
INTRODUCTION
Fusion proteins resulting from chromosomal rearrangements are known to drive the pathogenesis of a variety of hematological neoplasms such as chronic myeloid leukemia (CML). CML is a malignant myeloproliferative disorder driven by hematopoietic stem cells that acquire a reciprocal translocation between the long arms of chromosomes 9 and 22: t(9;22)(q34;q11) [1–3]. This translocation generates the BCR-ABL oncogenic fusion protein, which harbors constitutive tyrosine kinase activity. The characteristic expression of this oncoprotein transforms the hematopoietic progenitor cells by activating downstream signaling proteins that increase cell survival and proliferation [4].
CML treatments are based predominantly on the use of tyrosine kinase inhibitors (TKI), especially Imatinib mesylate, which have been highly effective at increasing the life expectancy of CML patients in recent decades. However, CML eradication is still hindered by the emergence of TKI resistant cells [5–7]. The mechanisms of Imatinib resistance can be BCR-ABL-dependent (gene amplification or point mutations) or BCR-ABL-independent [8]. A definitive and effective therapeutic strategy is therefore still needed for these patients [9].
Recently, Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/Cas9 technology has initiated a new era of genome editing by overcoming the limitations of earlier methods [10, 11] that are inefficient, time-consuming and labor-intensive processes, hindering their general application [12, 13]. The CRISPR/Cas9 system has been widely used to generate genetic mutations in human cells [14–18] and to correct mutated genes [19–22]. The CRISPR/Cas9 system has created new possibilities for modeling diseases in vitro and in vivo, but few studies of CRISPR/Cas9 have explored the possibility of editing or correcting fusion genes resulting from chromosomal translocations [23–25].
BCR/ABL fusion, which is responsible for CML pathogenesis, is an ideal target for CRISPR-Cas9-mediated gene therapy. Moreover, the CRISPR/Cas9 system has been overwhelmingly adopted as a powerful tool for creating in vivo and in vitro models and new therapeutic opportunities for treating human diseases, including hematological conditions [26]. Thus, the present study shows a CRISPR-Cas9 application for truncating the specific BCR-ABL fusion (p210) at the genetic level in a cellular model. In addition, functional studies were performed to assess the effects of p210 on cell cycle and apoptosis. In vivo experiments were carried out to determine whether its tumorigenic capacity was also blocked by the use of mouse xenografts. To our knowledge, this is the first time that? CRISPR/Cas9 genomic editing system has been used to modify the BCR/ABL fusion gene successfully preventing its possible oncogenic effects.
RESULTS
The CRISPR/CAS9 system efficiently and specifically disrupts the human oncogene BCR/ABL fusion in an oncogene-dependent cell line
Boff-p210 is an oncogene-dependent cell line in which the expression of the human BCR/ABL oncogenic fusion confers the ability to survive and proliferate in the absence of IL-3 [27]. Cell cycle analysis of Boff-p210 cultured in the absence of IL-3 confirmed this capacity, in contrast to the Baf/3 un-manipulated parental cell line, which needs IL-3 to survive and proliferate [28]. The Boff-p210 genome was edited using CRISPR-Cas9 technology to truncate the BCR/ABL oncogene and inactivate its malignant potential.
To assess human gene editing using CRISPR/Cas9 technology in a p210 oncogene-dependent cell line, Boff-p210 cells were transduced with a lentivirus expressing a constitutive Cas9, thereby establishing the Boff-p210 Cas9 cell line. Cas9 nuclease activity was then assessed by transducing Boff-p210 Cas9 and its parental cell line with a plasmid encoding both GFP and the sgRNA against GFP [29]. After ten days, FACS analysis showed, upon transduction with this vector, an ~80% reduction in the frequency of GFP-positive cells in the active Cas9-expressing cell line compared with Boff-p210 parental cells (Figure 1A), indicating an efficient expression of active Cas9 nuclease in Boff-p210 Cas9 cells.
