
INTRODUCTION
Outbreaks of catastrophic swine diseases have drawn extensive attention to the considerable economic and social losses that affect a wide range of swine farms locally and the industry worldwide. Current vaccination strategies and antiviral drugs cannot effectively control swine diseases, such as porcine reproductive and respiratory syndrome (PRRS), which is mainly caused by PRRS virus (PRRSV), based on PRRSV itself is easy to mutate with time, PRRSV has the diversity of the genotypes. The complex genetic diversity brings a challenge to increase the efficacy of current PRRS vaccines [1, 2]. The innate immune response is the first line of host defense against infections. Type I interferons (IFNs) (primarily IFN-α/β) are induced to boost the immune response, protecting hosts from viral and nonviral pathogens [3]. IFNs can inhibit a variety of viruses, bacteria, parasites, and bacterial and viral co-infections. Therefore, the role of IFNs has been focused on in the prevention and treatment of swine diseases in recent years. Initiation of IFN genes transcription depends on the timely recognition of pathogen-associated molecular patterns through host pattern recognition receptors within endosomes (such as Toll-like receptors, TLRs) or in the cytosol (including cytosolic RNA sensors, RNA helicases, and cytosolic DNA sensors) [4]. Recognition triggers distinct signaling pathways that converge on the activation of transcription factors called IFN regulatory factors (IRFs) [3], which include nine members (IRF-1 to 9). Among the IRF family, IRF1, IRF3, and IRF7 are characterized as acting early in regulating type I IFNs expression [3]. Moreover, IRF3 and IRF7 are two important members of IRFs family, having high homology. They play an important regulatory role in the expression and secretion of type I IFN for antiviral functions [5].
Moreover, IRF3 and IRF7 are primarily responsible for TLR4-IFN signaling through the TIR domain-containing adaptor protein pathway [6]. During Lactobacillus acidophilus infection, IRF1, IRF3, and IRF7 are involved in IFN-β responses in dendritic cells triggered by myeloid differentiation factors (MyD88) [7]. These results suggest that multiple IRF members often work together to coordinately regulate type I IFN gene activation, although the molecular mechanisms involved remain unclear. IRF1, IRF3, and IRF7 might simultaneously regulate IFN gene expression in certain tissues or cell types through similar signaling pathways, such as the MyD88 pathway [8]. Notably, IRF3 and IRF7 are necessary for the retinoic acid-inducible gene I-like receptors mediated IFN response [9], as well as for the MyD88-mediated IFN response [10]. Moreover, IFN suppression by viruses may be achieved through blocking IRF3/IRF7 activity [11]. IRF3/IRF7 undergo phosphorylation and enter the nucleus through nuclear localization signals, increasing IFN activity [12].
IRF3/IRF7 has been shown to regulate cell proliferation, apoptosis, inflammation, innate immune responses and insulin resistance [13–15]. Both can be activated by LPS, which exists mainly in the cell wall of Gram-negative bacteria. LPS binds to its receptor TLR4, located on the cell surface, activating TLR4-mediated signal transduction pathways, then activating downstream TRIF-dependent signal pathway, leading to IRF3/IRF7 phosphorylation, which regulates expression and secretion of type I IFNs.
Currently, studies on swine IRF3 and IRF7, especially regarding their function, are very few. To better understand IRF3/IRF7 function in preventing and resolving swine diseases, we constructed the eukaryotic expression vectors PB-IRF3 and PB-IRF7, and transfected them into PK15 cells to obtain IRF3/IRF7 overexpression. Meanwhile, we explored the effects of swine IRF3/IRF7 on TLR4 signaling pathway, using LPS alone, or in combination with a TBK1 inhibitor (LiCl). We found that overexpress IRF3/IRF7 in PK15 cells could activate TLR4 signal pathway through regulating MyD88, TBK1, NFκB and TRAF6 expression. Moreover, it promoted IFNα and inhibited IL-6 expression, further suppressing the inflammatory response. These results demonstrated that overexpression of swine IRF3/IRF7 could decrease the inflammatory response through TLR4 signaling pathway, and participate in type I interferons production.
RESULTS
Observation of transfection efficiency
The PB vector carries the GFP gene. When the PB vector is integrated into the host cell genome, GFP protein is expressed in the host cells, which can be seen using a fluorescence microscope. We clearly observed green fluorescence in the PBR3, PBF7, and PBv transfected, but not in the non-transfected, PK15 cells [Figure 1]. These results indicated that vectors with the GFP gene were successfully transfected into cells and integrated into the genome. Through screening, we obtained stable transfected cell lines.
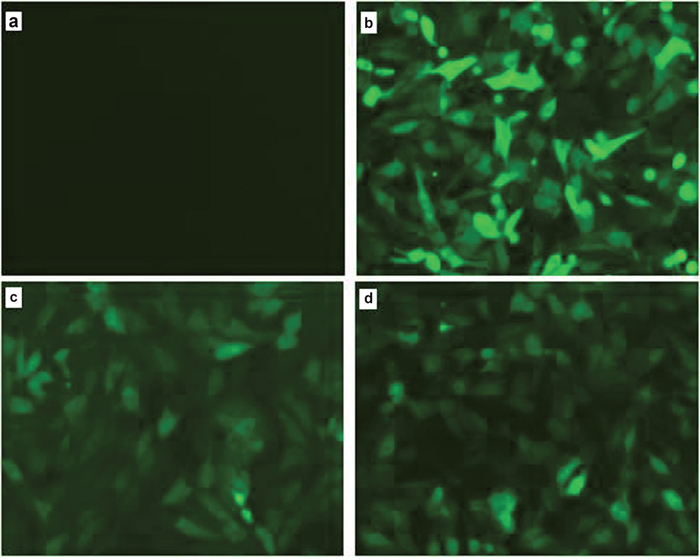
Figure 1: Observation of green fluorescence in PK15 cells transfected with IRF3/IRF7 expression vectors. Green fluorescence was observed clearly using fluorescence microscopy in the PBv (b), PBR3 (c) and PBF7 (d) cells, but not in non-transfected PK15 cells (a).
