
INTRODUCTION
Trichloroethylene (TCE) is a chlorinated hydrocarbon that has long been used as an industrial solvent. Because of inappropriate handling and transportation, TCE has become a major environmental contaminant. Both the U.S. Environmental Protection Agency (EPA) and the International Agency for Research on Cancer (IARC) have characterized TCE as a human carcinogen. TCE is mainly metabolized in the liver and causes hepatic toxicity by altering the status of cytochrome P450 [1]. Occupational exposure to TCE has been linked to severe liver dysfunction [2], and epidemiological studies suggested a strong association of TCE exposure with liver cancer [3, 4]. In animal studies, long-term oral gavage with TCE induced hepatocellular proliferation, mitochondrial dysfunction, and glycogen depletion [5]. We have previously identified SET (I2PP2A) also known as protein phosphatase type 2A inhibitor II [6] as a key mediator of TCE-induced liver cell apoptosis [7, 8]. However, whether SET-mediated changes in protein phosphorylation patterns are involved in TCE-induced cytotoxicity remains unknown. Using quantitative phosphoproteomic methods, we characterized SET-related protein phosphorylation profiles and found that dephosphorylation and upregulation of nucleolin was crucial for TCE-induced hepatic cell apoptosis.
RESULTS
SET knockdown alters nucleolin phosphorylation status in TCE-treated cells
To assess the influence of SET on protein phosphorylation changes induced by TCE, we characterized protein phosphorylation by mass spectrometry in control and SET-siRNA-transfected (SET-knockdown) L-02 cells. After treatment with TCE, 26 phosphosites on 16 proteins were found to be altered in SET-competent, control L-02 cells, whereas 15 phosphosites on 12 proteins showed differential changes in SET-knockdown L-02 cells (Table 1 and Table 2). Among these changes, the phosphorylation of nucleolin residues S184 and S206 was decreased in control cells, and increased instead after SET-knockdown.
Table 1: Identification of TCE induced abnormal (de)-phosphorylation of 27 sites in 16 proteins
Uniport accessions |
Protein descriptions |
Fold change# |
p value |
Phosphorylated peptides* |
phosphorylated sites |
|---|---|---|---|---|---|
Q9UQ35 |
Serine/arginine repetitive matrix protein 2 |
-1.214 |
0.036429 |
RGEGDAPFSEPGTT STQRPSpSPETATK |
S323 |
Q9UQ35 |
Serine/arginine repetitive matrix protein 2 |
-1.74236 |
0.02238 |
SATRPpSpPSPER |
S351 S353 |
Q9UQ35 |
Serine/arginine repetitive matrix protein 2 |
-1.28055 |
0.040114 |
SLpTpRSPPAIR |
T2069 S2071 |
P46821 |
Microtubule-associated protein 1B |
-1.49788 |
0.010365 |
ESpSPLpYSPTFSDSTSAVK |
S1793 S1797 |
P19338 |
Nucleolin |
-1.85542 |
0.005201 |
AAAAAPApSEDEDDED DEDDEDDDDDEEDDpS EEEAMETTPAK |
S184 S206 |
P49736 |
DNA replication licensing factor MCM2 |
-1.39098 |
0.004499 |
GLLYDpSDEEDEERPAR |
S139 |
O00193 |
Small acidic protein |
-1.81218 |
0.036586 |
RSApSPDDDLGSSNWEAA DLGNEER |
S17 |
Q15149-4 |
Isoform 4 of Plectin |
-3.62735 |
0.029429 |
TSpSEDNLYLAVLR |
S21 |
Q9UHB6 |
LIM domain and actin-binding protein 1 |
-2.06584 |
0.018827 |
QQpSPQEPK |
S698 |
