
INTRODUCTION
Malignant pleural mesothelioma (MPM) is a rare but aggressive neoplasm that has been the subject of considerable attention given its strong relationship to asbestos and its dismal outcome despite continuous efforts to intensify its treatment. Chemotherapy, radiotherapy, and surgery have been proven ineffective as single treatment modalities, but combined modality regimes have yielded minor improvement in disease progression [1]. However, no definite treatment guidelines have been established, because of the poor quality of the published evidence [2]. In particular, the impact of adjuvant high-dose hemithoracic radiotherapy after surgical complete resection following initial neoadjuvant chemotherapy remains controversial [3–6]. In addition, a recent phase 2 trial of postoperative radiotherapy in MPM showed no survival benefit [7]. It is also reported that, despite some small improvements in local control of patients with postoperative radiation, regional relapse remains the predominant pattern of tumor recurrence after multimodality treatment [8].
Photon beams are routinely used for external-beam radiotherapy. In order to predict and avoid radiation injury to healthy tissues, dose limitations for such tissues and organs have been described. During the past decades, innovative delivery techniques for photon irradiation, which is the most often used modality for radiation treatments, have evolved that allow dose accumulation within precisely contoured targets while uninvolved structures in close vicinity can be spared. Three-dimensional treatment planning, stereotactic setups and intensity-modulated radiotherapy (IMRT) have substantially improved dose deposition with minimized exposure of adjacent organs. However, some recent reports have still shown high rates of local in-field or marginal recurrences and significant toxicity after adjuvant radiotherapy using photon IMRT (16-48%) [9–11]. As the principal and unalterable physical characteristics of photons imply gradual energy deposition, a new modality – particle irradiation – has been suggested to continuously replace photon treatments: the physical and biological properties of particle irradiation facilitate dose-escalated treatments with significantly reduced dose exposure to uninvolved peritumoral tissues and organs.
The repeatedly reported notion of post-radiation tumor recurrences occurring mostly within areas of prior photon dose exposure has raised the question of whether mesothelioma cells might increase their motility and migrate from radiation fields before lethal doses have been administered during photon radiotherapy. Several authors have demonstrated in other malignancies, such as lung cancer [12–14], prostate cancer [15], melanoma [16], fibrosarcomas [17], and glioma [18], that low doses of photon irradiation can stimulate mechanisms of cellular migration and alter the peritumoral stroma into a promigratory milieu through integrin induction [12, 17], transformation of extracellular matrix proteins [13], activation of matrix-metalloproteinases [15, 16, 18], and stimulation of chemokine secretion. Among the many different chemokines, SDF-1α has repeatedly been identified to account for radiation-induced motility in thoracic malignancies [14].
Therefore, we hypothesized that low doses of photon irradiation might promote the migration of MPM cells. The purpose of this study was to investigate the motility of MPM cells after low doses of photon and particle irradiation and to elucidate the mechanisms of the detected phenotypes.
RESULTS
Mesothelioma cell migration was promoted by serum components
In order to analyze the migration ability of mesothelioma, modified Boyden chamber assays were performed to analyze transmigration of H28 and H226 mesothelioma cells through 8-μm pore size polycarbonate membranes coated with collagen I and IV. Both cell lines were attracted by serum in a concentration-dependent manner (5% and 10%). Moreover, SDF-1α, which is a typical pro-migratory serum component, also promoted the migration of H28 through collagen I- and IV-coated membranes (p=0.007 and p<0.001, respectively) and H226 cells through collagen I-coated membranes (p<0.001) (Figure 1).
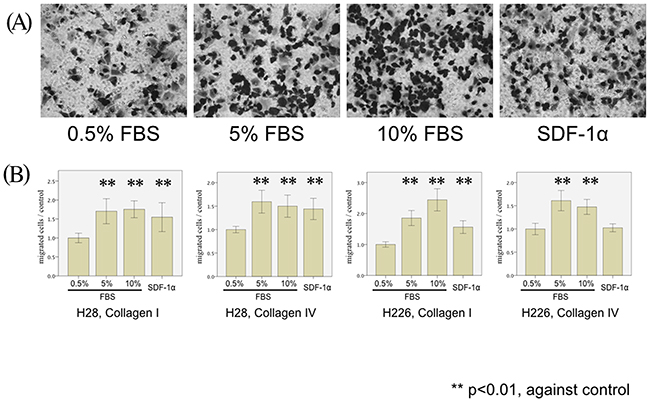
Figure 1: MPM cell migration stimulated by FBS and SDF-1α. (A) H226 cells are microscopically analyzed after the 5-hour migration assay using 0.5% FBS (left), 5% FBS (center left), 10% FBS (center right), and SDF-1α (0.1μg/ml) (right) as chemoattractants and Collagen I-coated membranes (magnification x20). (B) H28 cell migration through Collagen I- (left Figure) and Collagen IV- (middle left figure) coated membranes and H226 cell migration through Collagen I- (middle right Figure) and Collagen IV- (right Figure) coated membranes were examined in the modified Boyden chamber experiments. The number of stimulated MPM cells (H28 and H226) transmigrated through collagen I- and collagen IV-coated membranes was counted. In each Figure, the average cell counts after the 5-hour migration assay using 0.5% FBS is defined as a control, and the normalized ratio of each cell count to the control is calculated.
