
Introduction
Bladder cancer, particularly muscle invasive bladder cancer (MIBC), is amongst the most common and deadliest genitourinary cancers [1]. The mainstay treatment for advanced stage tumours includes surgery and cisplatin-based chemotherapeutic regimens [1], which fail in avoiding tumour relapse and disease progression. Tremendous efforts have been put in the establishment of biomarker panels for early diagnosis, follow-up, patient stratification, prognosis, treatment selection and development of targeted therapeutics [2]. However, the highly heterogeneous molecular nature of bladder tumours has hampered true developments in this field [3]. Moreover, bladder cancer remains mostly an “orphan disease” in terms of targeted therapeutics, leading to few improvements in patient’s overall survival over the last decade [2, 4]. More detailed information on the clinicopathological nature of bladder tumours and critical aspects in disease management have been recently reviewed [5]. A schematic illustration of bladder cancer staging and grading is shown in Figure 1.
Several decades of glycobiology research have disclosed the existence of profound alterations in the glycosylation patterns of bladder tumours, reflecting specific changes in glycan biosynthetic pathways, glycosyltransferases expression, amongst other factors [6]. These events often lead to novel protein and lipid glycoforms, either by incomplete or neo-synthesis of glycan epitopes, that cannot be found in the corresponding healthy tissues and preneoplastic lesions. These events play a key role in tumour progression by affecting ligand-receptor interactions, and interfering with regulation of cell signaling, adhesion, migration, proliferation, angiogenesis, and immune responses [6]. Moreover, cancer-associated glycans may be actively secreted into bodily fluids (e.g. blood and urine) or shed from apoptotic and necrotic cancer cells [7]. As such, glycans and abnormally glycosylated molecules (e.g. proteins and lipids) hold tremendous value for non-invasive cancer detection, while membrane bound glycans may be used to selectively target tumour sites and specific cancer cells. Nevertheless, the structural complexity and heterogeneity of oligosaccharides, and the lack of analytical methods for elucidating structures still pose a major difficulty when addressing the glycome, glycolipidome and glycoproteome [8]. Still, a plethora of mass spectrometry-based analytical approaches have been developed to address these challenges [8, 9] and the standardization of high-throughput glycomics is expected to boost our knowledge on bladder cancer glycobiology in the near future.
Based on these considerations, the present review comprehensively summarizes the clinical significance of the main biomarkers arising from over forty years of bladder cancer glycobiology research and establishes the milestones towards clinical applications. Ultimately, we discuss the need to integrate glycans in holistic panomics models for precision oncology, namely the molecular-based individualization of patient care.
Glycosylation signatures in bladder cancer: biological and clinical implications
Glycosylation is the most frequent, complex and plastic post-translational modification of secreted and membrane-bound proteins, as well as a common substitution in lipids at the cell membrane [10]. Glycans are secondary gene products resulting from the coordinated action of nucleotide sugar transporters, glycosyltransferases and glycosidases in the endoplasmic reticulum (ER) and Golgi apparatus (GA) of mammalian cells [10]. Glycans are involved in several structural, modulatory, molecular mimicry and recognition roles including protein folding, stability, adhesion and trafficking, as recently reviewed [11]. Alterations in glycosylation patterns are common features of solid tumours, being detected even in pre-malignant lesions [12]. Generally, the most frequently described cancer-related glycosylation modifications include the synthesis of highly branched and heavily sialylated glycans, the premature termination of biosynthesis, resulting in the expression of short-chained forms, and the expression de-novo of glycosidic antigens of foetal type [13]. These structural motifs are mostly associated with: i) altered glycogenes expression [14, 15]; ii) impaired glycosyltransferases’ chaperone function [16]; iii) altered glycosidase/glycosyltransferase activity [15]; iv) reorganization of glycosyltransferases topology [17, 18]; v) bioavailability of sugar nucleotide donors and cofactors [19]; vi) alterations on the conformation of peptide backbone or on the nascent glycan chain structure [19]. The resultant aberrant and cancer-associated glycans seem to be implicated in the activation of oncogenic pathways [20], establishment of tumour-tolerogenic immune responses [21], and in epithelial-to-mesenchymal transition (EMT), a crucial milestone towards invasion and metastasis [22, 23]. Thus, many glycoepitopes, and their related glycosidases/glycosyltransferases, can be considered relevant tumour-associated antigens [24, 25], with possible clinical significance in bladder cancer. Therefore, the following sections will focus on these key findings in bladder cancer glycobiology (summarized in Supplementary Table 1 - Supplementary material). Given their structural complexity and broad distribution, known cancer-associated glycogenes, glycosyltransferases and glycans will be presented in the context of specific classes of biomolecules (glycoproteins, glycolipids, proteoglycans).
Protein glycosylation
Two main classes of glycans can be found altered in cancer cell-surface proteins, namely N-glycans, attached to the peptide sequence via an asparagine (Asn) residue, and O-glycans, attached by a N-acetylgalactosamine (GalNAc) residue to the hydroxyl group of a serine (Ser) or threonine (Thr) residue.
Cancer-associated N-glycans
Protein N-glycosylation takes place in the ER, where the oligosaccharide transferase complex (OSTase) scans nascent proteins for Asn-X-Ser/Thr “sequons” (“X” stands for any amino acid residue except proline), and transfers a precursor glycan (Glc3Man9GlcNAc2-) from dolichol pyrophosphate to Asn residues [26]. At this point, all N-glycans share a common core structure (Manα1-6(Manα1-3)Manβ1-4GlcNAcβ1-4GlcNAcβ1-Asn-X-Ser/Thr), which is further processed in the ER and GA by several glycosyltransferases and glycosidases, yielding mature core structures that may be classified into three major N-glycan types (oligomannose, complex, and hybrid, Figure 2). The O-3 linked Man residues in hybrid and complex N-glycans may be further O-4 substituted with N-acetylglucosamine (GlcNAc) residues by GlcNAcT-III (GnT-III) to yield bisecting core structures. The introduction of the bisecting GlcNAc residue by GnT-III alters the composition and conformation of the N-glycan, resulting in the suppression of further processing and elongation [27, 28]. More highly branched N-glycans may be generated by the action of different GlcNActransferases (GnT-IV, -V, -VI). These structures may be further elongated with galactose, poly-N-acetyllactosamine, sialic acid, and fucose residues. Particularly, N-glycans frequently exhibit Lewis (Le) blood group related antigens (Lea, Lex, Leb and Ley) and corresponding sialylated structures or ABO(H) blood group determinants as terminal epitopes. Similar terminal structures may also be found in O-glycans (Figure 2). Other sugar modifications may include phosphorylation, O-acetylation of sialic acids, and O-sulfation of galactose and N-acetylglucosamine residues, thereby increasing the structural complexity of the glycome [29].
Several N-glycan alterations have been described in bladder tumours, including changes in branching and terminal structures through oversialylation, fucosylation (Supplementary Table 1), which will be discussed in detail in the following sections.
N-glycans branching
Alterations in N-glycans branching resulting from impaired GnTs expression have been evaluated in the context of bladder cancer prognosis. Namely, increased GnT-III, N-glycans bisection and GnT-IV expression were associated with higher disease stage and grade in bladder cancer patients [30]. Conversely, decreased GnT-V expression, responsible by O-6 N-glycans branching, was found associated with higher bladder tumour grade and stage, shorter disease-free survival and bladder cancer recurrence [31, 32]. Moreover, low GnT-V expression was found to predict shorter cause-specific survival of bladder cancer patients while overexpression of O-6 branched N-linked oligosaccharides was associated with lower tumour stage, suggesting that these findings could be applied to risk stratification [32]. The opposing associations of GnT-III and GnT-V in bladder cancer prognosis can be explained by the antagonistic effect of their enzymatic activity [28]. Contrasting with the findings for bladder cancer, reduced GnT-III and increased GnT-V expressions have been found to promote metastasis in different cancer models [33-36] yet no consensus exists between GnT-V expression and prognosis in gastric [35, 36], oral squamous cell [37] and endometrial cancers [38]. These observations suggest GnT-V/III evaluation may hold potential for bladder cancer prognosis and ultimately targeted therapeutics, which warrants confirmation in future studies.
b) N-glycans terminal structures (also found in protein O-glycans and glycolipids)
Amongst the most common cancer-associated structural features are alterations of terminal glycan epitopes. In fact, the first reported glycosylation alterations in bladder cancer were the loss of ABO(H) blood group determinants in advanced stage carcinomas of secretor individuals [39, 40], as well as changes in Lewis antigens patterns.
The ABO(H) blood group system consists of terminal oligosaccharide antigens carried by glycoproteins or glycolipids in hematopoietic or epithelial cells [41]. Their biosynthesis is presumed to be controlled by the ABO(H), Se, H, Le, and X blood group genes [41]. These antigens are present on normal bladder epithelium of secretor individuals but not on some low-grade and early-stage papillary urothelial carcinomas [42]. Moreover, initially expressing tumours lose these cell surface antigens upon local recurrence, progression to invasion or metastization [42]. As such, the possibility that loss of genetically predicted blood group antigens precedes the development of recurrent, invasive or metastatic bladder cancer has been extensively explored [43]. Studies have shown that abnormally low or absent expression of these epitopes is frequently found in high grade and invasive bladder disease [44-46] and associated with bladder tumour progression and shorter recurrence-free survival [47]. Furthermore, loss of tissue ABO(H) antigens in the initial biopsy of bladder carcinomas predicts a much greater chance of subsequent invasion than in tumours with detectable ABO(H) antigens [44, 45, 47]. However, a significant number of patients whose initial tumours were reported as blood group antigen negative failed to develop an invasive tumour [47]. It is possible that these conflicting results may, at least in part, be explained by differences in methodology, interpretation, or both. Moreover, the loss of activity of the A and B gene-encoded transferases in bladder tumours from blood group A and B individuals was reported, which explains the deletion of these antigens in bladder tumours [48]. In addition, the loss of the ABO(H) gene and/or its promoter hypermethylation is a specific marker for urothelial carcinoma [39]. In summary, alterations in ABO(H) accompanying bladder malignant transformation and disease dissemination are well established surrogate markers of profound alterations in glycosylation pathways, constituting important starting points for more in depth structural studies.
The ABO(H) determinants have biosynthetic and structural similarities with Lewis antigens, including the fucosylated type 1 Lewisa (Galβ(1-3)GlcNAc[Fuc3b1;(1-4)]) and type 2 Lewisx (Galβ(1-3)GlcNAc[Fuc3b1;(1-4)]). Several authors have associated Lewisa and Lewisx expression patterns with malignant transformations of the bladder, reporting significantly lower expression of this antigen in healthy urothelium when compared to invasive tumours [44, 46]. As such, reduced expression of Lewisa and Lewisx was associated with higher tumour grade and invasion [44] and shorter recurrence-free survival [49]. As such, the expression of these antigens can be associated with worse bladder cancer phenotypes. Moreover, Lewisa antigen expression patterns change at an early neoplastic stage, suggesting that Lewisa determination might be useful in the diagnosis of very early premalignant changes in the urothelium [49]. In addition, scoring Lewisa expression allows the sub-classification of histologically identical tumours into prognostically different groups, pointing to a relationship between the pathological grade and stage of the evaluated tumours and a morphological and functional de-differentiation [49]. Given this, Lewisa antigen is a valuable functional marker of the malignant potential in superficial bladder cancer. In turn, the Lewisx antigen is not expressed in normal urothelium, except for occasional umbrella cells [46, 50], but has been found in the majority of invasive tumours, regardless of blood type and secretor status of the individuals studied [46]. Lewisy is expressed in both normal urothelium and bladder tumours, yet its expression was associated with bladder tumour invasion capability [46]. Nevertheless, the number of studies concerning Lewis antigens in bladder cancer is still scarce to withdraw conclusions about their biological and clinical significance.
c) Oversialylation and fucosylation (also occurring in protein O-glycans and glycolipids)
Oversialylation of cancer cells often stem from the overexpression of sialylated Lewis antigens sialyl lewisa (SLea; the CA19-9 antigen) and sialyl lewisx (SLex), which can be found as terminal epitopes of N-glycans, O-glycans and glycolipids [51]. SLea/x are specific ligands for E- and P-selectins in endothelial cells, thereby promoting the adhesion of malignant cells to the endothelium and the metastatic cascade [50-52]. These antigens also thought to play a role in tumour growth, invasion, and angiogenesis [51, 53]. In line with these observations, the overexpression of SLea and SLex have also been associated with bladder cancer malignant potential. Particularly, serum overexpression of SLea was associated with higher stage, grade and invasion [53] while tissue loss/reduction of SLea expression was associated with higher atypia grade [50]. SLex has been closely link to invasive and metastatic potential of primary bladder tumours and correlated with shorter 5-year and 7-year survival rates [54], but another study demonstrated no associations between SLex with grade or stage in urothelial carcinoma of the renal pelvis, ureter, and urinary bladder [50]. The disialylated form of Lea (termed disialyl-Lewisa, dSLea) was described as preferentially expressed in non-malignant cells, and may be useful for distinguishing benign from malignant diseases mostly expressing SLea [55]. Supporting these observations, the overall increase in cell-surface sialic acid content was shown to reduce the attachment of metastatic tumour cells to the extracellular matrix [56]. These observations support the need for a comprehensive interrogation of bladder cancer cells “sialome” towards understanding tumour progression and dissemination. Moreover, future studies should explore the biological and clinical relevance of structurally identical sialylated forms in the context of bladder cancer.
Fucosylation is another common modification involving oligosaccharides on glycoproteins and glycolipids [57]. Particularly, the quantitative glycome analysis of N-glycan patterns in bladder cancer cells often reveals significant differences in N-glycan fucosylation compared to normal cells. Namely, bladder cancer cells (KK47, YTS1, J82, T24) showed high expression of complex core-fucosylated N-glycans and low expression of terminally fucosylated N-glycans [58]. Nevertheless, the implications of these differential fucosylation patterns in bladder cancer malignancy have been so far poorly explored. The transcript levels of fucosyltransferase (FUT) VI (FUT-VI) and FUT-VII from invasive and non-invasive bladder tumours were also explored using RT-PCR. Particularly, bladder cancer cell lines from invasive tumours that maintained their metastatic properties showed high levels of both enzymes, and cell lines from non-invasive tumours (KK-47) or normal bladder epithelia (HCV-29) were negative for FUT-VI and FUT-VII [54]. These evidences suggest that FUT-VI/-VII expression associates with more malignant cancer cell phenotypes. Another study has described β1-integrin activation by alpha1,2-fucosyltransferase 1 (FUT-I)-mediated fucosylation in J82 human bladder cancer cells, thereby enhancing bladder cancer adhesion and subsequent metastasis [59]. As such, changes in bladder cancer fucosylation patterns seem to be associated with tumour invasion and progression to metastization in cancer cell lines, suggesting that these changes could provide novel strategies for cancer therapy.
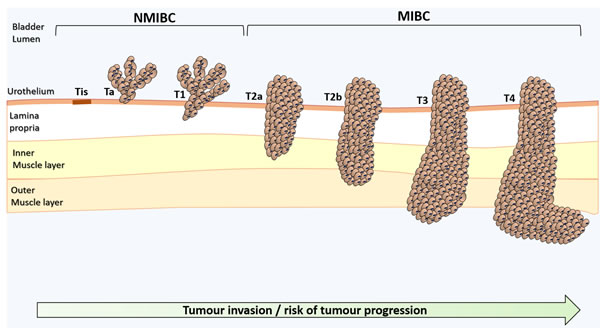
Figure 1: Schematic representation of bladder cancer stage and grade. The stage of the primary tumour (T) is based on the extent of penetration or invasion into the bladder wall. Regarding tumour grading, bladder lesions can be classified as urothelial papilloma (a benign lesion), papillary urothelial neoplasm of low malignant potential (PUNLMP), low-grade papillary urothelial carcinoma and high-grade papillary urothelial carcinoma. Of note, PUNLMP lesions do not have cytological features of malignancy and have a very low risk of progression. Nevertheless, they show high tendency to recur. Tis, Tumour in situ: ‘‘flat tumour’’; Ta, Non-invasive papillary carcinoma; T1, Tumour invades sub-epithelial connective tissue; T2, Tumour invades muscle; T2a, Tumour invades superficial muscle (inner half); T2b, Tumour invades deep muscle (outer half); T3, Tumour invades perivesical tissue; T4, Tumour invades any of the following: prostate, uterus, vagina, pelvic or abdominal wall.
Cancer-associated O-glycosylation
The most common form of cell-surface protein O-glycosylation results from the transfer of a GalNAc residue from a UDP-GalNAc donor to either serine or threonine in a given polypeptide chain (O-GalNAc glycosylation), originating the monosaccharide Tn antigen. This reaction is catalysed by several UDP-GalNAc:polypeptide N-acetylgalactosaminyl transferases (ppGalNAc-Ts) in the ER, in a substrate dependent manner [60]. As opposed to N-glycosylation, no consensus sequence is required for ppGalNAc-Ts recognition. The Tn antigen is generally extended with a Gal residue by Gal-transferase (β(1-3)-galactosyltransferase, C1Gal-T1 or T-synthase) and cosmc chaperone, originating the disaccharide Thomsen-Friedenreich or T antigen (Galβ1-3GalNAcα-O-Ser/Thr, core 1). Alternatively, Tn and T antigens can be sialylated by sialyltransferases, forming the sialyl-Tn (STn), sialyl-T and disialyl-T antigens. Sialylation stops any further processing of the oligosaccharide chain, prompting short-chain GalNAc-type O-glycans expression [60]. Alternatively, core 1 may be extended originating cores 2-4 (Figure 3), which are precursors for a vast array of more extended oligosaccharides and terminal structures, similar to the ones found in mature N-glycans.
Recently, a precision mapping of human O-GalNAc glycoproteome has revealed over 6000 glycosites in more than 600 O-glycoproteins, the majority of which of membrane origin [61], greatly expanding our view on the O-glycoproteome and its functional role. Alterations in O-glycosylation pathways are a common hallmark of malignant transformations, frequently amplified at the cell-surface as a result of the high number of O-glycosylation sites presented by mucins [62]. Such events are particularly pronounced in adenocarcinomas, due to the overexpression of these molecules [63]. While hindered by extended glycosylation in healthy and benign tissues, simple mucin-type O-GalNAc glycans are uncovered in most human carcinomas, including bladder cancer [45, 64-67].