Overexpression of swine IRF3/IRF7 genes in PK15 cells
We constructed the eukaryotic expression vectors PB-IRF3 and PB-IRF7, and transfected them into PK15 cells. To evaluate IRF expression efficiency, we detected the expression of IRF3/IRF7 using real-time RT-PCR. IRF3 and IRF7 mRNA expression were significantly higher in PBR3 and PBR7 cells than in PBv cells (P< 0.001) [Figure 2a, 2b]. Thus, IRF3/IRF7 was successfully transfected into PK15 cells and efficiently overexpressed.
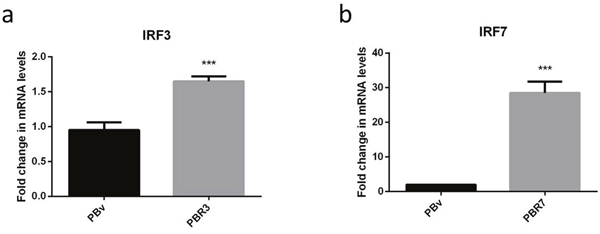
Figure 2: Overexpression of the swine IRF3/IRF7 gene in PK15 cells. (a and b) demonstrate the overexpression of IRF3 and IRF7 mRNA in PBR3 and PBv and PBR7 and PBv cells, respectively. GAPDH was used as an internal housekeeping gene *** P < 0.001 vs. PBv.
Swine IRF3/IRF7 genes induce an anti-inflammatory response
IRF3/IRF7 plays an important role in type I IFN production. Both exist in multiple types of cells, mainly in the cytoplasm in an inactive form. If cells are stimulated, IRF3/IRF7 are phosphorylated and change conformation, translocating to the nucleus. First, we explored the effect of swine IRF3/IRF7 overexpression on IFNα and IL-6 production. As shown in Figure 3a and Figure 4a, compared with control cells, LPS increased IFNα mRNA abundance in PBR3/PBR7 and PBv cells. Pretreatment with LiCl followed by LPS, significantly downregulated IFNα mRNA expression in both PBR3/PBR7 and PBv cells (P< 0.001). Notably, in all treatments, IFNα mRNA expression in PBR3/PBR7 was higher than that in PBv. These results suggested that IRF3/IRF7 can enhance IFNα mRNA expression, especially during inflammatory reactions, and illustrated that IRF3/IRF7 exerted an important role in promoting type I interferons production.
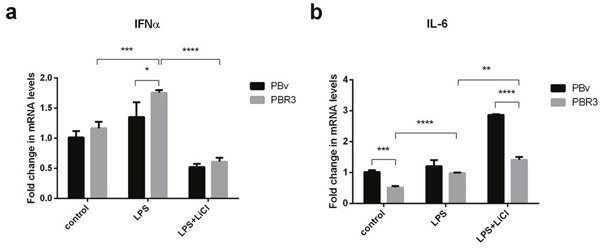
Figure 3: Effect of swine IRF3 on the expression of IFNα and IL-6. The expression of IFNα (a) and IL-6 (b) in PBR3 and PBv cells induced by LPS (50 μg/ml) with or without LiCl (20 mM) pretreatment. * P < 0.05, ** P < 0.01, *** P < 0.001 vs. PBv or between conditions.
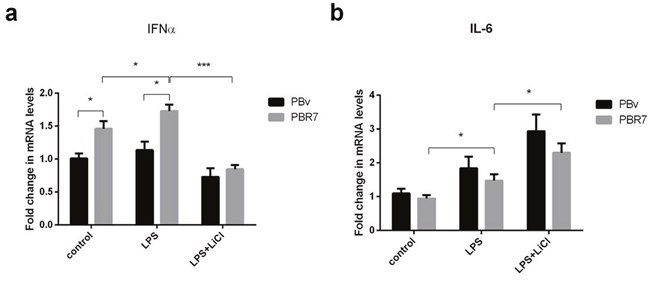
Figure 4: Effect of swine IRF7 on the expression of IFNα and IL-6. The expression of IFNα (a) and IL-6 (b) in PBR7 and PBv cells induced by LPS (50 μg/ml) with or without LiCl (20 mM) pretreatment. * P < 0.05, *** P < 0.001 vs. PBv or between conditions.
The changes in IL-6 mRNA were different from IFNα mRNA, as shown in Figure 3b and Figure 4b. Compared with the control group, LPS increased IL-6 mRNA abundance in PBR3/PBR7 and PBv cells. However, after blocking the TBK1 pathway, LPS still increased IL-6 mRNA expression both in PBR3/PBR7 and PBv cells (P< 0.001). In all treatments, IL-6 mRNA expression in PBR3/PBR7 was lower than that in PBv. These results suggested that IRF3/IRF7 can inhibit IL-6 mRNA expression, especially during inflammatory reactions, independent of the TBK1 pathway. These results suggest that IRF3/IRF7 can attenuate the inflammatory response.