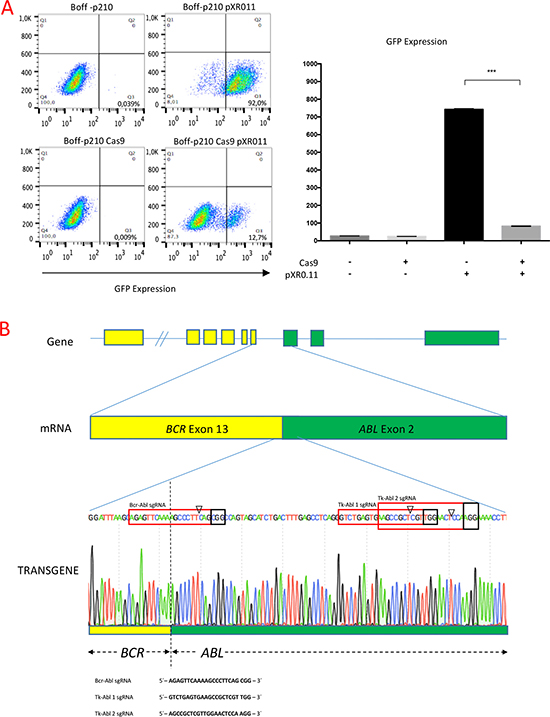
Figure 1: (A) Generation of the Boff-p210 Cas9 cell line. (Left panel) FACS plots showing the lower frequency of GFP-positive cells in Cas9-expressing cells compared with parental cells, both transduced with pXPR011. (Right panel) Quantification of GFP expression (mean ± SD). (B) Schematic representation of BCR/ABL fusion transgene and the sequences of sgRNAs for editing. Schematic representation of the BCR/ABL fusion transgene, and the sequences of three sgRNAs designed to edit BCR/ABL. One of them, Bcr-Abl sgRNA (highlighted in the red box) had 10 bp complementary to the BCR sequence and 10 bp complementary to the ABL gene, providing high specificity with the fusion gene sequence. Tk-Abl 1 and Tk-Abl 2 sgRNA were complementary to the ABL sequence. The arrowhead indicates the expected Cas9 cleavage site. PAM (highlighted in the black box) is the protospacer-adjacent motif required for Cas9 nuclease activity.
Three custom-designed single guide RNAs (sgRNAs) were used to genetically inactivate the BCR/ABL oncogene. These specific sgRNAs direct Cas9 to the BCR/ABL fusion sequence (Bcr-Abl sgRNA) or to the Abelson tyrosine kinase sequence (Tk-Abl 1 sgRNA and Tk-Abl 2 sgRNA), 40 nucleotides downstream of the fusion point (Figure 1B). Three individual lentiviral infection assays were performed with each sgRNA to generate three different Boff-p210 clones with the edited BCR/ABL oncogene, establishing three edited pools of Boff-p210 Cas9 cells.
Sanger sequencing showed the presence of indel mutations at the expected locations in all the CRISPR-Cas9 assays with each p210 sgRNA, while no changes were observed with mock sgRNA (Figure 2A). Tracking of Indels by Decomposition (TIDE) analysis identified BCR-ABL sgRNA as the most efficient sgRNA of those tested, with 85% of the Boff-p210 Cas9 cell pool edited (Bcr-Abl-EP hereafter) (Figure 2B, Table 1). Likewise, the algorithm predicted different patterns of genome repair, mainly deletions in the 11 bases adjacent to the cleavage point. The most frequently predicted mutations were an 8-bp deletion (18.5%), a 1-bp insertion (17.5%), an 11-bp deletion (10.2%) and a 1-bp deletion (9.1%) (Figure 2B, Table 1).