Low-doses of photon irradiation enhance mesothelioma cell migration
We investigated SDF-1α-induced transmigration of both mesothelioma cell lines through membranes coated with collagen I and IV following single doses of photon irradiation (1 Gy and 2 Gy): Irradiation of both H28 and H226 cells with single photon doses of 1 Gy and 2 Gy increased transmigration through collagen I- and collagen IV-coated filters. Compared with non-irradiated cells, a significant increase of cell migration through collagen I-coated filters was noted after 1 Gy and 2 Gy in H28 cells (p=0.009 and p=0.003, respectively), and on collagen IV-coated filters after 2 Gy (p<0.001). In H226 cells, irradiation with 1 Gy and 2 Gy increased transmigration through both collagen I and IV-coated filters (p<0.001 and p=0.003, respectively) (Figure 2).
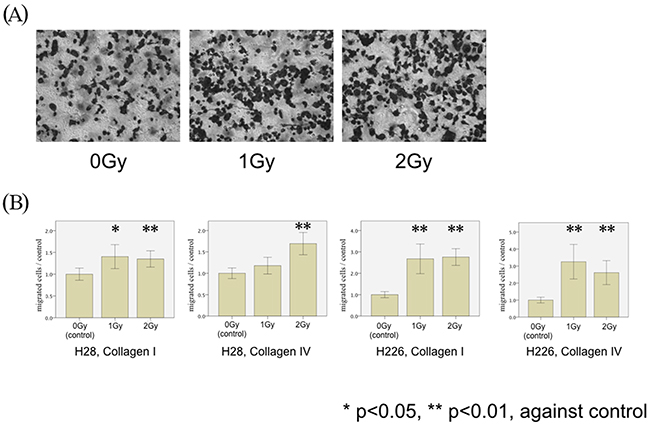
Figure 2: MPM cell migration following photon irradiation. (A) H226 cells are microscopically analyzed without (left) and with irradiation by single doses of 1 Gy (middle) and 2 Gy (right) in modified Boyden chamber experiments using Collagen I-coated membranes (magnification x20). (B) H28 cell migration through Collagen I- (left Figure) and Collagen IV- (middle left Figure) coated membranes and H226 cells through Collagen I- (middle right Figure) and Collagen IV- (right Figure) coated membranes were examined in the modified Boyden chamber experiments. The number of irradiated MPM cells (H28 and H226) transmigrated through collagen I- and collagen IV-coated membranes was counted. The average number of migrated cells in the non-irradiated condition is defined as a control, and the normalized ratio of each cell count to the control is calculated.
Photon irradiation increased the expression of CXCR4, but not of α3, α5, or β1 integrins
In order to clarify the underlying mechanism of photon-induced hypermigration, FACS analyses were performed to investigate alterations in integrin expression caused by irradiation. No significant modifications of α3, α5, or β1 integrin expression were observed in both H28 and H226 cells following photon irradiation (Figure 3A). On the other hand, the expression of CXCR4 was significantly increased on H28 cells after single doses of 2 Gy photon irradiation (p=0.005) and on H226 cells after single doses of 1 Gy and 2 Gy photon irradiation (p<0.001 and p=0.033, respectively) (Figure 3A). Therefore, CXCR4 was selected for subsequent investigations.
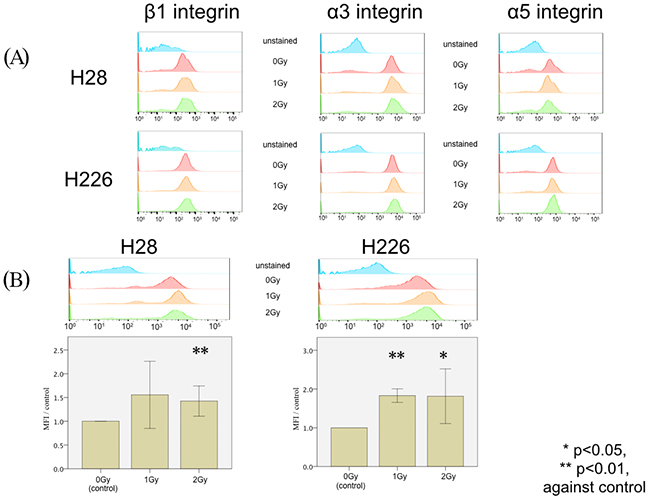
Figure 3: FACS analysis of photon-induced stimulation of expression in cell surface markers. (A) FACS analysis of the expression of β1 integrin (left column), α3 integrin (middle column), and α5 integrin (right column) does not reveal any difference among H28 (upper row) or H226 cells (lower row) without irradiation, with 1 Gy irradiation, and with 2 Gy irradiation. Unstained cells were analyzed in the same experiment as a negative control. (B) FACS histogram analysis of CXCR4 expression (upper row) upon a single dose of 1 Gy irradiation and 2 Gy irradiation in H28 cells (left) and H226 cells (right) compared to non-radiated cells. Data are given as the normalized ratio of CXCR4 expression compared to non-irradiated cells.
Inhibition of CXCR4 significantly impaired photon-induced hypermigration
In order to inhibit CXCR4, we selected AMD3100 in this study. The exposure of both cell lines to AMD3100 24 hours before the migration assays almost fully reversed the detected phenotype of photon-induced migration and significantly inhibited the previously radiation-enhanced migration through collagen I/IV-coated membranes of both H28 and H226 cells after exposition to photon radiation with 2 Gy (H28: p=0.024 and p=0.016; H226: p<0.001 and p=0.001, respectively). However, AMD3100 did not affect the migration of non-irradiated MPM cells (Figure 4).
