (ATCC, USA) were cultured in DMEM culture medium with 10% fetal bovine serum, in an atmosphere containing 5% CO2 in air at 37°C. Adenoviruses were generated and the supernatant of adenovirus-infected cells was harvested as previously described [35]. Then the supernatant was added to the iMC23 cells [36], which were cultured under the same condition as JB6 cells. For tyrosinase activity assay, iMC23 cells were treated with 1% TritonX-100/PBS for 30min and centrifuged. 50 μL supernatant was added with 10μL 2mg/ml L-DOPA and incubated at 37°C for 2 hours. The sample was measured at 480nm for absorbance.
Reverse transcription and polymerase chain reaction (RT-PCR)
Total RNA was extracted from the skin specimens per the instruction for use of Trizol reagent. RT-PCR was performed per the instruction for use of the reverse transcription kit (TOYOBO, Japan) and PCR MIX kit (Tiangen, China). The primers for β-catenin were designed as: 5’-ATCACTGAGCCTGCCATCTG-3’ (sense); 5’-GTTGCCACGCCTTCATTCC -3’ (antisense), with a 581bp product in length. The annealing temperature was set at 63°C.
Western-blot
Proteins were extracted from the skin specimens by using of RIPA lysis buffer (Beyontime, China). The extracts were vortexed, centrifuged at 12000×g for 10min. The concentration was determined by using a BCA protein concentration determination kit (Beyontime, China). Then the extracts were added with the loading buffer and incubated in boiling water for 5 min, followed by electrophoresis (concentrating gel 80V, 30 min; separating gel 100V, 90 min). The proteins were transferred to a PVDF membrane (250 mA, 2 h), which was treated with methanol for 1 min and electrotransfer buffer for 15 min before use. After wash with TBST (5min × 5), the PVDF membrane was blocked with 5% defatted milk (2 h), incubated with rabbit anti-β-catenin polyclonal antibody (1:1000; Abcam, USA) at 4°C for overnight, and secondary antibody (1:10000; Boster, China) at 37°C for 1 h, then developed with chemoluminescence.
β-gal staining and immunohistochemistry
The back skins from Dct-Lac-Z CD1 transgenic mice were fixed at 4°C in 4% paraformaldehyde for 1 h, and stained with x-gal (Beyotime, China) for 24 h. After wash, the skin specimens were embedded in paraffin and sliced into 5 μm sections, which were incubated with rabbit anti-β-catenin polyclonal antibody (1:600; Abcam, USA) at 4°C for overnight, and the secondary antibody (Zhongshan, China) at 37oC for 1h. The samples were developed with DAB (Zhongshan, China).
Data acquisition and statistical analysis
RT-PCR and Western blot images were analyzed with Quantity One software to acquire the optical density of the bands. The relative expression level was normalized at the ratio of optical density of target band against that of internal control band. All experiments in this study were repeated at least three times. Data were expressed as x̄±S and One-way ANOVA of data using SPSS10.0 software.
Author contributions
Conception and design: FD, XL, TY, ZZ
Development of methodology: FD
Acquisition of data: YX, YL, HG, HX, CH, ZZ
Analysis and interpretation of data: FD, ML
Writing, review, and/or revision of the manuscript: FD, ML
ACKNOWLEDGMENTS
We thank Prof. Ian Jackson (MRC Human Genetics Unit) for the Dcr-LacZ transgenic mice.
CONFLICTS OF INTEREST
The authors declare that no conflicts of interest exist.
FINANCIAL SUPPORT
This work was supported by research grants from the National Natural Science Foundation of China (81271770 and 81502371 to TY and FD) and Chongqing Science and Technology Commission (cstc2016jcyjA0333 to FD). Mingxing Lei is supported by projects funded by China Postdoctoral Science Foundation (2016M590866), Fundamental Research Funds for the Central Universities (106112015CDJRC231206), Special Funding for Postdoctoral Research Projects in Chongqing (Xm2015093), and a fellowship from the China Scholarship Council (2011605042).
REFERENCES
1. Lei M, Chuong CM. STEM CELLS. Aging, alopecia, and stem cells. Science. 2016; 351: 559-60. doi: 10.1126/science.aaf1635.
2. Matsumura H, Mohri Y, Binh NT, Morinaga H, Fukuda M, Ito M, Kurata S, Hoeijmakers J, Nishimura EK. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science. 2016; 351: aad4395. doi: 10.1126/science.aad4395aad4395.
3. Bai XF, Lei MX, Shi JZ, Yu Y, Qiu WM, Lai XD, Liu YX, Yang T, Yang L, Widelitz RB, Chuong CM, Lian XH. Roles of GasderminA3 in Catagen-Telogen Transition during Hair Cycling. Journal of Investigative Dermatology. 2015; 135: 2162-72. doi: 10.1038/jid.2015.147.