Q9UHB6 |
LIM domain and actin-binding protein 1 |
-5.05972 |
0.000401 |
QQpSPQEPK |
S698 |
Q8NE71 |
ATP-binding cassette sub-family F member 1 |
-2.06663 |
0.007659 |
KAEQGpSEEEGEGEEEEE EGGESK |
S228 |
Q8NE71 |
ATP-binding cassette sub-family F member 1 |
-1.83936 |
0.009453 |
QQPPEPEWIGDGESTpSPSDK |
S22 |
Q8WX93 |
Palladin |
-1.94712 |
0.012633 |
IApSDEEIQGTK |
S893 |
P55081 |
Microfibrillar-associated protein 1 |
1.496243 |
0.009119 |
IVEPEVVGEpSpDSEVEGDAWR |
S116 S118 |
O95218 |
Zinc finger Ran-binding domain-containing protein 2 |
-2.58179 |
0.019206 |
ENVEYIEREEpSDGEYDEFGR |
S120 |
Q13541 |
Eukaryotic translation initiation factor 4E-binding protein 1 |
1.054209 |
0.008982 |
RVVLGDGVQLPPGDYpST TPGGTLFSpTTPGGTR |
S37 T46 |
Q13442 |
28 kDa heat- and acid-stable phosphoprotein |
-1.36106 |
0.012011 |
SLDpSDpESEDEEDDYQQK |
S60 S63 |
Q92688 |
Acidic leucine-rich nuclear phosphoprotein 32 family member B |
-2.38103 |
0.04378 |
KREpTDDEGEDD |
T244 |
P11137 |
Microtubule-associated protein 2 |
1.324371 |
0.032474 |
VDHGAEIITQpSPGR |
S1782 |
Q9H3N1 |
Thioredoxin-related transmembrane protein 1 |
-2.01394 |
0.049445 |
KVEEEQEADEEDVpSEEEAESK |
S247 |
# positive fold changes indicate up-regulation and negative fold changes indicate down-regulation.
* the amino acid letters after the lower case of “p” indicate phosphorylation sites.
Table 2: Identification of SET mediated abnormal (de)-phosphorylation of 15 sites in 12 proteins in TCE treated L-02 cells
Uniport accessions |
Protein descriptions |
Fold change# |
p value |
Phosphorylated peptides* |
phosphorylated animo acids |
|---|---|---|---|---|---|
P07900-2 |
Isoform 2 of Heat shock protein HSP 90-alpha |
1.512643 |
0.010983 |
ESEDKPEIEDVGpSDEEEEK |
S385 |
P19338 |
Nucleolin |
3.542418 |
0.005201 |
AAAAAPApSEDEDDEDDEDDE DDDDDEEDDpSEEEAMETTPAK |
S184 S206 |
Q15149-4 |
Isoform 4 of Plectin |
-1.22808 |
0.029429 |
TSpSEDNLYLAVLR |
S21 |
Q14247-2 |
Isoform 2 of Src substrate cortactin |
1.310994 |
0.034449 |
TQpTPPVSPAPQPTEER |
T364 |
Q9UHB6 |
LIM domain and actin-binding protein 1 |
1.908012 |
0.018827 |
QQpSPQEPK |
S698 |
Q8NE71 |
ATP-binding cassette sub-family F member 1 |
1.234261 |
0.007659 |
KAEQGpSEEEGEGE EEEEEGGESK |
S228 |
Q8WX93 |
Palladin |
2.832776 |
0.012633 |
IApSDEEIQGTK |
S893 |
E9PRY8 |
Elongation factor 1-delta |
2.171604 |
0.041186 |
KPATPAEDDEDDDIDL FGpSDNEEEDKEAAQLR |
S578 |
Q13541 |
Eukaryotic translation initiation factor 4E-binding protein 1 |
1.52283 |
0.008982 |
RVVLGDGVQLPPGDYp STTPGGTLFSpTTPGGTR |
S37 T46 |
Q13442 |
28 kDa heat- and acid-stable phosphoprotein |
2.686815 |
0.012011 |
SLDpSDpESEDEEDDYQQK |
S60 S63 |
Q92688 |
Acidic leucine-rich nuclear phosphoprotein 32 family member B |
2.008763 |
0.04378 |
KREpTDDEGEDD |
T244 |
O94804 |
Serine/threonine-protein kinase 10 |
1.42668 |
0.039742 |
QVAEQGGDLpSPAANR |
S438 |
* the lower case of letter “p” before the amino acid represents the position of the phosphorylation and the amino acid is which to have been phosphorylated.
# The fold change data were log2 transformed, thus negative numbers indicated decreasing and positive numbers indicated increasing of phosphorylation.
