
INTRODUCTION
Glycoconjugate vaccine, composed of polysaccharides (PSs) covalently linked with carrier protein, can significantly elicit PS-specific antibodies especially IgG [1-4]. Recent studies have placed much emphasis on glycoconjugate modification to enhance its antibody response [5-11]. Glycosylation site selection, as a significant factor leading to heterogeneities of glycoconjugate structure, draws more and more attentions for its impact on the immunogenicity of glycoconjugate vaccine [9, 12].
Recent studies have revealed that glycoconjugate is trimmed into peptides and glycopeptides in antigen presenting cells (APCs). Herein, peptides bind to major histocompatibility complex II (MHCII) and then are presented to T cells, glycopeptides bind to MHCII via their peptide epitopes and then their PS epitopes are presented to T cells [9]. It is reasonable to deduct that different locations of glycosylation sites could lead to diverse presentation of peptides and glycopeptides to APCs and then to be presented to T cells, which might influence the immunogenicity of glycoconjugate vaccine.
To elucidate the relationship between glycosylation connectivity and immunogenicity of glycoconjugate vaccine, in this study, anti-E. coli O157:H7 glycoconjugate O-PS-MBP with defined connectivity, and three selected peptide segments derived from O-PS-MBP were synthesized. In terms of their immunogenic properties in BALB/c mice, we explored different roles of peptides and glycopeptides trimmed from O-PS-MBP on the magnitude and breadth of PS-specific IgG antibodies.
RESULTS
Synthesis of glycoconjugate O-PS-MBP with defined glycosylation sites
To distinguish peptides and glycopeptides generated from glycoconjugate, one simple way is using glycosylation site-specific glycoconjugate. Site-selective chemical conjugation methods have been emerging in recent years, but are almost largely empirical process with high production cost [13-18]. In this study, we investigated the utility of protein glycan coupling technology (PGCT) to produce homogenic glycoconjugate via one-shot in vivo E. coli fermentation [19]. Campylobacter jejuni (C. jejuni) oligosaccharyltransferase PglB, the key enzyme involved in PGCT, is able to transfer PSs to the asparagine residue of the amino acid sequence D/E-X-N-Y-S/T (X, Y≠P) within carrier protein [19, 20]. In this study, maltose-binding protein (MBP) was selected as the carrier protein, because of its defined structure and well-proven immunological enhancement in our previous findings [21]. MBP was modified with a C-terminal peptide sequence containing four consecutive DQNAT motifs, reported as the optimal acceptor sequence recognized by PglB [22]. Then, the modified MBP together with PglB and E. coli O157:H7 O-PS were co-expressed in E. coli strain CLM24, to synthesize O-PS-MBP in vivo (Figure 1 and Supplementary Figure 1) [21].
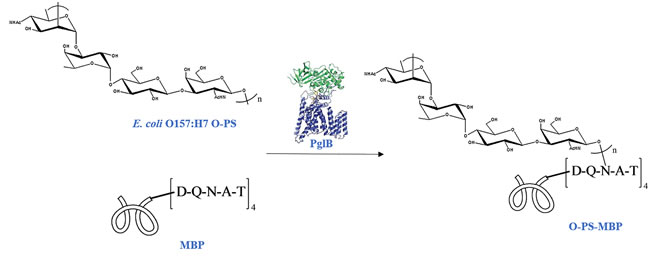
Figure 1: Schematic for biosynthesis of O-PS-MBP. In E. coli CLM24 periplasm, transmembrane protein PglB catalyzes the conjugation of E. coli O157:H7 O-PS with the reducing end GalNAc to the asparagine residue amino acid sequence (DQNAT)4 within carrier protein MBP.
