
INTRODUCTION
Gastric cancer is a major health threat in China and other countries [1–3]. Although significant progresses have been made in pathological mechanism research and therapeutic strategies for this devastating disease, the prognosis is still poor [4–6]. Its incidence, on the other hand, has been steadily rising in China [7] and possible other countries [1, 8]. The application of the conventional cytotoxic agents and molecular-targeted agents is not satisfactory when facing pre-existing and/or acquired resistance [5, 6, 9]. Therefore, research focus is to understand the molecular mechanisms of gastric cancer tumorigenesis and progression, and to identify novel oncotarget proteins for this malignancy [5, 6, 9].
Autophagy is important in the progression of a number of cancers [10–15]. Although some studies suggested that autophagy might promote cancer cell death, many others have proposed that autophagy is indeed important for cancer cell survival and apoptosis-resistance [13, 14, 16, 17]. Among all the autophagy-related genes (ATGs), Ulk1 (UNC51-like kinase 1) serves as a key upstream signaling for autophagy initiation [18–21]. This serine-threonine kinase is the mammalian orthologue of yeast ATG1 [18–21]. Ulk1 forms a complex with Atg13, FIP200, and Atg101, which is required to start autophagy [18–21]. It expression and potential functions in human gastric cancer were tested in the current study.
RESULTS
Ulk1 over-expression in fresh human gastric cancer tissues
We first tested expression of Ulk1 in human gastric cancer specimens. Tissue lysates were prepared from the fresh gastric cancer samples. Quantitative real-time PCR (“qRT-PCR”) assay was performed to test Ulk1 mRNA expression in above lysates. Results in Figure 1A demonstrated that Ulk1 mRNA level was significantly elevated in the gastric cancer tissues (“Tumor”, n=12), as compared to that in the surrounding normal gastric epithelial tissues (“Normal”, n=12) (Figure 1A). Ulk1 mRNA level in cancer tissues was about four times higher than that in the normal tissues (Figure 1A). Western blotting assay was applied next to test Ulk1 protein expression in above tissues. Quantified blot results integrating total 12 tissue samples confirmed significant Ulk1 protein upregulation in cancer tissues (Figure 1B). Together, these results demonstrate Ulk1 over-expression in fresh human gastric cancer tissues.
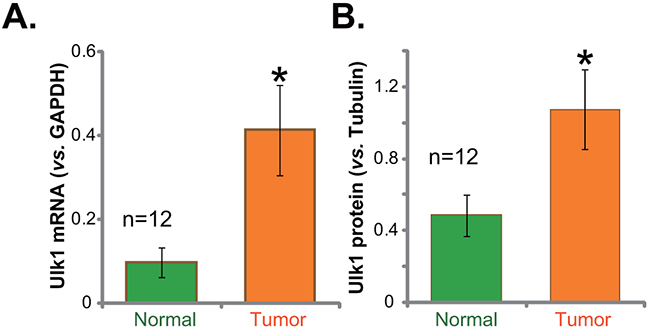
Figure 1: Ulk1 over-expression in fresh human gastric cancer tissues. The fresh human gastric cancer tissues (“Tumor”, n=12) and the surrounding normal gastric epithelial tissues (“Normal”, n=12) were lysed; Expressions of Ulk1 mRNA (A) qRT-PCR assay and Ulk1 protein (Western blotting assay) were tested. Ulk1 protein expression was quantified (B) vs. loading Tubulin, n=12. *p<0.05 vs. “Normal”.
Ulk1 over-expression in human gastric cancer cells
Next, we tested expression of Ulk1 in human gastric cancer cells. In this study, three distinct gastric cancer cell lines, AGS, HGC-27, and SNU601, were cultured. qRT-PCR assay was again employed to test Ulk1 mRNA expression in the cancer cell lines, and its level was compared with that in GES-1 gastric mucosal epithelial cells (non-cancerous normal cells [22]). Results in Figure 2A demonstrated that Ulk1 mRNA level was significantly higher in the above gastric cancer cells than that in GES-1 epithelial cells. Consequently, Ulk1 protein expression was also increased in cancer cells (Figure 2B). Quantification results showed about 2-3 times higher of Ulk1 protein expression in cancer cells (vs. GES-1 epithelial cells, Figure 2B). Among all the tested cancer cell lines, AGS cells expressed highest level of Ulk1 (Figure 2A and 2B). This cell line was chosen for further studies.