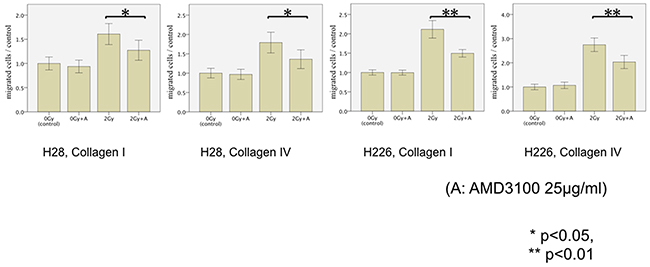
Figure 4: MPM cell migration following photon irradiation and treatment of cells with AMD3100 (25μg/ml). The transmigration of H28 cells through Collagen I-coated membrane (left), H28 cells through Collagen IV-coated membrane (middle left), H226 cells through Collagen I-coated membrane (middle right), and H226 cells through Collagen IV-coated membrane (right) is examined in the modified Boyden chamber experiments In each Figure, the average cell count of non-irradiated cells is defined as a control. Results are displayed as normalized ratio of cell counts compared to the control.
Carbon-ion irradiation did not stimulate CXCR4-SDF1α signaling
Next, we focused on the effect of carbon-ion irradiation instead of photon irradiation. We detected a minor, insignificant increase in migration with both cell lines following carbon ion doses of 0.3 and 1 Gy (Figure 5A). The relative carbon ion irradiation-associated stimulation of cell migration was significantly inferior to all corresponding photon doses (Figure 5A, */**). To rule out, the observed differences were due to radiation-specific differences in cell survival, we performed clonogenic survival experiments. Both photon and carbon ion irradiation significantly impaired clonogenicity; but there was no significant difference in clonogenic cell survival between both cell lines after irradiation with corresponding photon and carbon doses (Figure 5B). Therefore, cell survival did not account for the result of the radiation modality-specific migratory findings. Moreover, Annexin-V assays showed that no significant difference was found between either cell lines with and without irradiation (data not shown). From these data, loss of clonogenicity or apoptosis were not responsible for the detected migration phenotypes. In addition, FACS analysis revealed that carbon ion irradiation did not stimulate CXCR4 expression on the surface of either cell line as opposed to the photon-induced upregulation of CXCR4 (Figure 5C).
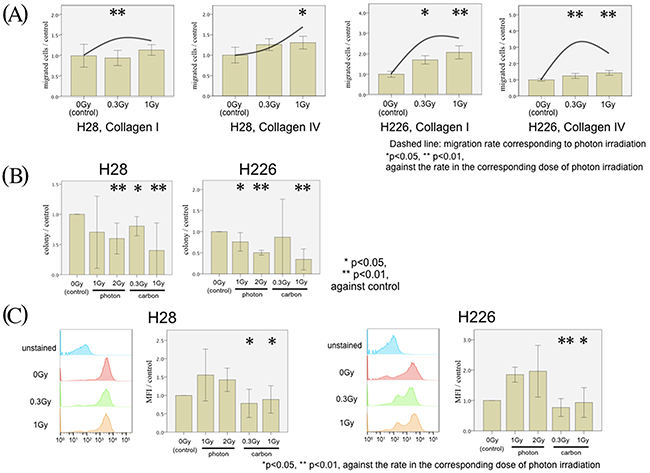
Figure 5: MPM cell migration following carbon irradiation. (A) H28 cell migration through Collagen I- (left figure) and Collagen IV- (middle left figure) coated membranes and H226 cells through Collagen I- (middle right figure) and Collagen IV- (right figure) coated membranes are examined in the modified Boyden chamber experiments. Transmigrated MPM cells without irradiation (left bar) with 0.3 Gy carbon ion irradiation (middle bar) and with 1 Gy carbon ion irradiation (right bar) were counted. In each Figure, the average of the cell count without irradiation is defined as the control, and the normalized ratio of each cell count to the control is analyzed. The mean migration rate of corresponding photon irradiation is presented as a dashed line. (B) Analysis of clonogenic survival with and without photon and carbon ion irradiation in H28 and H226 cells. (C) In each Figure, MFI of the CXCR4 expression without irradiation is defined as a control, and the normalized ratio of each MFI to the control is calculated.
Hypothesizing that the difference in SDF-1α/CXCR4 signaling was responsible for the observed photon vs. carbon ion phenotype, we next performed ELISA, western blots, and knock-down experiments with siRNA to substantiate our findings. CXCR4 ELISA assays confirmed the photon increase of CXCR4 translation in both photon-irradiated cell lines, whereas no such increase was detected in the carbon ion experiments (Figure 6A).
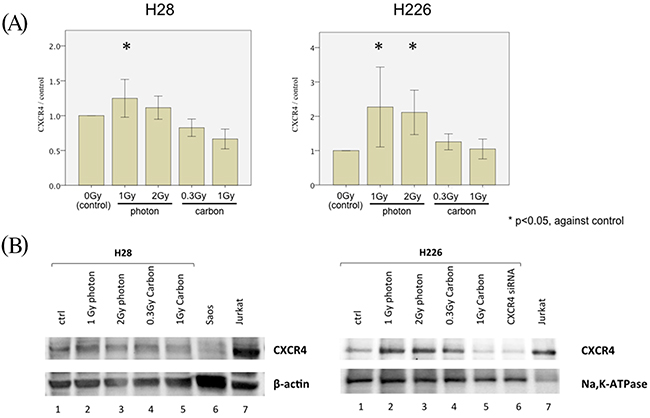
Figure 6: CXCR4 regulation of MPM cell migration following irradiation. (A) CXCR4 ELISA measuring the concentration of CXCR4 among the total protein lysates generated from H28 cells (left) and H226 cells (right). Cells were treated with a single dose of 1 Gy and 2 Gy photon-irradiation or 0.3 Gy and 1 Gy carbon ion irradiation prior to protein lysate generation. In each Figure, the concentration of CXCR4 is depicted as normalized ratio of measured CXCR4 concentration in analyzed samples to the CXCR4 concentration in protein lysate generated from non-irradiated cells (defined as control). (B) Western blot analysis using 20μg of total protein from H28 cells (left) and 20μg membrane-extracted protein from H226 cells (right). H28 and H226 cells were irradiated with a single dose of 1 Gy photon, 2 Gy photon, 0.3 Gy carbon ions and 1 Gy carbon ions before generation of protein lysates. Furthermore, protein was extracted from H226 cells following CXCR4 siRNA treatment. Beta-actin and Na+-K+ ATPase are used as loading controls of total protein and membrane-extracted protein, respectively. Saos-2 cell lysate are included as negative control and Jurkat cell lysate as positive control for CXCR4 protein expression.