IgG antibodies generated by O-PS-MBP
BALB/c mice were immunized with O-PS-MBP with two booster injections on days 14 and 28. On day 35, sera were collected and titrated against O-PS and MBP respectively in ELISA assays (Table 1). The results showed an average anti-O-PS IgG titer of 4.37 and an average anti-MBP IgG titer of 5.84. When compared the anti-MBP IgG titers between MBP and O-PS-MBP immunized mice, O-PS had slight interference on the immunogenicity of O-PS-MBP, even with no significant difference (Table 1, P > 0.05), which could be explained by an average anti-MBP response which masks the contribution of O-PS on individual peptide segments of O-PS-MBP.
Table 1: Mean titers of serum anti-O-PS and anti-MBP antibodies elicited in mice by injection of MBP and O-PS-MBP.
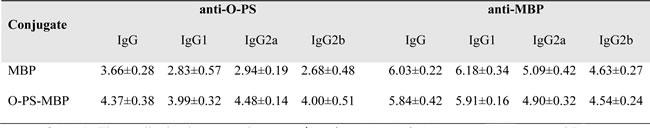
NOTE. The antibody titer was shown as log10(cutoff value). The cutoff value was ODnegative control×2.1.
O-PS shows interference on anti-peptide IgG
To further investigate the interference of O-PS on peptides at different locations of O-PS-MBP, three nonadeca peptide segments GS-1, GS-2 and GS-3 from O-PS-MBP were synthesized, herein, GS-1 containing the glycosylation sites, GS-2 adjacent to the glycosylation sites and GS-3 far from the glycosylation sites (Figure 2 and Supplementary Figure 2). In other words, GS-1 and GS-2 contained peptide epitopes of glycopeptides, while GS-3 contained peptide epitopes of vacant peptide sequences. The anti-GS-1, GS-2, GS-3 IgG titers in mice immunized with O-PS-MBP and MBP were evaluated, respectively (Figure 3A). The results showed that anti-GS-1 and GS-2 IgG titers decreased significantly when MBP was linked with O-PS (P < 0.001), whereas anti-GS-3 IgG response in immunization of O-PS-MBP had no significant difference compared with that of MBP, reflecting that O-PS blocked the generation of IgG antibodies against GS-1 and GS-2 but not GS-3. This phenomenon confirmed the mechanism that peptide epitopes of glycopeptides helped to present PS epitopes to T cells [9]. For that reason, the number of peptide epitopes themselves presenting to T cells reduced, which led to a decline of anti-peptide epitope IgG response.
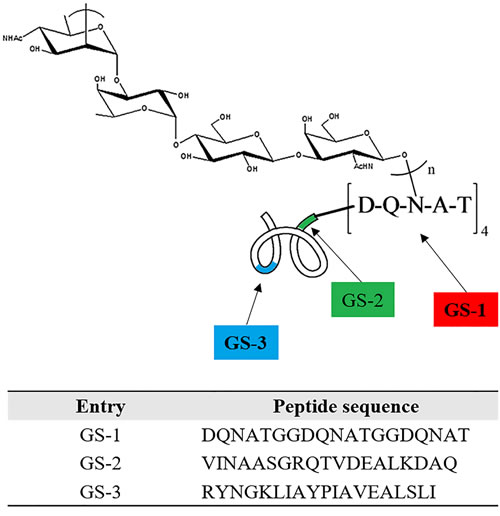
Figure 2: Schematic for peptides GS1, GS2 and GS3. GS-1: contains the glycosylation sites. GS-2: adjacent to the glycosylation sites with 2 amino acid space from (DQNAT)4. GS-3: remote from the glycosylation sites with 251 amino acid space from (DQNAT)4. See amino acid sequence of protein carrier MBP in Supporting Information for specific sequence and location details.
As for the breadth of the IgG response, the anti-O-PS IgG was dominantly IgG2a (P < 0.05), but anti-MBP IgG was dominantly IgG1 (P < 0.0001). When compared the anti-MBP IgG subclass (IgG1, IgG2a, IgG2b) titers between MBP and O-PS-MBP immunized mice, O-PS had also slight interference on the immunogenicity of O-PS-MBP, even with no significant difference (Table 1, P > 0.05). When focusing on individual peptides, we found that anti-GS-1 and GS-2 IgG2a titers decreased dramatically when MBP was linked with O-PS (Figure 3B-3D, P < 0.001), but not GS-3. These results can be explained that, due to GS-1 and GS-2 contained peptide epitopes of glycopeptides, the decline of anti-GS-1 and GS-2 IgG2a contributed to the increase of anti-O-PS IgG2a, then leading to that the anti-O-PS IgG was dominantly IgG2a. In other words, in glycopeptides, the IgG subclass profile against peptide epitope was consistent with that against O-PS.