Swine IRF3/IRF7 actives the TLR4 signaling pathway
LPS activates multiple TLR4-mediated signal transduction pathways, including a downstream MyD88-independent pathway, leading to IRF3 and IRF7 phosphorylation, which regulates expression and secretion of type I IFNs. Since IRF3 and IRF7 regulate IFNα and IL-6 production, we investigated whether IRF3/IRF7 also have effects on upstream molecules in the TLR4-mediated signaling pathway using real-time PCR. As shown in Figure 5 and Figure 6, we found that changes in MyD88, TRAF6, TBK1 and NFκB mRNA were similar in all treatments. Compared with the control group, LPS increased MyD88, TRAF6, TBK1 and NFκB mRNA levels in PBR3/PBR7 and PBv cells. Pretreatment with LiCl prior to LPS stimulation, downregulated MyD88, TRAF6, TBK1 and NFκB mRNA expression both in PBR3/PBR7 and PBv cells. In all treatments, MyD88, TRAF6, TBK1 and NFκB mRNA expression in PBR3/PBR7 were higher than those in PBv. Our results suggested that swine IRF3/IRF7 could activate TLR4 signaling pathway.
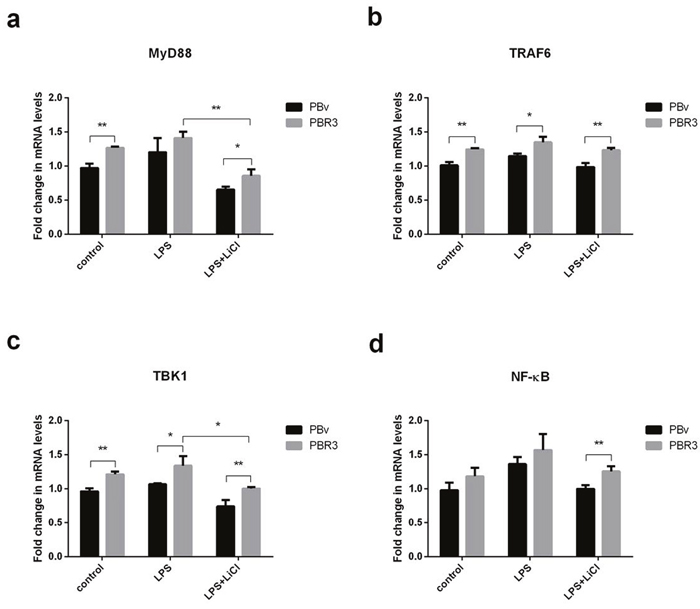
Figure 5: Effect of swine IRF3 overexpression on TLR4 signaling pathway components. Graphs demonstrate the expression of MyD88 (a), TRAF6 (b), TBK1 (c) and NFκB (d) in PBR3 and PBv cells stimulated with LPS (50 μg/ml) with or without LiCl (20 mM) pretreatment. * P < 0.05, ** P < 0.01 vs. PBv or between treatments.
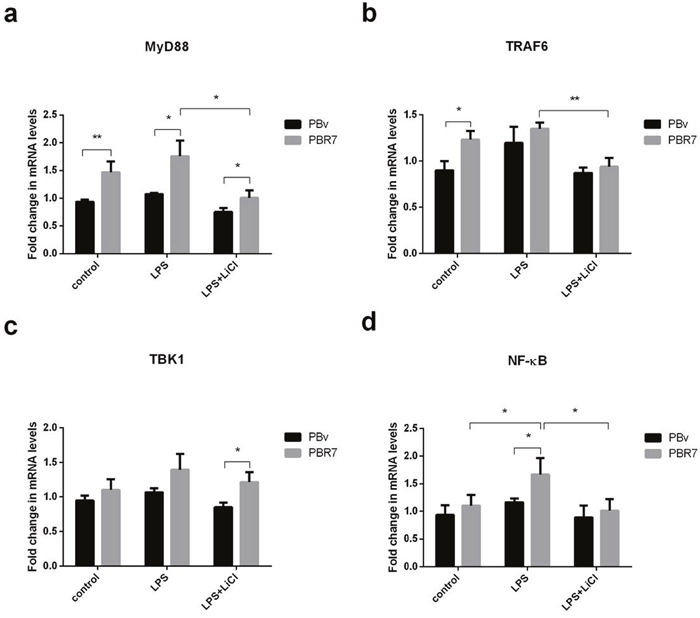
Figure 6: Effect of swine IRF7 on the expression of components of the TLR4 signaling pathway. Graphs demonstrate the expression of MyD88 (a), TRAF6 (b), TBK1 (c) and NFκB (d) in PBR7 and PBv cells stimulated with LPS (50 μg/ml) with or without LiCl (20 mM) pretreatment. * P < 0.05, ** P < 0.01 vs. PBv or between conditions.
DISCUSSION
Transposable elements play an important role through “cut-and-paste” of target DNA during genome editing, leading to product insertional mutagenesis or transgenesis [16, 17]. Piggybac (PB) is a DNA transposon which was originally isolated from the genomes of baculoviruses that infect cabbage looper moth Trichoplusianini [18]. PB inserts TTAA sequences at target sites, displaying little selectivity for specific position of the genome and a modest superiority for the sensitivity of DNase I region [19, 20]. A small modification of PB would be effective in helping integration into safe sites. Previous studies have found that PB fused to a DNA binding domain remained active, which can distinguish insertion from innate sites [21, 22]. In this experiment, PB transposons as transgene vectors were successfully constructed using the eukaryotic expression vector PB-IRF3 and PB-IRF7, and then the PBase and PB vector plasmid with target gene were co-transfected into PK15 cells. After puromycin-selection, we eliminated non-transfected cells. All cells were clearly observed using fluorescence microscopy after transfection, suggesting that PB-IRF3/IRF7 and the PB-vector were effectively transfected into PK15 cells. From the results of transfection in this study, we found that the efficiency of integration of exogenous genes was affected by the size of DNA fragments, it may be that shorter fragmented genes are more easily integrated into the host. Real-time PCR results showed that IRF3 and IRF7 mRNA expression were obviously higher in PBR3/PBR7 than those in PBv (P< 0.001).