Western blots for CXCR4 in membrane-bound proteins from H226 cells identified the responsible and siRNA-suppressible protein band for CXCR4. Subsequent western blots from both cell lines confirmed an upregulation of CXCR4 translation following photon irradiation with 1 and 2 Gy, while no change in protein levels was detected through western blots following carbon ion doses (Figure 6B).
To address the possible contribution of the CXCR4-ligand SDF-1α, we performed ELISA assays which showed that the concentration of soluble SDF-1α in the supernatants of irradiated cells was below the detection limits in all settings of the cultured cells (data not shown).
DISCUSSION
We showed that the common clinical finding of marginal MPM recurrences after radiotherapy may be related to photon-induced ECM-based tumor cell hypermigration towards CXCR4-ligands and that this phenomenon may be inhibited either by the addition of CXCR4-antagonists or by replacement of photon irradiation with carbon ion irradiation.
Given the anatomical and pathological charac-teristics in MPM, surgical resection will inevitably result in an incomplete resection of the tumor microscopically [19]. Nevertheless, surgical macroscopic complete resection prolongs survival and, therefore, it plays a significant role in the multimodal therapy of MPM [20]. Photon radiotherapy, commonly administered in the adjuvant setting, is intended to enhance local control rates – but it has repeatedly been reported to cause inacceptable toxicity and to fail in controlling the disease – especially at the borders of prior radiation fields. Speculations about photon irradiation promoting locoregional relapses have previously been supported by findings from several other tumor cell lines.
Signaling between SDF-1α and CXCR4 contributes to tumor growth, angiogenesis, invasion and metastases in numerous malignancies [21–23]. In addition, irradiation-enhanced invasiveness of non-small lung cancer and breast cancer has been attributed to the SDF-1α/CXCR4 interaction [14, 24]. Moreover, the relationship between MPM and the SDF-1α/CXCR4 chemotactic axis has been reported in previous reports [25, 26]. Therefore we selected the CXCR4- SDF-1α pathway as a promising candidate to elucidate the underlying mechanisms of radiation-altered cell migration. To our best knowledge, our study was the first report to demonstrate that photon-induced migration in MPM cells was primarily caused by increased expression of SDF-1α -binding CXCR4.
Since prior findings in irradiated primary brain tumor cells [27, 28], NSCLC cells [29], and non-malignant cells [30] had revealed that increases of integrin cell surface expressions were responsible for radiation-induced hypermigration, we also decided to analyze the expression of various integrins in MPM cells. However, our results showed that no radiation-associated changes of either α3-, α5-, or β1-family integrin expressions, which had previously been reported as collagen I- and IV-binding integrins in MPM [31–33], were responsible for the detected phenotype of photon-increased hypermigration in MPM cells. Therefore, we concluded that the finding of photon-induced MPM cell migration was related to photon-induced increases in responsiveness towards chemoattractants rather than increased MPM cell interactions with the ECM.
Our preclinical data suggest two possible options to overcome this treatment-related adverse event: One option could be the concurrent use of a SDF-1α/CXCR4 inhibitor with photon irradiation. From our results, AMD3100, an FDA approved antagonist of CXCR4, is effective to prevent radiation-promoted migration of MPM cells in vitro; therefore, it may be assumed that it may be a promising agent to be investigated in clinical trials that might reduce the rate of marginal/local recurrences in MPM patients following radiotherapy.
A further option may be changing the radiation modality from photon to carbon ion radiotherapy. Carbon ion irradiation offers several advantages when compared with conventional photon radiotherapy, including higher relative biologic effectiveness in tumor cell killing and a more accurate dose distribution, which contributes to the low risk of side effect [34–37], although carbon ion treatment facilities are rare and cause significantly higher treatment costs than conventional photon facilities. Moreover, it has been reported that low doses of carbon ion irradiation attenuate the ability of migration in other malignancies, such as lung cancer [38, 39], pancreatic cancer [40], glioma [27, 41], and fibrosarcoma [18]; therefore, it is believed that carbon ion irradiation may have a different effect on radiation-induced migration compared with conventional photon irradiation. Especially, carbon ion irradiation is known to induce specific subcellular alterations including unique gene expression or enzyme activity profiles. The underlying mechanisms of these alterations have previously been reported, suggesting the activation of the Akt signaling pathway [39] and the inhibition of MMP-2 activity [18, 42]. Our results showed that carbon-ion irradiation did not promote the migration of MPM cell lines, nor did it increase the expression of CXCR4, suggesting that carbon ion irradiation is expected to prevent the recurrence of MPM. For this reason, the use of carbon ion irradiation for MPM patients, especially in an adjuvant lower-dose radiotherapy, should have positive effects and be tested in prospective clinical trials.
Our study has several limitations. First, this study is only an in vitro study, which may not accurately reflect the in vivo situation. Moreover, the effect of photon irradiation on the microenvironment surrounding the tumor should not be ignored. In order to confirm this mechanism and to address the contribution of microenvironmental modifications through radiation, radiation experiments in animal models with human MPM xenografts would be a promising approach for a subsequent corroborating study [43].