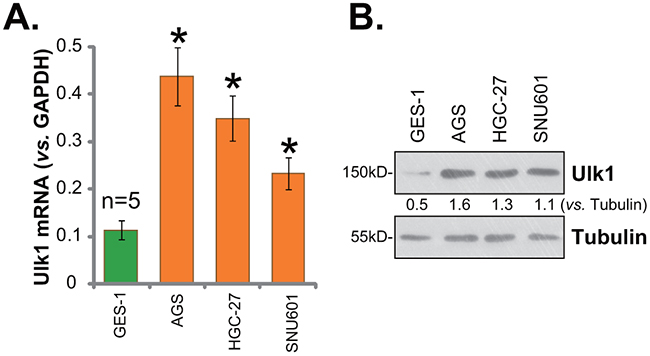
Figure 2: Ulk1 over-expression in human gastric cancer cells. Human gastric cancer cell lines (AGS, HGC-27, and SNU601) and the GES-1 gastric mucosal epithelial cells were subjected to qRT-PCR assay (A) and Western blotting assay (B) to test Ulk1 expression. * p<0.05 vs. GES-1 cells. Experiments in this figure were repeated three times, and similar results were obtained.
Ulk1 shRNA knockdown inhibits gastric cancer cell survival
In order to study the function of Ulk1 in gastric cancer cell survival, shRNA strategy was employed. As demonstrated, two distant lentiviral Ulk1 shRNAs (“1#” and “2#”), with non-overlapping targeting sequence, were applied here. As shown in Figure 3A and 3B, the two Ulk1 shRNAs (“1#” and “2#”) both efficiently downregulated Ulk1 in AGS gastric cancer cells. Ulk1 protein (Figure 3A) and Ulk1 mRNA (Figure 3B) were both decreased after infection of the shRNA. Notably, Ulk1 shRNA-2# was significantly more efficient than Ulk1 shRNA-1# in silencing Ulk1 (Figure 3A and 3B). MTT assay was applied next to test survival of above cells. Results in Figure 3C demonstrated that Ulk1 shRNA knockdown decreased MTT viability OD of AGS cells, indicating an anti-survival activity by Ulk1 shRNA. Meanwhile, AGS cell proliferation, tested by the BrdU ELISA assay, was also inhibited after Ulk1 knockdown (Figure 3D). On the other hand, basal cell apoptosis intensity was enhanced following Ulk1 depletion in AGS cells (Figure 3E). Cell apoptosis was quantified via the Histone DNA ELISA assay (Figure 3E). Notably, Ulk1 shRNA-2# was more dramatic than Ulk1 shRNA-1# in inhibiting AGS cell survival (Figure 3C), proliferation (Figure 3D), and promoting cell apoptosis (Figure 3E).
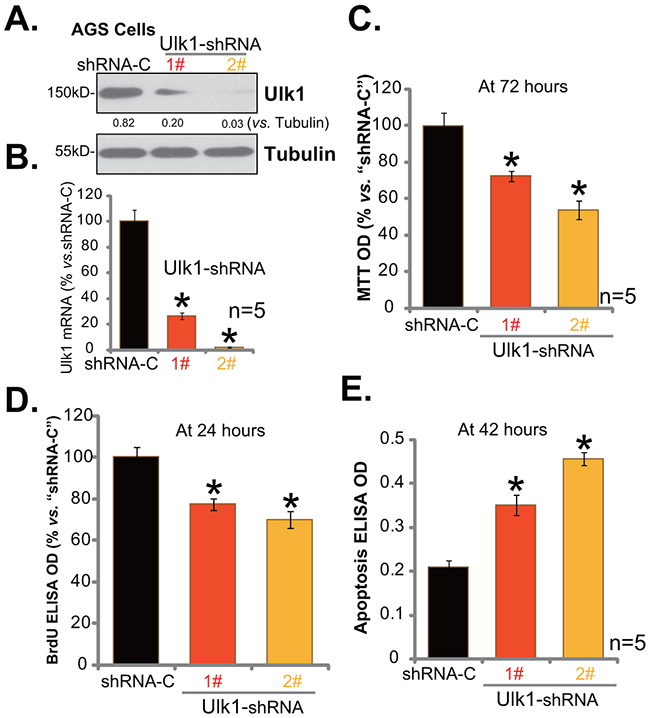
Figure 3: Ulk1 shRNA knockdown inhibits gastric cancer cell survival. Expressions of Ulk1 protein (A) and Ulk1 mRNA (B) in AGS cells with lentiviral Ulk1 shRNA (“1#” or “2#”) or scramble control shRNA (“shRNA-C”) were shown. Above cells were also subjected to MTT assay (C), BrdU ELISA assay (D) and Histone DNA ELISA assay (E) to test cell survival, proliferation and apoptosis, respectively. For these assays, exact same number of viable cells with listed shRNA was plated initially (At 0 hour). *p<0.05 vs. “shRNA-C” cells. Experiments in this figure were repeated four times, and similar results were obtained.