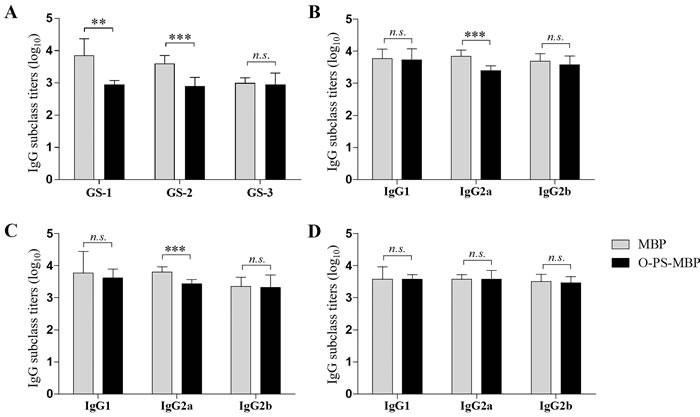
Figure 3: Anti-peptide IgG and subclass titers. A. Anti-peptide IgG titers. B. Anti-GS-1 IgG subclass titers. C. Anti-GS-2 IgG subclass titers. D. Anti-GS-3 IgG subclass titers. Differences were indicated with symbols (**: P < 0.01; ***: P < 0.001; n.s.: P > 0.05). Results were expressed as the arithmetic mean ±SD indicated by error bars.
Peptide priming effect
Inversely, to explore the impact of GS-1, GS-2, GS-3 on anti-O-PS IgG antibodies generated by O-PS-MBP, we primed the mice with GS-1-BSA, GS-2-BSA, GS-3-BSA (conjugating to a protein can enhance the immunogenicity of peptides), respectively, and then immunized the mice all with glycoconjugate O-PS-MBP. After the immunization, no significant difference was detected in anti-O-PS IgG titers between the mice primed with PBS and BSA, reflecting that BSA has no interference for eliciting anti-O-PS IgG titers by O-PS-MBP (Figure 4). Furthermore, the mice primed with GS-1-BSA or GS-2-BSA, rather than those primed with GS-3-BSA, induced higher anti-O-PS IgG titers compared with those primed with BSA (Figure 4). All above results showed that the peptide epitopes from glycopeptides had significant priming effects for eliciting PS-specific IgG titers. Only peptides at and near the glycosylation sites, not all peptides trimmed from carrier protein, had significant priming effects for eliciting PS-specific IgG titers by glycoconjugate vaccine.
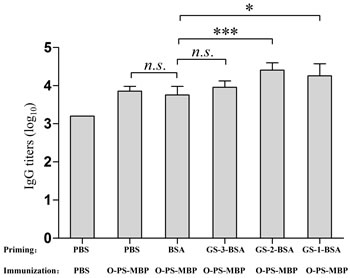
Figure 4: Anti-O-PS IgG titers. Sera from priming with peptides and immunization of O-PS-MBP were tested for anti-O-PS IgG titers. Differences were indicated with symbols (*: P < 0.05; ***: P < 0.001; n.s.: P > 0.05). Results were expressed as the arithmetic mean ±SD indicated by error bars.