As another limitation, SDF-1α/CXCR4 signaling is not the only pathway to promote tumor cell migration. Other pathways may contribute simultaneously, and the SDF-1α/CXCR4 pathway may work indirectly through further up- and downstream signaling pathways, such as the interaction of other chemokines and chemokine receptors. In this study, we only examined the SDF-1α/ CXCR4 pathway and the possible role of integrins, as they have repeatedly been identified to be responsible for radiation-increased cell motility and as there is a pharmacological agent, FDA-approved AMD3100, that can be administered to reverse the observed events.
In order to exclude the potential contribution of further pathways, comprehensive analysis, for example through DNA microarray, RNA-ChiP, or Proteomic analysis, would be required. In the present manuscript, we decided to not perform such experiments, as the detected phenotype was successfully inhibited by adding AMD3100.
In conclusion, low doses of photon irradiation promoted MPM cell migration through the increased expression of CXCR4 with subsequently increased SDF-1α/CXCR4 signaling. Clinically, this might enhance the risk of tumor cell spread and infiltration and, therefore, explain prior disappointing results from clinical trials investigating photon radiotherapy in MPM patients. The administration of the CXCR4 antagonist AMD3100 effectively inhibited this increased migration. As an alternative to photon irradiation, carbon ion irradiation did not significantly promote migration. Therefore, our finding suggest that the co-administration of the clinically already available CXCR4 antagonist AMD3100 concurrently to photon irradiation or, alternatively, the replacement of photon irradiation with carbon ion irradiation may be two possible solutions to establish and enhance the clinical benefit of radiation treatments in MPM patients.
MATERIALS AND METHODS
Reagents and cell lines
H28 and H226 mesothelioma cells were purchased from ATCC and maintained at 37°C and 5% CO2 in RPMI 1640 medium supplemented with 1% Penicillin/Streptomycin and 10% fetal bovine serum (FBS; Biochrom, Berlin, Germany). Twenty-four hours before the migration assay, cells were serum starved in RPMI 1640 medium containing 1% Penicillin/Streptomycin and 0.5% FBS. Cell passaging was performed every week.
Stromal cell-derived factor 1α (SDF-1α) was purchased from Gibco (Eggenstein, Germany). The CXC chemokine receptor 4 (CXCR4) antagonist AMD3100 was purchased from Sigma Aldrich (Munich, Germany). To block CXCR4, cells were exposed to AMD3100 at a concentration of 25μg/ml 24 hours before the migration assay.
For FACS analysis, PE-labeled anti-human CXCR4 antibody (555974), PE-labeled anti-human β1 antibody (556049), PE-labeled anti-human α3 antibody (556025), PE-labeled anti-human α5 antibody (555617), and isotope controls corresponding to these antibodies were purchased from BD Bioscience (Heidelberg, Germany). For Western blot analysis, anti- CXCR4 antibody (clone 12G5), and anti- β-actin (clone BA3R) were purchased from Thermo Fisher Scientific (Darmstadt, Germany). Anti- Na+-K+ ATPase (clone EP1845Y) was purchased from Abcam (Cambridge, UK).
Migration assays using membranes coated with extracellular matrix proteins
For migration assays, polycarbonate membranes with 8-μm pores were coated with 0.5 μg/cm2 Collagen I (Corning, Bodenheim, Germany) and 0.5 μg/cm2 collagen IV (Corning, Bodenheim, Germany) and stored overnight at 4°C before the experiments. Next, 2 × 104 cells were loaded into the upper chamber of a 48-well modified microchemotaxis chamber (Multiwell Chemotaxis Chamber, Neuro Probe). The lower well contained cell culture medium containing 5% FBS, 10% FBS, or SDF-1α (0.1μg/ml), as indicated. An 8-μm pore size polycarbonate membrane separated the lower and upper chambers. After 5 hours of incubation at 37ºC, transmigrated cells on the lower chamber side were stained with methylene blue and counted with a Leica DC300F microscope. The number of invading cells was counted using a phase-contrast microscope. Two fields were randomly selected per well, and the number of the cells was recorded by an investigator blinded to experimental set-up.
Irradiation
Photon irradiation was performed by a biological cabinet X-ray irradiator with 320 kV and 12.50 mA (X-RAD 320 Precision X-ray Inc., N. Bradford, Conn.) at single doses of 1 and 2 Gy (110 cGy/min dose rate).
Carbon-ion irradiation was performed at the Heidelberg Ion Therapy Center using the raster scanning technique developed by Haberer et al. [44] at the horizontal beam line (Siemens AG). An extended Bragg peak (dose average LET, 103 keV/μm) was adjusted using a 30-mm acrylic shield and discharged single doses of 0.3 and 1 Gy. Cell monolayers were positioned in the middle of the extended Bragg peak.
Irradiation was performed at room temperature 24 hours before the experiments were started, as we previously reported [17]. For functional assessment of irradiated cells, cell viability was confirmed using trypan blue staining, and only viable cells were used for migration assay.
Assuming a relative biological effectiveness between 2 and 3, we chose single doses of 0.3 and 1 Gy for carbon-ion irradiations to realize the same biological effect as photon irradiation performed with single doses of 1 and 2 Gy.
FACS analysis
Twenty-four hours after irradiation, cells were fixed in 4% paraformaldehyde (PFA) in PBS and preserved at 4ºC. The fixative was removed by two washes with PBS immediately before staining. Cells were stained with a PE-labeled antibody against β1, α3, and α5 integrins; CXCR4; and matching isotope controls. H28 and H226 mesothelioma cells were analyzed with a flow cytometer (FACS Canto™ II, BD Biosciences, Heidelberg, Germany) and FlowJo software (TreeStar, Ashland, OR, USA). The results are displayed in histogram plots and assessed via subsequent quantitative analyses for the mean fluorescence intensity (MFI).