To confirm the above results, lysates from SET-siRNA-transfected L-02 cells (SET-siRNA1 and SET-siRNA4) were analyzed by phosphate-affinity SDS-PAGE. The presence of Phos-Tag™ in the acrylamide gels reduces the mobility of phosphorylated proteins, allowing further separation of differentially phosphorylated proteins. The results indicated that following TCE treatment, nucleolin was dephosphorylated and upregulated in L-02 cells, while knockdown of SET prevented these changes (Figure 1).
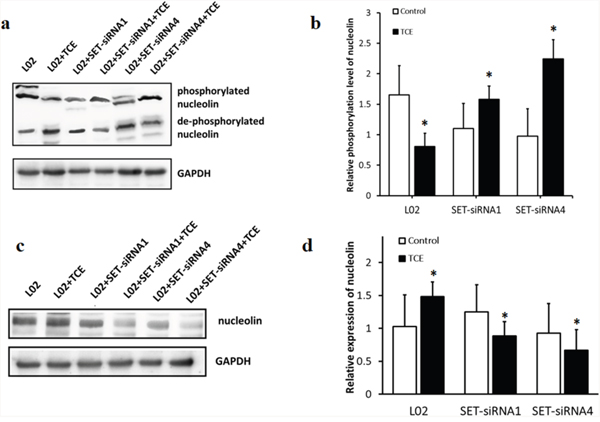
Figure 1: SET decreases phosphorylation of nucleolin and increases its expression in TCE-exposed liver cells. L-02 cells were interfered with two different SET-siRNA oligonucleotides. (a) phosphorylated and unphosphorylated nucleolin were separated by SDS-PAGE; (b) after TCE treatment, the phosphorylation of nucleolin decreased in control L-02 cells and increased after SET-siRNA-transfection (*: P < 0.05); (c) nucleolin expression, protein band image; (d) after TCE treatment, the expression of nucleolin increased in control L-02 cells and decreased after SET-siRNA-transfection (*: P < 0.05, compared with control). Note that the pattern of nucleolin expression was inversely correlated with its pattern of phosphorylation.
Nucleolin dephosphorylation prevents its binding to c-myc
C-myc was reported to be a major transcriptional regulator of nucleolin, while nucleolin, in turn, negatively regulates the transcription of c-myc [9, 10]. Since the overall phosphorylation status of nucleolin was found to be decreased in TCE-treated L-02 liver cells, and this was prevented by SET knockdown, co-IP experiments were performed to determine whether the phosphorylation of nucleolin affected its binding to c-myc. Results indicated that inhibition of c-myc downregulated nucleolin expression (Figure 2), and TCE-induced nucleolin dephosphorylation inhibited in turn its binding to c-myc (Figure 3).
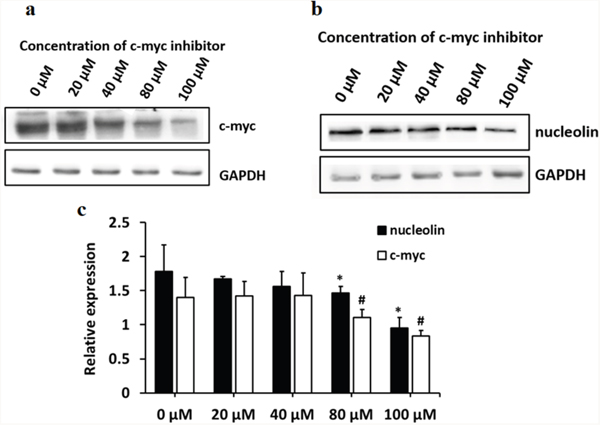
Figure 2: c-myc controls the expression of nucleolin in L-02 cells. L-02 cells were treated with different concentrations of 10058-F4, a c-myc inhibitor. (a) c-myc protein band; (b) nucleolin protein band; (c) both c-myc and nucleolin expression were significantly decreased in cells treated with c-myc inhibitor concentrations of 80 μM and 100 μM (#: P < 0.05, relative expression of c-myc, compared with 0 μM; (*: P < 0.05, relative expression of nucleolin, compared with 0 μM).