DISCUSSION
The importance of glycosylation site selection on the immunogenicity of glycoconjugate vaccine is not obvious because most of these glycoconjugates are made by random chemical methods. In this study, the immunogenicity results of glycoconjugate O-PS-MBP with defined glycosylation sites shows that glycopeptides rather than peptides trimmed from glycoconjugate have an important role in generating PS-specific IgG antibodies. Alternatively, the number and location of glycosylation sites could result in presenting of different glycopeptides to T cells and generating diverse PS-specific antibodies. Based on our findings, for an extension of recently proposed mechanism for adaptive immune system activation by glycoconjugate vaccine (mentioned in introduction), glycoconjugate is trimmed into peptides and glycopeptides in APC. Those peptide epitopes help present PS epitopes to T and B cells to generate PS-specific antibodies. Correspondingly, those peptide epitopes could not present themselves to T and B cells to generate peptide epitope-specific antibodies. Meanwhile, the immunogenicity properties of peptide epitopes are transferred to PS epitopes. In other words, the profile of generated PS-specific antibodies depends on the characteristics of peptide epitopes (Figure 5).
Carrier protein priming/suppression refers to improved/reduced PS-specific response to glycoconjugate vaccine in an individual previously immunized with carrier protein compared with whom not. It is well believed that priming with carrier protein enhances the PS-specific IgG response by increasing the number of protein-specific T cells, which help the proliferation and differentiation of PS-specific B cells [23, 24]. However, based on our findings, only peptides at and near the glycosylation sites, not all peptides trimmed from carrier protein, had significant priming effects for eliciting PS-specific IgG titers by glycoconjugate vaccines. This is a good explanation for a clinical phenomenon: high hapten density of the carrier protein results in priming. Reversely, low ratio of hapten to carrier protein results in suppression.
In conclusion, we have utilized a glycosylation site-selective conjugation method suitable for the synthesis of glycoconjugate O-PS-MBP, which allows a better investigation of the immunological mechanism of glycoconjugate vaccine. We found that the peptide epitopes from the glycopeptide fragments promote the generation of PS-specific IgG and apply their own subclass distribution properties on the PS-specific IgG subclass distribution. By altering the location of the glycosylation sites, the PS-specific IgG titer and subclass profile of glycoconjugate vaccine could be controlled.
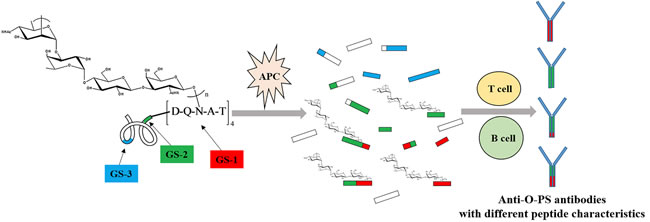
Figure 5: The proposed mechanism of adaptive immune system activation by glycoconjugate O-PS-MBP. O-PS-MBP is trimmed into peptides and glycopeptides in APC. Herein, glycopeptides have two parts, peptide epitopes and PS epitopes. Those peptide epitopes help present PS epitopes to T and B cells to generate PS-specific antibodies. Correspondingly, peptide epitopes could not present themselves to T and B cells to generate peptide epitope-specific antibodies. Meanwhile, the immunogenicity properties of peptide epitopes are transferred to PS epitopes. In other words, the profile of generated PS-specific antibodies depends on the characteristics of peptide epitopes.
MATERIALS AND METHODS
Ethics statement
This study was carried out in accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocols were approved by the Ethics Committee of Shandong University School of Medicine (No. 001 in 2011 for Animal Ethics Approval) and all efforts were made to minimize sufferings.
Bacterial strains, plasmids and growth conditions
E. coli DH5α was used for cloning of plasmids. E. coli CLM24 was used for glycoconjugate expression experiments. All E. coli strains were grown in Luria-Bertani (LB) broth. Ampicillin (100 μg/ml), chloramphenicol (25 μg/ml) and spectinomycin (50 μg/ml) were used for plasmids selection as needed. All strains and plasmids used in this study are listed in Supplementary Table 1.