Apoptosis assay
Apoptosis of H28/H226 was determined by an Annexin V-PE apoptosis detection kit (BD Biosciences). This assay was performed 24 hours after photon or carbon irradiation, according to the manufacturer’s protocol.
Clonogenic survival assays
Cells were plated and allowed to attach for 24 hours before irradiation. After treatment, cells were grown for 14 days to allow for colony formation. Colonies were fixed with 25% acetic acid in methanol and stained with 0.5% crystal violet solution. Colonies consisting of more than 50 cells were then counted on a light microscope. All clonogenic assays were performed in triplicates.
ELISA
Concentration of CXCR4 in total cell lysate generated from H28 and H226 cell lines were determined by corresponding ELISA assays (R&D Systems, Wiesbaden, Germany), according to the manufacturers’ protocols.
Western blot analysis
Cells were grown to 80% confluency in T-75 flask and then treated with the respective irradiation. Total protein and plasma membrane fraction from cells were isolated using Total Protein Extraction Kit and Minute Plasma Membrane Protein Isolation Kit (Invent Biotechnologies, Eden Prairie, MN), respectively. Protein content of the supernatant was determined by the Bradford method with a commercial protein assay reagent (Bio-Rad, Munich, Germany). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis was performed with gels containing 10% polyacrylamide. Proteins were transferred to nitro cellulose membranes. Nitro cellulose membrane was incubated for 1h with 5% BSA in Tris-buffered saline (TBS) and probed with the CXCR4 antibody and the anti- Na+-K+ ATPase over night at 4°C in Tris-buffered saline (TBS) containing 0.1% Tween 20 and 5% BSA. All antibodies were diluted as the recommendation of manufacturer. Membranes were subsequently incubated with secondary antibody conjugated to horseradish peroxidase for 1 hour at room temperature, and immunocomplexes were visualized by enhanced chemiluminescence (Thermo Fisher Scientific). Saos-2 cell lysate (sc-2235) and Jurkat cell lysate (sc-24788), both purchased from Santa Cruz Biotechnology (Heidelberg, Germany), were included in Western blot analysis as negative and positive control for CXCR4 protein expression, respectively.
RNA silencing of CXCR4
CXCR4 silencing small inhibitory RNA oligon-eucleotides (Thermo Fisher Scientific) were administered according to the manufacturer’s protocol. CXCR4 silen-cing small inhibitory RNA represents a mixture of two sequences, 5′-GGCAGUCCAUGUCAUCUAC-3′, and 5′-GUAGAUGACAUGGACUGCC- 3′. Fifty picomol per milliliter of each inhibitory siRNA was administered 24 hours before irradiation.
Statistical analysis
All experiments were performed at least three times in triplicate in each individual experimental set-up. Comparisons between two groups were performed with Student’s t-test (SPSS 23.0; SPSS Inc., Chicago, IL, USA). p-Values less than 0.05 were considered to be statistically significant.
CONFLICTS OF INTEREST
No conflict declared.
GRANT SUPPORT
This work was supported by Uehara foundation fellowship for research abroad and by institutional research funding.
REFERENCES
1. Singhal S, Kaiser LR. Malignant mesothelioma: options for management. Surg Clin North Am. 2002; 82: 797–831.
2. Treasure T, Utley M. Ten traps for the unwary in surgical series: a case study in mesothelioma reports. J Thorac Cardiovasc Surg. 2007; 133: 1414–8.
3. Rea F, Marulli G, Bortolotti L, Breda C, Favaretto AG, Loreggian L, Sartori F. Induction chemotherapy, extrapleural pneumonectomy (EPP) and adjuvant hemi-thoracic radiation in malignant pleural mesothelioma (MPM): feasibility and results. Lung Cancer. 2007; 57: 89–95.
4. Flores RM, Krug LM, Rosenzweig KE, Venkatraman E, Vincent A, Heelan R, Akhurst T, Rusch VW. Induction chemotherapy, extrapleural pneumonectomy, and postoperative high-dose radiotherapy for locally advanced malignant pleural mesothelioma: a phase II trial. J Thorac Oncol. 2006; 1: 289–95.
5. Krug LM, Pass HI, Rusch VW, Kindler HL, Sugarbaker DJ, Rosenzweig KE, Flores R, Friedberg JS, Pisters K, Monberg M, Obasaju CK, Vogelzang NJ. Multicenter phase II trial of neoadjuvant pemetrexed plus cisplatin followed by extrapleural pneumonectomy and radiation for malignant pleural mesothelioma. J Clin Oncol. 2009; 27: 3007–13.
6. Thieke C, Nicolay NH, Sterzing F, Hoffmann H, Roeder F, Safi S, Debus J, Huber PE. Long-term results in malignant pleural mesothelioma treated with neoadjuvant chemotherapy, extrapleural pneumonectomy and intensity-modulated radiotherapy. Radiat Oncol. 2015; 10: 267.
7. Stahel RA, Riesterer O, Xyrafas A, Opitz I, Beyeler M, Ochsenbein A, Früh M, Cathomas R, Nackaerts K, Peters S, Mamot C, Zippelius A, Mordasini C, et al. Neoadjuvant chemotherapy and extrapleural pneumonectomy of malignant pleural mesothelioma with or without hemithoracic radiotherapy (SAKK 17/04): a randomised, international, multicentre phase 2 trial. Lancet Oncol. 2015; 16: 1651–8.