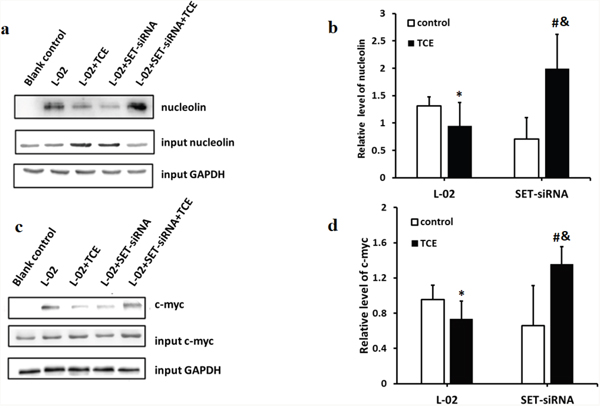
Figure 3: Dephosphorylation impairs the ability of nucleolin to bind to c-myc. (a) c-myc protein band eluted from immobilized nucleolin; (b) the ability of c-myc to capture nucleolin was impaired in TCE-treated L-02 liver cells, but improved however after SET knockdown (*: P < 0.05, compared with control; &: P < 0.05, compared with TCE-treated L-02 cells); (c) nucleolin protein band eluted from immobilized c-myc; (d) the ability of nucleolin to capture c-myc was impaired by TCE treatment in L-02 cells; this interaction improved after SET knockdown (*: P < 0.05, compared with control; &: P < 0.05, compared with TCE treated, L-02 control cells).
Nucleolin knockdown attenuates SET-mediated hepatic cell apoptosis induced by TCE exposure
To test the hypothesis that SET-mediated nucleolin overexpression contributes to TCE-induced apoptosis, L-02 cells were transfected with lentivirus-containing siRNAs against nucleolin. Western-blot analysis indicated that nucleolin was successfully down-regulated in L-02 cells (Figure 4). TCE-induced apoptosis of L-02 cells was suppressed after SET and nucleolin knockdown (Figure 5). These data suggest that nucleolin expression contributes to SET-mediated apoptosis in liver cells exposed to TCE.
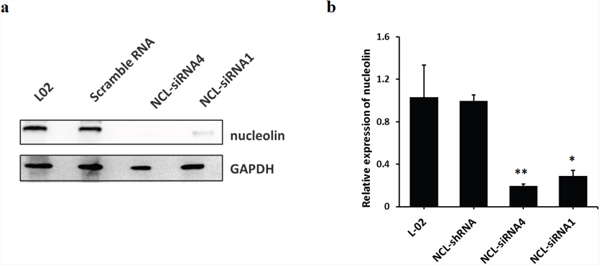
Figure 4: Stable knockdown of nucleolin in L-02 cells. (a) nucleolin protein band; (b) successful nucleolin knock down in L-02 cells by lentivirus-mediated RNA interference. The efficacy of NCL-siRNA4 was higher than that of NCL-siRNA1 (*: P < 0.05, compared with L-02 control cells; **: P < 0.01, compared with L-02 control cells).
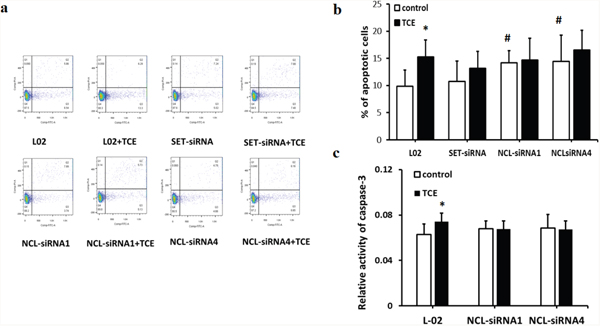
Figure 5: SET and nucleolin siRNAs attenuate TCE-induced apoptosis in human liver cells. (a) flow cytometry analysis of cell apoptosis by PI/Annexin-V staining; (b, c) TCE-induced apoptosis in L-02 cells was attenuated after SET knockdown and nucleolin knockdown (*: P < 0.05, compared with control,; #: P < 0.05, compared with L-02 cells); (c) caspase-3 activity assay indicates inhibition of apoptosis upon nucleolin knockdown in TCE-treated L-02 cells (P < 0.05).