4. Lei M, Yang T, Lai X, Bai X, Qiu W, Lian X, Yang L. Upregulation of interfollicular epidermal and hair infundibulum beta-catenin expression in Gsdma3 mutant mice. Acta Histochem. 2013; 115: 63-9. doi: 10.1016/j.acthis.2012.04.002.
5. Chueh SC, Lin SJ, Chen CC, Lei MX, Wang LM, Widelitz R, Hughes MW, Jiang TX, Chuong CM. Therapeutic strategy for hair regeneration: hair cycle activation, niche environment modulation, wound-induced follicle neogenesis, and stem cell engineering. Expert Opinion on Biological Therapy. 2013; 13: 377-91. doi: 10.1517/14712598.2013.739601.
6. Lei M, Bai X, Yang T, Lai X, Qiu W, Yang L, Lian X. Gsdma3 is a new factor needed for TNF-alpha-mediated apoptosis signal pathway in mouse skin keratinocytes. Histochem Cell Biol. 2012; 138: 385-96. doi: 10.1007/s00418-012-0960-1.
7. Lei M, Gao X, Yang L, Yang T, Lian X. Gsdma3 gene is needed for the induction of apoptosis-driven catagen during mouse hair follicle cycle. Histochem Cell Biol. 2011; 136: 335-43. doi: 10.1007/s00418-011-0845-8.
8. Lei MX, Chuong CM, Widelitz RB. Tuning Wnt Signals for More or Fewer Hairs. Journal of Investigative Dermatology. 2013; 133: 7-9. doi: 10.1038/jid.2012.446.
9. Plikus MV, Mayer JA, de la Cruz D, Baker RE, Maini PK, Maxson R, Chuong CM. Cyclic dermal BMP signalling regulates stem cell activation during hair regeneration. Nature. 2008; 451: 340-4. doi: 10.1038/nature06457.
10. Liao CP, Booker RC, Morrison SJ, Le LQ. Identification of hair shaft progenitors that create a niche for hair pigmentation. Genes Dev. 2017; 31: 744-56. doi: 10.1101/gad.298703.117.
11. Chang CY, Pasolli HA, Giannopoulou EG, Guasch G, Gronostajski RM, Elemento O, Fuchs E. NFIB is a governor of epithelial-melanocyte stem cell behaviour in a shared niche. Nature. 2013; 495: 98-102. doi: 10.1038/nature11847.
12. Lim X, Nusse R. Wnt signaling in skin development, homeostasis, and disease. Cold Spring Harb Perspect Biol. 2013; 5. doi: 10.1101/cshperspect.a008029.
13. Rabbani P, Takeo M, Chou W, Myung P, Bosenberg M, Chin L, Taketo MM, Ito M. Coordinated activation of Wnt in epithelial and melanocyte stem cells initiates pigmented hair regeneration. Cell. 2011; 145: 941-55. doi: 10.1016/j.cell.2011.05.004.
14. Guo H, Lei M, Li Y, Liu Y, Tang Y, Xing Y, Deng F, Yang K. Paracrine Secreted Frizzled-Related Protein 4 Inhibits Melanocytes Differentiation in Hair Follicle. Stem Cells Int. 2017; 2017: 2857478. doi: 10.1155/2017/2857478.
15. Dorsky RI, Raible DW, Moon RT. Direct regulation of nacre, a zebrafish MITF homolog required for pigment cell formation, by the Wnt pathway. Genes Dev. 2000; 14: 158-62.
16. Brack AS, Conboy MJ, Roy S, Lee M, Kuo CJ, Keller C, Rando TA. Increased Wnt signaling during aging alters muscle stem cell fate and increases fibrosis. Science. 2007; 317: 807-10. doi: 10.1126/science.1144090.
17. Narita M, Nunez S, Heard E, Narita M, Lin AW, Hearn SA, Spector DL, Hannon GJ, Lowe SW. Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell. 2003; 113: 703-16.
18. Zhang DY, Wang HJ, Tan YZ. Wnt/beta-catenin signaling induces the aging of mesenchymal stem cells through the DNA damage response and the p53/p21 pathway. PLoS One. 2011; 6: e21397. doi: 10.1371/journal.pone.0021397.
19. Lei MX, Guo HY, Qiu WM, Lai XD, Yang T, Widelitz RB, Chuong CM, Lian XH, Yang L. Modulating hair follicle size with Wnt10b/DKK1 during hair regeneration. Experimental Dermatology. 2014; 23: 407-13. doi: 10.1111/exd.12416.
20. Ye J, Yang T, Guo H, Tang Y, Deng F, Li Y, Xing Y, Yang L, Yang K. Wnt10b promotes differentiation of mouse hair follicle melanocytes. Int J Med Sci. 2013; 10: 691-8. doi: 10.7150/ijms.6170.
21. Nishimura EK, Granter SR, Fisher DE. Mechanisms of hair graying: incomplete melanocyte stem cell maintenance in the niche. Science. 2005; 307: 720-4. doi: 10.1126/science.1099593.