Construction of recombinant plasmids
The plasmid pYES1L-E. coli O157:H7 O-PS was constructed by inserting E. coli O157:H7 O antigen gene cluster and its upstream genes (including wcaM, z3206, galF) and downstream genes (including gnd, ugd, wzz, hisl) into plasmid pYES1L (GENEART® High-Order Genetic Assembly System, Invitrogen) [21]. The plasmid pACT3-PglB was constructed by inserting pglb gene from C. jejuni NCTC 11168 into plasmid pACT3 at the SmaI-SalI site. The plasmid pBAD24-MBP was constructed by inserting DNA encoding MBP, O-PS acceptor peptide tag GT (i.e., N-DQNATGGDQNATGGDQNATGGDQNAT-C) and a His6 tag (i.e., N-HHHHHH-C) into plasmid pBAD24 at the SmaI-SalI site.
Synthesis and characterization of peptides
The peptides GS-1, GS-2 and GS-3 were synthesized by Bankpeptide, Inc. China and detected by electrospray ionization-mass spectroscopy (ESI-MS) with a Shimadzu LCMS-IT-TOF 8030 spectrometer.
BALB/c mice immunization
Six-week-old female BALB/c mice were injected subcutaneously with 30 μg (protein quantity) of O-PS-MBP or equivalent MBP per mouse, each group contained 8 mice. The mice were injected with three doses of 2 weeks’ interval. Six-week-old female BALB/c mice were injected subcutaneously with 30 μg (peptide quantity) of BSA conjugated peptides (GS-1-BSA, GS-2-BSA, GS-3-BSA) or controls (BSA, PBS) per mouse, each group contained 8 mice. The mice were injected with three doses of 2 weeks’ interval. Then, all groups of mice were injected subcutaneously with 30 μg (protein quantity) of O-PS-MBP per mouse. The mice were injected with two doses of 2 weeks’ interval. O-PS-MBP and MBP were formulated in PBS (pH 7.4) with Freund’s complete adjuvant (FCA) (Sigma) for the first immunization or Freund’s incomplete adjuvant (FIA) (Sigma) for the other immunizations. Seven days after the final immunization, the blood were taken from the tail vein of mice and centrifuged at 3,000g for 30 min to obtain sera.
Analysis of IgG responses
The IgG responses against E. coli O157:H7 O-PS, MBP or peptides induced by different formulations, were measured by ELISA as described previously [21, 25]. Briefly, the 96-well plates (Costar® Polystyrene High Binding Plate 3590) were coated with 100 μl of 1 μg/ml E. coli O157:H7 LPS in 0.05 M Na2CO3 (pH 9.8) or 10 μg/ml MBP in PBS (pH 7.4) or 10 μg/ml peptides (GS-1, GS-2, GS-3) in 0.05 M Na2CO3 (pH 9.8), followed by incubation at 4oC overnight. The coated plates were then washed with 250 μl PBS + Tween (PBST) (pH 7.4) for five times and then blocked with 2% (w/v) BSA in PBS for 2 h at room temperature. After washing as above, two-fold serial dilutions of sera in PBS were then added to each column. Plates were incubated for 2 h at room temperature and washed again. HRP-goat anti-mouse IgG, IgG1, IgG2a, IgG2b (Abcam) appropriately diluted in PBST were added and the plates were incubated for 1 h at room temperature. After another washing, 100 μl TMB solution was added to the plates and incubated for 15 min before 100 μl of 1 M HCl was added. The plates were read at OD450. Sera titers were expressed as the log10(maximal dilution fold) corresponding to a cut-off value at ODnegative control×2.1.
Statistical analysis
All figures and statistical analyses were generated using the program GraphPad Prism version 5.0. Data were shown as means ± standard deviation (SD). When two groups were compared, t test was used for analyses. One-way ANOVA was used to test for statistical significance of differences between three experimental groups.
Abbreviations
PS: polysaccharide; APCs: antigen presenting cells; MHCII: major histocompatibility complex II; PGCT: protein glycan coupling technology; MBP: maltose-binding protein; LB: Luria-Bertani; FCA: Freund’s complete adjuvant; FIA: Freund’s incomplete adjuvant; SD: standard deviation; C. jejuni: Campylobacter jejuni.