DISCUSSION
SET is amulti-functional protein that specifically inhibits phosphatase 2A in eukaryotic cells, and has been linked to liver carcinogenesis [11]. Our previous studies suggested that SET is as a key mediator of TCE-induced apoptosis in hepatocytes, although the specific mechanisms remained unclear [8, 12, 13]. Using phosphoproteomics analysis, we found that the phosphorylation status of nucleolin was differently affected (increased) upon siRNA-mediated SET knockdown in TCE-exposed human liver L-02 cells. We also found that nucleolin phosphorylation was negatively associated with its own expression. Our data showed that dephosphorylation of nucleolin impaired its ability to bind c-myc, a major transcription factor that induces the expression of many genes, including nucleolin. Since in its phosphorylated state nucleolin represses c-myc activity, dephosphorylation of nucleolin resulted in an increase in its own expression. Furthermore, our study underscored a major role of nucleolin in TCE-mediated liver cell toxicity, as nucleolin knockdown attenuated L-02 cell apoptosis following treatment with TCE (Figure 6). Thus, our results identify nucleolin as a SET-regulated phosphoprotein critically involved in TCE-induced liver cell toxicity.
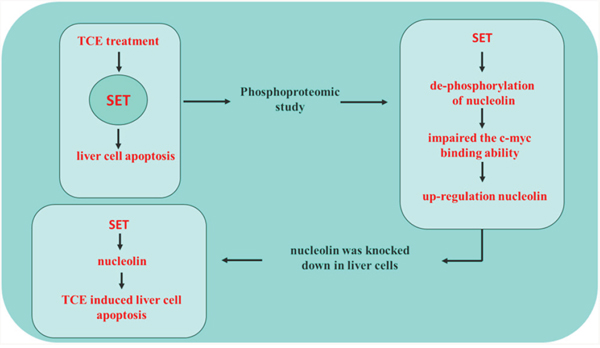
Figure 6: SET mediates TCE-induced liver cell apoptosis through upregulation of nucleolin. After TCE exposure, SET up-regulates nucleolin by promoting its dephosphorylation, which impairs its ability to bind its transcriptional activator, c-myc, further leading to liver cell toxicity.
Nucleolin is an abundant, multifunctional phosphoprotein mainly located in the nucleolus. It is involved in chromatin structure stabilization, rRNA maturation, ribosome assembly, and nucleo-cytoplasmic transport [14]. Cell division cycle 2 kinase (cdc2)-mediated phosphorylation of nucleolin was found to be important for cell mitosis [15, 16]. A potential contribution of nucleolin to carcinogenesis has been highlighted by past investigations. For instance, nucleolin was shown to promote transformation of non-small cell lung cancer by regulating cytosolic phospholipase 2A [17]. Another study showed that phosphorylation-induced redistribution of nucleolin could promote tumor metastasis through the PI3K/Akt pathway in colorectal carcinoma [18]. Nucleolin has also been observed to suppress Fas-mediated apoptosis by blocking the binding of Fas ligands to Fas [19]. Of potential therapeutic interest, a small amount of nucleolin has been found to be expressed on cell membranes, serving as a receptor for PEGylated, poly-lysine packaged DNA particles [20].
In summary, the present study demonstrated that SET-mediated dephosphorylation of nucleolin in TCE-treated human L-02 liver cells promotes nucleolin expression by suppressing its c-myc binding ability. Importantly, SET and nucleolin knockdown prevented TCE-induced apoptosis in these cells. Although further work is needed to characterize in detail the mechanisms underlying these observations, the present findings provide important insights into the molecular bases of TCE hepatotoxicity, and will hopefully help guide prevention and treatment efforts.
MATERIALS AND METHODS
Reagents
The human liver HL-7702 (L-02) cell line was purchased from the Institute of Cell Biology of the Chinese Academy of Sciences (Shanghai, China). SET-siRNA transfected L-02 cells were derived from a previous study [8]. The iTRAQ 8 Plex Labeling Kit was purchased from SCIEX (Framingham, MA, U.S.). Reagents for cell culture, titanium dioxide spin columns, Co-IP kit, Alexa Fluor® 488 Annexin V/Dead Cell Apoptosis Kit, C18 columns, and electro-generated chemical luminescence substrates were obtained from Thermo Scientific (Rockford, IL, U.S.). Sequencing grade trypsin, triethylammonium bicarbonate (TEAB), acetonitrile, isopropyl alcohol, pyrroline, and ammonium hydroxide were obtained from Sigma-Aldrich (Shanghai, China). The Phos-Tag™ reagent was purchased from Wako Pure Chemicals (Richmond, VA, U.S.). Antibodies against nucleolin and c-myc were purchased from Abcam (Cambridge, MA, U.S.). Antibody against GAPDH were purchased from Santa Cruz Biotechnology (Dallas, TX, U.S.). Reagents for Western-blot were obtained from Bio-Rad (Hercules, CA, U.S.).