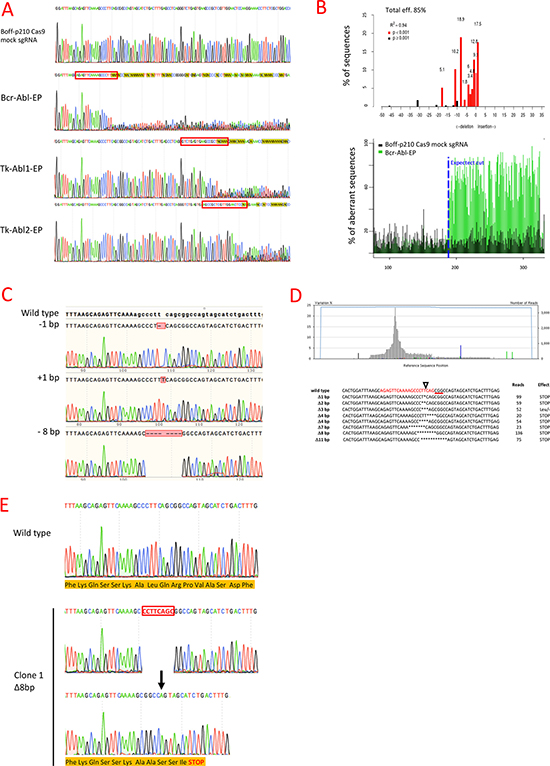
Figure 2: Genome editing of BCR/ABL in the Boff-P210 cell line. (A) Sanger sequencing of the BCR/ABL fusion region in Boff-p210 cells. The Boff-p210 cells expressing mock sgRNA, used as a control, had a wild type sequence, while cells expressing Bcr-Abl sgRNA (Bcr-Abl-EP), Tk-Abl1 sgRNA (Tk-Abl1-EP) and Tk-Abl2 sgRNA (Tk-Abl1-EP) showed a mixture of sequences around the expected Cas9 cleavage point. (B) TIDE decomposition algorithm analysis of the edited sequence in Bcr-Abl-EP cells, showing high editing efficiency at the expected cleavage point. The lower panel illustrates the aberrant sequence signal in Boff-p210 cells (black) and Boff-p210-edited cells (green) and the expected cleavage site (vertical dotted line). (C) DNA sequences of DNA post-PCR cloning colonies of Boff-p210-edited cell DNA. We compare three induced mutations with the original sequence. (D) Next-generation sequencing of the transgene target region. The upper panel shows the percentages of each base pair in the sequenced region. The lower panel shows commonly sequenced variants for CRISPR-targeted Bcr/Abl transgene in Boff-p210 cells. The sgRNA-target site is displayed in red, the PAM sequence as text underlined in red, and the cleavage site is indicated by a triangle. The number of reads per variant and the predicted effect of the mutation are shown. (E) In silico analysis of single cell-derived clone. Clone 1 showed a 8 bp deletion resulting in a stop codon and in a premature end of translation. It was selected to establish the Bcr-Abl-SC cell line.