Author contributions
M.C., Z.M. and P.G.W. designed the experiments. M.C., Z.M., H.Z. and X.L. prepared manuscript; Z.M. and H.Z. performed the experiments; Z.M., H.Z., X.L. and M.C. analyzed the data. All authors reviewed and approved the final version.
CONFLICTS OF INTEREST
The authors declare no competing financial interest.
FUNDING
This work was financially supported by the National Natural Science Foundation of China (No. 31270983) and National Institutes of Health (No. R01 AI083754).
REFERENCES
1. Mond JJ, Lees A, Snapper CM. T cell-independent antigens type 2. Annu Rev Immunol. 1995; 13: 655-92. doi: 10.1146/annurev.iy.13.040195.003255.
2. Stein KE. Thymus-independent and thymus-dependent responses to polysaccharide antigens. J Infect Dis. 1992 (Suppl 1); 165:S49–52.
3. Avery OT, Goebel WF. Chemo-Immunological Studies on Conjugated Carbohydrate-Proteins : Ii. Immunological Specificity Of Synthetic Sugar-Protein Antigens. J Exp Med. 1929; 50: 533-50.
4. Rappuoli R, De Gregorio E. A sweet T cell response. Nat Med. 2011; 17: 1551-2. doi: 10.1038/nm.2587.
5. Jackson LA, Baxter R, Reisinger K, Karsten A, Shah J, Bedell L, Dull PM, Group VPS. Phase III comparison of an investigational quadrivalent meningococcal conjugate vaccine with the licensed meningococcal ACWY conjugate vaccine in adolescents. Clin Infect Dis. 2009; 49: e1-10. doi: 10.1086/599117.
6. Southern J, Borrow R, Andrews N, Morris R, Waight P, Hudson M, Balmer P, Findlow H, Findlow J, Miller E. Immunogenicity of a reduced schedule of meningococcal group C conjugate vaccine given concomitantly with the Prevenar and Pediacel vaccines in healthy infants in the United Kingdom. Clin Vaccine Immunol. 2009; 16: 194-9. doi: 10.1128/CVI.00420-08.
7. Halperin SA, Gupta A, Jeanfreau R, Klein NP, Reisinger K, Walter E, Bedell L, Gill C, Dull PM. Comparison of the safety and immunogenicity of an investigational and a licensed quadrivalent meningococcal conjugate vaccine in children 2-10 years of age. Vaccine. 2010; 28: 7865-72. doi: 10.1016/j.vaccine.2010.09.092.
8. Granoff DM, Anderson EL, Osterholm MT, Holmes SJ, McHugh JE, Belshe RB, Medley F, Murphy TV. Differences in the immunogenicity of three Haemophilus influenzae type b conjugate vaccines in infants. J Pediatr. 1992; 121: 187-94.
9. Avci FY, Li X, Tsuji M, Kasper DL. A mechanism for glycoconjugate vaccine activation of the adaptive immune system and its implications for vaccine design. Nat Med. 2011; 17: 1602-9. doi: 10.1038/nm.2535.
10. Seppala I, Makela O. Antigenicity of dextran-protein conjugates in mice. Effect of molecular weight of the carbohydrate and comparison of two modes of coupling. J Immunol. 1989; 143: 1259-64.
11. Buskas T, Li Y, Boons GJ. The immunogenicity of the tumor-associated antigen Lewis(y) may be suppressed by a bifunctional cross-linker required for coupling to a carrier protein. Chemistry. 2004; 10: 3517-24. doi: 10.1002/chem.200400074.
12. Stefanetti G, Hu QY, Usera A, Robinson Z, Allan M, Singh A, Imase H, Cobb J, Zhai H, Quinn D, Lei M, Saul A, Adamo R, et al. Sugar-Protein Connectivity Impacts on the Immunogenicity of Site-Selective Salmonella O-Antigen Glycoconjugate Vaccines. Angew Chem Int Ed Engl. 2015; 54: 13198-203. doi: 10.1002/anie.201506112.