Cell culture and treatment
L-02 cells and SET-siRNA-transfected L-02 cells were cultured in RPMI-1640 medium supplemented with 12% fetal bovine serum, 100 μg/ml streptomycin and 100 units/ml penicillin at 37°C with 5% CO2. The cells were treated with TCE at a concentration of 8 mmol/L, which was chosen based on previous observations [21]. TCE was pre-dissolved in 50 μL of DMSO and added to 10 mL of culture medium. Treatment lasted 24 h. Separately, L-02 cells were treated with different concentrations of a c-myc inhibitor (10058-F4, Santa Cruz) for 24 h.
Phosphoproteomics study
After treatment with TCE cells were harvested and centrifuged at 1,200 rpm. Proteins were extracted by PhosphoSafe Extraction Reagent (Thermo Scientific). Protein buffer was then replaced by TEAB using 3kDa filtration devices (Millipore). Total protein levels were quantified using a BCA protein quantification kit (Thermo Scientific). Proteins were digested with trypsin and digestion products were labeled with iTRAQ (8-plex); a pooled sample from all groups served as the internal control. The TiO2-enriched phosphopeptides were separated using liquid chromatography (3 μm, 100 Å, 75 μm i.d. × 15 cm, Acclaim PepMap100, C18, Dionex) and analyzed using mass spectrometry (TripleTOF 5600 System, AB SCIEX). Data were processed and searched against the Uniprot human protein database (a total of 71,434 entries) with the MASCOT search engine. All reporter ion intensities groups were fractioned to internal standard, and the limma package for R 3.1.1 (obtained from Bioconductor, http://bioconductor.org) was used for quantitative analysis of phosphopeptides.
Western blot analysis
Proteins were extracted for Western blot analysis using PhosphoSafe Extraction Reagent and quantified using a BCA protein quantification kit. Equal amounts (50 μg of each sample) of protein extracts were electrophoresed on Phos-Tag™ / 10% SDS-PAGE gels (containing 0.2 mmol/L MnCl2 and 0.1 mmol/L Phos-Tag™). The samples were then transferred onto PVDF membranes and blocked with 5% skimmed milk powder in TBST. Antibodies against nucleolin and c-myc were diluted 1:1,000. Anti–GAPDH antibody was diluted 1:3,000 in TBST (0.2% Tween-20 in TBS). The membranes were incubated with diluted antibodies for 1 h at room temperature, washed 3 times with TBST, and incubated with HRP-conjugated goat anti-mouse antibodies (diluted 1:3,000 in TBST) for 1 h at room temperature. After exposure to ECL substrate, protein band images were captured by ImageQuant RT ECL System (GE Healthcare) and analyzed using ImageQuant TL-1D Analysis Tool. Overall phosphorylation levels were calculated as the ratio between the sum of phosphorylated isoforms and total proteins.
Co-immunoprecipitation assay
First, 10 μg of antibodies against c-myc and nucleolin were conjugated and cross-linked to protein A/G plus agarose (Pierce Crosslink IP Kit, Thermo). Proteins were extracted from TCE-treated cells in accordance with the manufacturer’s instructions and incubated with cross-linked antibodies.