Exogenous over-expression of Ulk1 promotes gastric cancer cell survival
Based on the above results, it is speculated that Ulk1 over-expression might possibly promote gastric cancer cell survival and proliferation. Thus, an Ulk1 expression vector, tagged with green fluorescence protein (GFP), was constructed. This construct was transfected to AGS cells. Western blotting assay results in Figure 4A confirmed expression of Ulk1-GFP in the transfected stable AGS cells. Ulk1 mRNA level was also remarkably elevated in the Ulk1-GFP-expressing cells (Figure 4B). Significantly, exogenous over-expression of Ulk1 indeed promoted AGS cell survival (Figure 4C, tested by MTT assay) and proliferation (Figure 4D, tested by the BrdU ELISA assay). Meanwhile, basal apoptosis intensity, or apoptosis ELISA OD, was decreased following Ulk1 over-expression in AGS cells (Figure 4E). Therefore, exogenous over-expression of Ulk1 promotes AGS cell survival.
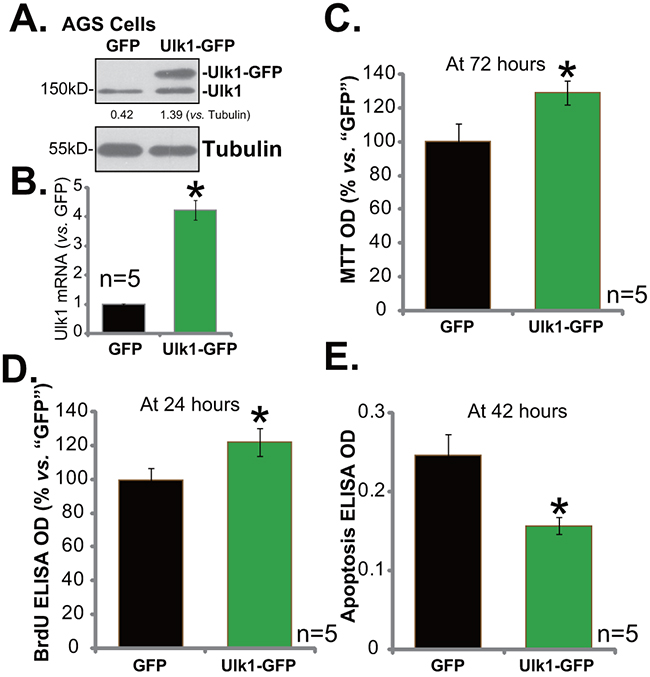
Figure 4: Exogenous over-expression of Ulk1 promotes gastric cancer cell survival. Expressions of Ulk1 protein (A) and Ulk1 mRNA (B) in AGS cells with exogenous Ulk1 (GFP-tagged) or empty vector (pSuper-puro-GFP, “GFP”) were shown. Above cells were also subjected to MTT assay (C), BrdU ELISA assay (D) and Histone DNA ELISA assay (E) to test cell survival, proliferation and apoptosis, respectively. For the assays, exact same number of viable cells with Ulk1-GFP or empty vector was plated initially (At 0 hour). *p<0.05 vs. “GFP” cells. Experiments in this figure were repeated three times, and similar results were obtained.
Ulk1 over-expression in gastric cancer correlates with patients’ T classification and cancer relapse
To further test Ulk1 expression, we examined its level in paraffin-embedded tissues via immunohistochemistry (IHC) staining. A total of 145 patients with gastric cancer (112 males and 33 females; mean age 58.9 years, Table 1), administrated at author’s hospital, were enrolled. Among all the patients, 25, 34 and 86 patients were at stage I, II and III, respectively (based on AJCC TNM staging system). Their paraffin-embedded cancer tissues were tested.