Table 1: TIDE algorithm predicted indels induced by each sgRNA
Guide |
Edition efficiency |
TIDE Predicted Indels |
|---|---|---|
Bcr-Abl sgRNA |
85% |
+1 bp (17.5%), –1 bp (9.1%), –2 bp (4.8%) –3 bp (3.4%), –4 bp (6%), –6 bp (1.8%), –8 bp (18.9%), –11 bp (10.2%), –18 bp (5.1%) |
TK-ABL 1 sgRNA |
54,6% |
+1 bp (14.9%), –1 bp (8%) –2 bp (5.2%), –10 bp (17.6%) |
TK-ABL 2 sgRNA |
68,8% |
+1 bp (30.8%), –1 bp (5.9%), –2 bp (4.8%), –4 bp (15.2%), –14 bp (5.1%) |
Mock sgRNA |
0% |
Similar indels were obtained with the TK-Ab l 1 and TK-Abl 2 sgRNAs, although the overall predicted editing efficiencies (54.6% and 68.8%, respectively) were lower than that of Bcr-Abl sgRNA (Table 1). Thus, Bcr-Abl sgRNA was selected to confirm the indel mutations generated by the CRISPR/Cas9 system. Target DNA from the Bcr-Abl-EP cells was subcloned by PCR. Colonies were sequenced by the Sanger method to confirm that the mutations carried by the clones corresponded to the alterations predicted by the TIDE algorithm: 1-bp insertion, 1-bp or 8-bp deletions (Figure 2C). These findings suggest that these mutations are frequently generated when the cell repairs the Cas9-induced cleavage. Moreover, next-generation sequencing (NGS) was performed to sequence the target region of the Bcr-Abl-EP cells (coverage = 3386X). Several indels were detected around the expected Cas9 cleavage point (Figure 2D). The most frequent indels (> 5%) were frame-shift mutations producing a stop codon, except for the TTC-3bp-deletion (52 reads), which caused the loss of a leucine, but did not alter the downstream reading frame (Figure 2D). Therefore, most of the edited Bcr-Abl-EP cells showed mutations that produce truncated p210 oncoprotein (Figure 2D).
It is of note that no altered sequences were found in any of the top five potential off-target genes of Bcr-Abl sgRNA (Table 2), suggesting that the activity of the CRISPR/Cas9 system designed in our experimental procedures is highly specific.
Table 2: Analysis of the genomic potential off-targets of Bcr-Abl sgRNA
Gene |
Annotation |
Target |
Forward |
Reverse |
|---|---|---|---|---|
Rcor1 |
NM_198023 |
GGAAATCACAAGCCCTTCAGCAG |
ATGGCAGCTCTCCCAATCAACA |
TACAAGCACCAGGTACAATGA |
Epg5 |
NM_001195633 |
AGTGTTGAATAACCCTTCAGGGG |
CAGTAGCAGTGTTGCCCTCCA |
CACACCTTCTCTTAGTCTATGG |
Gimap7 |
NM_146167 |
AGAACTCAAAGGCCCTGCAGGAG |
CTAAGCAACATGTGGACCACT |
GGAGCTGAAGGGAGTTAGGG |
Zfp457 |
NM_001003666 |
AGTGTACAAAAGCTCTTCAGTGG |
GCGACTAAGATGGTTTCAATAG |
GCACAGCACCTCAAGGTCAG |
Alg10b |
NM_001033441 |
AGAGCCCACAACCCCTTCAGCGG |
AGTGCTCCTAAGGGGCTTAGT |
GAAGTAAGCGGCATTGCTTATAG |
Establishment of a single cell derived-cell line carriying a specific CRISPR/CAS9-mediated mutation in the BCR/ABL oncogene
To generate a clone of Boff-p210 cells with truncated BCR-ABL oncoprotein, single cells from the Bcr-Abl-EP cell line, were sorted by FACS. 41 single-cell clones were analyzed by Sanger sequencing, which revealed that most of them (32/41) had an edited sequence (Supplementary Table 1). Four of these 32 clones, showing indel mutations that generated a premature stop codon near the Cas9 cleavage point, were choose to functional analysis (Supplementary Figure 2A). One of them (Clone 1), which carried an 8-bp deletion at the expected cleavage point, was selected to establish the cell line known as Bcr-Abl-SC (single edited cell-derived Boff-p210-Cas9), whose frame-shift mutation was predicted by the TIDE algorithm and confirmed by both PCR-subcloning and NGS (Figure 2E). In silico analysis of the mutation in clone 1 resulted in the generation of a stop codon downstream of the Cas9 cleavage point, leading to the alteration of the open reading frame and the subsequently premature end of translation (Figure 2E).