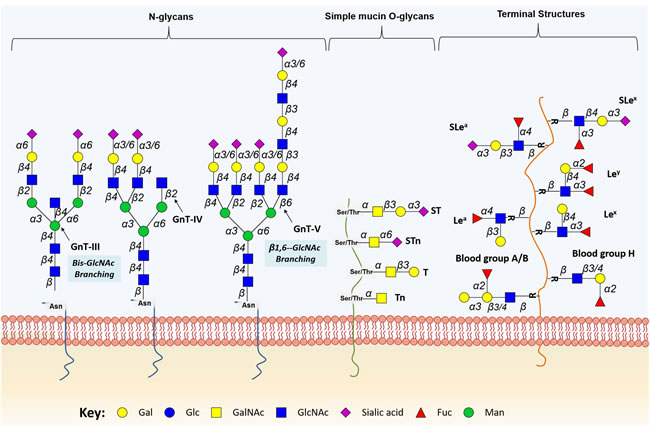
Figure 2: Schematic representation of protein-associated glycan structures relevant in bladder cancer. The figure represents specific N-linked and O-linked glycan structures, as well as terminal Lewis and sialylated Lewis structures that have biological significance in bladder cancer. Key enzymes mediating the addition of specific sugars are also shown. Protein N-glycan alterations include the β1-6 branching of N-glycans in result of GlcNAcT-V (GnT-V) overexpression, and the addition of bisecting GlcNAc branches by GlcNAcT-III (GnT-III) glycosyltransferases. Alterations in O-glycosylation pathways are also a common hallmark of malignant transformations of the bladder. Herein, we represent the overexpression of simple mucin-type O-glycans and their sialylated counterparts, T, sialyl T (ST), Tn and sialyl Tn (STn) antigens. Altered expression of terminal structures is also a common feature of bladder tumours. Namely, the abnormally low or absent expression of ABO(H) blood group determinants is frequently found in high grade and invasive disease. Carbohydrate terminal Lewis antigens are significantly under-expressed in healthy urothelium when compared to bladder tumours and are also highlighted here. Lewis type 1 antigens include Lewisa (Lea), and sialyl Lewisa (SLea), while the type 2 group includes Lewisx (Lex) and sialyl Lewisx (Slex).
a) Premature stop in O-glycosylation
Perhaps the most studied cancer-associated O-glycans are the Tn antigen, its sialylated counterpart sialyl-Tn (STn) and the T antigen. They result from a premature stop in protein O-glycosylation and are classically termed simple mucin-type O-glycans, reflecting their overexpression in cancer-associated mucins [68]. Nevertheless, these alterations can also be significantly observed in other densely O-glycosylated proteins of relevant importance in bladder cancer, namely CD44 and different types of integrins [69, 70]. Several reports attribute the expression of simple mucin-type O-glycans to a disorganisation of secretory pathway organelles in cancer cells, mutations on Cosmc, a gene encoding a molecular chaperone of T-synthase [16, 71], and absence or altered expression and/or activity of glycosyltransferases [72]. In particular, the overexpression of ST6GalNAc-I has been found to promote the premature sialylation of the Tn antigen and consequent formation of the STn antigen in bladder cancer [64, 69]. Specifically, the STn antigen is absent in the healthy urothelium, while being present in more than 70% of high-grade NMIBC and MIBC, denoting a cancer specific nature [64]. This post-translational modification of cell surface proteins is mostly expressed in non-proliferative tumour areas, known for their high resistance to cytostatic agents currently used to improve the overall survival of advanced stage bladder cancer patients [64]. Recently, a novel STn-dependent mechanism for chemotherapeutic resistance of gastric cancer cells to cisplatin has been described, in which STn protects cancer cells against chemotherapeutic-induced cell death by decreasing the interaction of cell surface glycan receptors with galectin-3 and increasing its intracellular accumulation [73]. Nevertheless, the relationship between chemoresistance and STn overexpression remains to be fully explored in bladder cancer. Furthermore, STn expression is significantly higher in MIBC when compared to NMIBC, denoting its association with muscle invasion and poor prognosis [20]. Studies in vitro have further demonstrated that this antigen plays an important role in bladder cancer cell migration and invasion through mechanisms so far unexplored [64, 69]. Recent glycoproteomics studies of bladder cancer cell models highlighted that STn was mainly present in integrins and cadherins, further reinforcing a possible role for this glycan in adhesion, cell motility and invasion [69]. Also, recent work from our group has demonstrated the presence of STn in lymph node and distant metastasis, strengthening the notion that STn expression may influence cancer cell motility and metastization (unpublished data). Furthermore, STn-expressing bladder cancer cells have shown the ability to induce a tolerogenic microenvironment by impairing dendritic cells maturation, allowing cancer cells to evade innate and adaptive immune system responses [21]. Interestingly, the tolerogenic effect of short-chained O-glycans has also been correlated with bladder tumour metastasis through a mechanism in which MUC1 carrying core 2 O-glycans functions as a molecular shield against NK cells attack, thereby promoting metastization [74]. In addition, STn expression in bladder cancer tissues has been used in combination with other surrogate markers of tumour aggressiveness envisaging patient stratification regarding disease stage and therapeutic benefit. Specifically, expression of STn and sialyl-6-T (s6T), a sialylated form of T antigen, are independent predictive markers of BCG treatment response and were found useful in the identification of patients who could benefit more from this immunotherapy [75]. Moreover, STn was found to be a marker of poor prognosis in bladder cancer and, in combination with PI3K/Akt/mTOR pathway evaluation, holds potential to improve disease stage stratification [20]. In turn, it was observed that the reduction of Tn antigen expression was associated with higher bladder cancer stage [67].
Several reports associated the presence of T antigens with higher grade, stage and poor prognosis in bladder cancer [66, 76], suggesting that these antigens may be surrogate markers of profound cellular alterations. Also, there is growing evidences linking the overexpression of ST3Gal.I, the enzyme responsible for T antigen sialylation, with higher stage and poor prognosis [65]. Moreover, the expression of T antigen is significantly associated with higher risk for subsequent recurrences with deep muscle invasion and metastatic involvement of regional lymph nodes [67]. In agreement with these observations, we have recently reported that short-chain O-glycans are preferentially accumulated in hypoxic tumour areas [69], known to harbor more malignant sub-populations. It has been suggested that HIF-1α directly or indirectly modulates the expression of glycosyltransferases involved in the initial steps of O-glycosylation while repressing core elongation, thereby promoting an accumulation of precursor structures [69]. The fact that these simple glycans are absent, significantly under-expressed or restricted to some cell types in healthy tissues, makes them ideal diagnostic and therapeutic targets for bladder cancer therapy [77].
Overexpression of cancer-associated membrane glycoproteins
Alteration in N- and O-glycosylation and other types of protein glycans are often amplified in cancer cells by the overexpression of key cancer-associated glycoproteins. Namely, HER2 (also known as ErbB2 or HER2/neu) is an heavily glycoprotein [78], member of the EGF receptor (EGFR) family, that is overexpressed in several malignancies, including advanced stage bladder cancer [79-81]. Curiously, the incidence of HER2 overexpression in bladder cancer (12.4%) is even higher than that found in breast carcinomas (10.5%), where it is associated with tumour aggressiveness, prognosis and responsiveness to therapy [81]. In fact, HER2 expression is also associated with poor prognosis in bladder cancer [82]. Thus, HER2 could serve as a useful biomarker for clinical prediction and trials of anti-HER2 agents are warranted in patients with advanced bladder cancer. Nevertheless, the glycosylation of HER2 in bladder cancer remains to be addressed, which would be critical for the establishment of a more sensitive and specific biomarker.
EpCAM, also known as CD326, is a glycoprotein predominantly located in intercellular spaces of epithelial, progenitor and normal stem cells [83, 84]. This transmembrane macromolecule regulates both normal and cancer-associated cellular adhesion, proliferation, differentiation, migration and invasion [84, 85]. Its expression is associated with increased tumour stage and grade, as well as with poor prognosis and decreased overall survival in bladder cancer patients [86, 87]. Despite these evidences, the glycosylation pattern of EpCAM in bladder cancer has also not yet been evaluated.
Frequently, cancer cells also overexpress galectins, N-acetyllactosamine-binding glycoproteins yielding either one or two carbohydrate-recognition domains. Galectins cross-link glycoproteins depending on their glycan structures and concentrations, forming galectin-glycan molecular lattices [88]. Particularly, the correlation between increased galectin expression and tumour progression is proposed to be linked to their interaction with poly-N-acetyllactosamines on matrix proteins such as laminin, aiding cellular invasion [89]. Moreover, these glycoproteins are known to modulate cell growth, differentiation, adhesion, and apoptosis [90-92]. The altered expression of galectins has been implicated in bladder cancer malignancy [93], and both galectin-1, -2, -3, and -8 were suggested as potential disease markers and possible targets for bladder cancer therapy [94]. Specifically, galectin-1 is a possible independent prognostic marker of urothelial carcinoma [95], with its positive immuno-expression being significantly correlated with tumour stage, grade, vascular invasion and nodal status [96]. Moreover, galectin-1 mRNA and protein levels are markedly increased in most high-grade bladder tumours compared with low-grade and normal bladder tissue [97, 98]. Furthermore, this glycoprotein is associated with bladder cancer cell invasion by mediating the activity of MMP9 through the Ras-Rac1-MEKK4-JNK-AP1 signalling pathway [95]. Recently, a photodynamic therapeutic approach targeting galectin-1 in bladder cancer cells and xenografts has inhibited tumour growth and enabled selective cytotoxicity in cancer cells, preventing undesired phototoxicity in the surrounding healthy tissues [99]. This study ultimately suggests that galectin-1 constitutes a valid bladder cancer cell biomarker capable of being used in effective targeted therapies. In turn, galectin-3 mRNA and protein levels were also found increased in bladder tumours when compared with normal urothelium [94, 97, 98, 100]. Moreover, galectin-3 levels are increased in invasive tumours compared with non-muscle invasive lesions [101-103]. Furthermore, its expression patterns are also correlated with tumour stage, grade, proliferation (Ki67), apoptosis (apopdetek and bcl-2), and overall survival in patients with T1G3 tumours [101]. These observations suggest a role for galectin-3 as a biomarker for bladder cancer staging and prognosis. In succession, galectin-7 was pointed as a predictive marker of chemosensitivity to cisplatin in urothelial cancer [104]. Finally, the loss of galectin-8 in bladder tumours increases tumour recurrence, while decreased immunohistochemical staining is associated with higher tumour stage and grade [105]. As such, the loss of galectin-8 might be an early step in the development of malignant lesions of the bladder and is a significant independent predictor of recurrence [105].
Several studies have recently pointed out the unique biological properties of basal-like bladder tumour cell subpopulations in their anchorage-independent growth ability and their association to poorly differentiated bladder cancer [106]. In this context, CD44, a member of the transmembrane glycoprotein family commonly implicated in cell-cell and cell-matrix interactions, cell proliferation, differentiation, migration, angiogenesis, presentation of cytokines, chemokines, and growth factors to the corresponding receptors, docking of proteases at the cell membrane, and cell survival [107-109], has been implicated as a cancer stem cell (CSC) marker in several malignancies [110-114]. Particularly, both CD44 and its splicing variants have been involved in bladder cancer carcinogenesis and progression. CD44+ cells exhibit an enhanced capacity to form xenografts in immunocompromised mice as well as chemoresistance compared to CD44− cells [115, 116]. CD44v6, a CD44 isoform containing the CD44v6 exon, has also been shown increased in bladder CSCs [117, 118]. CD44v6 expression on CSCs is supported by a study that correlates CD44v6 expression on bladder cancer cell lines with stem cell properties [119]. Both expression levels of CD44 and CD44v6 were higher in invasive bladder tumours than in pre-invasive tumours and normal urothelium [120]. Also, CD44 and CD44v6 upregulation is associated with higher tumour grade and stage [120, 121]. However, other studies have demonstrated an inverse association between CD44v6 expression and bladder cancer grade and stage [121, 122]. Moreover, the loss of CD44v6 expression was demonstrated as an independent factor for increased recurrence and shorter overall survival [123]. Also, the loss of CD44 expression was associated with shorter progression-free survival [124]. These discrepancies can be explained by the lack of standard immunohistochemical assays, the use of antibodies with different specificities, and differences in the clinicopathological status of bladder tumours used in the different studies. Therefore, integrative and standardized studies are necessary to elucidate the role of CD44 and CD44v6 in bladder cancer, as they hold an important biological and clinical value and may serve as therapeutic targets. In turn, CD44 variant 9 (CD44v9) overexpression has been associated with shorter progression-free and cancer-specific survival in bladder cancer [125], likely impacting invasion and migration via the epithelial-mesenchymal transition (EMT). Therefore, its expression might be a useful predictive biomarker in basal-type muscle invasive and high-risk NMIBC [125]. Nevertheless, the specific glycosylation patterns of CD44 in the context of bladder cancer also remains an open research topic.
Mucins are large membrane-bound glycophosphoproteins, commonly overexpressed in several malignancies [126], including bladder cancer [127-129]. Mucin 1 (MUC1) is restricted to the apical membranes of umbrella cells in normal urothelium, while there is an aberrant MUC1 expression in basal and intermediate layers of neoplastic epithelium [128, 130]. Additionally, the pattern, intensity and depth of MUC1 immunostaining are correlated with bladder cancer grade [129]. Notwithstanding, other study reported no correlation of MUC1 expression with survival, tumour stage or grade [131]. Yet, patients overexpressing MUC1 only had a favourable survival when HER3 was also overexpressed [131]. This may be at least partially explained by the existence of several MUC1 glycoforms, including underglycosylated, sialylated, and fully glycosylated forms. As previously mentioned, several studies have been focusing on the identification of extracellular cell surface markers for urothelial CSCs, envisaging diagnosis and drug targeting. Of note, it has been shown that urothelial CSCs are enriched in an MUC1−CD44v6+ subpopulation of cells. This conclusion was based on the observation that MUC1− and CD44v6+ cells were only present in the basal layer of normal urothelium, which is thought to comprise urothelial stem cells. Subsequently, MUC1− and CD44v6+ cells were isolated, and a slightly increased clonogenicity was observed for these cells compared with unsorted bladder tumour cells [117]. Expression of other mucins such MUC2 and MUC6 were associated with a less aggressive behavior of bladder tumours and demonstrated to be useful predictors of better bladder cancer survival while MUC4 demonstrated an opposite role [129]. In addition, MUC16 STn+ glycoforms, characteristic of ovarian cancers, were recently described for the first time in bladder cancer and demonstrated to be expressed in a subset of advanced-stage bladder tumours facing worst prognosis [132]. Nevertheless, with the exception of MUC16, the specific glycosylation patterns of this class of glycoproteins also remains unknown in bladder cancer.
Integrins are a family of transmembrane adhesion receptors for extracellular matrix components participating in the metastatic cascade. Particularly, normal urothelium presents a polarized expression of alpha6beta4 integrin (ITGA6) on basal cells, while neoplastic urothelium frequently overexpresses this receptor [133]. Moreover, the evaluation of alpha6beta4 integrin tumour expression may provide valuable prognostic information on bladder cancer patients clinical outcome, since patients with alpha6beta4 integrin overexpression hold a significantly worst survival [133]. Throughout EMT-driven carcinogenesis, disseminated cancer cells often acquire a stem cell-like self-renewal capability [134, 135]. Moreover, during EMT, epithelial markers such as ITGAV (αv integrin receptors) are upregulated in several solid tumours [136-138], including bladder cancer with a trend increase in ITGAV expression with disease stage and grade [139]. Furthermore, the functional inactivation of ITGAV (targeting with the integrin receptor antagonist GLPG0187 or knockdown of ITGAV) leads to a less malignant bladder cancer phenotype with significantly impaired migration, EMT response, clonogenicity and a reduction in the size of the stem/progenitor pool. In line with these in vitro observations, knockdown of ITGAV or treatment with GLPG0187 significantly inhibited metastasis and secondary tumour growth [140]. In turn, a central role was also suggested for the beta1-integrin subunit in forming the cell-cell and cell-matrix bonds necessary for adhesion, extravasation and migration of bladder cancer cells [141] through enhanced transmission and generation of contractile forces [142] and possible microenvironmental involvement [69]. Despite its role in bladder carcinogenesis there are also no reports about the specific glycosylation of this class of glycoproteins.
In summary, increased levels of several glycoproteins have been associated with the severity of disease and as part of the molecular signature of more malignant bladder cancer sub-populations. These events not only amplify structural alterations that stem from deregulations in glycosylation pathways but also synergically contribute together with altered glycosylation, to a net effect favouring disease progression. Nevertheless, a comprehensive and context-oriented glycomapping of relevant glycoproteins has not been provided yet, which would be crucial for achieving highly specific cancer biomarkers holding true therapeutic potential. Moreover, the glycomic mapping of relevant glycoproteins may provide highly cancer-specific epitopes in comparison to glycans or glycoproteins alone. This would pave the way for designing more effective targeted therapeutics for more malignant bladder cancer cells.
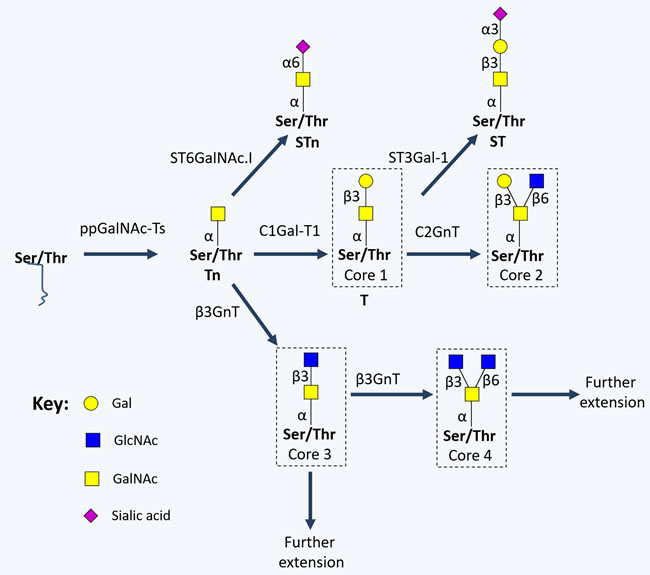
Figure 3: Schematic representation of short-chained O-linked glycan structures. The addition of specific sugar monomers to Ser/Thr residues of a protein backbone begins with the action of polypeptide N-acetylgalactosamine transferases (ppGalNAc-Ts; a family of 20 enzymes, including GalNAc-T1, GalNAc-T2, GalNAc-T3, GalNAc-T4, GalNAc-T5 and GalNAc-T6) giving rise to the Tn antigen, which is generally extended with a Gal residue by C1Gal-T1, originating the Thomsen-Friedenreich or T antigen (core 1). Alternatively, Tn and T antigens can be sialylated by α2,3-sialyltransferases (ST3Gal-Ts) and α-GalNAc ST6Gal-I (ST6GalNAc-I), forming the sialyl-Tn (STn), and sialyl-T antigens. On the other hand, core 1 may be extended originating cores 2-4 by the action of N-acetylglucosamine (GlcNAc) transferases (GnTs; such as GnT-III, GnT-V, core 2 GnTs (C2GnTs) and β3GnT).
Proteoglycan glycosylation
Proteoglycans are structurally and functionally complex glycoconjugates, exhibiting one or more high molecular weight glycosaminoglycan (GAG) chains covalently attached to a protein core [143]. These structures can be found as: i) transmembrane syndecans or glypicans, at the cell surface; ii) hyalectans (aggrecan, versican, brevican and neurocan) or small leucine-rich proteoglycans (decorin, biglycan and lumican) at the extracellular matrix (ECM); iii) basement membrane proteoglycans (perlecan, agrin and collagen XVIII) [144]. Serglycin is the only characterized proteoglycan found at intracellular level, normally in secretory compartments [145].
The biosynthesis and modification of proteoglycans occurs in the Golgi apparatus (GA) through the action of glycosyltransferases, sulfotransferases, epimerases, sulfatases, glycosidases, and heparanases, revealing multiple layers of regulation of these macromolecules [143]. The length and structure of each GAG chain may differ greatly within a certain proteoglycan molecule, while the number of chains linked to the protein core is determined by the number of sugar attachment sites, marked by Ser-Gly dipeptide motifs [143, 146]. The biosynthesis of GAGs, such as chondroitin sulfate, heparan sulfate, dermatan sulfate, hyaluronic acid, and heparin is initiated by the sequential addition of four monosaccharides (Xyl, Gal and GlcA) to a Ser-Gly motif on the core protein. Then, the sugar chains are extended by the addition of two alternating monosaccharides containing an acetylated or sulfated hexosamine (GalNAc, GlcNAc) and uronic acid (GlcA acid or idoA) [143]. In the case of keratan sulfate, the GAG is initiated as N-linked or O-linked repeating disaccharides, and extended by the addition of N-acetyl-glucosamine and galactose residues [143]. Once synthesized, the GAGs are linked to a core protein and proteoglycans are transported from the GA to the cell surface or ECM [144, 147]. Notably, unlike all other GAGs, hyaluronic acid is primarily found as a free sugar chain at the ECM, and its synthesis is epigenetically regulated [148]. Interestingly, hyalectans have the ability to bind hyaluronic acid through their N-terminal globular domain (G1), therefore increasing ECM complexity [149]. Of note, proteins such as MHC class II invariant chain, transferrin receptor, thrombomodulin and CD44 can be considered proteoglycans, since some of their alternative splicing variants present GAG-initiation sites [150]. Other proteoglycans like endocan and versican also present alternatively spliced forms with variable sugar modifications [150]. In particular, a versican variant without chondroitin sulphate attachment sites has been described, [149].