Table 1: Correlation between Ulk1 expression and clinical features of gastric cancer
Patients characteristics |
Ulk1(low) |
Ulk1(high) |
P value |
||
|---|---|---|---|---|---|
cases |
% |
cases |
% |
||
Age (years) |
0.653 |
||||
≤60 |
16 |
20% |
64 |
80% |
|
>60 |
15 |
23.10% |
50 |
76.90% |
|
Sex |
0.979 |
||||
Male |
24 |
21.40% |
88 |
78.60% |
|
Female |
7 |
21.20% |
26 |
78.80% |
|
T classification |
0.014* |
||||
T1 |
8 |
50.00% |
8 |
50.00% |
|
T2 |
7 |
23.30% |
23 |
76.70% |
|
T3 |
1 |
6.30% |
15 |
93.80% |
|
T4a |
15 |
18.10% |
68 |
81.90% |
|
N classification |
0.737 |
||||
N0 |
12 |
25.50% |
35 |
74.50% |
|
N1 |
6 |
20.70% |
23 |
79.30% |
|
N2 |
8 |
22.20% |
28 |
77.80% |
|
N3 |
5 |
15.20% |
28 |
84.80% |
|
Stage |
0.227 |
||||
I |
8 |
32.00% |
17 |
68% |
|
II |
8 |
23.50% |
26 |
76.50% |
|
III |
15 |
17.40% |
71 |
76.50% |
|
Lauren classification |
0.224 |
||||
Intestinal |
20 |
24.40% |
62 |
75.60% |
|
Mixed |
5 |
29.40% |
12 |
70.60% |
|
Diffuse |
6 |
13.00% |
40 |
87.00% |
|
N classification |
0.56 |
||||
Poor |
7 |
15.60% |
38 |
84.40% |
|
M-P |
18 |
25.00% |
54 |
75.00% |
|
M |
6 |
23.10% |
20 |
76.90% |
|
W-M |
0 |
0% |
2 |
100% |
|
Vessel invasion |
0.15 |
||||
No |
26 |
24.30% |
81 |
75.70% |
|
Yes |
5 |
13.20% |
33 |
86.80% |
|
Relapse |
0.007* |
||||
No |
27 |
27.80% |
70 |
72.20% |
|
Yes |
4 |
8.30% |
44 |
91.70% |
|
Ulk1 was stained as buffy color in the cytoplasm. High expression of Ulk1 correlated with T classification (P<0.05) and cancer relapse (P<0.05). Ulk1 expression did not correlate with age, sex, N classification, stage, Lauren classification, differentiation, histological grade, and Vessel invasion (P>0.05).
The IHC staining assay results showed that Ulk1 was mainly expressed in the cytoplasm of cancer cells (Figure 5A and 5B). Again, Ulk1 IHC intensity was significantly higher in the cancer tissues (“Tumor”, Figure 5A) than that in the surrounding normal epithelial tissues (“Normal”, Figure 5B). As summarized in Table 1, the majority (114 out of 145) gastric cancers showed high Ulk1 expression, and the other 31 patients showed low Ulk1 expression. Table 1 demonstrated the correlation between Ulk1 expression and patients’ clinic-pathological characteristics. Among all the clinic features, high Ulk1 expression was significantly correlated with T classification (P=0.014) and cancer relapse (P=0.007). In accordance with chi-square test, the Ulk1 overexpressed cancers have higher recurrence rate compared with that with low level of Ulk1. Notably, high Ulk1 expression did not correlate with age, sex, N classification, stage, Lauren classification, differentiation, histological grade, and Vessel invasion (P > 0.05) (Table 1). These results suggest that high Ulk1 expression in gastric cancer is correlated with patients’ T classification and cancer relapse.
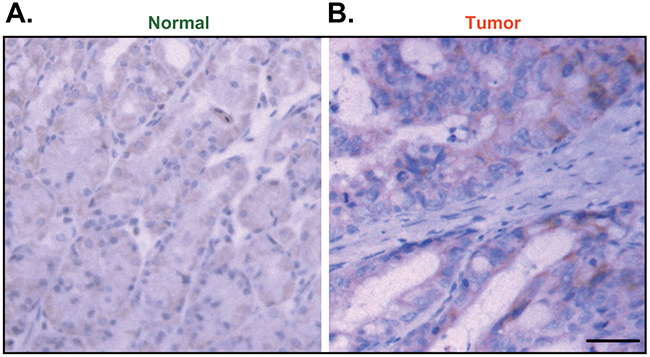
Figure 5: Immunohistochemistry (IHC) staining of Ulk1 in paraffin-embedded tissues. Representative Ulk1 IHC images in paraffin-embedded gastric cancer tissues (“Tumor”, B) and surrounding normal epithelial tissues (“Normal”, A) (Bar=100 μm).