IFNs are essential for both innate and adaptive immunity, they play an important role in initially recognizing virus infection and evoking antiviral responses by the immune system. IFN combines with their receptors on the surface of cells, subsequently initiating signaling cascades through Janus kinase-induced signal activators and transducers. The IFN signaling pathway exerts major roles in regulating the expression of various IFN-stimulated genes, which are mainly associated with innate immunity, and thus determine the immune state of the body. Different ligands can bind to TLRs that activate the transcription factors NFκB, IRF3 and IRF7, which are all required for initiating the production of IFNα/β [23]. IRF3/IRF7 is inactive form in the cytoplasm, best known as an important regulator of type I IFN response [24, 25]. When cells are stimulated, IRF3/IRF7 are phosphorylated and change conformation, translocating to the nucleus.
LPS is a principal component of the cell wall in Gram-negative bacteria [26]. LPS recognition and signal transduction are key events in the host immune reaction, and are related to many inflammatory diseases. LPS can activate multiple signal transduction pathways. In the presence of LPS-binding protein (LBP) and CD14, LPS binds with TLR4, recruiting adaptor protein MyD88 and interleukin-1 (IL-1) receptor-associated kinase, followed phosphorylated and activated tumor necrosis factor receptor-associated factor-6 (TRAF6). The activated TRAF6 enhances the activity of the transcription factor NFκB, leading to activation of MAP kinase pathways. Finally, cells produce and release many pro-inflammatory cytokines, such as tumor necrosis factor α (TNFα), IL-6 and IL-8 [27–30]. Lithium salt is a widely used glycogen synthase kinase-3β (GSK3β) inhibitor and an effective drug for treating inflammatory diseases [31, 32]. Furthermore, previous studies have demonstrated that GSK3β is involved in innate immune reactions [33]. For example, GSK3β has been documented as a major regulator maintaining the balance of pro- and anti-inflammatory actions associated with TLR stimulation [34]. Wang et al. demonstrated that lithium decreases IFN-β levels and attenuates antivirus role of host through targeting TBK1 [35].
IRF3 and IRF7 are central players of the signaling cascade leading to IFN type I induction [36]. The activated IRF3 bind to the promoters of IFN type I upregulate IFN-α/β transcription, associating with transcriptional coactivators [37, 38]. Rocca and coworkers showed that the virus induced the inhibition of transcription of the IRF3 gene, decreasing IRF3 translocation to the nucleus furthor inhibits IFN- α/β production [39]. However, Bauthofer et al demonstrated that IRF3 degradation via a proteasome-dependent mechanism, rather than inhibiting the phosphorylation and translocation of IRF3 in classical swine fever virus infected. IRF7 expression, however, is not affected.
In this paper, we explored the effect of overexpression of swine IRF3/IRF7 on IFNα and IL-6 production. Results showed (Figure 3a and Figure 4a) that LPS increased IFNα mRNA abundance with and without overexpression IRF3/7 cells. LiCl significantly downregulated IFNα mRNA expression in both PBR3/PBR7 and PBv (P< 0.001). In all treatments, overexpression IRF3/7 increased IFNα mRNA expression. These results suggested that IRF3/IRF7 can enhance IFNα mRNA expression, especially during inflammatory reactions, at the same time also illustrated that IRF3/IRF7 exerted an important role in activating type I interferon response.
IRF3/7 promotes type I IFN transcription in the nucleus [6, 40]. IFNs are secreted binding to specific receptors as an endocrine or autocrine manner, triggering the activation of hundreds of IRFs that participate in antiviral pathways [40, 41]. Recently, Ramírez-Carvajal has demonstrated that a constitutively active fusion protein of porcine IRF3 and IRF7 completely protects swine against foot and mouth disease by inducing a strong type I IFN response [42, 43].
The changes in IL-6 mRNA were different from IFNα mRNA, as shown in Figure 3b and Figure 4b. LPS increased IL-6 mRNA abundance with and without overexpression IRF3/7 cells. After blocking the TBK1 pathway, LPS still increased IL-6 mRNA expression both in PBR3/PBR7 and PBv cells (P<0.001). In all treatments, overexpression IRF3/7 decreased IL-6 mRNA. These results suggested that IRF3/IRF7 can inhibit IL-6 mRNA expression, especially during inflammatory reactions independent of the TBK1 pathway. These data suggested that swine IRF3/IRF7 attenuate inflammatory responses. However, Li et al found that IRF7 overexpressing in human microglia increased IL-6 secretion in tissues and serums, promoting signal transducers and activators of transcription 3 (stat3)-IL-6 signaling activation [44]. The conflict conclusion may result from different cell and microenvironment, as well as different detection method.