Proteoglycans present high affinities for various ECM constituents and cell adhesion molecules, playing a crucial role in intercellular interactions [144]. These glycoconjugates can also bind growth factors, cytokines and chemokines, allowing them to escape proteolysis. Some can also act as co-receptors for growth factors and tyrosine kinase receptors, changing the duration of their signaling reactions or lowering their activation thresholds [143, 144]. Therefore, the altered expression of proteoglycans, including syndecan-1, neuropilins, versican, chondroitin sulfate proteoglycan 6, decorin, biglycan, endocan, hyaluronic acid and its metabolic enzymes, has been linked to several cancers and, specifically, with bladder cancer carcinogenesis, metastasis and prognosis (Figure 4, Supplementary Table 1).
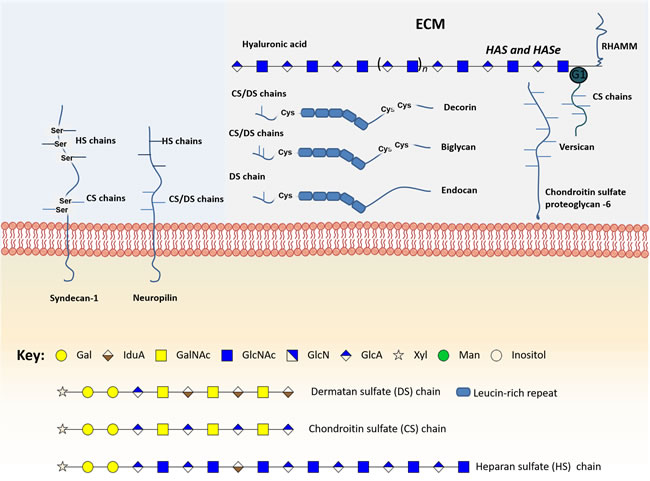
Figure 4: Schematic representation of the main glycomolecules with biological relevance in bladder cancer. The figure represents specific proteoglycans that have one or more glycosaminoglycan (GAG) chains, consisting of linear co-polymers of acidic disaccharide repeating units such as chondroitin sulfate (DS), heparan sulfate (HS) and dermatan sulfate (DS). These glycomolecules can be found attached to the outer leaflet of the plasma membrane (Syndecan-1) or in the extracellular matrix (Versican, Chondroitin sulfate proteoglycan-6, decorin and biglycan). Particularly, some of these structures can bind to each other through their N-terminal globular domain (G1), therefore increasing extracellular matrix (ECM) complexity. Of note, hyaluronic acid is the only GAG primarily found as a free sugar chain in the extracellular matrix. Hyaluronic acid synthases (HAS) and Hyaluronidases (HAse) constantly degrade and remodel hyaluronic acid molecules largely affecting ECM dynamics. Some glycoproteins can also be found linked to the cell membrane through a glycosylphosphatidylinositol (GPI) anchor, an example is glypican-3.
Cancer-associated transmembrane proteoglycans
Syndecans are a family of heparin sulfate proteoglycans, commonly presenting three to five heparin sulfate chains, and are known to modulate cellular adhesion, migration, proliferation, differentiation, and growth factor signaling [151]. These macromolecules are commonly found at bladder cancer cell surfaces, along with other transmembrane proteoglycans and glypicans [152, 153]. Syndecans can also be found in their soluble form, due to a post-translational modification causing the release of their ectodomains through juxtamembrane region proteolysis [154]. Particularly, syndecan-1 (CD138), frequently expressed in epithelial cells and some leukocytes [155], was found to be increased in bladder cancer patients serum and stroma, especially in muscle-invasive cases [156-158]. Serum overexpression of syndecan-1 was associated with lymph node metastasis, while stromal overexpression was related with poorer overall survival [158]. The loss of transmembrane syndecan-1 expression in tumour cells was related to higher tumour stage and grade [159, 160], as well as reduced recurrence-free survival in bladder cancer [160, 161]. Still, high-grade superficial, and deep invasive bladder carcinomas were also characterized by elevated expression of syndecan-1, while low-grade and non-invasive phenotypes do not [161]. Cytoplasmic overexpression of syndecan-1 in cancer cells, often accentuated close to the nucleus, was demonstrated in Ta tumours compared to normal urothelium, suggesting a failure in intracellular trafficking caused by the loss of functional syndecan-1 [162]. This directly affects carcinogenesis through the loss of cellular adhesion properties, thereby promoting more invasive phenotypes [160]. Also, syndecan-1 altered expression can affect tumour cells via junB-FLIP long signals, involving apoptosis resistance and increased proliferation [161]. Simultaneous loss of syndecan-1 expression in tumour cells and its overexpression in high-stage and high-grade bladder cancer patients serum suggest the importance of syndecan-1 in tumour progression; therefore, this molecule could be a new therapeutic target in human urinary bladder cancer [158].
Neuropilins are co-receptors of two structurally and functionally unrelated ligands classes, the class 3 semaphorins and selected VEGF family members [163]. Neuropilin-1 has multiple heparan and/or chondroitin/dermatan sulfate GAG chains [163]. Recent reports demonstrate neuropilin-1 expression on non-endothelial cells in bladder urothelium [164], as well as its overexpression in high grade/stage bladder tumours [165]. Moreover, neuropilin-1 upregulation was associated with shorter overall survival in bladder cancer patients [165]. In addition, neuropilin-2 is expressed in neural and endothelial cells and, upon ligand stimulation, induces neural development and the growth of newly formed blood and lymphatic vessels [163]. Overexpression of neuropilin-2 demonstrated to have prognostic value in bladder cancer, as it was associated with shorter overall and cancer-specific survival and earlier cancer-specific death after transurethral resection and radiochemotherapy [166]. Additionally, the co-expression of neuropilin-2 and the family member VEGF-C is also a prognostic marker for overall survival of bladder cancer patients [166]. Therefore, syndecan-1 and neuropilins may play an important role in the progression of bladder cancer and their altered expression may serve as a biomarker for prognosis.
Cancer-associated extracellular matrix proteoglycans
Versican, also known as chondroitin sulfate proteoglycan 2, a central component of cancer-related inflammation, is highly expressed in metastatic bladder carcinomas and its overexpression is correlated with poor survival [167]. In tumour cell lines, versican overexpression was associated with increased cell migration and tumour stage [168]. A correlation between versican overexpression, RhoGTP dissociation inhibitor 2 (RhoGDI2) underexpression, metastasis and poor clinical outcome was also demonstrated [167, 169]. Particularly, RhoGDI2 underexpression and versican overexpression are associated with metastasis through the involvement of macrophages and the CCL2/CCR2 signaling axis [167, 169]. In fact, RhoGDI2 is a regulator of several Rho GTPases that play important roles in cell cycle progression, neovascularization, invasiveness, and metastasis [170]. Therefore, targeting this mechanism may provide novel therapeutic strategies for delaying the appearance of clinical metastasis [170].
The role of decorin, a key component of the tumour stroma, in cancer progression and its therapeutic potential has been the focus of several studies. Increased secretion of decorin in the MB49/MB49-I murine bladder cancer model and in muscle-invasive tumours was associated with the promotion of angiogenesis and tumour cell invasiveness [171]. Nevertheless, other studies demonstrate a possible tumour suppressor role for decorin, where bladder tumour tissues are entirely devoid of decorin expression while non-malignant stromal areas express this proteoglycan [172, 173]. A mechanism through which decorin exerts its tumour suppressor role has been proposed, where decorin may act as a natural antagonist of the oncogene insulin-like growth factor receptor I (IGF-IR) [173, 174]. Therefore, in bladder tumours, the loss of decorin expression eliminates IGF-IR activity and signaling repression, promoting cellular motility, invasion, and cancer progression [173, 174].
Biglycan is a small leucine-rich proteoglycan with immune and growth factor activity modulating properties, as well as matrix assembly involvement [175]. This proteoglycan has been demonstrated to be overexpressed on invasive bladder cancer tissue [172, 176]. Interestingly, while biglycan overexpression is associated with higher tumour stages and muscle invasiveness, it’s up-regulation was related with tumour cell proliferation inhibition and increased patients’ 10-year survival [176].
Endocan, also known as endothelial cell-specific molecule 1, is a secreted proteoglycan that has a single dermatan sulfate side chain attached to serine 137 and demonstrated to be highly elevated on tumour vessels from invasive bladder cancer tissues [177]. Moreover, its expression correlated with stage, and invasiveness as well as predicted a shorter recurrence-free survival time in non-invasive bladder cancers [177]. Therefore, endocan expression impacts the prognosis of bladder cancer patients and, as described ahead, also is a possible diagnosis marker.
Hyaluronic acid (HA), an unsulfated anionic linear GAG, its implicated in cell adhesion, migration and angiogenesis [178]. Particularly, hyaluronidases (HAse) are enzymes that hydrolyze HA molecules into small angiogenic fragments, participating in the degradation of tumour surrounding ECM, and enabling cancer cells invasion and dissemination [178]. As such, HA, HAses (e.g. HYAL1), hyaluronic acid synthases (HAS) 1, 2 and 3, as well as hyaluronic acid receptors (e.g. CD44 and receptor for hyaluronan-mediated motility, RHAMM) have been suggested as possible diagnosis and prognosis biomarkers. Also, both HAS, HYAL1, CD44 and RHAMM were found to be overexpressed in bladder cancer tissues [179, 180]. In addition, HYAL1 expression was also correlated with disease-specific mortality and recurrence [179, 181]. Finally, elevated expression of RHAMM was found in invasive bladder tumours and associated with poor prognosis, due to increased tumour cell proliferation and shorter overall and disease-specific survival [180].
Alterations in cancer-associated proteoglycans were demonstrated as associated with bladder cancer progression and may present prognosis value, yet more studies are necessary in order to confirm these associations and transpose these markers to clinical practice.
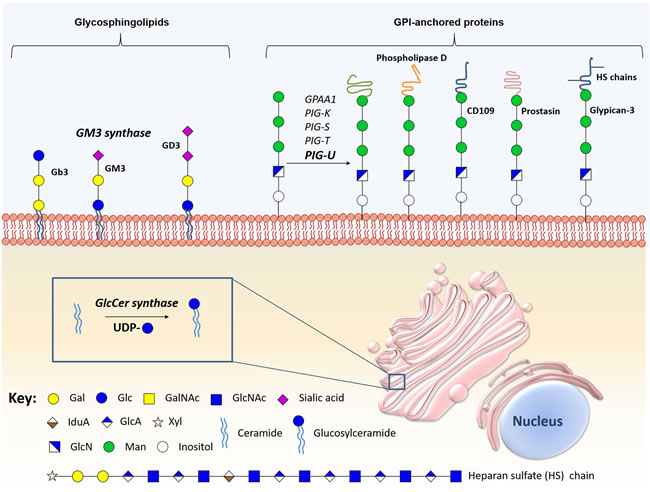
Figure 5: Schematic representation of the main biologically relevant glycosphingolipids and glycosylphosphatidylinositol-anchored proteins in bladder cancer. The figure represents certain glycosphingolipids, especially the sialic acid-containing glycomolecules, glycosylphosphatidylinositol-anchored proteins, and the enzymes implicated in the synthesis and hydrolysis of these conjugates have been implicate in bladder cancer malignancy.
Lipid glycosylation
Glycolipids are a major class of glycoconjugates that include glycosphingolipids (GSLs) and GPI anchors.
Alterations in sphingolipids’ glycosylation
Glycosphingolipids (GSLs) are neutral or anionic molecules composed by a hydrophilic glycan covalently β-linked via glucose (glucosylceramide) or galactose (galactosylceramide) to the terminal hydroxyl group of a hydrophobic ceramide backbone [182, 183]. Specifically, GSL biosynthesis is initiated in the ER with the condensation of sphingosine and acyl-CoA by a group of six ceramide synthases, giving rise to a long-chain amino alcohol base (sphingosine) in amide linkage to a fatty acid, namely a ceramide lipid [183, 184]. Ceramide can then be galactosylated by galactosylceramide synthase, to produce galactosylceramide, which in turn can be transported to the GA where it is sialylated to produce GM4 ganglioside, or sulfated to produce sulfogalactolipids [185-187]. Also, ceramide is frequently glucosylated in the GA by glucosylceramide synthase to form glucosylceramide, the core structure of 90% of GSLs [185-187]. Subsequently, the C-4 hydroxyl of glucosylceramide can be galactosylated by β4-galactosyltransferases V and VI, forming lactosylceramide [188]. Once produced, lactosylceramide will serve as the metabolic precursor of more than 300 structurally different classes of complex GSLs through the action of specific glycosyltransferases and sulfotransferases, depending on nucleotide sugar donors availability [189]. Particularly, lactosylceramide is a template for: 1) GA2, through β,4-N-acetylgalactosylaminyltransferase (B4GALNT1) activity; 2) GM3 ganglioside, through α-2,3-sialyltransferase (ST3GAL5Gb3); 3) Gb3, by α-1-4-galactosyltransferase (A4GALT) activity; and, 4) Lc3, by the β-1,3-N-acetylglucosaminyltransferase (B3GNT5) [189]. GSLs glycan chains can be further extended and terminated with structural moieties similar to those found in glycoproteins, namely and Lewis blood type antigens. After synthesis, these structures leave the GA and are redirected to the plasma membrane [190], constituting approximately 5% of all membrane lipids
GSLs are also implicated in key cellular functions, such as cell adhesion, proliferation, differentiation, apoptosis, motility and immune recognition [191-193]. Particularly, ceramide is implicated in apoptosis and regulates several cell cycle and senescence pathways [194]. Consequently, GSLs have received considerable attention as promising biomarkers for disease progression, as well as pharmacological targets for bladder cancer therapy. Pioneering studies demonstrated that glucosylceramide and glucosylceramide synthase are overexpressed in several multidrug resistant cancer cell lines, being related with drug resistance [195-197]. Particularly, in bladder cancer, glucosylceramide synthase overexpression was demonstrated to be associated with higher histologic grade [198]. In accordance, the overexpression of this enzyme is an indicator of poor prognosis, showing associations with lymph node metastasis, reduction in the 5-year overall and disease-free survival [198]. The glycosphingolipid composition of human bladder cancer tissue has been assessed, showing large amounts of ganglioside GM3 in superficial bladder tumours, but not in invasive tumours [199]. This overexpression can be caused by the simultaneous overexpression of GM3 synthase and downregulation of both Gb3 and GD3 synthases [199]. Moreover, high levels of GM3 are associated with reduced invasive potential [199], proliferation, motility, tumour growth and increased apoptosis [200]. Since exposure to exogenous GM3 have been proved to inhibit tumour cell lines proliferation and adhesion, this approach was proposed as bladder cancer therapy. Also, the direct instillation of GM3 in orthotopic models inhibited tumour growth [201]. It has also been reported that the expression of GM2, GM3, or GM2/GM3 complexes inhibited bladder cancer cell motility and growth [202] (Figure 5, Supplementary Table 1).
Alterations in glycosylphosphatidylinositol-anchored molecules
The anchoring of proteins and proteoglycans to cell membranes through the lipid portion of a GPI anchor is a conserved post-translational modification [203]. These anchors present conserved core structures consisting of ethanolamine phosphate, three mannose monomers, and a non-N-acetylated glucosamine attached to a inositol phospholipid (EtNP-6Manα-2Manα-6Manα-4GlNα-6myoInositol-P-lipid) [204]. This backbone can be modified with phosphoethanolamine and/or various glycan side-branches. More detailed information about the biosynthesis of this class of glycans may be found in [204-206] and has been summarized in Figure 5.
Alterations in several enzymes involved in glycosylation of GPI-anchored molecules such the multi-protein transaminase complex were first mentioned as relevant in cancer after the discovery of the oncogenic activity of PIG-U in bladder cancer [207, 208]. This was demonstrated through the induction of tumourigenesis mediated by PIG-U overexpression in mice [208]. In addition, PIG-U overexpression in vitro was correlated with increased cell proliferation and upregulation of GPI-anchored proteins, such as urokinase receptor, increasing STAT-3 phosphorylation and subsequent cellular migration, and apoptosis [208]. PIG-U overexpression is also associated with higher tumour grade and muscle invasion, suggesting its role in tumour development and progression [209]. Moreover, overexpression of PIG-U is an independent predictor of recurrence for superficial bladder cancer [209]. Consequently, the expression of PIG-U and other multi-protein transaminase complex subunits was explored and confirmed in different cancer types using microarrays [210].
GPI-anchored proteins are almost exclusively located on cell surfaces, and are functionally diverse, presenting key roles in cell-cell interaction, adhesion, host defense, and signaling transduction [204, 211]. Thus, these proteins have been explored regarding their potential as biomarkers for carcinogenesis and metastatic potential in bladder cancer (Figure 5, Supplementary Table 1). The expression of GPI-specific phospholipase D, a highly specific GPI-anchored enzyme, is significantly increased in highly malignant murine bladder carcinoma cells when compared to less malignant controls [212]. In addition, CD109, a GPI-anchored glycoprotein that negatively regulates the transforming growth factor (TGF)-β/Smad signaling in vitro, is overexpressed in the basal layer of NMIBC and low-grade tumours. Interestingly, CD109 shows a similar expression pattern to cancer stem cell marker CD44, and its overexpression was associated with better cancer-specific survival [213]. Prostasin, a GPI-anchored serine protease crucial for epithelial differentiation [214] and epidermal growth factor receptor (EGFR) proteolysis [215], was shown to be downregulated in high-grade urothelial bladder cancer cell lines [216]. This loss of expression was associated with EMT, marked by a reduced E-cadherin expression and loss of epithelial morphology, which may have implications in the invasive potential and resistance to anti-EGFR therapy [216]. However, the clinical relevance of prostasin was not yet evaluated in bladder cancer. Notwithstanding, multi-protein transaminase complex subunits and GPI-anchored proteins have potential to serve as markers of tumourigenesis and metastatic capability, being relevant targets for bladder cancer therapy.
Beyond GPI-anchored proteins, alterations in glypicans, a class of GPI-anchored proteoglycans, also have been studied in cancer. Glypicans are a family of GPI-anchored heparan sulfate proteoglycans known to interact with growth factors through heparan sulfate chains [217]. This class of proteoglycans is predominantly expressed during fetal development, being critical to organogenesis [218]. Moreover, glypicans were described in several cancers [219, 220], including bladder cancer. Particularly, glypican-3 is expressed in squamous cell and invasive urothelial carcinomas; however, it is not a good biomarker for diagnosis using tumour tissues [217]. Furthermore, glypican-3 expression was not associated with tumour stage, grade, lymph node metastasis, concomitant CIS, soft tissue surgical margins, disease recurrence or cancer specific mortality after radical cystectomy [221].
In conclusion, alterations in PIG-U and GPI-anchored proteins seem to be promising regarding their prognosis value while GPI-anchored proteoglycans appear to be bad prognosis biomarkers. Notwithstanding, further studies are necessary to evaluate these GPI-anchored molecules and enzymes from multi-protein transaminase complex in bladder cancer biological and clinical context.