Obliteration of the BCR/ABL oncogene by CRISPR-CAS9 in the BOFF-P210 cell line leads to an absence of the oncogenic protein
To confirm that the CRISPR-Cas9 system efficiently truncates BCR/ABL oncogene, the presence of the BCR-ABL oncoprotein was tested by western blot analysis. A single band, corresponding to the ABL protein (140 kDa), was detected in Baf/3 (Figure 3A), while Boff-p210 also showed a 210-kDa band corresponding to a BCR-ABL fusion. No changes were observed in either BCR-ABL or ABL expression in pools of edited cells (Figure 3A). Similar results were obtained for the TK-Abl sgRNAs cell lines (Supplementary Figure 1A). In contrast, BCR-ABL protein expression was undetectable in the Bcr-Abl-SC cells (Figure 3A, Supplementary Figure 2B).
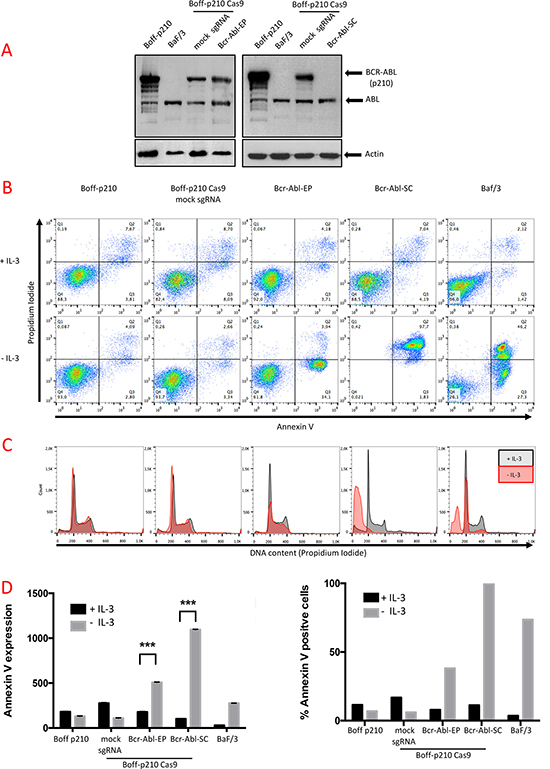
Figure 3: Functional analysis of Boff-p210-edited cells. (A) Western blot analysis of BCR-ABL expression. Abl expression (140 kDa) was observed in all cells. A 210 kDa band, corresponding BCR-ABL expression was observed in unedited Boff-p210 cells (parental and sgRNA mock controls) and in Bcr-Abl-EP cells. In contrast, Bcr-Abl-SC and Baf/3 cells showed no expression of BCR-ABL. (B) Annexin V/propidium iodide labeling of Boff-p210 cells after four days in culture in the presence or absence of IL-3. Unedited and mock sgRNA-expressing cells showed IL-3-independent growth. Baf/3 as Bcr-Abl-EP cells showed greater annexin V labeling after IL-3 withdrawal. This effect was stronger in cell lines derived from a single-edited cell. (C) Flow cytometric analysis of Bcr-Abl-EP cells. No changes were observed in the cell cycle phases of Boff-p210 or the mock cells, grown with or without IL-3. High frequencies of cells in the subG0 phase were found in the single edit derived-cells cultured without IL-3, as in Baf/3 cells. (D) Quantification of annexin V labeling (Left graph, represented as mean ± SEM; ***p < 0.001) and percentage of Boff-p210-positive cells (right graph).
In vitro ablation of BCR/ABL by the CRISPR/CAS9 system in tumorigenic p210 cells induces cell cycle arrest and impairs survival
To study the consequences of in vitro obliteration of the BCR/ABL oncogene by the CRISPR-Cas9 system, the processes of cell-cycle progression and apoptosis were analyzed in Boff-p210-edited cells in the presence or absence of IL-3 for 4 days.