LPS activates multiple TLR4-mediated signal transduction pathways, activating the downstream MyD88-independent signaling pathway, leading to IRF3/IRF7 phosphorylation, and regulation of the expression and secretion of type I IFNs. Components of swine viruses inhibit IRF3-complex formation in the nucleus [45], which prevents binding to the IFN-β promoter regions and inhibits the assembly of RNA polymerase II of IFN gene expression. In addition, miR-23 induced type I IFN expression through activation of IRF3/IRF7, whichmight further inhibit virus infection [46]. Since IRF3/IRF7 can regulate IFNα and IL-6 production, we investigated whether IRF3/IRF7 has effects on upstream molecules of the TLR4-mediated signaling pathway. As shown in Figure 5 and Figure 6, we found that MyD88, TRAF6, TBK1 and NFκB mRNA variation trends were similar in all treatments. LPS increased MyD88, TRAF6, TBK1 and NFκB mRNA abundance in PBR3/PBR7 and PBv cells. Pretreatment with LiCl significantly downregulated LPS-induced MyD88, TRAF6, TBK1 and NFκB mRNA expression both in PBR3/PBR7 and PBv cells. In all treatments, MyD88, TRAF6, TBK1 and NFκB mRNA expression in PBR3/PBR7 were higher than those in PBv cells. Our results suggested that swine IRF3/IRF7 could activate TLR4 signaling pathway.
MATERIALS AND METHODS
Reagents
LPS (E. coli O55:B5), puromycin and LiCl were purchased from Sigma (St Louis, MO, USA). Dulbecco’s modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were obtained from Gibco (Life Technologies, Carlsbad, CA, USA). Trizol reagent, Prime Script RT reagent kit, SYBR Premix Ex Taq, and pMD19-T were purchased from TaKaRa Bio Inc. (Shiga, Japan). PB vector and PB transposes were obtained from SBI Ltd. (Palo Alto, CA, US). Other reagents were purchased from Sino Pharm Chemical Reagent Ltd. (Shanghai, China).
Plasmids
Construction of the eukaryotic expression vectors PB-IRF3 and PB-IRF7 were as follows. Total RNA was obtained from swine tissues using Trizol reagent, and then cDNA was synthetized using the extracted RNA. IRF3 and IRF7 fragments were amplified and the PCR products were purified using a gel extraction kit. The purified products were ligated into pMD19-T plasmids, and used to transform E. coli DH5a cells. Recombinant plasmids were extracted from bacterial colonies, and the presence of IRF3 or IRF7 sequence identified in plasmids by agarose gel electrophoresis. The length of IRF3 and IRF7 sequences were 1260 bp and 1470 bp, respectively. IRF3 and IRF7 cDNA was subcloned from pMD19-T-IRF3 or pMD19-T-IRF7 into the PB vector (that contained green fluorescent protein (GFP) and puromycin resistance genes) after double restriction enzyme digestion. The obtained recombinant expression vectors were called PB-IRF3 and PB-IRF7. Nhe I and EcoR I were used to digest PB-IRF3 or PB-IRF7, and then identified them through agarose gel electrophoresis. Sequencing using both forward and reverse primers was performed to confirm the above recombinant vectors.
Cell culture, transfection, and photograph
PK15 porcine kidney epithelial cells were maintained in DMEM medium including 10% FBS (v/v) and 1% penicillin/streptomycin (v/v), and cultured at 37°C with 5% CO2 (MCO-5AC CO2 Incubator, Sanyo, Tokyo, Japan). Prior to transfection experiments, cells were plated at a density of 2×105 cells per well in six-well plates. The PK15 cells were transfected with PB-IRF3 or PB-IRF7 constructs in parallel with empty vectors using lipofectamine 2000 reagent (Life Technologies) complying with the manufacturer’s instructions. Based on the transfecting plasmids, cells were divided into four groups: PB-IRF3, PB-IRF7, PB-vector and non-transfection. Twelve h after transfection, photomicropgraphs were taken through a DAS microscope (Leitz DM RB) with a charged coupled device camera (C4880; Hamamatsu, Japan) to detect GFP expression. The transfected cells were selected for on 5 μg/mL puromycin for 2 weeks. Cells that were not transfected with the target gene were killed, and we achieved stable transfection cell lines, named PK15-PBIRF3 (PBR3), PK15-PBIRF7 (PBR7), and PK15-PBvector (PBv).
Transfection efficiency and overexpression detection
The PB vector carries both the purine gene and the GFP gene. Therefore, when the PB vector was transfected into cells and integrated into the genome, GFP protein was expressed in the host cells. After 12 h of transfection, the cells were observed under the fluorescence microscope to evaluate transfection efficiency. Overexpression of target genes (IRF3, IRF7) were detected using RT-PCR.
Cell treatment
The stably transfected cell lines, PBR3, PBR7, and PBv, were separated into 3 groups: control, LPS, and LPS+LiCl. Cells were exposed to LPS (50 μg/ml) for 6 h in the LPS group. Cells in the LPS + LiCl group were treated with 20 mMLiCl (an inhibitor of TBK1, which inhibits IRF3/7 activity, and interferes with IFN secretion) for 6 h before the addition of LPS (50 μg/ml) for 6 h. Cells in the control group were cultured in basal medium. After treatment, cells were collected to carry out TLR4 signaling pathway and inflammatory markers analysis using real-time qPCR.
cDNA synthesis and PCR
Trizol reagent was used to extract total RNA from cultured cells. A reverse transcriptase kit was used to carry out cDNA synthesis. PCR analysis was performed with a SYBR Green PCR Kit using a real-time fluorescence quantitative PCR instrument (Eppendorf Mastercycle reprealplex, Hamburg, Germany), with a reaction volume of 20 μL. The PCR program was as follows: 95°C 20 s, 60°C 30 s, and 72°C 20 s, for 40 cycles. Swine mRNA specific primers are listed in Table 1. The synthesis of all primers was carried out by Shanghai Sangon Co. Ltd. (Shanghai, China). Each sample was analyzed in triplicate, and RT-PCR results were analyzed and evaluated using the relative quantity Ct method. Melting curve analysis was used to confirm the specificity of primers. The expression of the target gene was normalized as the ratio of target gene/GAPDH mRNA.