DISCUSSION
Although sustained and intensified autophagy could promote cell death (“autophagic cell death”), minor and moderate autophagy, seen in the majority of occasions, could promote cell survival [12–15]. In those cases, cellular components are digested to provide nutrients and energies for cell survival [12–15]. This process shall help cells to survival under conditions like hypoxia and nutrient/growth factor deprivation, which are frequently happening in cancer cells [23–25].
The process of autophagy starts with formation of the double membrane vesicles (autophagosomes) in the cytoplasm, which degrade cytoplasmic materials by acidic lysosomal hydrolases [26]. Ulk1 is vital for autophagy initiation, via integrating signals from upstream sensors, including mTOR (mammalian target of rapamycin) and AMPK (AMP-activated protein kinase) [18]. For example, AMPK phosphorylates and activates Ulk1 to initiate autophagy [18, 21]. On the other hand, mTOR phosphorylates Ulk1 at different sites of AMPK, and inhibits Ulk1 activity [18, 21] to shut down autophagy. Recent studies have also proposed other mechanisms of Ulk1 in promoting cell autophagy and also mitophagy [10].
Our in vitro studies showed that knockdown of Ulk1 by targeted shRNAs inhibited AGS cell survival and proliferation, but inducing cell apoptosis. On the other hand, exogenous over-expression of Ulk1 promoted AGS cell survival and proliferation. These evidences implied that high Ulk1 expression in human gastric cancer cells should be pro-survival.
Studies testing Ulk1 expression in cancer cells have not been able to reach consistent conclusion. Pike et al., showed that transcriptional up-regulation of Ulk1 is important for cancer cell survival [27], and that loss of Ulk1 could promote necrotic cancer cell death. Same study implied that Ulk1 mRNA upregulation is associated with poor prognosis of breast cancer [27]. On the other hand, Tang et al., suggested that low Ulk1 expression is associated with progression of certain breast cancers [28]. We showed that Ulk1 (mRNA and protein) expression was significantly elevated in 12 different fresh human gastric cancer tissues. Meanwhile, Ulk1 over-expression was also noticed in several gastric cancer cell lines. IHC staining assay results further confirmed that 114 gastric cancer tissues showed high Ulk1 expression, and only 31 patients showed low Ulk1 expression. Importantly, high Ulk1 expression in gastric cancer was significantly correlated with patients’ T classification and cancer relapse. These in vitro and in vivo evidences suggest that over-expressed Ulk1 could be an important oncotarget protein for human gastric cancer. Further studies will be needed to enlarge the sample size to further support this conclusion. Patients’ survival data should also be followed.
MATERIALS AND METHODS
Cell culture
The established human gastric cancer cell lines, AGS, HGC-27, and SNU601, as well as the GES-1 gastric mucosal epithelial cells were obtained from the Cell Bank of CAS (Shanghai, China) at Dec 2013. Cells were maintained in RPMI/DMEM medium with 10% fetal bovine serum (FBS). The verification of each single cell line was described previously [29]. The reagents for cell culture were from Gibco (Shanghai, China).
Reagents and antibodies
Puromycin was provided by Sigma (Shanghai, China). All the antibodies in this study were purchased from Cell Signaling Tech (Danvers, MA).
Fresh human gastric cancer tissues
Surgery-isolated human gastric cancer tissues were washed. Tumor tissues and surrounding normal tissues were separated. A total of twelve patients with gastric cancer were enrolled (8 male, 4 female, 42-67 years old). These patients received no other therapies prior to the surgery. Fresh tissues were stored in liquid nitrogen. Tissue lysis buffer (Biyuntian, Wuxi, China) was applied to homogenate the tissues [30–33]. The protocols using human samples were approved by the Internal Review Board (IRB) and Ethics Board (EB) of authors’ institutions. The written-informed consent was obtained each and every participant. All studies were conducted according to the principles expressed in the Declaration of Helsinki and national/international guidelines.