Prognosis glycomarkers for bladder cancer
The most challenging aspect in bladder cancer management stems from its highly heterogeneous nature, irrespectively of disease stage. This constitutes a true obstacle for individualized therapeutic decision, disease monitoring and prognosis with tremendously negative impact on patient care and life expectancy. In this context, several clinical studies have unveiled glycan-associated signatures (glycans, glycan binding proteins, glycosyltranserases, heavily glycosylated proteins/lipids/GPI-anchored molecules) associated with more aggressive cancer cell phenotypes, tumour recurrence, progression, metastization, or decreased overall survival. Particularly, in bladder cancer, the presence/overexpression of GnT-III/IV, Lex, Ley, SLex, STn, ST6GalNAc.I, T-antigen, ST3Gal.I, HER2, EpCAM, galectin-1, galectin-3, CD44, CD44v9, MUC1, MUC4, ITGA6, ITGAV, neuropilin-1, neuropilin-2, versican, decorin, biglycan, endocan, HYAL1, hyaluronic acid synthase 1, RHAAM, glucosylceramide synthase, and PIG-U was associated with more aggressive phenotypes and/or poor prognosis [20, 30, 46, 50, 54, 64-67, 76, 82, 86, 87, 96, 97, 101-103, 120, 121, 124, 125, 129, 133, 139, 165-168, 171, 172, 176, 177, 179-181, 198, 208, 209]. By another hand, (over)expression of β-1-6GalNAc antennae, galectin-7, CD44v6, MUC2, MUC6, glycolipid enzyme GM3, and CD109 were linked to a less aggressive phenotype and/or better prognosis [32, 104, 121, 122, 129, 199, 213]. In turn, the loss of GnT-V, ABO(H) terminal structures, sLea, Tn, galectin-8 and prostasin is associated with more aggressive cancer phenotypes, making them potential markers of poor prognosis [31, 32, 44-47, 50, 67, 105, 158-160, 200, 216]. Yet, glypican-3 tissue expression was not associated with aggressive phenotype nor prognosis [217, 221] and studies for syndecan-1 demonstrated contradictory results [158-162, 222], warranting elucidation of its prognostic and diagnostic role. For other molecules, the co-expression with another allowed to have or enhanced a significant prognostic and therapeutic outcome impact, such as Lea/Leb antigens [44, 49], s6T/STn [75] neuropilin-2/VEGF-C [166], Gb3/GD3 synthases [199] and GPI-specific phospholipase D/H-ras oncogene [212] (Figure 6 and 7, Supplementary Table 1).
While these studies support the potential of glycans for patient stratification significant bias hampers conclusive remarks to support introduction in clinical practice. Namely, most studies differ in cohort size and distribution of the cohort by bladder cancer type, stage and grade, which often translate into conflicting results. Moreover, most studies disregard patient ethnicity, lack endpoints standardization and rely on different biological samples, as well as in different sample processing and analysis techniques. Nevertheless, several molecular markers hold promise for becoming widely available and cost-effective tools for a more reliable risk assessment. Thus, efforts need to be conducted to validate glycobiomarkers in larger series for prospective use, ideally in the context of prospective clinical multicenter randomized trials, using current clinicopathological parameters for risk assessment. The inclusion of relevant glycobiomakers in current patient stratification parameters may help accurately assess patient’s prognosis and response to different treatment options. Therefore, future studies should also evaluate the impact of glycomarkers in the therapeutic outcome and as novel targets for bladder cancer therapy. Finally, recent evidences support the need to integrate this knowledge in multiplex risk assessment tool combining standard clinicopathological factors with molecular markers [223] envisaging individualization of responses. Although much effort has been spent on glycobiomarkers research, clinicians and medicinal chemists rarely consider glycans as biological targets or drugs [224], hindering advances in bladder cancer management and in the development of targeted therapeutics. Notwithstanding, this unfamiliarity is beginning to change as improved methods for carbohydrate synthesis [225-227], sequencing [228, 229], and biological analysis [230-232] become more sensitive and widely available.
Exploring glycans for non-invasive bladder cancer detection
Non-invasive disease detection remains a major challenge despite the pressing need for tools capable of reducing the burden of disease-follow up, treatment monitoring and early detection [233]. The active secretion of cancer-associated glycoconjugates into bodily fluids, such as urine and blood, or shed from apoptotic and necrotic cancer cells holds tremendous potential to address this challenge [7, 51]. Accordingly, several FDA-approved cancer biomarkers for non-invasive cancer detection, follow-up and prognosis are either glycans, such as CA19-9 (SLea), or heavily glycosylated glycoproteins, such as CA125 (MUC16), CA15-3 (MUC1), CA-72-4 (tumour-associated glycoprotein 72, TAG-72), PSA (prostate-specific antigen), and CEA (carcinoembryonic antigen) [233]. In bladder cancer, serum CA19-9 is a marker of aggressiveness and advanced stage disease, being almost invariably raised in patients with metastatic cancer. As such, it constitutes a valuable marker of poor prognosis [234]. Notwithstanding, its value as a screening tool has been opposed by its low sensitivity (29%) [234]. On the other hand, urinary CA19-9 is a better screening parameter, with optimum sensitivity and specificity, than its serum counterpart for diagnosis of low grade and early stage bladder cancer. Furthermore, it can be suggested that urinary CA19-9 can be used as better prognostic marker for low grade bladder cancer than its serum equivalent [235]. Moreover, urinary CA19-9 levels could be a new effective diagnostic tool for bladder cancer patients with both Le and Se alleles. Particularly, 70% of bladder cancer patients with both Le and Se alleles presented CA19-9 levels over the cut-off value, and only 16% of patients with other urological conditions were over the cut-off [236]. Furthermore, simultaneous elevation of CA19-9 and CEA serum levels correlated with tumour invasion and grade in patients with CA19-9-expressing urothelial carcinomas [53]. In addition, SLea antigen has been observed in bladder dysplasia, Tis, non-invasive, and invasive carcinomas of the bladder [53, 236, 237], suggesting that these patients may present elevated CA19-9, which warrants confirmation in broad clinical studies.
Considering the cancer-associated glycoproteins, galectins also hold potential for bladder cancer detection. In fact, bladder cancer patient’s serum levels of galectin-3 are considerably higher than control groups, and are correlated with tumour type, stage and grade [98, 100, 103]. Particularly, patients with high-grade urothelial carcinoma have higher serum levels of galectin-3 than those with low-grade tumours [103]. Moreover, patients with muscle-invasive tumours also have higher serum levels of galectin-3 than those with Ta tumours [103], conferring to this glycoprotein a diagnostic and stratification value for bladder cancer patients. In addition, serum levels of galectin-3 also have diagnostic value for bladder squamous cell carcinoma (SCC) patients [103]. More recently, a multiplexed immunosensor has been developed for the detection of specific biomarkers galectin-1 and lactate dehydrogenase B present in different grades of bladder cancer cell lysates. This approach has allowed not only the identification of different grades of bladder cancer cells but also the real time detection of multiple analytes on a single chip, providing more practical benefits for clinical diagnosis [238].
Regarding proteoglycans, syndecan-1 was also explored as a diagnosis biomarker in a non-invasive urine-based assay, however with low predictive value (sensitivity of 70%, specificity of 48% and accuracy of 59%) [222]. Therefore, further validation is warranted in larger and prospective studies. The chondroitin sulfate proteoglycan 6, also known as structural maintenance of chromosomes 3 (SMC3), has also been found overexpressed in bladder cancer. Moreover, when used in combination with six gene transcripts (insulin-like growth factor-binding protein 7, sorting nexin 16, cathepsin D, chromodomain helicase DNA-binding protein 2, nell-like 2, and tumour necrosis factor receptor superfamily member 7), it could discriminate bladder cancer from control blood samples with a sensitivity of 83% (95% confidence interval, 67-93%) and a specificity of 93% (95% confidence interval, 76-99%), constituting a possible blood diagnosis biomarker [239]. Beyond the prognosis role of endocan, recently this proteoglycan also demonstrated non-invasive diagnosis potential for bladder cancer. In fact, serum expression of endocan could discriminated bladder cancer patients with a sensitivity of 50% and specificity of 77% while the urinary endocan expression resulted in a sensitivity of 62% and specificity of 71% [240]. Moreover, HA and its related enzymes may have potential for detecting bladder cancer patients. In fact, increased HA levels are detected in all bladder cancer grades, while HAse levels are preferentially elevated in urothelial bladder cancer grades 2 and 3 [241, 242]. Lokeshwar et al. and Passerotti et al. have reported an optimal sensitivity (91.9% and 81.9%, respectively) and specificity (92.8% and 80.5%, respectively) of the HA test for bladder cancer detection [243, 244]. Urinary HAse measurement demonstrated a 100% sensitivity and 88.8% specificity to detect grade 2 and 3 bladder tumours [242]. Measurements of both urinary HA and HAse (the HA-HAse test) demonstrated an 89% sensitivity and 83% specificity for detecting urothelial bladder cancer [241]. Similarly, other study evaluated this non-invasive method in urine samples, showing that the HA test has 83.1% sensitivity, 90.1% specificity and 86.5% accuracy to detect bladder cancer [245]. Also, the urinary HAse test demonstrated 81.5% sensitivity, 83.8% specificity and 82.9% accuracy in detecting grade 2 and 3 bladder cancer [245]. Other studies also reported high sensitivity and specificity for detecting bladder cancer using both urinary HA and HAse by a non-invasive approach [246-249]. In fact, the sensitivity of HA-HAse test demonstrated to be superior to ImmunoCyt® and cytology (83.3% versus 63.3% and 73.0%, respectively), as well as to BTA STAT® test (94.0% versus 61.0%). In turn, specificity was comparable between HA-HAse and ImmunoCyt® or cytology (78.1% versus 75.0% and 79.7%, respectively) and between HA-HAse and BTA STAT® test (63.0% versus 74.0%) for detecting bladder cancer or bladder cancer recurrence [248, 249]. The evaluation of HAS1 and HA expression resulted in a 79% and 88% sensitivity, as well an 83.3% and 100% specificity, respectively, for detecting bladder cancer. Moreover, both expressions correlated with a positive HA urine test [247]. In addition, the combined expression of HAS2-HYAL1 detected bladder cancer with overall sensitivity of 85.4% and 79.5% specificity, predicting recurrence within 6 months [179] (Figure 6).
Despite significant research efforts and promising upfront results, neither glycans nor glycoconjugates have yet been approved for non-invasive bladder cancer detection. Again, reduced study dimensions, biased patient series and variations in detection methods are amongst the factors hampering the generalization of these approaches. As such, comprehensive clinical studies on the glycome, glycoproteome and glycolipidome of bodily fluids are warranted to broaden our understanding about alterations accompanying malignant transformations, disease progression and dissemination. Moreover, efforts should be undertaken to incorporate glycans in broad biomarker panels envisaging highly sensitive and specific detection methods.
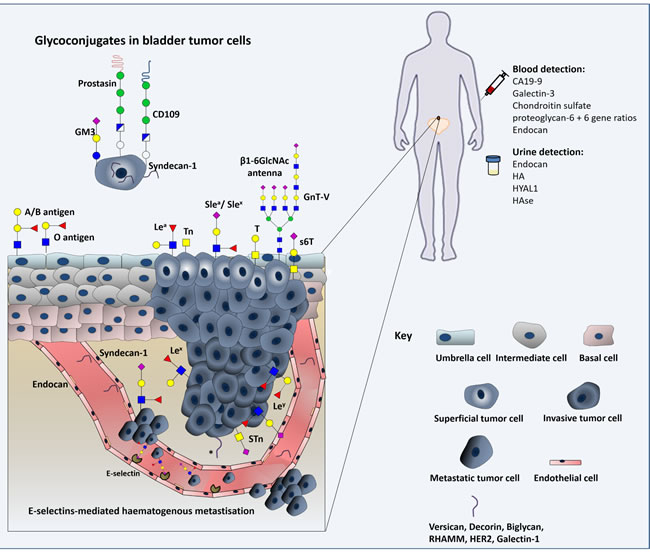
Figure 6: Schematic representation of the glycomolecule-mediated metastization model and diagnostic value of glycans. Herein we represent the process of tumour cell invasion, dissociation and metastization in which glycans interfere with cell-cell adhesion and haematogenous tumour cell spread. We emphasize the modification of epithelial cadherin with β1,6-N-acetylglucosamine (β1,6GlcNAc)-branched N-glycan structures, the loss of ABO(H) blood group determinants, changes in Lewis antigens patterns, and the oversialylation of glycans resulting in the over-expression of simple mucin type O-GalNAc glycans. Furthermore, expression of glycolipids, proteoglycans and gangliosides in cancer cell membranes can modulate signal transduction, activating various cellular pathways that induce tumour growth and progression. As such, some of these relevant glycomolecules are represented as well. The diagnostic value of some of these macromolecules is also highlighted.
Glycomics and Glycoproteomics: insights towards precision medicine
The establishment of clinically useful glycan-based/assisted molecular models have been significantly delayed by serious analytical limitations. In fact, the majority of the studies presented to this date are target-directed and based on immune assays with antibodies and/or lectins whose specificity for the target ligands has not been fully disclosed yet. These aspects delay the generalization of glycans biomarker potential. Moreover, most studies are one-dimensional, failing to provide a comprehensive overview of the glycome in clinical settings. The low abundance of relevant glycoforms in biological milieus also poses a major analytical difficulty, which adds to the significant structural diversity and complexity presented by this class of biomolecules.
In the past decade, mass spectrometry has emerged as a core analytical technology to interrogate the glycome due to the rapid advance in resolution, mass accuracy, sensitivity, and reproducibility provided by modern hybrid mass analyzers. Currently, glycans and glycoconjugates analysis relies on hyphenated techniques comprehending separations by liquid chromatography or capillary electrophoresis, as well as detection by electrospray tandem mass spectrometry (ESI-MS) and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-MS). Complementary tandem experiments are required for unambiguous assignments. Even though, the identification of isomeric and/or isobaric species remains a challenging task. Nevertheless, it has been demonstrated that certain isomeric glycans produce characteristic product ion spectra that can be used for identification, irrespectively of the type of mass spectrometer [250, 251]. Moreover, the introduction of graphitized LC columns has significantly improved the analysis of native glycans by mass spectrometry [252]. More in depth insights on analytical advances in glycomics may be found in recent reviews on the subject [253-255]. Recent developments in matrix-assisted laser desorption/ionization (MALDI) mass spectrometry imaging (MSI) on formalin-fixed paraffin-embedded (FFPE) tissue sections may also provid a key tool for evaluating spatio-temporal investigation of glycosylation changes in cancer tissues. [269]. However, while the field still struggles with technical difficulties associated with variations in accuracy depending on analytical method and variety of mass spectrometry architectures, significant efforts are ongoing to standardize protocols and implement robust glycoanalytical platforms [256, 257]. In addition, rapid expansion of high-throughput mass spectrometry studies has generated significant amounts of experimental “omics” data that require more sophisticated bioinformatics tools and databases. In this context, several groups have started the development of algorithms for computerized annotation of mass spectra and fragmentation data, as revised by Hu et al. [258]. However, while significant advances have been observed for glycan analysis, glycopeptide data interpretation remains immature compared to proteomics data analysis. This is partly due to lack of consensus regarding the best way of estimate the false discovery rate, and the existence of multiple formats of data storage. Nevertheless, guidelines for reporting mass spectrometry-based glycoanalytic data are being developed [257, 259].
Despite the enormous potential of glycomics, few quantitative and comprehensive studies were conducted on bladder cancer. Yang et al. quantitatively analyzed and compared glycan expression patterns in normal and bladder cancer cells through an integrated methodology using lectin microarray and mass spectrometry [58]. It has been demonstrated that SLex and high mannose-type N-glycans were highly expressed in bladder cancer cells [58]. In addition, a high expression of core-fucosylated N-glycans but a low expression of terminally fucosylated N-glycans was observed in bladder cancer cells [58]. In turn, Pocheć et al. determined the N-glycan patterns of integrin α3β1 in bladder cancer cells compared to normal bladder cells using an integrated methodology of lectin-binding assays and mass spectrometry [260]. Accordingly, bladder cancer cell-associated integrins have been found to express high-mannose, hybrid and predominantly complex type N-oligosaccharides, as well as the sialylated tetra-antennary complex type glycan Hex7HexNAc6FucSia4 [260]. Weather by focusing on the whole proteome or in a single glycoprotein, both studies have given an important in vitro view of the glycome pattern of bladder tumour cells compared to a normal state, paving the way for new and more comprehensive studies.
In summary, early glycan-based clinical studies have created the molecular basis to drive the emerging area of advanced glycomics in bladder cancer. We expect that the generalization of these approaches leads to the discovery of key glycan biomarkers with clinical and therapeutic potential in the near future. The incorporation of glycogenomics and glycoinformatics datasets are expected to accelerate a comprehensive understanding of the glycome. Furthermore, the integration with other “omics” will be crucial to deepen the understanding of glycosylation’s role in human systems and provide models capable of disclosing the polymorphic nature of disease and ultimately help tailoring medical decisions and achieve precision medicine settings.
Concluding remarks and future perspectives
Bladder cancer is a heterogeneous disease encompassing distinct biological features and clinical outcomes. This is responsible for elevated recurrence rates, often accompanied by disease progression facing existing treatments. Moreover, it has hampered the establishment of precision medicine settings capable of molecular-based individualization of disease management. These aspects make bladder cancer one of the costliest malignancies to manage, constituting a burden to both patients and healthcare systems. We believe that true advances in this field will require an integrative panomics approach capable of providing robust models for molecular-based patient tailored clinical decisions. So far, most efforts have been put in genomics, transcriptomics and proteomics fields, with upfront enthusiastic results; however, over 40 years of glycobiology research has yet to retrieve solid evidences capable of boosting clinical implementation. Even though many studies have highlighted glycans and glyconjugates (glycoproteins, glycolipids and proteoglycans) holding true clinical potential, few have engaged in a comprehensive interrogation of the glycome. Notwithstanding, the relevance of these molecular entities for disease progression and dissemination suggests potential for more in depth targeted omics studies. Moreover, the standardization of glycomics protocols backed by high-throughput analytical and novel bioinformatics tools opens now a unique opportunity for real advances in this area. Therefore, the field must now focus on large scale multicentric translational studies integrating glycomics data with novel molecular findings, including recently proposed models for bladder cancer risk-stratification [261-263]. Moreover, our understanding on the glycobiology of chemoresistance, formation of the pre-metastatic and metastatic niches is still scarce and warrant careful evaluation on the near future. Complementary understanding of glycan biosynthesis pathways and the biological significance of these alterations will be warranted, envisaging theragnostic applications. Taking into consideration the glycome’s strong dependence on the microenvironment and physiological status, significant efforts should put on developing models capable of recreating tumour glycoheterogeneity. Patient derived xenografts have been proven useful in this context [264] and may constitute valuable tools for understanding the dynamics of glycosylation in malignancy as well as for the identification of prognostic glycobiomarkers. Ultimately, this will be of key importance for developing targeted therapeutics while exploring the cell-surface nature of glycans. Such approaches would strongly benefit from the identification of glycoconjugates (proteins, peptides or lipids) yielding cancer-associated carbohydrate antigens, which would significantly narrow biomarker specificity for malignant cells. While the development of novel high-affinity glycan ligands, namely humanized monoclonal antibodies and antibody fragments, for theragnostics applications still poses a major task due to the complex structural nature of these molecules, several advances in cell glycoengeneiring and glycosynthesis [265-267] hold potential to overcome these limitations. Finally, it has been highlighted that glycans play a key role in immune modulation, especially by favoring tumour tolerogenic mechanisms [21, 268]. A deeper understanding of influence of glycosylation in immunological mechanisms is a hot research topic that will pave the way for circumventing these events and for more effective and less toxic immunotherapies. In summary, an intervention roadmap has been established to boost glycobiology towards omics settings capable of generating key data to improve the management of bladder cancer patients.