Recombination of lentiviral vectors and generation of stable nucleolin knockdown cells
Four siRNA oligonucleotides targeting the mRNA of nucleolin (NCL-siRNA), as well as a scrambled RNA oligonucleotide, were designed and synthesized with BamHI and EcoRI restriction site enzymes (Sangon Biotech Co., Ltd.; Shanghai, China). The two siRNAs with the highest interference performance were used in subsequent studies (NCL- siRNA1: gatccGCGATGAGGATGACGAAGATTTCAAGAGAATCTTCGTCATCCTC ATCGTTTTTT, NCL-siRNA4: gatccGGAAGACGGTGAAATTGATTTCAAGAGAATCAATTTCACCGTCTTCC TTTTTT). The siRNAs and scrambled RNA oligonucleotides were cloned into pLVX-shRNA1 plasmids. The pseudotyped lentiviruses were produced by co-transfecting recombinant siRNA plasmids with Clontech Lenti-X HT packaging vectors into 293T cells. After 48 h, the lentivirus-containing supernatants were collected by centrifugation at 1,200 rpm for 10 min. L-02 cells were treated with supernatants for 24 h and stable nucleolin knockdown cells were isolated after 2 weeks of selection with puromycin (2 μg/mL). Knockdown efficiency was verified by Western blot analysis. SET-siRNA production and RNA interference efficacy have been described and validated in our previous studies (Liu et al. 2007; Yang et al. 2012).
Apoptosis assays
PI/Annexin V- staining assay
After TCE treatment, approximately 5 × 105 of control and NCL-siRNA-transfected liver cells were collected using low speed centrifugation (1,200 rpm for 5 min at room temperature). The cells were washed twice with PBS, resuspended in the Alexa Fluor® 488 Annexin V binding buffer and then stained with PI/ Alexa Fluor 488Annexin V-at room temperature in the dark for 15 min. Apoptosis levels were measured using a flow cytometer (BD FACSCanto™ II).
Caspase-3 activity assay
Caspase-3 activity was measured using a Caspase-3 activity kit in accordance with the manufacturer’s instructions (Biovision). Cells were collected and washed 3 times in ice-cold PBS, re-suspended in lysis buffer, incubated on ice for 20 min, and then centrifuged at 12,000g at 4°C for 10 min. The supernatant was kept and protein concentration was quantified using a BCA protein quantification kit (Thermo). Then 100 μg of protein extracts were diluted with dilution buffer and incubated with DEVD-pNA substrate at 37°C for 1 h. The samples’ absorbance was measured at 400 nm using a TECAN Infinite M1000 microplate reader.
CONFLICTS OF INTEREST
The authors do not have any conflicts of interest or financial interests to disclose.
FUNDING INFORMATION
This work was supported by the National Natural Science Foundation of China (NDFC) [81273126], the Project of Guangdong Natural Science Foundation [2016A030310033], and the Project of Shenzhen Basic Research Plan [JCYJ20150402102135491, JCYJ20150402102135497, JCYJ20160428142316603].
REFERENCES
1. Lash LH, Putt DA, Huang P, Hueni SE, Parker JC. Modulation of hepatic and renal metabolism and toxicity of trichloroethylene and perchloroethylene by alterations in status of cytochrome P450 and glutathione. Toxicology. 2007; 235:11–26.
2. Xu X, Yang R, Wu N, Zhong P, Ke Y, Zhou L, Yuan J, Li G, Huang H, Wu B. Severe hypersensitivity dermatitis and liver dysfunction induced by occupational exposure to trichloroethylene. Ind Health. 2009; 47:107–12.
3. Karami S, Bassig B, Stewart PA, Lee KM, Rothman N, Moore LE, Lan Q. Occupational trichloroethylene exposure and risk of lymphatic and haematopoietic cancers: a meta-analysis. Occup Environ Med. 2013; 70:591–99.
4. Hansen J, Sallmén M, Seldén AI, Anttila A, Pukkala E, Andersson K, Bryngelsson IL, Raaschou-Nielsen O, Olsen JH, McLaughlin JK. Risk of cancer among workers exposed to trichloroethylene: analysis of three Nordic cohort studies. J Natl Cancer Inst. 2013; 105:869–77.
5. Sano Y, Nakashima H, Yoshioka N, Etho N, Nomiyama T, Nishiwaki Y, Takebayashi T, Oame K. Trichloroethylene liver toxicity in mouse and rat: microarray analysis reveals species differences in gene expression. Arch Toxicol. 2009; 83:835–49.
6. Switzer CH, Cheng RY, Vitek TM, Christensen DJ, Wink DA, Vitek MP. Targeting SET/I(2)PP2A oncoprotein functions as a multi-pathway strategy for cancer therapy. Oncogene. 2011; 30:2504–13.
7. Liu J, Huang H, Xing X, Xi R, Zhuang Z, Yuan J, Yang F, Zhao J. Comparative proteomic analysis on human L-02 liver cells treated with varying concentrations of trichloroethylene. Toxicol Ind Health. 2007; 23:91–101.