Table 1: Primers used for RT-PCR amplification
Gene |
Primers |
Accession No. |
|---|---|---|
IRF3 |
F: 5’-TCATCGAAGATCTGATTGCCTTC-3’ |
NM_213770.1 |
NFκB |
F: 5’-CCCATGTAGACAGCACCACCTATGAT-3’ |
NM_001048232.1 |
IRF7 |
F: 5’-CTGCGATGGCTGGATGAA-3’ |
NM_001097428.1 |
MyD88 |
F: 5’-GCTGTAGGGGGAATGTGTGT-3’ |
XM_013992286.1 |
TRAF6 |
F: 5’-GGGAACGATACGCCTTACAA-3’ |
NM_001105286.1 |
IL-6 |
F: 5’-CTGGCAGAAAACAACCTGAACC-3’ |
NM_001252429.1 |
TBK1 |
F: 5’-ACAGATTTTGGTGCAGCCAG-3’ |
NM_001105292.1 |
IFNα |
F: 5’-GGTGCATGAGATGCTCCA-3’ |
XM_013993516.1 |
TLR4 |
F: 5’-CCCTGACAACATCCCCACAT-3’ |
XM_013986843.1 |
GAPDH |
F: 5’-GGAGAACGGGAAGCTTGTCA-3’ |
NM_001206359.1 |
Statistical analysis
All results are presented as the mean ± standard deviation. SPSS (Statistical Package for the Social Sciences) statistical software (version 13.0) was used to analyze data (IBM SPSS Inc., Armonk, NY, USA). One-way analysis of variance (ANOVA) was employed to analyze the differences among groups. P-values of P< 0.05 were taken to indicate statistically significant differences.
Abbreviations
LPS=lipopolysaccharide; LiCl=lithium chloride; PRRS=porcine reproductive and respiratory syndrome; PRRSV=porcine reproductive and respiratory syndrome virus; IFNs=Type I interferons; TLRs=Toll-like receptors; IRFs= Type I interferons regulatory factors; PB=Piggyback; LBP=LPS-binding protein; MyD88=myeloid differentiation factors; IL-1=interleukin-1; TNFα=tumor necrosis factor α; GSK3β=glycogen synthase kinase-3β; DMEM=Dulbecco’s modified Eagle’s medium; FBS=fetal bovine serum; GFP=green fluorescent protein; PBR3=PK15-PBIRF3; PBR7=PK15-PBIRF7; PBv=PK15-PBvector; ANOVA=one-way analysis of variance; TRAF6=tumor necrosis factor receptor-associated factor-6; stat3=signal transducers and activators of transcription 3.
Author contributions
He-Ping Li and Yue-Ying Wang designed the experiments and wrote manuscript. Pei-Ge Chen, Yan-Jing Guan, Guang-Ming Zha, and Xian-Qin Jiao conrtibuted to the experiments and writing. He-Shui Zhu, and Cheng-Yu Zhang analyzed the data and cultured cells. All authors have read and approved the final manuscript.
ACKNOWLEDGMENTS
The authors are grateful to Guo-Yu Yang for his suggestions in vector construction and Yu-jie Guo support in preparing this paper.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
FUNDING
This work was supported by the National Key Research and Development Program of China (2016YFD0500503), National Natural Science Fund (31470120). This project was partially supported by the Natural Science Fund of Henan Province (112102310705, 15A230022, 14A230010, 152300410070, 15A230017) and Zhengzhou City Natural Science Fund (141PPTGG407). The funding sources had no involvement in study design; in the collection, analysis and interpretation of data; in the writing of the report; and in the decision to submit the article for publication.
REFERENCES
1. Kimman TG, Cornelissen LA, Moormann RJ, Rebel JM, Stockhofe-Zurwieden N. Challenges for porcine reproductive and respiratory syndrome virus (PRRSV) vaccinology. Vaccine. 2009; 27:3704-18. doi: 10.1016/j.vaccine.2009.04.022.
2. Frossard JP, Hughes GJ, Westcott DG, Naidu B, Williamson S, Woodger NG, Steinbach F, Drew TW. Porcine reproductive and respiratory syndrome virus: genetic diversity of recent British isolates. Vet Microbiol. 2013; 162:507-18. doi: 10.1016/j.vetmic.2012.11.011.
3. Tamura T, Yanai H, Savitsky D, Taniguchi T. The IRF family transcription factors in immunity and oncogenesis. Annu Rev Immunol. 2008; 26:535-84. doi: 10.1146/annurev.immunol.26.021607.090400.
4. Gurtler C, Bowie AG. Innate immune detection of microbial nucleic acids. Trends Microbiol. 2013; 21:413-20. doi: 10.1016/j.tim.2013.04.004.
5. Lin L, Pan S, Zhao J, Liu C, Wang P, Fu L, Xu X, Jin M, Zhang A. HSPD1 interacts with IRF3 to facilitate interferon-beta induction. PLoS One. 2014; 9:e114874. doi: 10.1371/journal.pone.0114874.
6. Honda K, Yanai H, Negishi H, Asagiri M, Sato M, Mizutani T, Shimada N, Ohba Y, Takaoka A, Yoshida N, Taniguchi T. IRF-7 is the master regulator of type-I interferon-dependent immune responses. Nature. 2005; 434:772-777. doi: 10.1038/nature03464.
7. Weiss G, Maaetoft-Udsen K, Stifter SA, Hertzog P, Goriely S, Thomsen AR, Paludan SR, Frøkiær H. MyD88 drives the IFN-β response to Lactobacillus acidophilus in dendritic cells through a mechanism involving IRF1, IRF3, and IRF7. J Immunol. 2012; 189:2860-8. doi: 10.4049/jimmunol.1103491.