22. Qiu W, Tang H, Guo H, Lei M, Yan H, Lian X, Wu J. 12-O-tetradecanoylphorbol-13-acetate activates hair follicle melanocytes for hair pigmentation via Wnt/beta-catenin signaling. Cell Tissue Res. 2016; 366: 329-40. doi: 10.1007/s00441-016-2450-6.
23. Qiu W, Lei M, Tang H, Yan H, Wen X, Zhang W, Tan R, Wang D, Wu J. Hoxc13 is a crucial regulator of murine hair cycle. Cell Tissue Res. 2016; 364: 149-58. doi: 10.1007/s00441-015-2312-7.
24. Qiu W, Yang K, Lei M, Yan H, Tang H, Bai X, Yang G, Lian X, Wu J. SCF/c-kit signaling is required in 12-O-tetradecanoylphorbol-13-acetate-induced migration and differentiation of hair follicle melanocytes for epidermal pigmentation. Cell Tissue Res. 2015; 360: 333-46. doi: 10.1007/s00441-014-2101-8.
25. Wang N, Yang T, Li J, Lei M, Shi J, Qiu W, Lian X. The expression and role of c-Myc in mouse hair follicle morphogenesis and cycling. Acta Histochem. 2012; 114: 199-206. doi: 10.1016/j.acthis.2011.04.009.
26. Chen CC, Murray PJ, Jiang TX, Plikus MV, Chang YT, Lee OK, Widelitz RB, Chuong CM. Regenerative Hair Waves in Aging Mice and Extra-Follicular Modulators Follistatin, Dkk1, and Sfrp4. Journal of Investigative Dermatology. 2014; 134: 2086-96. doi: 10.1038/jid.2014.139.
27. Nalapareddy K, Nattamai KJ, Kumar RS, Karns R, Wikenheiser-Brokamp KA, Sampson LL, Mahe MM, Sundaram N, Yacyshyn MB, Yacyshyn B, Helmrath MA, Zheng Y, Geiger H. Canonical Wnt Signaling Ameliorates Aging of Intestinal Stem Cells. Cell Rep. 2017; 18: 2608-21. doi: 10.1016/j.celrep.2017.02.056.
28. Fujimaki S, Wakabayashi T, Takemasa T, Asashima M, Kuwabara T. The regulation of stem cell aging by Wnt signaling. Histol Histopathol. 2015; 30: 1411-30. doi: 10.14670/HH-11-657.
29. Lim X, Tan SH, Yu KL, Lim SB, Nusse R. Axin2 marks quiescent hair follicle bulge stem cells that are maintained by autocrine Wnt/beta-catenin signaling. Proc Natl Acad Sci U S A. 2016; 113: E1498-505. doi: 10.1073/pnas.1601599113.
30. Collins CA, Kretzschmar K, Watt FM. Reprogramming adult dermis to a neonatal state through epidermal activation of beta-catenin. Development. 2011; 138: 5189-99. doi: 10.1242/dev.064592.
31. Qiu W, Lei M, Li J, Wang N, Lian X. Activated hair follicle stem cells and Wnt/beta-catenin signaling involve in pathnogenesis of sebaceous neoplasms. Int J Med Sci. 2014; 11: 1022-8. doi: 10.7150/ijms.8383.
32. Lang D, Lu MM, Huang L, Engleka KA, Zhang M, Chu EY, Lipner S, Skoultchi A, Millar SE, Epstein JA. Pax3 functions at a nodal point in melanocyte stem cell differentiation. Nature. 2005; 433: 884-7. doi: 10.1038/nature03292.
33. Yang H, Adam RC, Ge Y, Hua ZL, Fuchs E. Epithelial-Mesenchymal Micro-niches Govern Stem Cell Lineage Choices. Cell. 2017; 169: 483-96 e13. doi: 10.1016/j.cell.2017.03.038.
34. Lei M, Schumacher LJ, Lai YC, Juan WT, Yeh CY, Wu P, Jiang TX, Baker RE, Widelitz RB, Yang L, Chuong CM. Self-organization process in newborn skin organoid formation inspires strategy to restore hair regeneration of adult cells. Proc Natl Acad Sci U S A. 2017. doi: 10.1073/pnas.1700475114.
35. Lei MX, Lai XD, Bai XF, Qiu WM, Yang T, Liao XL, Chuong CM, Yang L, Lian XH, Zhong JLL. Prolonged overexpression of Wnt10b induces epidermal keratinocyte transformation through activating EGF pathway. Histochemistry and Cell Biology. 2015; 144: 209-21. doi: 10.1007/s00418-015-1330-6.
36. Yang K, Chen J, Jiang W, Huang E, Cui J, Kim SH, Hu N, Liu H, Zhang W, Li R, Chen X, Kong Y, Zhang J, et al. Conditional immortalization establishes a repertoire of mouse melanocyte progenitors with distinct melanogenic differentiation potential. J Invest Dermatol. 2012; 132: 2479-83. doi: 10.1038/jid.2012.145.