8. Kostron A, Friess M, Crameri O, Inci I, Schneiter D, Hillinger S, Stahel R, Weder W, Opitz I. Relapse pattern and second-line treatment following multimodality treatment for malignant pleural mesothelioma. Eur J Cardiothorac Surg. 2016; 49: 1516–23.
9. Gomez DR, Hong DS, Allen PK, Welsh JS, Mehran RJ, Tsao AS, Liao Z, Bilton SD, Komaki R, Rice DC. Patterns of failure, toxicity, and survival after extrapleural pneumonectomy and hemithoracic intensity-modulated radiation therapy for malignant pleural mesothelioma. J Thorac Oncol. 2013; 8: 238–45.
10. Minatel E, Trovo M, Bearz A, Di Maso M, Baresic T, Drigo A, Barresi L, Furlan C, Del Conte A, Bruschi G, Fontana P, Pagan V, Franchin G. Radical radiation therapy after lung-sparing surgery for malignant pleural mesothelioma: survival, pattern of failure, and prognostic factors. Int J Radiat Oncol Biol Phys. 2015; 93: 606–13.
11. Krayenbuehl J, Dimmerling P, Ciernik IF, Riesterer O. Clinical outcome of postoperative highly conformal versus 3D conformal radiotherapy in patients with malignant pleural mesothelioma. Radiat Oncol. 2014; 9: 32.
12. Ishihara S, Haga H, Yasuda M, Mizutani T, Kawabata K, Shirato H, Nishioka T. Integrin beta1-dependent invasive migration of irradiation-tolerant human lung adenocarcinoma cells in 3D collagen matrix. Biochem Biophys Res Commun. 2010; 396: 651–5.
13. Jung JW, Hwang SY, Hwang JS, Oh ES, Park S, Han IO. Ionising radiation induces changes associated with epithelial-mesenchymal transdifferentiation and increased cell motility of A549 lung epithelial cells. Eur J Cancer. 2007; 43: 1214–24.
14. Gu Q, He Y, Ji J, Yao Y, Shen W, Luo J, Zhu W, Cao H, Geng Y, Xu J, Zhang S, Cao J, Ding WQ. Hypoxia-inducible factor 1α (HIF-1α) and reactive oxygen species (ROS) mediates radiation-induced invasiveness through the SDF-1α/CXCR4 pathway in non-small cell lung carcinoma cells. Oncotarget. 2015; 6: 10893–907. doi: 10.18632/oncotarget.3535.
15. Agemy L, Harmelin A, Waks T, Leibovitch I, Rabin T, Pfeffer MR, Eshhar Z. Irradiation enhances the metastatic potential of prostatic small cell carcinoma xenografts. Prostate. 2008; 68: 530–9.
16. Kaliski A, Maggiorella L, Cengel KA, Mathe D, Rouffiac V, Opolon P, Lassau N, Bourhis J, Deutsch E. Angiogenesis and tumor growth inhibition by a matrix metalloproteinase inhibitor targeting radiation-induced invasion. Mol Cancer Ther. 2005; 4: 1717–28.
17. Rieken S, Habermehl D, Mohr A, Wuerth L, Lindel K, Weber K, Debus J, Combs SE. Targeting ανβ3 and ανβ5 inhibits photon-induced hypermigration of malignant glioma cells. Radiat Oncol. 2011; 6: 132.
18. Ogata T, Teshima T, Kagawa K, Hishikawa Y, Takahashi Y, Kawaguchi A, Suzumoto Y, Nojima K, Furusawa Y, Matsuura N. Particle irradiation suppresses metastatic potential of cancer cells. Cancer Res. 2005; 65: 113–20.
19. Takuwa T, Hasegawa S. Current surgical strategies for malignant pleural mesothelioma. Surg Today. 2016; 46: 887–94.
20. Rusch V, Baldini EH, Bueno R, De Perrot M, Flores R, Hasegawa S, Klepetko W, Krug L, Lang-Lazdunski L, Pass H, Weder W, Sugarbaker DJ; Participants in 2012 International Mesothelioma Interest Group Congress. The role of surgical cytoreduction in the treatment of malignant pleural mesothelioma: meeting summary of the International Mesothelioma Interest Group Congress, September 11-14, 2012, Boston, Mass. J Thorac Cardiovasc Surg. 2013; 145: 909–10.
21. Kajiyama H, Shibata K, Terauchi M, Ino K, Nawa A, Kikkawa F. Involvement of SDF-1α/CXCR4 axis in the enhanced peritoneal metastasis of epithelial ovarian carcinoma. Int J Cancer. 2008; 122: 91–9.
22. Yasumoto K, Koizumi K, Kawashima A, Saitoh Y, Arita Y, Shinohara K, Minami T, Nakayama T, Sakurai H, Takahashi Y, Yoshie O, Saiki I. Role of the CXCL12/CXCR4 axis in peritoneal carcinomatosis of gastric cancer. Cancer Res. 2006; 66: 2181–7.
23. Phillips RJ, Burdick MD, Lutz M, Belperio JA, Keane MP, Strieter RM. The stromal derived factor-1/CXCL12-CXC chemokine receptor 4 biological axis in non-small cell lung cancer metastases. Am J Respir Crit Care Med. 2003; 167: 1676–86.
24. Gong HY, Hu WG, Hu QY, Li XP, Song QB. Radiation-induced pulmonary injury accelerated pulmonary metastasis in a mouse model of breast cancer. Oncol Lett. 2015; 10: 3613–8.
25. Lau BW, Kane AB. SDF1/CXCL12 is involved in recruitment of stem-like progenitor cells to orthotopic murine malignant mesothelioma spheroids. Anticancer Res. 2010; 30: 2153–60.