13. Crotti S, Zhai HL, Zhou J, Allan M, Proietti D, Pansegrau W, Hu QY, Berti F, Adamo R. Defined Conjugation of Glycans to the Lysines of CRM197 Guided by their Reactivity Mapping. Chembiochem. 2014; 15: 836-43. doi: 10.1002/cbic.201300785.
14. Hu QY, Allan M, Adamo R, Quinn D, Zhai HL, Wu GX, Clark K, Zhou J, Ortiz S, Wang B, Danieli E, Crotti S, Tontini M, et al. Synthesis of a well-defined glycoconjugate vaccine by a tyrosine-selective conjugation strategy. Chemical Science. 2013; 4: 3827-32. doi: 10.1039/c3sc51694f.
15. Adamo R, Hu QY, Torosantucci A, Crotti S, Brogioni G, Allan M, Chiani P, Bromuro C, Quinn D, Tontini M, Berti F. Deciphering the structure-immunogenicity relationship of anti-Candida glycoconjugate vaccines. Chemical Science. 2014; 5: 4302-11. doi: 10.1039/c4sc01361a.
16. Boutureira O, Bernardes GJ. Advances in chemical protein modification. Chem Rev. 2015; 115: 2174-95. doi: 10.1021/cr500399p.
17. Grayson EJ, Bernardes GJ, Chalker JM, Boutureira O, Koeppe JR, Davis BG. A coordinated synthesis and conjugation strategy for the preparation of homogeneous glycoconjugate vaccine candidates. Angew Chem Int Ed Engl. 2011; 50: 4127-32. doi: 10.1002/anie.201006327.
18. Spicer CD, Davis BG. Selective chemical protein modification. Nat Commun. 2014; 5: 4740. doi: 10.1038/ncomms5740.
19. Ihssen J, Kowarik M, Dilettoso S, Tanner C, Wacker M, Thony-Meyer L. Production of glycoprotein vaccines in Escherichia coli. Microb Cell Fact. 2010; 9: 61. doi: 10.1186/1475-2859-9-61.
20. Feldman MF, Wacker M, Hernandez M, Hitchen PG, Marolda CL, Kowarik M, Morris HR, Dell A, Valvano MA, Aebi M. Engineering N-linked protein glycosylation with diverse O antigen lipopolysaccharide structures in Escherichia coli. Proc Natl Acad Sci U S A. 2005; 102: 3016-21. doi: 10.1073/pnas.0500044102.
21. Ma Z, Zhang H, Shang W, Zhu F, Han W, Zhao X, Han D, Wang PG, Chen M. Glycoconjugate vaccine containing Escherichia coli O157:H7 O-antigen linked with maltose-binding protein elicits humoral and cellular responses. PLoS One. 2014; 9: e105215. doi: 10.1371/journal.pone.0105215.
22. Chen MM, Glover KJ, Imperiali B. From peptide to protein: comparative analysis of the substrate specificity of N-linked glycosylation in C. jejuni. Biochemistry. 2007; 46: 5579-85. doi: 10.1021/bi602633n.
23. Kurikka S. Priming with diphtheria-tetanus-pertussis vaccine enhances the response to the Haemophilus influenzae type b tetanus conjugate vaccine in infancy. Vaccine. 1996; 14: 1239-42.
24. Granoff DM, Holmes SJ, Osterholm MT, McHugh JE, Lucas AH, Anderson EL, Belshe RB, Jacobs JL, Medley F, Murphy TV. Induction of immunologic memory in infants primed with Haemophilus influenzae type b conjugate vaccines. J Infect Dis. 1993; 168: 663-71.
25. Iwashkiw JA, Fentabil MA, Faridmoayer A, Mills DC, Peppler M, Czibener C, Ciocchini AE, Comerci DJ, Ugalde JE, Feldman MF. Exploiting the Campylobacter jejuni protein glycosylation system for glycoengineering vaccines and diagnostic tools directed against brucellosis. Microb Cell Fact. 2012; 11: 13. doi: 10.1186/1475-2859-11-13.