Immunohistochemistry (IHC) staining
A total of 145 gastric cancer tissues (paraffin-embedded), collected from Dec 2012-Dec 2013, were analyzed via IHC analysis. As described [34, 35], the staining was performed on cryostat sections (4 μm) of fixed human gastric cancer tissues (or the surrounding normal tissues). Primary antibody (anti-Ulk1, 1:50) and horseradish peroxidase (HRP)-coupled secondary antibody (Santa Cruz) were added to the tissue slides. The peroxidase activity was visualized via the 3-amino-9-ethyl-carbazol (AEC) and MAYER solutions (Merck). Tissue samples with rich cells and easily observable spots were considered valid, where as those with folded slices, coloring failure, and spots without cells were considered invalid. The degree of immuno-staining was reviewed and scored independently by two observers based on the intensity of staining. Staining intensity was graded according to the following criteria: 0 (no staining), 1 (weak staining =light yellow), 2 (moderate staining= yellow brown), and 3 (strong staining= brown). Moderate and strong staining were utilized to define tumors with high Ulk1 expression, and no and weak staining were used to indicate low Ulk1 expression.
RNA extraction and real-time PCR
As reported [29, 34, 36], total RNA from cell/tissue lysates was obtained via the Trizol reagents (Invitrogen). Quantitative Real Time-PCR (“qRT-PCR”) assay was performed. The PCR reaction mixture contained SYBR Master Mix (Applied Bio-system), 0.5 μg RNA and 100 nM primers. The ABI Prism 7500 Fast Real-Time PCR system (Foster City, CA) was employed for PCR assay. The primers for Ulk1 mRNA and GAPDH mRNA were described previously [27]. Melt curve analysis was tested to analyze product melting temperature. GAPDH was always analyzed as the reference gene. The 2−ΔΔCt method was applied to quantify targeted expression change within samples [34, 36].
Western blotting assay
As previously described [29, 32, 37, 38], protein lysates (30 μg each lane) were separated by SDS-page gel (10%), and were transferred to the polyvinylidene difluoride (PVDF) membranes (Millipore). After blocking, membranes were incubated with specific primary and secondary antibodies. Enhanced chemiluminescence (ECL) reagents (Amersham, Shanghai, China) were applied for detection of the band. ImageJ software was utilized to quantify total gray of each band, and its level was normalized to Tubulin (loading).
MTT assay
Cell survival was tested by the routine 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay as described previously [35].
BrdU ELISA assay of cell proliferation
Cells with applied treatment were incubated with BrdU (10 μM, Cell Signaling Tech, Shanghai, China). BrdU incorporation was determined in the ELISA format [33]. ELISA OD was utilized as an indicator or cell proliferation.
Histone DNA ELISA assay
Quantification of cell apoptosis by the Histone DNA ELISA assay was reported in our previous studies [38, 32].
Ulk1 shRNA knockdown
The two distinct Ulk1 lentiviral shRNAs (packed in GV428 vector), targeting non-overlapping sequence of human Ulk1, were designed by the Genepharm (Shanghai, China). These two shRNAs were added directly to AGS cells (with 50-60% of confluence). After 24 hours, virus-containing medium was replaced with fresh complete medium. Stable AGS cells were then selected by puromycin (2.5 μg/mL, Sigma) for another 3 days. Expression of Ulk1 in the stable cells was verified by Western blotting assay and qRT-PCR assay.
Exogenous Ulk1 over-expression
The Ulk1 cDNA, provided again by Genepharm, was cloned into the pSuper-puro-GFP vector [29], which was transfected into AGS cells via Lipofectamine 2000 (Invitrogen). After 24 hours, cells were re-plated on selection medium containing puromycin (2.5 μg/mL, Sigma). Expression of Ulk1 (GFP-tagged) in the resulting cells was tested by Western blotting assay and qRT-PCR assay.
Statistical analyses
The SPSS software 15.0 version was used for statistical analysis. The χ2 test was applied to analyze the relationship between Ulk1 expression and patients’ clinical features. P values for all comparisons were two-tailed. Statistical significance was defined as P <0.05 for all tests.
Author contributions
All the listed authors in the study carried out the experiments, participated in the design of the study and performed the statistical analysis, conceived of the study, and participated in its design and coordination and helped to draft the manuscript.
ACKNOWLEDGMENTS
This work is supported by the National Natural Science Foundation (Grant numbers: 81472786); The Six Talents Peak Project of Jiangsu Province (Grant numbers: 2012-WSN-012); Suzhou Municipal Health Bureau projects (Grant number: LCZX201318); The Foundation of tumor clinical and basic research team (KYC005)
.CONFLICTS OF INTEREST
The authors have no conflict of interests.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer Statistics, 2017. CA Cancer J Clin. 2017; 67:7-30.