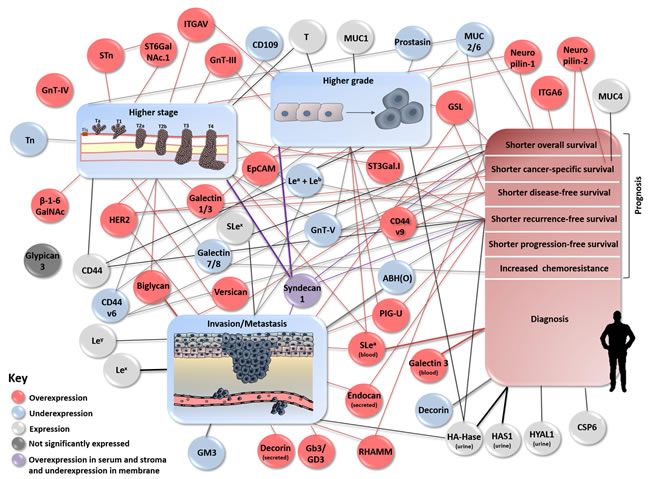
Figure 7: Schematic representation describing associations between (altered) expression of glycans and glycoconjugates and bladder tumour stage, grade, invasion/metastasis, patients’ diagnosis and prognosis. The figure clusters using a gradient of colored circles and lines the biological and clinical role of the altered expression of glycans and glycoconjugates in bladder cancer.
Abbreviations
BCG: bacillus Calmette-Guérin; CSC: cancer stem cell; ECM: extracellular matrix; EGFR: epidermal growth factor receptor; EMT: epithelial-to-mesenchymal transition; ER: endoplasmic reticulum; FDA: Food and Drug Administration; FUT-I: alpha1,2-fucosyltransferase 1; FUT-VI: alpha1,3-fucosyltransferase VI; GA: Golgi apparatus; GalNAc: N-acetylgalactosamine; GlcNAc: N-acetylglucosamine; GPAA1: GPI anchor attachment 1; HAS: hyaluronic acid synthases; IGF-IR: insulin-like growth factor receptor I; ITGA6: alpha6beta4 integrin; ITGAV: αv integrin receptors; MIBC: muscle invasive bladder cancer; MUC1: mucin 1; MVAC: methotrexate, vinblastine, cisplatin and doxorubicin; NMIBC: non-muscle invasive bladder cancer; PD-L1: programmed death-ligand 1; PGAP1: post-GPI attachment to proteins 1; ppGalNAc-Ts: UDP-GalNAc:polypeptide N-acetylgalactosaminyl transferases; NMP22: nuclear matrix protein; OSTase: oligosaccharide transferase complex; RHAMM: hyaluronan-mediated motility; RhoGDI2: RhoGTP dissociation inhibitor 2; SLea: sialyl lewisa; SLex: sialyl lewisx; STn: sialyl-Tn; TGF: transforming growth factor; TURBT: transurethral resection of the bladder tumour.
Author contributions
LL, LLS and JAF presented the idea of making this review. RA, AP, CG, EF, MN, LL and JAF structured the manuscript. RA, AP, CG, EF and MN searched the state of art. RA and AP produced the first draft of the review. All authors contributed with alterations on the manuscript. All alterations were approved by LL, LLS and JAF.
Acknowledgments
The authors which to acknowledge the Portuguese Foundation for Science and Technology (FCT) for the PhD grants SFRH/BD/105355/2014 (Rita Azevedo), SFRH/BD/111242/2015 (Andreia Peixoto), SFRH/BD/127327/2016 (Cristiana Gaiteiro), SFRH/BD/103571/2014 (Elisabete Fernandes) and Postdoctoral grants SFRH/BPD/101827/2014 (Luis Lima) and SFRH/BPD/111048/2015 (José Alexandre Ferreira). FCT is co-financed by European Social Fund (ESF) under Human Potential Operation Programme (POPH) from National Strategic Reference Framework (NSRF). We also acknowledge the research grants from Research Center of Portuguese Oncology Institute of Porto (CI-IPOP 129 and 58-2016).
Conflicts of interest
The authors declare no potential conflict of interests.
References
1. Witjes JA, Comperat E, Cowan NC, De Santis M, Gakis G, Lebret T, Ribal MJ, Van der Heijden AG, Sherif A, European Association of U. EAU guidelines on muscle-invasive and metastatic bladder cancer: summary of the 2013 guidelines. Eur Urol. 2014; 65: 778-92. doi: 10.1016/j.eururo.2013.11.046.
2. Netto GJ. Molecular biomarkers in urothelial carcinoma of the bladder: are we there yet? Nat Rev Urol. 2012; 9: 41-51. doi: 10.1038/nrurol.2011.193.
3. Chen C, Qi XJ, Cao YW, Wang YH, Yang XC, Shao SX, Niu HT. Bladder Tumor Heterogeneity: The Impact on Clinical Treatment. Urol Int. 2015; 95: 1-8. doi: 10.1159/000370165.
4. Azevedo R, Ferreira JA, Peixoto A, Neves M, Sousa N, Lima A, Santos LL. Emerging antibody-based therapeutic strategies for bladder cancer: A systematic review. J Control Release. 2015; 214: 40-61. doi: 10.1016/j.jconrel.2015.07.002.
5. Sanli O, Dobruch J, Knowles MA, Burger M, Alemozaffar M, Nielsen ME, Lotan Y. Bladder cancer. Nat Rev Dis Primers. 2017; 3: 17022. doi: 10.1038/nrdp.2017.22.
6. Stowell SR, Ju T, Cummings RD. Protein glycosylation in cancer. Annu Rev Pathol. 2015; 10: 473-510. doi: 10.1146/annurev-pathol-012414-040438.
7. Anderson NL, Anderson NG. The human plasma proteome: history, character, and diagnostic prospects. Mol Cell Proteomics. 2002; 1: 845-67.
8. Mechref Y, Hu Y, Garcia A, Hussein A. Identifying cancer biomarkers by mass spectrometry-based glycomics. Electrophoresis. 2012; 33: 1755-67. doi: 10.1002/elps.201100715.
9. Ghazarian H, Idoni B, Oppenheimer SB. A glycobiology review: carbohydrates, lectins and implications in cancer therapeutics. Acta Histochem. 2011; 113: 236-47. doi: 10.1016/j.acthis.2010.02.004.
10. Spiro RG. Protein glycosylation: nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology. 2002; 12: 43R-56R.
11. Varki A. Biological roles of glycans. Glycobiology. 2017; 27: 3-49. doi: 10.1093/glycob/cww086.
12. Hakomori S. Tumor Malignancy Defined by Aberrant Glycosylation and Sphingo(glyco)lipid Metabolism. Cancer Research. 1996; 56: 5309-18.
13. Gruszewska E, Chrostek L. The alterations of glycosylation in malignant diseases. [Article in Polish]. Pol Merkur Lekarski. 2013; 34: 58-61.
14. Guda K, Moinova H, He J, Jamison O, Ravi L, Natale L, Lutterbaugh J, Lawrence E, Lewis S, Willson JK, Lowe JB, Wiesner GL, Parmigiani G, et al. Inactivating germ-line and somatic mutations in polypeptide N-acetylgalactosaminyltransferase 12 in human colon cancers. Proc Natl Acad Sci U S A. 2009; 106: 12921-5. doi: 10.1073/pnas.0901454106.
15. Vazquez-Martin C, Cuevas E, Gil-Martin E, Fernandez-Briera A. Correlation analysis between tumor-associated antigen sialyl-Tn expression and ST6GalNAc I activity in human colon adenocarcinoma. Oncology. 2004; 67: 159-65. doi: 10.1159/000081003.
16. Ju T, Lanneau GS, Gautam T, Wang Y, Xia B, Stowell SR, Willard MT, Wang W, Xia JY, Zuna RE, Laszik Z, Benbrook DM, Hanigan MH, et al. Human tumor antigens Tn and sialyl Tn arise from mutations in Cosmc. Cancer Res. 2008; 68: 1636-46. doi: 10.1158/0008-5472.CAN-07-2345.
17. Gill DJ, Tham KM, Chia J, Wang SC, Steentoft C, Clausen H, Bard-Chapeau EA, Bard FA. Initiation of GalNAc-type O-glycosylation in the endoplasmic reticulum promotes cancer cell invasiveness. Proc Natl Acad Sci U S A. 2013; 110: E3152-61. doi: 10.1073/pnas.1305269110.
18. Rivinoja A, Hassinen A, Kokkonen N, Kauppila A, Kellokumpu S. Elevated Golgi pH impairs terminal N-glycosylation by inducing mislocalization of Golgi glycosyltransferases. J Cell Physiol. 2009; 220: 144-54. doi: 10.1002/jcp.21744.
19. Pinho SS, Reis CA. Glycosylation in cancer: mechanisms and clinical implications. Nat Rev Cancer. 2015; 15: 540-55. doi: 10.1038/nrc3982.
20. Costa C, Pereira S, Lima L, Peixoto A, Fernandes E, Neves D, Neves M, Gaiteiro C, Tavares A, Gil da Costa RM, Cruz R, Amaro T, Oliveira PA, et al. Abnormal Protein Glycosylation and Activated PI3K/Akt/mTOR Pathway: Role in Bladder Cancer Prognosis and Targeted Therapeutics. PLoS One. 2015; 10: e0141253. doi: 10.1371/journal.pone.0141253.
21. Carrascal MA, Severino PF, Guadalupe Cabral M, Silva M, Ferreira JA, Calais F, Quinto H, Pen C, Ligeiro D, Santos LL, Dall’Olio F, Videira PA. Sialyl Tn-expressing bladder cancer cells induce a tolerogenic phenotype in innate and adaptive immune cells. Mol Oncol. 2014; 8: 753-65. doi: 10.1016/j.molonc.2014.02.008.
22. Hakomori S. Glycosylation defining cancer malignancy: new wine in an old bottle. Proc Natl Acad Sci U S A. 2002; 99: 10231-3. doi: 10.1073/pnas.172380699.
23. Guo J, Li X, Tan Z, Lu W, Yang G, Guan F. Alteration of N-glycans and expression of their related glycogenes in the epithelial-mesenchymal transition of HCV29 bladder epithelial cells. Molecules. 2014; 19: 20073-90. doi: 10.3390/molecules191220073.
24. Cazet A, Julien S, Bobowski M, Burchell J, Delannoy P. Tumour-associated carbohydrate antigens in breast cancer. Breast Cancer Res. 2010; 12: 204. doi: 10.1186/bcr2577.
25. Hauselmann I, Borsig L. Altered tumor-cell glycosylation promotes metastasis. Front Oncol. 2014; 4: 28. doi: 10.3389/fonc.2014.00028.
26. Helenius A, Aebi M. Intracellular functions of N-linked glycans. Science. 2001; 291: 2364-9.
27. Zhao YY, Takahashi M, Gu JG, Miyoshi E, Matsumoto A, Kitazume S, Taniguchi N. Functional roles of N-glycans in cell signaling and cell adhesion in cancer. Cancer Sci. 2008; 99: 1304-10. doi: 10.1111/j.1349-7006.2008.00839.x.
28. Zhao Y, Nakagawa T, Itoh S, Inamori K, Isaji T, Kariya Y, Kondo A, Miyoshi E, Miyazaki K, Kawasaki N, Taniguchi N, Gu J. N-acetylglucosaminyltransferase III antagonizes the effect of N-acetylglucosaminyltransferase V on alpha3beta1 integrin-mediated cell migration. J Biol Chem. 2006; 281: 32122-30. doi: 10.1074/jbc.M607274200.
29. Muthana SM, Campbell CT, Gildersleeve JC. Modifications of glycans: biological significance and therapeutic opportunities. ACS Chem Biol. 2012; 7: 31-43. doi: 10.1021/cb2004466.
30. Guo JM, Zhang XY, Chen HL, Wang GM, Zhang YK. Structural alterations of sugar chains in urine fibronectin from bladder cancer patients and its enzymatic mechanism. J Cancer Res Clin Oncol. 2001; 127: 512-9.
31. Takahashi T, Hagisawa S, Yoshikawa K, Tezuka F, Kaku M, Ohyama C. Predictive value of N-acetylglucosaminyltransferase-V for superficial bladder cancer recurrence. J Urol. 2006; 175: 90-3; discussion 3. doi: 10.1016/S0022-5347(05)00044-3.
32. Ishimura H, Takahashi T, Nakagawa H, Nishimura S, Arai Y, Horikawa Y, Habuchi T, Miyoshi E, Kyan A, Hagisawa S, Ohyama C. N-acetylglucosaminyltransferase V and beta1-6 branching N-linked oligosaccharides are associated with good prognosis of patients with bladder cancer. Clin Cancer Res. 2006; 12: 2506-11. doi: 10.1158/1078-0432.CCR-05-1938.
33. Taniguchi N, Miyoshi E, Ko JH, Ikeda Y, Ihara Y. Implication of N-acetylglucosaminyltransferases III and V in cancer: gene regulation and signaling mechanism. Biochim Biophys Acta. 1999; 1455: 287-300.
34. Granovsky M, Fata J, Pawling J, Muller WJ, Khokha R, Dennis JW. Suppression of tumor growth and metastasis in Mgat5-deficient mice. Nat Med. 2000; 6: 306-12. doi: 10.1038/73163.
35. Huang B, Wu Q, Ge Y, Zhang J, Sun L, Zhang Y, Fu L, Fan J, Wang Z. Expression of N-acetylglucosaminyltransferase V in gastric cancer correlates with metastasis and prognosis. Int J Oncol. 2014; 44: 849-57. doi: 10.3892/ijo.2014.2248.
36. Huang B, Sun L, Cao J, Zhang Y, Wu Q, Zhang J, Ge Y, Fu L, Wang Z. Downregulation of the GnT-V gene inhibits metastasis and invasion of BGC823 gastric cancer cells. Oncol Rep. 2013; 29: 2392-400. doi: 10.3892/or.2013.2373.
37. Seto K, Uchida F, Baba O, Yamatoji M, Karube R, Warabi E, Sakai S, Hasegawa S, Yamagata K, Yanagawa T, Onizawa K, Miyoshi E, Shoda J, et al. Negative expression of N-acetylglucosaminyltransferase V in oral squamous cell carcinoma correlates with poor prognosis. Springerplus. 2013; 2: 657. doi: 10.1186/2193-1801-2-657.
38. Yamamoto E, Ino K, Miyoshi E, Shibata K, Takahashi N, Kajiyama H, Nawa A, Nomura S, Nagasaka T, Kikkawa F. Expression of N-acetylglucosaminyltransferase V in endometrial cancer correlates with poor prognosis. Br J Cancer. 2007; 97: 1538-44. doi: 10.1038/sj.bjc.6604044.
39. Chihara Y, Sugano K, Kobayashi A, Kanai Y, Yamamoto H, Nakazono M, Fujimoto H, Kakizoe T, Fujimoto K, Hirohashi S, Hirao Y. Loss of blood group A antigen expression in bladder cancer caused by allelic loss and/or methylation of the ABO gene. Lab Invest. 2005; 85: 895-907. doi: 10.1038/labinvest.3700268.
40. Sheinfeld J, Reuter VE, Fair WR, Cordon-Cardo C. Expression of blood group antigens in bladder cancer: current concepts. Semin Surg Oncol. 1992; 8: 308-15. doi: 10.1002/ssu.2980080510.
41. Stanley P, Cummings RD. (2009). Structures Common to Different Glycans. In: Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P, Bertozzi CR, Hart GW and Etzler ME, eds. Essentials of Glycobiology. (Cold Spring Harbor (NY).
42. Bergman S, Javadpour N. The cell surface antigen A, B or O(H) as an indicator of malignant potential in stage A bladder carcinoma: preliminary report. J Urol. 1978; 119: 49-51.
43. Hakomori S. (2001). Tumour-associated carbohydrate antigens defining tumor malignancy: Basis for development of anti-cancer vaccines. In: Albert M and Wu KAPP, eds., pp. 369-402.
44. Summers JL, Coon JS, Ward RM, Falor WH, Miller AW 3rd, Weinstein RS. Lewis antigens in normal and neoplastic urothelium. Am J Pathol. 1985; 121: 176-83.
45. Summers JL, Coon JS, Ward RM, Falor WH, Miller AW 3rd, Weinstein RS. Prognosis in carcinoma of the urinary bladder based upon tissue blood group ABH and Thomsen-Friedenreich antigen status and karyotype of the initial tumor. Cancer Res. 1983; 43: 934-9.
46. Cordon-cardo C, Reuter VE, Lloyd KO, Determinants L, Sheinfeld J, Fair WR, Old LJ, Melamed MR. Blood Group-related Antigens in Human Urothelium : Enhanced Expression of Precursor , Le X , and Le Y Determinants in Urothelial Carcinoma. Cancer Res. 1988; 48: 4113-20.
47. Thorpe SJ, Abel P, Slavin G, Feizi T. Blood group antigens in the normal and neoplastic bladder epithelium. J Clin Pathol. 1983; 36: 873-82.
48. Orntoft TF, Wolf H, Watkins WM. Activity of the human blood group ABO, Se, H, Le, and X gene-encoded glycosyltransferases in normal and malignant bladder urothelium. Cancer Res. 1988; 48: 4427-33.
49. Juhl BR, Hartzen SH, Hainau B. Lewis a antigen in transitional cell tumors of the urinary bladder. Cancer. 1986; 58: 222-8.
50. Kajiwara H, Yasuda M, Kumaki N, Shibayama T, Osamura Y. Expression of carbohydrate antigens (SSEA-1, sialyl-Lewis X, DU-PAN-2 and CA19-9) and E-selectin in urothelial carcinoma of the renal pelvis, ureter, and urinary bladder. Tokai J Exp Clin Med. 2005; 30: 177-82.
51. Ferreira JA, Magalhaes A, Gomes J, Peixoto A, Gaiteiro C, Fernandes E, Santos LL, Reis CA. Protein glycosylation in gastric and colorectal cancers: Toward cancer detection and targeted therapeutics. Cancer Lett. 2017; 387: 32-45. doi: 10.1016/j.canlet.2016.01.044.
52. St Hill CA. Interactions between endothelial selectins and cancer cells regulate metastasis. Front Biosci (Landmark Ed). 2011; 16: 3233-51.
53. Hegele A, Mecklenburg V, Varga Z, Olbert P, Hofmann R, Barth P. CA19.9 and CEA in transitional cell carcinoma of the bladder: serological and immunohistochemical findings. Anticancer Res. 2010; 30: 5195-200.
54. Numahata K, Satoh M, Handa K, Saito S, Ohyama C, Ito A, Takahashi T, Hoshi S, Orikasa S, Hakomori SI. Sialosyl-Le(x) expression defines invasive and metastatic properties of bladder carcinoma. Cancer. 2002; 94: 673-85. doi: 10.1002/cncr.10268.
55. Miyazaki K, Ohmori K, Izawa M, Koike T, Kumamoto K, Furukawa K, Ando T, Kiso M, Yamaji T, Hashimoto Y, Suzuki A, Yoshida A, Takeuchi M, et al. Loss of disialyl Lewis(a), the ligand for lymphocyte inhibitory receptor sialic acid-binding immunoglobulin-like lectin-7 (Siglec-7) associated with increased sialyl Lewis(a) expression on human colon cancers. Cancer Res. 2004; 64: 4498-505. doi: 10.1158/0008-5472.CAN-03-3614.
56. Almaraz RT, Tian Y, Bhattarcharya R, Tan E, Chen SH, Dallas MR, Chen L, Zhang Z, Zhang H, Konstantopoulos K, Yarema KJ. Metabolic flux increases glycoprotein sialylation: implications for cell adhesion and cancer metastasis. Mol Cell Proteomics. 2012; 11: M112 017558. doi: 10.1074/mcp.M112.017558.
57. Miyoshi E, Moriwaki K, Nakagawa T. Biological function of fucosylation in cancer biology. J Biochem. 2008; 143: 725-9. doi: 10.1093/jb/mvn011.
58. Yang G, Tan Z, Lu W, Guo J, Yu H, Yu J, Sun C, Qi X, Li Z, Guan F. Quantitative glycome analysis of N-glycan patterns in bladder cancer vs normal bladder cells using an integrated strategy. J Proteome Res. 2015; 14: 639-53. doi: 10.1021/pr5006026.