8. Feng H, Zhang QM, Zhang YB, Li Z, Zhang J, Xiong YW, Wu M, Gui JF. Zebrafish IRF1, IRF3, and IRF7 differentially regulate IFNF1 and IFNF3 expression through assembly of homoor heteroprotein complexes. J Immunol. 2016; 197:1893-1904. doi: 10.4049/jimmunol.1600159.
9. Li S, Lu LF, Feng H, Wu N, Chen DD, Zhang YB, Gui JF, Nie P, Zhang YA. IFN regulatory factor 10 is a negative regulator of the IFN responses in fish. J Immunol. 2014; 193:1100-9. doi: 10.4049/jimmunol.1400253.
10. Iliev DB, Sobhkhez M, Fremmerlid K, Jørgensen JB. MyD88 interacts with interferon regulatory factor (IRF) 3 and IRF7 in Atlantic salmon (Salmo salar): transgenic SsMyD88 modulates the IRF-induced type I interferon response and accumulates in aggresomes. J Biol Chem. 2011; 286:42715-24. doi: 10.1074/jbc.M111.293969.
11. Beura LK, Sarkar SN, Kwon B, Subramaniam S, Jones C, Pattnaik AK, Osorio FA. Porcine reproductive and respiratory syndrome virus nonstructural protein 1betamodulates host innate immune response by antagonizing IRF3 activation. J Virol. 2010; 84:1574-1584. doi: 10.1128/JVI.01326-09.
12. Dragan AI, Hargreaves VV, Makeyeva EN, Privalov PL. Mechanisms of activation of interferon regulator factor 3: the role of C-terminal domain phosphorylation in IRF-3 dimerization and DNA binding. Nucleic Acids Res. 2007; 35:3525-3534. doi:10.1093/nar/gkm142.
13. Tian WL, Jiang ZX, Wang F, Guo R, Tang P, Huang YM, Sun L. IRF3 is involved inhuman acute myeloid leukemia through regulating the expression of miR-155. BiochemBiophys Res Commun. 2016; 478:1130-1135. doi: 10.1016/j.bbrc.2016.08.080.
14. Guinn Z, Lampe AT, Brown DM, Petro TM. Significant role for IRF3 in both Tcell and APC effector functions during T cell responses. Cell Immunol. 2016; 310:141-149. doi: 10.1016/j.cellimm.2016.08.015.
15. Kumari M, Wang X, Lantier L, Lyubetskaya A, Eguchi J, Kang S, Tenen D, Roh HC, Kong X, Kazak L, Ahmad R, Rosen ED. IRF3 promotes adipose inflammation andinsulin resistance and represses browning. J Clin Invest. 2016; 126:2839-2854. doi: 10.1172/JCI86080.
16. Yant SR, Meuse L, Chiu W, Ivics Z, Izsvak Z, Kay MA. Somatic integration and long-term transgene expression in normal and haemophilic mice using a DNA transposon system. Nat Genet. 2000; 25:35-41. doi:10.1038/75568.
17. Izsvák Z, Hackett PB, Cooper LJN, Ivics Z. Translating sleeping beauty transposition into cellular therapies: Victories and challenges. Bioessays. 2010; 32:756-767. doi: 10.1002/bies.201000027.
18. Cary LC, Goebel M, Corsaro BG, Wang H, Rosen E, Fraser MJ. Transposon mutagenesis of baculoviruses: analysis of Trichoplusiani transposon IFP2 insertions within the FP-locus of nuclear polyhedrosis viruses. Virology. 1989; 172:156-169. doi:10.1016/0042-6822(89)90117-7.
19. Wang H, Mayhew D, Chen X, Johnston M, Mitra RD. “Calling cards” for DNA-binding proteins in mammalian cells. Genetics. 2012; 190:941- 949. doi: 10.1534/genetics.111.137315.
20. Li X, Ewis H, Hice RH, Malani N, Parker N, Zhou L, Feschotte C, Bushman FD, Atkinson PW, Craig NL. A resurrected mammalian hAT transposable element and a closely related insect elementare highly active in human cell culture. Proc Natl Acad Sci. 2013; 110:E478-E487. doi: 10.1073/pnas.1121543109.
21. Taniguchi T, Ogasawara K, Takaoka A, Tanaka N. IRF family of transcription factors as regulators of host defense. Annu Rev Immunol. 2001; 19:623-655. doi:10.1146/annurev.immunol.19.1.623.
22. Honda K, Taniguchi T. IRFs: master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nature Reviews Immunology. 2006; 6:644-658. doi:10.1038/nri1900.
23. Baccala R, Hoebe K, Kono DH, Beutler B, Theofilopoulos AN. TLR-dependent and TLR-independent pathways of type I interferon induction in systemic autoimmunity. Nat Med. 2007; 13:543-551. doi:10.1038/nm1590.
24. Yanai H, Negishi H, Taniguchi T. The IRF family of transcription factors:Inception, impact and implications in oncogenesis. Oncoimmunology. 2012; 1:1376-1386. doi:10.4161/onci.22475.
25. Sato M, Suemori H, Hata N, Asagiri M, Ogasawara K, Nakao K, Nakaya T, Katsuki M, Noguchi S, Tanaka N, Taniguchi T. Distinct and essential roles oftranscription factors IRF-3 and IRF-7 in response to viruses for IFN-alpha/betagene induction. Immunity. 2000; 13:539-48. doi:10.1016/S1074-7613(00)00053-4.
26. Diks SH, Richel DJ, Peppelenbosch MP. LPS signal transduction: the picture is becoming more complex. Current Topics in Medicinal Chemistry. 2004; 4:1115-1126. doi: 10.2174/1568026043388141.