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016; 66:7-30.
3. Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, Jemal A, Yu XQ, He J. Cancer statistics in China, 2015. CA Cancer J Clin. 2016; 66:115-132.
4. Rugge M, Fassan M, Graham DY. Clinical guidelines: Secondary prevention of gastric cancer. Nat Rev Gastroenterol Hepatol. 2012; 9:128-129.
5. Wadhwa R, Song S, Lee JS, Yao Y, Wei Q, Ajani JA. Gastric cancer-molecular and clinical dimensions. Nat Rev Clin Oncol. 2013; 10:643-655.
6. Shah MA. Gastrointestinal cancer: targeted therapies in gastric cancer-the dawn of a new era. Nat Rev Clin Oncol. 2014; 11:10-11.
7. Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, Jemal A, Yu XQ, He J. Cancer statistics in China, 2015. CA Cancer J Clin. 2016.
8. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015; 65:5-29.
9. Smith K. Gastric cancer: new biologic therapy effective as second-line treatment in gastric cancer. Nat Rev Gastroenterol Hepatol. 2013; 10:694.
10. Reyjal J, Cormier K, Turcotte S. Autophagy and cell death to target cancer cells: exploiting synthetic lethality as cancer therapies. Adv Exp Med Biol. 2014; 772:167-188.
11. Nezis IP, Stenmark H. p62 at the interface of autophagy, oxidative stress signaling, and cancer. Antioxid Redox Signal. 2012; 17:786-793.
12. Li Y, Zhang J, Chen X, Liu T, He W, Chen Y, Zeng X. Molecular machinery of autophagy and its implication in cancer. Am J Med Sci. 2012; 343:155-161.
13. Kondo Y, Kanzawa T, Sawaya R, Kondo S. The role of autophagy in cancer development and response to therapy. Nat Rev Cancer. 2005; 5:726-734.
14. Janku F, McConkey DJ, Hong DS, Kurzrock R. Autophagy as a target for anticancer therapy. Nat Rev Clin Oncol. 2011; 8:528-539.
15. Amaravadi RK, Thompson CB. The roles of therapy-induced autophagy and necrosis in cancer treatment. Clin Cancer Res. 2007; 13:7271-7279.
16. Rubinsztein DC, Codogno P, Levine B. Autophagy modulation as a potential therapeutic target for diverse diseases. Nat Rev Drug Discov. 2012; 11:709-730.
17. Marino G, Niso-Santano M, Baehrecke EH, Kroemer G. Self-consumption: the interplay of autophagy and apoptosis. Nat Rev Mol Cell Biol. 2014; 15:81-94.
18. Kim J, Kundu M, Viollet B, Guan KL. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat Cell Biol. 2011; 13:132-141.
19. Khan SH, Kumar R. Role of an intrinsically disordered conformation in AMPK-mediated phosphorylation of ULK1 and regulation of autophagy. Mol Biosyst. 2012; 8:91-96.
20. Jung CH, Jun CB, Ro SH, Kim YM, Otto NM, Cao J, Kundu M, Kim DH. ULK-Atg13-FIP200 complexes mediate mTOR signaling to the autophagy machinery. Mol Biol Cell. 2009; 20:1992-2003.
21. Egan DF, Shackelford DB, Mihaylova MM, Gelino S, Kohnz RA, Mair W, Vasquez DS, Joshi A, Gwinn DM, Taylor R, Asara JM, Fitzpatrick J, Dillin A, et al. Phosphorylation of ULK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy. Science. 2011; 331:456-461.
22. Ji D, Zhang Z, Cheng L, Chang J, Wang S, Zheng B, Zheng R, Sun Z, Wang C, Liu R, Zhang X, Liu X, Wang X, et al. The combination of RAD001 and MK-2206 exerts synergistic cytotoxic effects against PTEN mutant gastric cancer cells: involvement of MAPK-dependent autophagic, but not apoptotic cell death pathway. PLoS One. 2014; 9:e85116.
23. Boya P, Gonzalez-Polo RA, Casares N, Perfettini JL, Dessen P, Larochette N, Metivier D, Meley D, Souquere S, Yoshimori T, Pierron G, Codogno P, Kroemer G. Inhibition of macroautophagy triggers apoptosis. Mol Cell Biol. 2005; 25:1025-1040.