59. Lu YC, Chen CN, Chu CY, Lu J, Wang BJ, Chen CH, Huang MC, Lin TH, Pan CC, Chen SS, Hsu WM, Liao YF, Wu PY, et al. Calreticulin activates beta1 integrin via fucosylation by fucosyltransferase 1 in J82 human bladder cancer cells. Biochem J. 2014; 460: 69-78. doi: 10.1042/BJ20131424.
60. Brockhausen I, Schachter H, Stanley P. (2009). O-GalNAc Glycans. In: Varki A, Cummings RD, Esko JD, Freeze HH, Stanley P, Bertozzi CR, Hart GW and Etzler ME, eds. Essentials of Glycobiology. (Cold Spring Harbor (NY).
61. Steentoft C, Vakhrushev SY, Joshi HJ, Kong Y, Vester-Christensen MB, Schjoldager KT, Lavrsen K, Dabelsteen S, Pedersen NB, Marcos-Silva L, Gupta R, Bennett EP, Mandel U, et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO J. 2013; 32: 1478-88. doi: 10.1038/emboj.2013.79.
62. Thanka Christlet TH, Veluraja K. Database analysis of O-glycosylation sites in proteins. Biophys J. 2001; 80: 952-60.
63. Kufe DW. Mucins in cancer: function, prognosis and therapy. Nat Rev Cancer. 2009; 9: 874-85. doi: 10.1038/nrc2761.
64. Ferreira JA, Videira PA, Lima L, Pereira S, Silva M, Carrascal M, Severino PF, Fernandes E, Almeida A, Costa C, Vitorino R, Amaro T, Oliveira MJ, et al. Overexpression of tumour-associated carbohydrate antigen sialyl-Tn in advanced bladder tumours. Mol Oncol. 2013; 7: 719-31. doi: 10.1016/j.molonc.2013.03.001.
65. Videira PA, Correia M, Malagolini N, Crespo HJ, Ligeiro D, Calais FM, Trindade H, Dall’Olio F. ST3Gal.I sialyltransferase relevance in bladder cancer tissues and cell lines. BMC Cancer. 2009; 9: 357. doi: 10.1186/1471-2407-9-357.
66. Yokoyama M, Ohoka H, Oda H, Oda T, Utsumi S, Takeuchi M. Thomsen-Friedenreich antigen in bladder cancer tissues detected by monoclonal antibody. Hinyokika Kiyo. 1988; 34: 255-8.
67. Langkilde NC, Wolf H, Clausen H, Kjeldsen T, Orntoft TF. Nuclear volume and expression of T-antigen, sialosyl-Tn-antigen, and Tn-antigen in carcinoma of the human bladder. Relation to tumor recurrence and progression. Cancer. 1992; 69: 219-27.
68. Tran DT, Ten Hagen KG. Mucin-type O-glycosylation during development. J Biol Chem. 2013; 288: 6921-9. doi: 10.1074/jbc.R112.418558.
69. Peixoto A, Fernandes E, Gaiteiro C, Lima L, Azevedo R, Soares J, Cotton S, Parreira B, Neves M, Amaro T, Tavares A, Teixeira F, Palmeira C, et al. Hypoxia enhances the malignant nature of bladder cancer cells and concomitantly antagonizes protein O-glycosylation extension. Oncotarget. 2016; 7:63138-63157. doi: 10.18632/oncotarget.11257.
70. Clement M, Rocher J, Loirand G, Le Pendu J. Expression of sialyl-Tn epitopes on beta1 integrin alters epithelial cell phenotype, proliferation and haptotaxis. J Cell Sci. 2004; 117: 5059-69. doi: 10.1242/jcs.01350.
71. Yoo NJ, Kim MS, Lee SH. Absence of COSMC gene mutations in breast and colorectal carcinomas. APMIS. 2008; 116: 154-5. doi: 10.1111/j.1600-0463.2008.00965.x.
72. Reis CA, Osório H, Silva L, Gomes C, David L. Alterations in glycosylation as biomarkers for cancer detection. J Clin Pathol. 2010; 63: 322-9.
73. Santos SN, Junqueira MS, Francisco G, Vilanova M, Magalhaes A, Baruffi MD, Chammas R, Harris AL, Reis CA, Bernardes ES. O-glycan sialylation alters galectin-3 subcellular localization and decreases chemotherapy sensitivity in gastric cancer. Oncotarget. 2016; 7:83570-83587. doi: 10.18632/oncotarget.13192.
74. Suzuki Y, Sutoh M, Hatakeyama S, Mori K, Yamamoto H, Koie T, Saitoh H, Yamaya K, Funyu T, Habuchi T, Arai Y, Fukuda M, Ohyama C, et al. MUC1 carrying core 2 O-glycans functions as a molecular shield against NK cell attack, promoting bladder tumor metastasis. Int J Oncol. 2012; 40: 1831-8. doi: 10.3892/ijo.2012.1411.
75. Lima L, Severino PF, Silva M, Miranda A, Tavares A, Pereira S, Fernandes E, Cruz R, Amaro T, Reis CA, Dall’Olio F, Amado F, Videira PA, et al. Response of high-risk of recurrence/progression bladder tumours expressing sialyl-Tn and sialyl-6-T to BCG immunotherapy. Br J Cancer. 2013; 109: 2106-14. doi: 10.1038/bjc.2013.571.
76. Limas C, Lange P. T-antigen in normal and neoplastic urothelium. Cancer. 1986; 58: 1236-45.
77. Cao Y, Stosiek P, Springer GF, Karsten U. Thomsen-Friedenreich-related carbohydrate antigens in normal adult human tissues: a systematic and comparative study. Histochem Cell Biol. 1996; 106: 197-207. doi: 10.1007/BF02484401.
78. Contessa JN, Bhojani MS, Freeze HH, Rehemtulla A, Lawrence TS. Inhibition of N-linked glycosylation disrupts receptor tyrosine kinase signaling in tumor cells. Cancer Res. 2008; 68: 3803-9. doi: 10.1158/0008-5472.CAN-07-6389.
79. Iqbal N, Iqbal N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol Biol Int. 2014; 2014: 852748. doi: 10.1155/2014/852748.
80. Kruger S, Weitsch G, Buttner H, Matthiensen A, Bohmer T, Marquardt T, Sayk F, Feller AC, Bohle A. HER2 overexpression in muscle-invasive urothelial carcinoma of the bladder: prognostic implications. Int J Cancer. 2002; 102: 514-8. doi: 10.1002/ijc.10731.
81. Yan M, Schwaederle M, Arguello D, Millis SZ, Gatalica Z, Kurzrock R. HER2 expression status in diverse cancers: review of results from 37,992 patients. Cancer Metastasis Rev. 2015; 34: 157-64. doi: 10.1007/s10555-015-9552-6.
82. Zhao J, Xu W, Zhang Z, Song R, Zeng S, Sun Y, Xu C. Prognostic role of HER2 expression in bladder cancer: a systematic review and meta-analysis. Int Urol Nephrol. 2015; 47: 87-94. doi: 10.1007/s11255-014-0866-z.
83. Balzar M, Winter MJ, de Boer CJ, Litvinov SV. The biology of the 17-1A antigen (Ep-CAM). J Mol Med (Berl). 1999; 77: 699-712.
84. Munz M, Baeuerle PA, Gires O. The emerging role of EpCAM in cancer and stem cell signaling. Cancer Res. 2009; 69: 5627-9. doi: 10.1158/0008-5472.CAN-09-0654.
85. Dolle L, Theise ND, Schmelzer E, Boulter L, Gires O, van Grunsven LA. EpCAM and the biology of hepatic stem/progenitor cells. Am J Physiol Gastrointest Liver Physiol. 2015; 308: G233-50. doi: 10.1152/ajpgi.00069.2014.
86. Bryan RT, Shimwell NJ, Wei W, Devall AJ, Pirrie SJ, James ND, Zeegers MP, Cheng KK, Martin A, Ward DG. Urinary EpCAM in urothelial bladder cancer patients: characterisation and evaluation of biomarker potential. Br J Cancer. 2014; 110: 679-85. doi: 10.1038/bjc.2013.744.
87. Brunner A, Prelog M, Verdorfer I, Tzankov A, Mikuz G, Ensinger C. EpCAM is predominantly expressed in high grade and advanced stage urothelial carcinoma of the bladder. J Clin Pathol. 2008; 61: 307-10. doi: 10.1136/jcp.2007.049460.
88. Rabinovich GA, Toscano MA, Jackson SS, Vasta GR. Functions of cell surface galectin-glycoprotein lattices. Curr Opin Struct Biol. 2007; 17: 513-20. doi: 10.1016/j.sbi.2007.09.002.
89. Rabinovich GA, Rubinstein N, Fainboim L. Unlocking the secrets of galectins: a challenge at the frontier of glyco-immunology. J Leukoc Biol. 2002; 71: 741-52.
90. Fortuna-Costa A, Gomes AM, Kozlowski EO, Stelling MP, Pavao MS. Extracellular galectin-3 in tumor progression and metastasis. Front Oncol. 2014; 4: 138. doi: 10.3389/fonc.2014.00138.
91. Funasaka T, Raz A, Nangia-Makker P. Galectin-3 in angiogenesis and metastasis. Glycobiology. 2014; 24: 886-91. doi: 10.1093/glycob/cwu086.
92. Boscher C, Dennis JW, Nabi IR. Glycosylation, galectins and cellular signaling. Curr Opin Cell Biol. 2011; 23: 383-92. doi: 10.1016/j.ceb.2011.05.001.
93. Liu FT, Rabinovich GA. Galectins as modulators of tumour progression. Nat Rev Cancer. 2005; 5: 29-41.
94. Langbein S, Brade J, Badawi JK, Hatzinger M, Kaltner H, Lensch M, Specht K, Andre S, Brinck U, Alken P, Gabius HJ. Gene-expression signature of adhesion/growth-regulatory tissue lectins (galectins) in transitional cell cancer and its prognostic relevance. Histopathology. 2007; 51: 681-90. doi: 10.1111/j.1365-2559.2007.02852.x.
95. Shen KH, Li CF, Chien LH, Huang CH, Su CC, Liao AC, Wu TF. Role of galectin-1 in urinary bladder urothelial carcinoma cell invasion through the JNK pathway. Cancer Sci. 2016; 107: 1390-8. doi: 10.1111/cas.13016.
96. Wu TF, Li CF, Chien LH, Shen KH, Huang HY, Su CC, Liao AC. Galectin-1 dysregulation independently predicts disease specific survival in bladder urothelial carcinoma. J Urol. 2015; 193: 1002-8. doi: 10.1016/j.juro.2014.09.107.
97. Cindolo L, Benvenuto G, Salvatore P, Pero R, Salvatore G, Mirone V, Prezioso D, Altieri V, Bruni CB, Chiariotti L. galectin-1 and galectin-3 expression in human bladder transitional-cell carcinomas. Int J Cancer. 1999; 84: 39-43.
98. Balan V, Nangia-Makker P, Raz A. Galectins as Cancer Biomarkers. Cancers. 2010; 2: 592-610.
99. Pereira PM, Silva S, Ramalho JS, Gomes CM, Girao H, Cavaleiro JA, Ribeiro CA, Tome JP, Fernandes R. The role of galectin-1 in in vitro and in vivo photodynamic therapy with a galactodendritic porphyrin. Eur J Cancer. 2016; 68: 60-9. doi: 10.1016/j.ejca.2016.08.018.
100. Sakaki M, Oka N, Nakanishi R, Yamaguchi K, Fukumori T, Kanayama HO. Serum level of galectin-3 in human bladder cancer. J Med Invest. 2008; 55: 127-32.
101. Canesin G, Gonzalez-Peramato P, Palou J, Urrutia M, Cordon-Cardo C, Sanchez-Carbayo M. Galectin-3 expression is associated with bladder cancer progression and clinical outcome. Tumour Biol. 2010; 31: 277-85. doi: 10.1007/s13277-010-0033-9.
102. Gendy HE, Madkour B, Abdelaty S, Essawy F, Khattab D, Hammam O, Nour HH. Diagnostic and Prognostic Significance of Serum and Tissue Galectin 3 Expression in Patients with Carcinoma of the Bladder. Curr Urol. 2014; 7: 185-90. doi: 10.1159/000365673.
103. El Gendy H, Madkour B, Abdelaty S, Essawy F, Khattab D, Hammam O, El Kholy A, Nour HH. Galectin 3 for the diagnosis of bladder cancer. Arab J Urol. 2014; 12: 178-81. doi: 10.1016/j.aju.2013.10.004.
104. Matsui Y, Ueda S, Watanabe J, Kuwabara I, Ogawa O, Nishiyama H. Sensitizing effect of galectin-7 in urothelial cancer to cisplatin through the accumulation of intracellular reactive oxygen species. Cancer Res. 2007; 67: 1212-20. doi: 10.1158/0008-5472.CAN-06-3283.
105. Kramer MW, Waalkes S, Serth J, Hennenlotter J, Tezval H, Stenzl A, Kuczyk MA, Merseburger AS. Decreased galectin-8 is a strong marker for recurrence in urothelial carcinoma of the bladder. Urol Int. 2011; 87: 143-50. doi: 10.1159/000328439.
106. Ohishi T, Koga F, Migita T. Bladder Cancer Stem-Like Cells: Their Origin and Therapeutic Perspectives. Int J Mol Sci. 2015; 17. doi: 10.3390/ijms17010043.
107. Naor D, Nedvetzki S, Golan I, Melnik L, Faitelson Y. CD44 in cancer. Crit Rev Clin Lab Sci. 2002; 39: 527-79. doi: 10.1080/10408360290795574.
108. Ponta H, Sherman L, Herrlich PA. CD44: from adhesion molecules to signalling regulators. Nat Rev Mol Cell Biol. 2003; 4: 33-45. doi: 10.1038/nrm1004.
109. Sneath RJ, Mangham DC. The normal structure and function of CD44 and its role in neoplasia. Mol Pathol. 1998; 51: 191-200.
110. Prince ME, Sivanandan R, Kaczorowski A, Wolf GT, Kaplan MJ, Dalerba P, Weissman IL, Clarke MF, Ailles LE. Identification of a subpopulation of cells with cancer stem cell properties in head and neck squamous cell carcinoma. Proc Natl Acad Sci U S A. 2007; 104: 973-8. doi: 10.1073/pnas.0610117104.
111. Takaishi S, Okumura T, Tu S, Wang SS, Shibata W, Vigneshwaran R, Gordon SA, Shimada Y, Wang TC. Identification of gastric cancer stem cells using the cell surface marker CD44. Stem Cells. 2009; 27: 1006-20. doi: 10.1002/stem.30.
112. Collins AT, Berry PA, Hyde C, Stower MJ, Maitland NJ. Prospective identification of tumorigenic prostate cancer stem cells. Cancer Res. 2005; 65: 10946-51. doi: 10.1158/0008-5472.CAN-05-2018.
113. Dalerba P, Dylla SJ, Park IK, Liu R, Wang X, Cho RW, Hoey T, Gurney A, Huang EH, Simeone DM, Shelton AA, Parmiani G, Castelli C, et al. Phenotypic characterization of human colorectal cancer stem cells. Proc Natl Acad Sci U S A. 2007; 104: 10158-63. doi: 10.1073/pnas.0703478104.
114. Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, Wicha M, Clarke MF, Simeone DM. Identification of pancreatic cancer stem cells. Cancer Res. 2007; 67: 1030-7. doi: 10.1158/0008-5472.CAN-06-2030.
115. Chan KS, Espinosa I, Chao M, Wong D, Ailles L, Diehn M, Gill H, Presti J Jr, Chang HY, van de Rijn M, Shortliffe L, Weissman IL. Identification, molecular characterization, clinical prognosis, and therapeutic targeting of human bladder tumor-initiating cells. Proc Natl Acad Sci U S A. 2009; 106: 14016-21. doi: 10.1073/pnas.0906549106.
116. Tatokoro M, Koga F, Yoshida S, Kawakami S, Fujii Y, Neckers L, Kihara K. Potential role of Hsp90 inhibitors in overcoming cisplatin resistance of bladder cancer-initiating cells. Int J Cancer. 2012; 131: 987-96. doi: 10.1002/ijc.26475.
117. Yang YM, Chang JW. Bladder cancer initiating cells (BCICs) are among EMA-CD44v6+ subset: novel methods for isolating undetermined cancer stem (initiating) cells. Cancer Invest. 2008; 26: 725-33. doi: 10.1080/07357900801941845.
118. Thanan R, Murata M, Ma N, Hammam O, Wishahi M, El Leithy T, Hiraku Y, Oikawa S, Kawanishi S. Nuclear localization of COX-2 in relation to the expression of stemness markers in urinary bladder cancer. Mediators Inflamm. 2012; 2012: 165879. doi: 10.1155/2012/165879.
119. Kuncova J, Kostrouch Z, Viale M, Revoltella R, Mandys V. Expression of CD44v6 correlates with cell proliferation and cellular atypia in urothelial carcinoma cell lines 5637 and HT1197. Folia Biol (Praha). 2005; 51: 3-11.
120. Omran OM, Ata HS. CD44s and CD44v6 in diagnosis and prognosis of human bladder cancer. Ultrastruct Pathol. 2012; 36: 145-52. doi: 10.3109/01913123.2011.651522.
121. Lipponen P, Aaltoma S, Kosma VM, Ala-Opas M, Eskelinen M. Expression of CD44 standard and variant-v6 proteins in transitional cell bladder tumours and their relation to prognosis during a long-term follow-up. J Pathol. 1998; 186: 157-64.
122. Klatte T, Seligson DB, Rao JY, Yu H, de Martino M, Garraway I, Wong SG, Belldegrun AS, Pantuck AJ. Absent CD44v6 expression is an independent predictor of poor urothelial bladder cancer outcome. J Urol. 2010; 183: 2403-8. doi: 10.1016/j.juro.2010.01.064.
123. Pauli C, Munz M, Kieu C, Mack B, Breinl P, Wollenberg B, Lang S, Zeidler R, Gires O. Tumor-specific glycosylation of the carcinoma-associated epithelial cell adhesion molecule EpCAM in head and neck carcinomas. Cancer Lett. 2003; 193: 25-32.
124. Stavropoulos NE, Filliadis I, Ioachim E, Michael M, Mermiga E, Hastazeris K, Nseyo UO. CD44 standard form expression as a predictor of progression in high risk superficial bladder tumors. Int Urol Nephrol. 2001; 33: 479-83.
125. Kobayashi K, Matsumoto H, Matsuyama H, Fujii N, Inoue R, Yamamoto Y, Nagao K. Clinical significance of CD44 variant 9 expression as a prognostic indicator in bladder cancer. Oncol Rep. 2016; 36: 2852-2860. doi: 10.3892/or.2016.5061.
126. Mehla K, Singh PK. MUC1: a novel metabolic master regulator. Biochim Biophys Acta. 2014; 1845: 126-35. doi: 10.1016/j.bbcan.2014.01.001.
127. Simms MS, Hughes OD, Limb M, Price MR, Bishop MC. MUC1 mucin as a tumour marker in bladder cancer. BJU Int. 1999; 84: 350-2.
128. Kaur S, Momi N, Chakraborty S, Wagner DG, Horn AJ, Lele SM, Theodorescu D, Batra SK. Altered expression of transmembrane mucins, MUC1 and MUC4, in bladder cancer: pathological implications in diagnosis. PLoS One. 2014; 9: e92742. doi: 10.1371/journal.pone.0092742.
129. Stojnev S, Ristic-Petrovic A, Velickovic LJ, Krstic M, Bogdanovic D, Khanh do T, Ristic A, Conic I, Stefanovic V. Prognostic significance of mucin expression in urothelial bladder cancer. Int J Clin Exp Pathol. 2014; 7: 4945-58.
130. Walsh MD, Hohn BG, Thong W, Devine PL, Gardiner RA, Samaratunga ML, McGuckin MA. Mucin expression by transitional cell carcinomas of the bladder. Br J Urol. 1994; 73: 256-62.