Annexin V and PI labeling were used to measure apoptosis in Boff-p210 cells by flow cytometry after 4 days in culture in the presence or absence of IL-3 (Figure 3B). We simultaneously analyzed the distribution of the cells in the different cell cycle phases by permeabilization followed by PI staining (Figure 3C). Non-tumorigenic Baf/3 cells rapidly died after 4 days in IL-3-withdrawn medium (Figure 3A). After 4 days of IL-3 deprivation, 73.3% of the Baf/f3 cells showed annexin V staining (Figure 3B, 3D), and there was clear evidence of oligonucleosomal DNA degradation (subG0 DNA content), characteristic of apoptosis (Figure 3C). In contrast, only 10–20% of the Boff-p210 or Boff-p210 Cas9 mock sgRNA cells, growing with or without IL-3, were positive for annexin V staining (Figure 3B, 3D). Similarly, no differences in the distribution of the cell cycle phases were observed in Boff-p210 and mock cells in the presence or absence of IL-3 (Figure 3C), with no evidence of cells in sub-G0. However, withdrawal of IL-3 in Bcr-Abl-EP cell cultures resulted in increased annexin V staining (7.89% in medium with IL-3; 38.04% in the absence of IL3), indicating that edited cells are IL3-dependent (Figure 3B, 3D). Accordingly, we observed a decrease in the G1, S and G2/M phases of the cell cycle in Bcr-Abl-EP cells in the absence of IL-3, while the presence of IL-3 maintained the cell viability of these cells. Boff-p210 and non-edited Boff-p210 cells (mock sgRNA assay) yielded similar results in the distribution of the cell cycle phases with or without IL3-conditioned medium (Figure 3C). A similar effect was observed in TK-ABL1-EP and TK-ABL2-EP cells in the absence of IL-3 (Supplementary Figure 1B).
To confirm that the loss of ability to grow in the absence of IL-3 observed in the pool of edited cells was due solely to the disruption of the BCR/ABL transgene, we assessed the effect of IL-3 withdrawal in an edited cell line derived from a single cell (Bcr-Abl-SC cells). Like control mock and Bcr-Abl-EP cells, Bcr-Abl-SC cells showed low expression of annexin V (11.23% of cells) and high viability in the presence of IL-3 (Figure 3B, 3C, 3D). However, after 4 days in IL-3-deprived culture, most cells (97.7%) showed a high level of annexin V staining (Figure 3B, 3D), accompanied by fragmented DNA (Figure 3C). The dramatic effect observed in the Bcr-Abl-SC cell line, similar to that of Baf/3 cells, suggests that there is a complete lack of BCR-ABL functional oncoprotein. These results were confirmed in three clones derived from a single-edited cell carrying a truncated BCR/ABL sequence in which IL-3 withdrawal produced a similar drastic effect (Supplementary Figure 2C).
CRISPR-CAS9 impairs the tumorigenic capacity of edited Boff-p210 selected cells
In order to determine the effects of Bcr/Abl oncogene disruption in vivo, the flanks of CB17C SCID mice were subcutaneously injected with Baf/3, Boff-p210, Bcr-Abl-EP cells, and Boff-p210 Cas9 mock sgRNA (Boff-p210 mock) cells. Mice injected with Boff-p210 cells and Boff-p210 mock cells, developed similar detectable tumors (mean mass: 663 mg and 445 mg respectively). In contrast, mice injected with Bcr-Abl-EP cells gave rise to significantly smaller (around 60%) subcutaneous tumors than those produced by the non-edited cells (mean mass: 172 mg; Figure 4A and 4B). As expected, Baf/3 cells showed no tumor growth in this period.