24. Wang Q, Wang X, Hernandez A, Hellmich MR, Gatalica Z, Evers BM. Regulation of TRAIL expression by the phosphatidylinositol 3-kinase/Akt/GSK-3 pathway in human colon cancer cells. J Biol Chem. 2002; 277:36602-36610.
25. Vivanco I, Sawyers CL. The phosphatidylinositol 3-Kinase AKT pathway in human cancer. Nat Rev Cancer. 2002; 2:489-501.
26. Yart A, Mayeux P, Raynal P. Gab1, SHP-2 and other novel regulators of Ras: targets for anticancer drug discovery? Curr Cancer Drug Targets. 2003; 3:177-192.
27. Pike LR, Singleton DC, Buffa F, Abramczyk O, Phadwal K, Li JL, Simon AK, Murray JT, Harris AL. Transcriptional up-regulation of ULK1 by ATF4 contributes to cancer cell survival. Biochem J. 2013; 449:389-400.
28. Tang J, Deng R, Luo RZ, Shen GP, Cai MY, Du ZM, Jiang S, Yang MT, Fu JH, Zhu XF. Low expression of ULK1 is associated with operable breast cancer progression and is an adverse prognostic marker of survival for patients. Breast Cancer Res Treat. 2012; 134:549-560.
29. Chen MB, Zhou ZT, Yang L, Wei MX, Tang M, Ruan TY, Xu JY, Zhou XZ, Chen G, Lu PH. KU-0060648 inhibits hepatocellular carcinoma cells through DNA-PKcs-dependent and DNA-PKcs-independent mechanisms. Oncotarget. 2016; 7:17047-17059. doi: 10.18632/oncotarget.7742.
30. Zhu YR, Zhou XZ, Zhu LQ, Yao C, Fang JF, Zhou F, Deng XW, Zhang YQ. The anti-cancer activity of the mTORC1/2 dual inhibitor XL388 in preclinical osteosarcoma models. Oncotarget. 2016; 7:49527-49538. doi: 10.18632/oncotarget.10389.
31. Zhen YF, Li ST, Zhu YR, Wang XD, Zhou XZ, Zhu LQ. Identification of DNA-PKcs as a primary resistance factor of salinomycin in osteosarcoma cells. Oncotarget. 2016; 7:79417-79427. doi: 10.18632/oncotarget.12712.
32. Lu PH, Chen MB, Ji C, Li WT, Wei MX, Wu MH. Aqueous Oldenlandia diffusa extracts inhibits colorectal cancer cells via activating AMP-activated protein kinase signalings. Oncotarget. 2016; 7:45889-45900. doi: 10.18632/oncotarget.9969.
33. Li ZW, Cai S, Liu Y, Yang CL, Tian Y, Chen G, Cao C. Over-expression of Galphai3 in human glioma is required for Akt-mTOR activation and cell growth. Oncotarget. 2016 Aug 1. doi: 10.18632/oncotarget.10995. [Epub ahead of print]
34. Chen MB, Yang L, Lu PH, Fu XL, Zhang Y, Zhu YQ, Tian Y. MicroRNA-101 down-regulates sphingosine kinase 1 in colorectal cancer cells. Biochem Biophys Res Commun. 2015; 463:954-960.
35. Chen MB, Jiang Q, Liu YY, Zhang Y, He BS, Wei MX, Lu JW, Ji Y, Lu PH. C6 ceramide dramatically increases vincristine sensitivity both in vivo and in vitro, involving AMP-activated protein kinase-p53 signaling. Carcinogenesis. 2015; 36:1061-1070.
36. Chen MB, Wei MX, Han JY, Wu XY, Li C, Wang J, Shen W, Lu PH. MicroRNA-451 regulates AMPK/mTORC1 signaling and fascin1 expression in HT-29 colorectal cancer. Cell Signal. 2014; 26:102-109.
37. Zhou C, Gu J, Zhang G, Dong D, Yang Q, Chen MB, Xu D. AMPK-autophagy inhibition sensitizes icaritin-induced anti-colorectal cancer cell activity. Oncotarget. 2017; 8:14736-14747. doi: 10.18632/oncotarget.14718.
38. Lu XS, Qiao YB, Li Y, Yang B, Chen MB, Xing CG. Preclinical study of cinobufagin as a promising anti-colorectal cancer agent. Oncotarget. 2017; 8:988-998. doi: 10.18632/oncotarget.13519.