131. Nielsen TO, Borre M, Nexo E, Sorensen BS. Co-expression of HER3 and MUC1 is associated with a favourable prognosis in patients with bladder cancer. BJU Int. 2015; 115: 163-5. doi: 10.1111/bju.12658.
132. Cotton S, Azevedo R, Gaiteiro C, Ferreira D, Lima L, Peixoto A, Fernandes E, Neves M, Neves D, Amaro T, Cruz R, Tavares A, Rangel M, et al. Targeted O-glycoproteomics explored increased sialylation and identified MUC16 as a poor prognosis biomarker in advanced stage bladder tumours. Mol Oncol. 2017; 11:895-912. doi: 10.1002/1878-0261.12035.
133. Grossman HB, Lee C, Bromberg J, Liebert M. Expression of the alpha6beta4 integrin provides prognostic information in bladder cancer. Oncol Rep. 2000; 7: 13-6.
134. Elshamy WM, Duhe RJ. Overview: cellular plasticity, cancer stem cells and metastasis. Cancer Lett. 2013; 341: 2-8. doi: 10.1016/j.canlet.2013.06.020.
135. Chang JT, Mani SA. Sheep, wolf, or werewolf: cancer stem cells and the epithelial-to-mesenchymal transition. Cancer Lett. 2013; 341: 16-23. doi: 10.1016/j.canlet.2013.03.004.
136. van der Horst G, van den Hoogen C, Buijs JT, Cheung H, Bloys H, Pelger RC, Lorenzon G, Heckmann B, Feyen J, Pujuguet P, Blanque R, Clement-Lacroix P, van der Pluijm G. Targeting of alpha(v)-integrins in stem/progenitor cells and supportive microenvironment impairs bone metastasis in human prostate cancer. Neoplasia. 2011; 13: 516-25.
137. Chen Q, Manning CD, Millar H, McCabe FL, Ferrante C, Sharp C, Shahied-Arruda L, Doshi P, Nakada MT, Anderson GM. CNTO 95, a fully human anti alphav integrin antibody, inhibits cell signaling, migration, invasion, and spontaneous metastasis of human breast cancer cells. Clin Exp Metastasis. 2008; 25: 139-48. doi: 10.1007/s10585-007-9132-4.
138. Trikha M, Zhou Z, Nemeth JA, Chen Q, Sharp C, Emmell E, Giles-Komar J, Nakada MT. CNTO 95, a fully human monoclonal antibody that inhibits alphav integrins, has antitumor and antiangiogenic activity in vivo. Int J Cancer. 2004; 110: 326-35. doi: 10.1002/ijc.20116.
139. Sachs MD, Rauen KA, Ramamurthy M, Dodson JL, De Marzo AM, Putzi MJ, Schoenberg MP, Rodriguez R. Integrin alpha(v) and coxsackie adenovirus receptor expression in clinical bladder cancer. Urology. 2002; 60: 531-6.
140. van der Horst G, Bos L, van der Mark M, Cheung H, Heckmann B, Clement-Lacroix P, Lorenzon G, Pelger RC, Bevers RF, van der Pluijm G. Targeting of alpha-v integrins reduces malignancy of bladder carcinoma. PLoS One. 2014; 9: e108464. doi: 10.1371/journal.pone.0108464.
141. Heyder C, Gloria-Maercker E, Hatzmann W, Niggemann B, Zanker KS, Dittmar T. Role of the beta1-integrin subunit in the adhesion, extravasation and migration of T24 human bladder carcinoma cells. Clin Exp Metastasis. 2005; 22: 99-106. doi: 10.1007/s10585-005-4335-z.
142. Mierke CT, Frey B, Fellner M, Herrmann M, Fabry B. Integrin alpha5beta1 facilitates cancer cell invasion through enhanced contractile forces. J Cell Sci. 2011; 124: 369-83. doi: 10.1242/jcs.071985.
143. Theocharis AD, Skandalis SS, Tzanakakis GN, Karamanos NK. Proteoglycans in health and disease: novel roles for proteoglycans in malignancy and their pharmacological targeting. FEBS J. 2010; 277: 3904-23. doi: 10.1111/j.1742-4658.2010.07800.x.
144. Sarrazin S, Lamanna WC, Esko JD. Heparan sulfate proteoglycans. Cold Spring Harb Perspect Biol. 2011; 3. doi: 10.1101/cshperspect.a004952.
145. Korpetinou A, Skandalis SS, Labropoulou VT, Smirlaki G, Noulas A, Karamanos NK, Theocharis AD. Serglycin: at the crossroad of inflammation and malignancy. Front Oncol. 2014; 3: 327. doi: 10.3389/fonc.2013.00327.
146. Afratis N, Gialeli C, Nikitovic D, Tsegenidis T, Karousou E, Theocharis AD, Pavao MS, Tzanakakis GN, Karamanos NK. Glycosaminoglycans: key players in cancer cell biology and treatment. FEBS J. 2012; 279: 1177-97. doi: 10.1111/j.1742-4658.2012.08529.x.
147. Couchman JR, Pataki CA. An introduction to proteoglycans and their localization. J Histochem Cytochem. 2012; 60: 885-97. doi: 10.1369/0022155412464638.
148. Vigetti D, Viola M, Karousou E, De Luca G, Passi A. Metabolic control of hyaluronan synthases. Matrix Biol. 2014; 35: 8-13. doi: 10.1016/j.matbio.2013.10.002.
149. Iozzo RV. Matrix proteoglycans: from molecular design to cellular function. Annu Rev Biochem. 1998; 67: 609-52. doi: 10.1146/annurev.biochem.67.1.609.
150. Prydz K, Dalen KT. Synthesis and sorting of proteoglycans. J Cell Sci. 2000; 113 Pt 2: 193-205.
151. Bernfield M, Gotte M, Park PW, Reizes O, Fitzgerald ML, Lincecum J, Zako M. Functions of cell surface heparan sulfate proteoglycans. Annu Rev Biochem. 1999; 68: 729-77. doi: 10.1146/annurev.biochem.68.1.729.
152. Kishibe J, Yamada S, Okada Y, Sato J, Ito A, Miyazaki K, Sugahara K. Structural requirements of heparan sulfate for the binding to the tumor-derived adhesion factor/angiomodulin that induces cord-like structures to ECV-304 human carcinoma cells. J Biol Chem. 2000; 275: 15321-9.
153. Couchman JR. Transmembrane signaling proteoglycans. Annu Rev Cell Dev Biol. 2010; 26: 89-114. doi: 10.1146/annurev-cellbio-100109-104126.
154. Teng YH, Aquino RS, Park PW. Molecular functions of syndecan-1 in disease. Matrix Biol. 2012; 31: 3-16. doi: 10.1016/j.matbio.2011.10.001.
155. Xian X, Gopal S, Couchman JR. Syndecans as receptors and organizers of the extracellular matrix. Cell Tissue Res. 2010; 339: 31-46. doi: 10.1007/s00441-009-0829-3.
156. Sanaee MN, Malekzadeh M, Khezri A, Ghaderi A, Doroudchi M. Soluble CD138/Syndecan-1 Increases in the Sera of Patients with Moderately Differentiated Bladder Cancer. Urol Int. 2015; 94: 472-8. doi: 10.1159/000364907.
157. Mennerich D, Vogel A, Klaman I, Dahl E, Lichtner RB, Rosenthal A, Pohlenz HD, Thierauch KH, Sommer A. Shift of syndecan-1 expression from epithelial to stromal cells during progression of solid tumours. Eur J Cancer. 2004; 40: 1373-82. doi: 10.1016/j.ejca.2004.01.038.
158. Szarvas T, Reis H, Kramer G, Shariat SF, Vom Dorp F, Tschirdewahn S, Schmid KW, Kovalszky I, Rubben H. Enhanced stromal syndecan-1 expression is an independent risk factor for poor survival in bladder cancer. Hum Pathol. 2014; 45: 674-82. doi: 10.1016/j.humpath.2013.10.036.
159. Miyake M, Lawton A, Dai Y, Chang M, Mengual L, Alcaraz A, Goodison S, Rosser CJ. Clinical implications in the shift of syndecan-1 expression from the cell membrane to the cytoplasm in bladder cancer. BMC Cancer. 2014; 14: 86. doi: 10.1186/1471-2407-14-86.
160. Kim JH, Park J. Prognostic significance of heme oxygenase-1, S100 calcium-binding protein A4, and syndecan-1 expression in primary non-muscle-invasive bladder cancer. Hum Pathol. 2014; 45: 1830-8. doi: 10.1016/j.humpath.2014.04.020.
161. Shimada K, Nakamura M, De Velasco MA, Tanaka M, Ouji Y, Miyake M, Fujimoto K, Hirao K, Konishi N. Role of syndecan-1 (CD138) in cell survival of human urothelial carcinoma. Cancer Sci. 2010; 101: 155-60. doi: 10.1111/j.1349-7006.2009.01379.x.
162. Aaboe M, Marcussen N, Jensen KM, Thykjaer T, Dyrskjot L, Orntoft TF. Gene expression profiling of noninvasive primary urothelial tumours using microarrays. Br J Cancer. 2005; 93: 1182-90. doi: 10.1038/sj.bjc.6602813.
163. Mythreye K, Blobe GC. Proteoglycan signaling co-receptors: roles in cell adhesion, migration and invasion. Cell Signal. 2009; 21: 1548-58. doi: 10.1016/j.cellsig.2009.05.001.
164. Saban R, Saban MR, Maier J, Fowler B, Tengowski M, Davis CA, Wu XR, Culkin DJ, Hauser P, Backer J, Hurst RE. Urothelial expression of neuropilins and VEGF receptors in control and interstitial cystitis patients. Am J Physiol Renal Physiol. 2008; 295: F1613-23. doi: 10.1152/ajprenal.90344.2008.
165. Cheng W, Fu D, Wei ZF, Xu F, Xu XF, Liu YH, Ge JP, Tian F, Han CH, Zhang ZY, Zhou LM. NRP-1 expression in bladder cancer and its implications for tumor progression. Tumour Biol. 2014; 35: 6089-94. doi: 10.1007/s13277-014-1806-3.
166. Keck B, Wach S, Taubert H, Zeiler S, Ott OJ, Kunath F, Hartmann A, Bertz S, Weiss C, Honscheid P, Schellenburg S, Rodel C, Baretton GB, et al. Neuropilin-2 and its ligand VEGF-C predict treatment response after transurethral resection and radiochemotherapy in bladder cancer patients. Int J Cancer. 2015; 136: 443-51. doi: 10.1002/ijc.28987.
167. Said N, Sanchez-Carbayo M, Smith SC, Theodorescu D. RhoGDI2 suppresses lung metastasis in mice by reducing tumor versican expression and macrophage infiltration. J Clin Invest. 2012; 122: 1503-18. doi: 10.1172/JCI61392.
168. Wu Y, Siadaty MS, Berens ME, Hampton GM, Theodorescu D. Overlapping gene expression profiles of cell migration and tumor invasion in human bladder cancer identify metallothionein 1E and nicotinamide N-methyltransferase as novel regulators of cell migration. Oncogene. 2008; 27: 6679-89. doi: 10.1038/onc.2008.264.
169. Said N, Theodorescu D. RhoGDI2 suppresses bladder cancer metastasis via reduction of inflammation in the tumor microenvironment. Oncoimmunology. 2012; 1: 1175-7. doi: 10.4161/onci.20594.
170. Cho HJ, Baek KE, Yoo J. RhoGDI2 as a therapeutic target in cancer. Expert Opin Ther Targets. 2010; 14: 67-75. doi: 10.1517/14728220903449251.
171. El Behi M, Krumeich S, Lodillinsky C, Kamoun A, Tibaldi L, Sugano G, De Reynies A, Chapeaublanc E, Laplanche A, Lebret T, Allory Y, Radvanyi F, Lantz O, et al. An essential role for decorin in bladder cancer invasiveness. EMBO Mol Med. 2013; 5: 1835-51. doi: 10.1002/emmm.201302655.
172. Sainio A, Nyman M, Lund R, Vuorikoski S, Bostrom P, Laato M, Bostrom PJ, Jarvelainen H. Lack of decorin expression by human bladder cancer cells offers new tools in the therapy of urothelial malignancies. PLoS One. 2013; 8: e76190. doi: 10.1371/journal.pone.0076190.
173. Iozzo RV, Buraschi S, Genua M, Xu SQ, Solomides CC, Peiper SC, Gomella LG, Owens RC, Morrione A. Decorin antagonizes IGF receptor I (IGF-IR) function by interfering with IGF-IR activity and attenuating downstream signaling. J Biol Chem. 2011; 286: 34712-21. doi: 10.1074/jbc.M111.262766.
174. Morrione A, Neill T, Iozzo RV. Dichotomy of decorin activity on the insulin-like growth factor-I system. FEBS J. 2013; 280: 2138-49. doi: 10.1111/febs.12149.
175. Iozzo RV, Schaefer L. Proteoglycans in health and disease: novel regulatory signaling mechanisms evoked by the small leucine-rich proteoglycans. FEBS J. 2010; 277: 3864-75. doi: 10.1111/j.1742-4658.2010.07797.x.
176. Niedworok C, Rock K, Kretschmer I, Freudenberger T, Nagy N, Szarvas T, Vom Dorp F, Reis H, Rubben H, Fischer JW. Inhibitory role of the small leucine-rich proteoglycan biglycan in bladder cancer. PLoS One. 2013; 8: e80084. doi: 10.1371/journal.pone.0080084.
177. Roudnicky F, Poyet C, Wild P, Krampitz S, Negrini F, Huggenberger R, Rogler A, Stohr R, Hartmann A, Provenzano M, Otto VI, Detmar M. Endocan is upregulated on tumor vessels in invasive bladder cancer where it mediates VEGF-A-induced angiogenesis. Cancer Res. 2013; 73: 1097-106. doi: 10.1158/0008-5472.CAN-12-1855.
178. Fraser JR, Laurent TC, Laurent UB. Hyaluronan: its nature, distribution, functions and turnover. J Intern Med. 1997; 242: 27-33.
179. Kramer MW, Escudero DO, Lokeshwar SD, Golshani R, Ekwenna OO, Acosta K, Merseburger AS, Soloway M, Lokeshwar VB. Association of hyaluronic acid family members (HAS1, HAS2, and HYAL-1) with bladder cancer diagnosis and prognosis. Cancer. 2011; 117: 1197-209. doi: 10.1002/cncr.25565.
180. Niedworok C, Kretschmer I, Rock K, Vom Dorp F, Szarvas T, Hess J, Freudenberger T, Melchior-Becker A, Rubben H, Fischer JW. The impact of the receptor of hyaluronan-mediated motility (RHAMM) on human urothelial transitional cell cancer of the bladder. PLoS One. 2013; 8: e75681. doi: 10.1371/journal.pone.0075681.
181. Kramer MW, Golshani R, Merseburger AS, Knapp J, Garcia A, Hennenlotter J, Duncan RC, Soloway MS, Jorda M, Kuczyk MA, Stenzl A, Lokeshwar VB. HYAL-1 hyaluronidase: a potential prognostic indicator for progression to muscle invasion and recurrence in bladder cancer. Eur Urol. 2010; 57: 86-93. doi: 10.1016/j.eururo.2009.03.057.
182. Hannun YA, Obeid LM. Principles of bioactive lipid signalling: lessons from sphingolipids. Nat Rev Mol Cell Biol. 2008; 9: 139-50. doi: 10.1038/nrm2329.
183. van Meer G, Voelker DR, Feigenson GW. Membrane lipids: where they are and how they behave. Nat Rev Mol Cell Biol. 2008; 9: 112-24. doi: 10.1038/nrm2330.
184. Mullen TD, Hannun YA, Obeid LM. Ceramide synthases at the centre of sphingolipid metabolism and biology. Biochem J. 2012; 441: 789-802. doi: 10.1042/BJ20111626.
185. Holthuis JC, Pomorski T, Raggers RJ, Sprong H, Van Meer G. The organizing potential of sphingolipids in intracellular membrane transport. Physiol Rev. 2001; 81: 1689-723.
186. Hanada K, Kumagai K, Yasuda S, Miura Y, Kawano M, Fukasawa M, Nishijima M. Molecular machinery for non-vesicular trafficking of ceramide. Nature. 2003; 426: 803-9. doi: 10.1038/nature02188.
187. Gault CR, Obeid LM, Hannun YA. An overview of sphingolipid metabolism: from synthesis to breakdown. Adv Exp Med Biol. 2010; 688: 1-23.
188. Kumagai T, Sato T, Natsuka S, Kobayashi Y, Zhou D, Shinkai T, Hayakawa S, Furukawa K. Involvement of murine beta-1,4-galactosyltransferase V in lactosylceramide biosynthesis. Glycoconj J. 2010; 27: 685-95. doi: 10.1007/s10719-010-9313-2.
189. Merrill AH Jr. Sphingolipid and glycosphingolipid metabolic pathways in the era of sphingolipidomics. Chem Rev. 2011; 111: 6387-422. doi: 10.1021/cr2002917.
190. Crespo PM, Iglesias-Bartolome R, Daniotti JL. Ganglioside GD3 traffics from the trans-Golgi network to plasma membrane by a Rab11-independent and brefeldin A-insensitive exocytic pathway. J Biol Chem. 2004; 279: 47610-8. doi: 10.1074/jbc.M407181200.
191. Hakomori SI. Cell adhesion/recognition and signal transduction through glycosphingolipid microdomain. Glycoconjugate Journal. 2000; 17: 143-51. doi: 10.1023/a:1026524820177.
192. Hakomori SI, Zhang Y. Glycosphingolipid antigens and cancer therapy. Chemistry & Biology. 1997; 4: 97-104.
193. Regina Todeschini A, Hakomori SI. Functional role of glycosphingolipids and gangliosides in control of cell adhesion, motility, and growth, through glycosynaptic microdomains. Biochim Biophys Acta. 2008; 1780: 421-33. doi: 10.1016/j.bbagen.2007.10.008.
194. Venable ME, Lee JY, Smyth MJ, Bielawska A, Obeid LM. Role of ceramide in cellular senescence. J Biol Chem. 1995; 270: 30701-8.
195. Lavie Y, Cao H, Bursten SL, Giuliano AE, Cabot MC. Accumulation of glucosylceramides in multidrug-resistant cancer cells. J Biol Chem. 1996; 271: 19530-6.
196. Lucci A, Cho WI, Han TY, Giuliano AE, Morton DL, Cabot MC. Glucosylceramide: a marker for multiple-drug resistant cancers. Anticancer Res. 1998; 18: 475-80.
197. Liu YY, Han TY, Giuliano AE, Cabot MC. Expression of glucosylceramide synthase, converting ceramide to glucosylceramide, confers adriamycin resistance in human breast cancer cells. J Biol Chem. 1999; 274: 1140-6.
198. Sun CC, Zhang Z, Zhang SY, Li J, Li ZL, Kong CZ. Up-regulation of glucosylceramide synthase in urinary bladder neoplasms. Urol Oncol. 2012; 30: 444-9. doi: 10.1016/j.urolonc.2010.04.012.
199. Kawamura S, Ohyama C, Watanabe R, Satoh M, Saito S, Hoshi S, Gasa S, Orikasa S. Glycolipid composition in bladder tumor: a crucial role of GM3 ganglioside in tumor invasion. Int J Cancer. 2001; 94: 343-7.
200. Watanabe R, Ohyama C, Aoki H, Takahashi T, Satoh M, Saito S, Hoshi S, Ishii A, Saito M, Arai Y. Ganglioside G(M3) overexpression induces apoptosis and reduces malignant potential in murine bladder cancer. Cancer Res. 2002; 62: 3850-4.
201. Wang H, Isaji T, Satoh M, Li D, Arai Y, Gu J. Antitumor effects of exogenous ganglioside GM3 on bladder cancer in an orthotopic cancer model. Urology. 2013; 81: 210 e11-5. doi: 10.1016/j.urology.2012.08.015.