27. Kawai T, Akira S. Toll-like receptor downstream signaling. Arthritis Res Ther. 2005; 7:12-19. doi:10.1186/ar1469.
28. Gao LY, Kwaik YA. The modulation of host cell apoptosis by intracellular bacterial pathogens. Trends Microbiol. 2000; 8:306-313. doi:10.1016/S0966-842X(00)01784-4..
29. Asehnoune K, Strassheim D, Mitra S, Kim JY, Abraham E. Involvement of PKCalpha/beta in TLR4 and TLR2 dependent activation of NF-kappa B. Cellular Signalling. 2005; 17:385-394. doi:10.1016/j.cellsig.2004.08.005.
30. Chen Y, Zhang C, Chiang SK, Wu T, Tso MO. Increased nuclear factor-kappa B p65 immunoreactivity following retinal ischemia and reperfusion injury in mice. J Neurosci Res. 2003; 72:125-131. doi:10.1002/jnr.10548.
31. Nassar A, Azab AN. Effects of lithium on inflammation. ACS Chem Neuro sci. 2014; 5:451-458. doi: 10.1021/cn500038f.
32. Castillo-Quan JI, Li L, Kinghorn KJ, Ivanov DK, Tain LS, Slack C, Kerr F, Nespital T, Thornton J, Hardy J, Bjedov I, Partridge L. Lithium promotes longevity through GSK3/NRF2-dependent hormesis. Cell Rep. 2016; 15:638-650. doi: 10.1016/j.celrep.2016.03.041.
33. Beurel E, Michalek SM, Jope RS. Innate and adaptive immune responses regulated by glycogen synthase kinase-3 (GSK3). Trends Immunol. 2010; 31:24. doi: 10.1016/j.it.2009.09.007.
34. Martin M, Rehani K, Jope RS, Michalek SM. Toll-like receptor mediated cytokine production is differentially regulated by glycogen synthase kinase 3. Nat Immunol. 2005; 6:777-784. doi:10.1038/ni1221.
35. Park MY, Mun ST. Carnosic acid inhibits TLR4-MyD88 signaling pathway inLPS-stimulated 3T3-L1 adipocytes. Nutr Res Pract. 2014; 8:516-520. doi: 10.4162/nrp.2014.8.5.516.
36. Kawai T, Akira S. Innate immune recognition of viral infection. Nat Immunol. 2006; 7:131-7. doi: 10.1038/ni1303.
37. Hiscott J, Pitha P, Genin P, Nguyen H, Heylbroeck C, Mamane Y, Algarte M, Lin R. Triggering the interferon response: the role of IRF-3 transcription factor. J Interferon Cytokine Res. 1999; 19:1-13. doi: 10.1089/107999099314360.
38. Suhara W, Yoneyama M, Kitabayashi I, Fujita T. Direct involvement of CREB-binding protein/p300 in sequence-specific DNA binding of virus-activated interferon regulatory factor-3 holocomplex. J Biol Chem. 2002; 277:22304-13. doi: 10.1074/jbc.M200192200.
39. La Rocca SA, Herbert RJ, Crooke H, Drew TW, Wileman TE, Powell PP. Loss of interferon regulatory factor 3 in cells infected with classical swine fever virus involves the N-terminal protease, Npro. J Virol. 2005; 79:7239-47. doi: 10.1128/JVI.79.11.7239-7247.2005.
40. Schneider WM, Chevillotte MD, Rice CM. Interferon-stimulated genes: a complex web of host defenses. Annu Rev Immunol. 2014; 32:513-45. doi: 10.1146/annurev-immunol-032713-120231.
41. Schoggins JW, Wilson SJ, Panis M, Murphy MY, Jones CT, Bieniasz P, Rice CM. A diverse range of gene products are effectors of the type I interferon antiviral response. Nature. 2011; 472:481-5. doi: 10.1038/nature09907.
42. Ramírez-Carvajal L, Diaz-San Segundo F, Ramirez-Medina E, Rodríguez LL, de Los Santos T. Constitutively Active IRF7/IRF3 Fusion Protein Completely Protects Swine against Foot-and-Mouth Disease. J Virol. 2016; 90:8809-21. doi: 10.1128/JVI.00800-16.
43. Ramírez-Carvajal L, Díaz-San Segundo F, Hickman D, Long CR, Zhu J, Rodríguez LL, de los Santos T. Expression of porcine fusion protein IRF7/3(5D) efficiently controls foot-and-mouth disease virus replication. J Virol. 2014; 88:11140-53. doi: 10.1128/JVI.00372-14.
44. Li Z, Huang Q, Chen H, Lin Z, Zhao M, Jiang Z. Interferon Regulatory Factor 7 Promoted Glioblastoma Progression and Stemness by Modulating IL-6 Expression in Microglia. J Cancer. 2017; 8:207-219. doi: 10.7150/jca.16415.
45. Kim O, Sun Y, Lai FW, Song C, Yoo D. Modulation of type I interferon induction by porcine reproductive and respiratory syndrome virus and degradation of CREB-binding protein by non-structural protein 1 in MARC-145 and HeLa cells. Virology. 2010; 402:315-326. doi: 10.1016/j.virol.2010.03.039.
46. Zhang Q, Guo XK, Gao L, Huang C, Li N, Jia X, Liu W, Feng WH. MicroRNA-23 inhibits PRRSV replication by directly targeting PRRSV RNA and possibly by upregulating type I interferons. Virology. 2014; 450-451:182-195. doi: 10.1016/j.virol.2013.12.020.