8. Yang X, Jiang Y, Li J, Hong WX, Wu D, Huang X, Huang H, Zhou L, Yang L, Yuan J, Zhuang Z, Liu J. Lentivirus-mediated silencing of I2PP2A through RNA interference attenuates trichloroethylene-induced cytotoxicity in human hepatic L-02 cells. Toxicol Lett. 2012; 209:232–38.
9. González V, Hurley LH. The C-terminus of nucleolin promotes the formation of the c-MYC G-quadruplex and inhibits c-MYC promoter activity. Biochemistry. 2010; 49:9706–14.
10. Greasley PJ, Bonnard C, Amati B. Myc induces the nucleolin and BN51 genes: possible implications in ribosome biogenesis. Nucleic Acids Res. 2000; 28:446–53.
11. Fukukawa C, Shima H, Tanuma N, Ogawa K, Kikuchi K. Up-regulation of I-2(PP2A)/SET gene expression in rat primary hepatomas and regenerating livers. Cancer Lett. 2000; 161:89–95.
12. Ren X, Yang X, Hong WX, Huang P, Wang Y, Liu W, Ye J, Huang H, Huang X, Shen L, Yang L, Zhuang Z, Liu J. Identification of the proteins related to SET-mediated hepatic cytotoxicity of trichloroethylene by proteomic analysis. Toxicol Lett. 2014; 227:12–19.
13. Hong WX, Yang L, Chen M, Yang X, Ren X, Fang S, Ye J, Huang H, Peng C, Zhou L, Huang X, Yang F, Wu D, et al. Proteomic analysis of trichloroethylene-induced alterations in expression, distribution, and interactions of SET/TAF-Iα and two SET/TAF-Iα-binding proteins, eEF1A1 and eEF1A2, in hepatic L-02 cells. Toxicol Appl Pharmacol. 2012; 263:259–72.
14. Ginisty H, Sicard H, Roger B, Bouvet P. Structure and functions of nucleolin. J Cell Sci. 1999; 112:761–72.
15. Dranovsky A, Vincent I, Gregori L, Schwarzman A, Colflesh D, Enghild J, Strittmatter W, Davies P, Goldgaber D. Cdc2 phosphorylation of nucleolin demarcates mitotic stages and Alzheimer’s disease pathology. Neurobiol Aging. 2001; 22:517–28.
16. Belenguer P, Caizergues-Ferrer M, Labbé JC, Dorée M, Amalric F. Mitosis-specific phosphorylation of nucleolin by p34cdc2 protein kinase. Mol Cell Biol. 1990; 10:3607–18.
17. Tsou JH, Chang KY, Wang WC, Tseng JT, Su WC, Hung LY, Chang WC, Chen BK. Nucleolin regulates c-Jun/Sp1-dependent transcriptional activation of cPLA2alpha in phorbol ester-treated non-small cell lung cancer A549 cells. Nucleic Acids Res. 2008; 36:217–27.
18. Wu DM, Zhang P, Liu RY, Sang YX, Zhou C, Xu GC, Yang JL, Tong AP, Wang CT. Phosphorylation and changes in the distribution of nucleolin promote tumor metastasis via the PI3K/Akt pathway in colorectal carcinoma. FEBS Lett. 2014; 588:1921–29.
19. Wise JF, Berkova Z, Mathur R, Zhu H, Braun FK, Tao RH, Sabichi AL, Ao X, Maeng H, Samaniego F. Nucleolin inhibits Fas ligand binding and suppresses Fas-mediated apoptosis in vivo via a surface nucleolin-Fas complex. Blood. 2013; 121:4729–39.
20. Chen X, Kube DM, Cooper MJ, Davis PB. Cell surface nucleolin serves as receptor for DNA nanoparticles composed of pegylated polylysine and DNA. Molecular therapy. 2008; 16:333-342.
21. Huang HY, Liu JJ, Xi RR, Xing XM, Yuan JH, Yang LQ, Tao GH, Gong CM, Zhuang ZX. An investigation of hormesis of trichloroethylene in L-02 liver cells by differential proteomic analysis. Mol Biol Rep. 2009; 36:2119–29.