202. Todeschini AR, Dos Santos JN, Handa K, Hakomori SI. Ganglioside GM2/GM3 complex affixed on silica nanospheres strongly inhibits cell motility through CD82/cMet-mediated pathway. Proc Natl Acad Sci U S A. 2008; 105: 1925-30. doi: 10.1073/pnas.0709619104.
203. Eisenhaber B, Bork P, Eisenhaber F. Post-translational GPI lipid anchor modification of proteins in kingdoms of life: analysis of protein sequence data from complete genomes. Protein Eng. 2001; 14: 17-25.
204. Ikezawa H. Glycosylphosphatidylinositol (GPI)-anchored proteins. Biol Pharm Bull. 2002; 25: 409-17.
205. Kinoshita T, Fujita M, Maeda Y. Biosynthesis, remodelling and functions of mammalian GPI-anchored proteins: recent progress. J Biochem. 2008; 144: 287-94. doi: 10.1093/jb/mvn090.
206. Zurzolo C, Simons K. Glycosylphosphatidylinositol-anchored proteins: Membrane organization and transport. Biochim Biophys Acta. 2016; 1858: 632-9. doi: 10.1016/j.bbamem.2015.12.018.
207. Montie JE. CDC91L1 (PIG-U) is a newly discovered oncogene in human bladder cancer. J Urol. 2005; 174: 869-70. doi: 10.1097/01.ju.0000171864.79218.0d.
208. Guo Z, Linn JF, Wu G, Anzick SL, Eisenberger CF, Halachmi S, Cohen Y, Fomenkov A, Hoque MO, Okami K, Steiner G, Engles JM, Osada M, et al. CDC91L1 (PIG-U) is a newly discovered oncogene in human bladder cancer. Nat Med. 2004; 10: 374-81. doi: 10.1038/nm1010.
209. Shen YJ, Ye DW, Yao XD, Trink B, Zhou XY, Zhang SL, Dai B, Zhang HL, Zhu Y, Guo Z, Wu G, Nagpal J. Overexpression of CDC91L1 (PIG-U) in bladder urothelial cell carcinoma: correlation with clinical variables and prognostic significance. BJU Int. 2008; 101: 113-9. doi: 10.1111/j.1464-410X.2007.07192.x.
210. Nagpal JK, Dasgupta S, Jadallah S, Chae YK, Ratovitski EA, Toubaji A, Netto GJ, Eagle T, Nissan A, Sidransky D, Trink B. Profiling the expression pattern of GPI transamidase complex subunits in human cancer. Mod Pathol. 2008; 21: 979-91. doi: 10.1038/modpathol.2008.76.
211. Fujita M, Jigami Y. Lipid remodeling of GPI-anchored proteins and its function. Biochim Biophys Acta. 2008; 1780: 410-20. doi: 10.1016/j.bbagen.2007.08.009.
212. Xiaotong H, Hannocks MJ, Hampson I, Brunner G. GPI-specific phospholipase D mRNA expression in tumor cells of different malignancy. Clin Exp Metastasis. 2002; 19: 291-9.
213. Hagikura M, Murakumo Y, Hasegawa M, Jijiwa M, Hagiwara S, Mii S, Hagikura S, Matsukawa Y, Yoshino Y, Hattori R, Wakai K, Nakamura S, Gotoh M, et al. Correlation of pathological grade and tumor stage of urothelial carcinomas with CD109 expression. Pathol Int. 2010; 60: 735-43. doi: 10.1111/j.1440-1827.2010.02592.x.
214. Leyvraz C, Charles RP, Rubera I, Guitard M, Rotman S, Breiden B, Sandhoff K, Hummler E. The epidermal barrier function is dependent on the serine protease CAP1/Prss8. J Cell Biol. 2005; 170: 487-96. doi: 10.1083/jcb.200501038.
215. Chen M, Chen LM, Lin CY, Chai KX. The epidermal growth factor receptor (EGFR) is proteolytically modified by the Matriptase-Prostasin serine protease cascade in cultured epithelial cells. Biochim Biophys Acta. 2008; 1783: 896-903. doi: 10.1016/j.bbamcr.2007.10.019.
216. Chen LM, Verity NJ, Chai KX. Loss of prostasin (PRSS8) in human bladder transitional cell carcinoma cell lines is associated with epithelial-mesenchymal transition (EMT). BMC Cancer. 2009; 9: 377. doi: 10.1186/1471-2407-9-377.
217. Gailey MP, Bellizzi AM. Immunohistochemistry for the novel markers glypican 3, PAX8, and p40 (DeltaNp63) in squamous cell and urothelial carcinoma. Am J Clin Pathol. 2013; 140: 872-80. doi: 10.1309/AJCP4NSKW5TLGTDS.
218. Saikali Z, Sinnett D. Expression of glypican 3 (GPC3) in embryonal tumors. Int J Cancer. 2000; 89: 418-22.
219. Grisaru S, Cano-Gauci D, Tee J, Filmus J, Rosenblum ND. Glypican-3 modulates BMP- and FGF-mediated effects during renal branching morphogenesis. Dev Biol. 2001; 231: 31-46. doi: 10.1006/dbio.2000.0127.
220. Hippo Y, Watanabe K, Watanabe A, Midorikawa Y, Yamamoto S, Ihara S, Tokita S, Iwanari H, Ito Y, Nakano K, Nezu J, Tsunoda H, Yoshino T, et al. Identification of soluble NH2-terminal fragment of glypican-3 as a serological marker for early-stage hepatocellular carcinoma. Cancer Res. 2004; 64: 2418-23.
221. Xylinas E, Cha EK, Khani F, Kluth LA, Rieken M, Volkmer BG, Hautmann R, Kufer R, Chen YT, Zerbib M, Rubin MA, Scherr DS, Shariat SF, et al. Association of oncofetal protein expression with clinical outcomes in patients with urothelial carcinoma of the bladder. J Urol. 2014; 191: 830-41. doi: 10.1016/j.juro.2013.08.048.
222. Urquidi V, Chang M, Dai Y, Kim J, Wolfson ED, Goodison S, Rosser CJ. IL-8 as a urinary biomarker for the detection of bladder cancer. BMC Urol. 2012; 12: 12. doi: 10.1186/1471-2490-12-12.
223. Lima L, Oliveira D, Ferreira JA, Tavares A, Cruz R, Medeiros R, Santos L. The role of functional polymorphisms in immune response genes as biomarkers of bacille Calmette-Guerin (BCG) immunotherapy outcome in bladder cancer: establishment of a predictive profile in a Southern Europe population. BJU Int. 2015; 116: 753-63. doi: 10.1111/bju.12844.
224. Ernst B, Magnani JL. From carbohydrate leads to glycomimetic drugs. Nat Rev Drug Discov. 2009; 8: 661-77. doi: 10.1038/nrd2852.
225. Boltje TJ, Buskas T, Boons GJ. Opportunities and challenges in synthetic oligosaccharide and glycoconjugate research. Nat Chem. 2009; 1: 611-22. doi: 10.1038/nchem.399.
226. Lepenies B, Yin J, Seeberger PH. Applications of synthetic carbohydrates to chemical biology. Curr Opin Chem Biol. 2010; 14: 404-11. doi: 10.1016/j.cbpa.2010.02.016.
227. Zhu X, Schmidt RR. New principles for glycoside-bond formation. Angew Chem Int Ed Engl. 2009; 48: 1900-34. doi: 10.1002/anie.200802036.
228. Alley WR Jr, Mann BF, Novotny MV. High-sensitivity analytical approaches for the structural characterization of glycoproteins. Chem Rev. 2013; 113: 2668-732. doi: 10.1021/cr3003714.
229. Zaia J. Mass spectrometry and the emerging field of glycomics. Chem Biol. 2008; 15: 881-92. doi: 10.1016/j.chembiol.2008.07.016.
230. Li B, Mock F, Wu P. Imaging the glycome in living systems. Methods Enzymol. 2012; 505: 401-19. doi: 10.1016/B978-0-12-388448-0.00029-2.
231. Laughlin ST, Bertozzi CR. Imaging the glycome. Proc Natl Acad Sci U S A. 2009; 106: 12-7. doi: 10.1073/pnas.0811481106.
232. Liang PH, Wu CY, Greenberg WA, Wong CH. Glycan arrays: biological and medical applications. Curr Opin Chem Biol. 2008; 12: 86-92. doi: 10.1016/j.cbpa.2008.01.031.
233. Kirwan A, Utratna M, O’Dwyer ME, Joshi L, Kilcoyne M. Glycosylation-Based Serum Biomarkers for Cancer Diagnostics and Prognostics. Biomed Res Int. 2015; 2015: 490531. doi: 10.1155/2015/490531.
234. Pall M, Iqbal J, Singh SK, Rana SV. CA 19-9 as a serum marker in urothelial carcinoma. Urol Ann. 2012; 4: 98-101. doi: 10.4103/0974-7796.95555.
235. Roy S, Dasgupta A, Kar K. Comparison of urinary and serum CA 19-9 as markers of early stage urothelial carcinoma. Int Braz J Urol. 2013; 39: 631-8. doi: 10.1590/S1677-5538.IBJU.2013.05.04.
236. Nagao K, Itoh Y, Fujita K, Fujime M. Evaluation of urinary CA19-9 levels in bladder cancer patients classified according to the combinations of Lewis and Secretor blood group genotypes. Int J Urol. 2007; 14: 795-9. doi: 10.1111/j.1442-2042.2007.01840.x.
237. Pal K, Roy S, Mondal SA, Chatterjee U, Tiwari P, Bera M. Urinary level of CA19-9 as a tumor marker in urothelial carcinoma of the bladder. Urol J. 2011; 8: 203-8.
238. Chuang CH, Wu TF, Chen CH, Chang KC, Ju JW, Huang YW, Van Nhan V. Lab on a chip for multiplexed immunoassays to detect bladder cancer using multifunctional dielectrophoretic manipulations. Lab Chip. 2015; 15: 3056-64. doi: 10.1039/c5lc00352k.
239. Osman I, Bajorin DF, Sun TT, Zhong H, Douglas D, Scattergood J, Zheng R, Han M, Marshall KW, Liew CC. Novel blood biomarkers of human urinary bladder cancer. Clin Cancer Res. 2006; 12: 3374-80. doi: 10.1158/1078-0432.CCR-05-2081.
240. Laloglu E, Aksoy H, Aksoy Y, Ozkaya F, Akcay F. The determination of serum and urinary endocan concentrations in patients with bladder cancer. Ann Clin Biochem. 2016; 53: 647-653. doi: 10.1177/0004563216629169.
241. Jamshidian H, Hashemi M, Nowroozi MR, Ayati M, Bonyadi M, Najjaran Tousi V. Sensitivity and specificity of urinary hyaluronic acid and hyaluronidase in detection of bladder transitional cell carcinoma. Urol J. 2014; 11: 1232-7.
242. Pham HT, Block NL, Lokeshwar VB. Tumor-derived hyaluronidase: a diagnostic urine marker for high-grade bladder cancer. Cancer Res. 1997; 57: 778-83.
243. Lokeshwar VB, Obek C, Soloway MS, Block NL. Tumor-associated hyaluronic acid: a new sensitive and specific urine marker for bladder cancer. Cancer Res. 1997; 57: 773-7.
244. Passerotti CC, Srougi M, Bomfim AC, Martins JR, Leite KR, Dos Reis ST, Sampaio LO, Ortiz V, Dietrich CP, Nader HB. Testing for urinary hyaluronate improves detection and grading of transitional cell carcinoma. Urol Oncol. 2011; 29: 710-5. doi: 10.1016/j.urolonc.2009.10.006.
245. Lokeshwar VB, Obek C, Pham HT, Wei D, Young MJ, Duncan RC, Soloway MS, Block NL. Urinary hyaluronic acid and hyaluronidase: markers for bladder cancer detection and evaluation of grade. J Urol. 2000; 163: 348-56.
246. Hautmann SH, Lokeshwar VB, Schroeder GL, Civantos F, Duncan RC, Gnann R, Friedrich MG, Soloway MS. Elevated tissue expression of hyaluronic acid and hyaluronidase validates the HA-HAase urine test for bladder cancer. J Urol. 2001; 165: 2068-74.
247. Golshani R, Hautmann SH, Estrella V, Cohen BL, Kyle CC, Manoharan M, Jorda M, Soloway MS, Lokeshwar VB. HAS1 expression in bladder cancer and its relation to urinary HA test. Int J Cancer. 2007; 120: 1712-20. doi: 10.1002/ijc.22222.
248. Hautmann S, Toma M, Lorenzo Gomez MF, Friedrich MG, Jaekel T, Michl U, Schroeder GL, Huland H, Juenemann KP, Lokeshwar VB. Immunocyt and the HA-HAase urine tests for the detection of bladder cancer: a side-by-side comparison. Eur Urol. 2004; 46: 466-71. doi: 10.1016/j.eururo.2004.06.006.
249. Lokeshwar VB, Schroeder GL, Selzer MG, Hautmann SH, Posey JT, Duncan RC, Watson R, Rose L, Markowitz S, Soloway MS. Bladder tumor markers for monitoring recurrence and screening comparison of hyaluronic acid-hyaluronidase and BTA-Stat tests. Cancer. 2002; 95: 61-72. doi: 10.1002/cncr.10652.
250. Ferreira JA, Domingues MR, Reis A, Monteiro MA, Coimbra MA. Differentiation of isomeric Lewis blood groups by positive ion electrospray tandem mass spectrometry. Anal Biochem. 2010; 397: 186-96. doi: 10.1016/j.ab.2009.10.034.
251. Everest-Dass AV, Abrahams JL, Kolarich D, Packer NH, Campbell MP. Structural feature ions for distinguishing N- and O-linked glycan isomers by LC-ESI-IT MS/MS. J Am Soc Mass Spectrom. 2013; 24: 895-906. doi: 10.1007/s13361-013-0610-4.
252. Jensen PH, Karlsson NG, Kolarich D, Packer NH. Structural analysis of N- and O-glycans released from glycoproteins. Nat Protoc. 2012; 7: 1299-310. doi: 10.1038/nprot.2012.063.
253. Han L, Costello CE. Mass spectrometry of glycans. Biochemistry (Mosc). 2013; 78: 710-20. doi: 10.1134/S0006297913070031.
254. Stavenhagen K, Kolarich D, Wuhrer M. Clinical Glycomics Employing Graphitized Carbon Liquid Chromatography-Mass Spectrometry. Chromatographia. 2015; 78: 307-20. doi: 10.1007/s10337-014-2813-7.
255. Hajba L, Csanky E, Guttman A. Liquid phase separation methods for N-glycosylation analysis of glycoproteins of biomedical and biopharmaceutical interest. A critical review. Anal Chim Acta. 2016; 943: 8-16. doi: 10.1016/j.aca.2016.08.035.
256. Struwe WB, Agravat S, Aoki-Kinoshita KF, Campbell MP, Costello CE, Dell A, Ten F, Haslam SM, Karlsson NG, Khoo KH, Kolarich D, Liu Y, McBride R, et al. The minimum information required for a glycomics experiment (MIRAGE) project: sample preparation guidelines for reliable reporting of glycomics datasets. Glycobiology. 2016; 26: 907-10. doi: 10.1093/glycob/cww082.
257. York WS, Agravat S, Aoki-Kinoshita KF, McBride R, Campbell MP, Costello CE, Dell A, Feizi T, Haslam SM, Karlsson N, Khoo KH, Kolarich D, Liu Y, et al. MIRAGE: the minimum information required for a glycomics experiment. Glycobiology. 2014; 24: 402-6. doi: 10.1093/glycob/cwu018.
258. Hu H, Khatri K, Klein J, Leymarie N, Zaia J. A review of methods for interpretation of glycopeptide tandem mass spectral data. Glycoconj J. 2016; 33: 285-96. doi: 10.1007/s10719-015-9633-3.
259. Bennun SV, Hizal DB, Heffner K, Can O, Zhang H, Betenbaugh MJ. Systems Glycobiology: Integrating Glycogenomics, Glycoproteomics, Glycomics, and Other ‘Omics Data Sets to Characterize Cellular Glycosylation Processes. J Mol Biol. 2016; 428: 3337-52. doi: 10.1016/j.jmb.2016.07.005.
260. Pochec E, Litynska A, Bubka M, Amoresano A, Casbarra A. Characterization of the oligosaccharide component of alpha3beta1 integrin from human bladder carcinoma cell line T24 and its role in adhesion and migration. Eur J Cell Biol. 2006; 85: 47-57. doi: 10.1016/j.ejcb.2005.08.010.
261. Choi W, Porten S, Kim S, Willis D, Plimack ER, Hoffman-Censits J, Roth B, Cheng T, Tran M, Lee IL, Melquist J, Bondaruk J, Majewski T, et al. Identification of distinct basal and luminal subtypes of muscle-invasive bladder cancer with different sensitivities to frontline chemotherapy. Cancer Cell. 2014; 25: 152-65. doi: 10.1016/j.ccr.2014.01.009.
262. Choi W, Czerniak B, Ochoa A, Su X, Siefker-Radtke A, Dinney C, McConkey DJ. Intrinsic basal and luminal subtypes of muscle-invasive bladder cancer. Nat Rev Urol. 2014; 11: 400-10. doi: 10.1038/nrurol.2014.129.
263. Hedegaard J, Lamy P, Nordentoft I, Algaba F, Hoyer S, Ulhoi BP, Vang S, Reinert T, Hermann GG, Mogensen K, Thomsen MB, Nielsen MM, Marquez M, et al. Comprehensive Transcriptional Analysis of Early-Stage Urothelial Carcinoma. Cancer Cell. 2016; 30: 27-42. doi: 10.1016/j.ccell.2016.05.004.
264. Bernardo C, Costa C, Amaro T, Goncalves M, Lopes P, Freitas R, Gartner F, Amado F, Ferreira JA, Santos L. Patient-derived sialyl-Tn-positive invasive bladder cancer xenografts in nude mice: an exploratory model study. Anticancer Res. 2014; 34: 735-44.
265. Cox KM, Sterling JD, Regan JT, Gasdaska JR, Frantz KK, Peele CG, Black A, Passmore D, Moldovan-Loomis C, Srinivasan M, Cuison S, Cardarelli PM, Dickey LF. Glycan optimization of a human monoclonal antibody in the aquatic plant Lemna minor. Nat Biotechnol. 2006; 24: 1591-7. doi: 10.1038/nbt1260.
266. Yamane-Ohnuki N, Kinoshita S, Inoue-Urakubo M, Kusunoki M, Iida S, Nakano R, Wakitani M, Niwa R, Sakurada M, Uchida K, Shitara K, Satoh M. Establishment of FUT8 knockout Chinese hamster ovary cells: an ideal host cell line for producing completely defucosylated antibodies with enhanced antibody-dependent cellular cytotoxicity. Biotechnol Bioeng. 2004; 87: 614-22. doi: 10.1002/bit.20151.
267. Lu J, Chu J, Zou Z, Hamacher NB, Rixon MW, Sun PD. Structure of FcgammaRI in complex with Fc reveals the importance of glycan recognition for high-affinity IgG binding. Proc Natl Acad Sci U S A. 2015; 112: 833-8. doi: 10.1073/pnas.1418812112.
268. van Kooyk Y, Rabinovich GA. Protein-glycan interactions in the control of innate and adaptive immune responses. Nat Immunol. 2008; 9: 593-601. doi: 10.1038/ni.f.203.
269. Everest-Dass AV, Briggs MT, Kaur G, Oehler MK, Hoffmann P, Packer NH. N-glycan MALDI Imaging Mass Spectrometry on Formalin-Fixed Paraffin-Embedded Tissue Enables the Delineation of Ovarian Cancer Tissues. Mol Cell Proteomics. 2016; 15: 3003-16. doi: 10.1074/mcp.M116.059816.