26. Li T, Li H, Wang Y, Harvard C, Tan JL, Au A, Xu Z, Jablons DM, You L. The expression of CXCR4, CXCL12 and CXCR7 in malignant pleural mesothelioma. J Pathol. 2011; 223: 519–30.
27. Rieken S, Habermehl D, Wuerth L, Brons S, Mohr A, Lindel K, Weber K, Haberer T, Debus J, Combs SE. Carbon ion irradiation inhibits glioma cell migration through downregulation of integrin expression. Int J Radiat Oncol Biol Phys. 2012; 83: 394–9.
28. Nalla AK, Asuthkar S, Bhoopathi P, Gujrati M, Dinh DH, Rao JS. Suppression of uPAR retards radiation-induced invasion and migration mediated by integrin β1/FAK signaling in medulloblastoma. PLoS One. 2010; 5: e13006.
29. Tsutsumi K, Tsuda M, Yazawa N, Nakamura H, Ishihara S, Haga H, Yasuda M, Yamazaki R, Shirato H, Kawaguchi H, Nishioka T, Ohba Y. Increased motility and invasiveness in tumor cells that survive 10 Gy irradiation. Cell Struct Funct. 2009; 34: 89–96.
30. Abdollahi A, Griggs DW, Zieher H, Roth A, Lipson KE, Saffrich R, Gröne HJ, Hallahan DE, Reisfeld RA, Debus J, Niethammer AG, Huber PE. Inhibition of αvβ3 integrin survival signaling enhances antiangiogenic and antitumor effects of radiotherapy. Clin Cancer Res. 2005; 11: 6270–9.
31. Klominek J, Sumitran Karuppan S, Hauzenberger D. Differential motile response of human malignant mesothelioma cells to fibronectin, laminin and collagen type IV: the role of beta1 integrins. Int J Cancer. 1997; 72: 1034–44.
32. Klominek J, Baskin B, Hauzenberger D. Platelet-derived growth factor (PDGF) BB acts as a chemoattractant for human malignant mesothelioma cells via PDGF receptor beta-integrin alpha3beta1 interaction. Clin Exp Metastasis. 1998; 16: 529–39.
33. Barth TF, Brüderlein S, Rinaldi N, Mechtersheimer G, Möller P. Pleural mesothelioma mimics the integrin profile of activated, sessile rather than detached mesothelial cells. Int J cancer. 1997; 72: 77–86.
34. Okada T, Kamada T, Tsuji H, Mizoe J, Baba M, Kato S, Yamada S, Sugahara S, Yasuda S, Yamamoto N, Imai R, Hasegawa A, Imada H, et al. Carbon ion radiotherapy: clinical experiences at National Institute of Radiological Science (NIRS). J Radiat Res. 2010; 51: 355–64.
35. Fokas E, Kraft G, An H, Engenhart-Cabillic R. Ion beam radiobiology and cancer: time to update ourselves. Biochim Biophys Acta. 2009; 1796: 216–29.
36. Schulz-Ertner D, Tsujii H. Particle radiation therapy using proton and heavier ion beams. J Clin Oncol. 2007; 25: 953–64.
37. Hamada N, Imaoka T, Masunaga S, Ogata T, Okayasu R, Takahashi A, Kato TA, Kobayashi Y, Ohnishi T, Ono K, Shimada Y, Teshima T. Recent advances in the biology of heavy-ion cancer therapy. J Radiat Res. 2010; 51: 365–83.
38. Ogata T, Teshima T, Inaoka M, Minami K, Tsuchiya T, Isono M, Furusawa Y, Matsuura N. Carbon ion irradiation suppresses metastatic potential of human non-small cell lung cancer A549 cells through the phosphatidylinositol-3-kinase/Akt signaling pathway. J Radiat Res. 2011; 52: 374–9.
39. Akino Y, Teshima T, Kihara A, Kodera-Suzumoto Y, Inaoka M, Higashiyama S, Furusawa Y, Matsuura N. Carbon-ion beam irradiation effectively suppresses migration and invasion of human non-small-cell lung cancer cells. Int J Radiat Oncol Biol Phys. 2009; 75: 475–81.
40. Fujita M, Otsuka Y, Imadome K, Endo S, Yamada S, Imai T. Carbon-ion radiation enhances migration ability and invasiveness of the pancreatic cancer cell, PANC-1, in vitro. Cancer Sci. 2012; 103: 677–83.
41. Goetze K, Scholz M, Taucher-Scholz G, Mueller-Klieser W. The impact of conventional and heavy ion irradiation on tumor cell migration in vitro. Int J Radiat Biol. 2007; 83: 889–96.
42. Takahashi Y, Teshima T, Kawaguchi N, Hamada Y, Mori S, Madachi A, Ikeda S, Mizuno H, Ogata T, Nojima K, Furusawa Y, Matsuura N. Heavy ion irradiation inhibits in vitro angiogenesis even at sublethal dose. Cancer Res. 2003; 63: 4253–7.
43. Asakura K, Izumi Y, Yamamoto M, Yamauchi Y, Kawai K, Serizawa A, Mizushima T, Ohmura M, Kawamura M, Wakui M, Adachi T, Nakamura M, Suematsu M, et al. The cytostatic effects of lovastatin on ACC-MESO-1 cells. J Surg Res. 2011; 170: e197–209.
44. Haberer T, Becher W, Schardt D, Kraft G. Magnetic scanning system for heavy ion therapy. Nucl Instruments Methods Phys Res Sect A Accel Spectrometers Detect Assoc Equip. 1993; 330: 296–305.