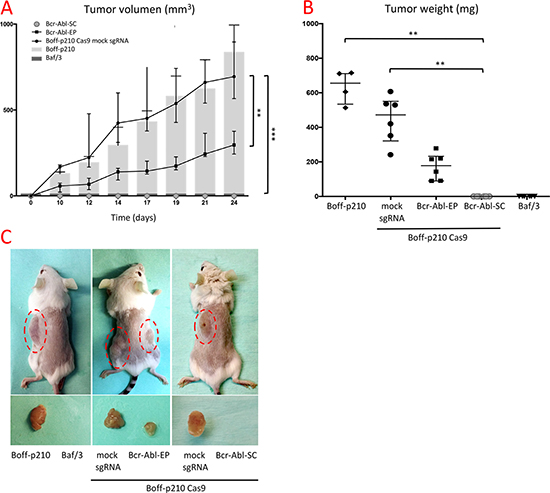
Figure 4: In vivo effects of CRISPR-mediated editing of the BCR/ABL oncogene. (A) Tumor growth (mm3) over the 24 days following subcutaneous cell injection. Similar tumor growth was observed in Boff-p210 (grey bars) and Boff-p210 Cas9 mock sgRNA (black dots line) injected cells. Tumors formed by Bcr-Abl-EP cells (black squares line) were half the size of those induced previously. Tumor growth was observed when the single edited cell-derived cells were injected (grey dotted line), as well Baf/3 cells (dark grey bars). The plot shows medians and ranges; ***p < 0.001). (B) After 24 days, mice were sacrificed and their tumor mass measured. The final tumor mass was reduced by half in the case of the edited pool cells (black squares), relative to controls (black dots). Bcr-Abl-SC (grey dots) and Baf/3 (black triangles) cells were unable to form a subcutaneous tumor. The plot shows medians and ranges; **p < 0.05). (C) External appearance of mice and developed tumors 24 days after subcutaneous cell injection.
Bcr-Abl-SC cells carrying an 8-bp deletion (clone 1, Supplementary Figure 2) were selected to test their tumorigenic capacity by injection into the flank of CB17C SCID mice. Non-edited Boff-p210 cells and Baf/3 cells were also injected as controls. 24 days post-injection, tumor growth was clearly detected in mice injected with non-edited Boff-p210 cells. Conversely, no tumor growth was observed in mice injected with cells derived from the single-edited cell line, or with Baf/3 cells (Figure 4A).
DISCUSSION
In this study, we have explored the ability use of the CRISPR/Cas9 technology to obliterate BCR-ABL fusion in order to determine its impact on the leukemic processes in in vitro and in xenograft models of CML. The CRISPR/Cas9 system has recently emerged as a powerful genome editing tool [14, 30]. Cas9 nuclease from Streptococcus pyogenes can be guided by means of simple base-pair complementarity between the first 20 nucleotides of an engineered sgRNA and a target genomic DNA sequence of interest [31, 32]. Recently, this technology has been used for gene correction of point mutations in cells [20, 21, 33]. However, to our knowledge, this is the first study in which CRISPR/Cas9 has been used to prevent the tumorigenic ability of an oncogenic fusion, such BCR/ABL, by inducing frame-shift mutations.
It is well known that the crucial genetic event in CML development is the generation of a t(9;22)(q34;q11) reciprocal chromosomal translocation [34], resulting mainly in the generation of the BCR-ABL (p210) oncoprotein with a constitutive kinase activity [35–37]. Herein, the tumorigenic Boff-p210 cell line is used as a cellular model, which is particularly suitable for BCR-ABL assays. This is an interleukin 3- (IL-3) independent cell line derived from the Baf/3 hematopoietic cell line [38]. Boff-p210 expresses the BCR-ABLp210 oncogene as a tetracycline-regulated transgene and can be genetically modified, expanded and then re-administered into immunocompromised mice. In the absence of tetracycline or doxycycline, BCR/ABL expression is constitutive, and confers on Boff-210 cells the ability to survive and proliferate in the absence of IL-3 [39].
A Boff-p210 cell line expressing constitutively active Cas9 was generated to genetically inactivate human BCR/ABL oncogenes, thereby to avoid its pathological effect on the Boff-p210 cell line. Three specific sgRNAs against the BCR/ABL fusion gene were designed as RNA guides for Cas9. One of them drives Cas9 nuclease against the BCR/ABL junction sequence and the other two drive it against the ABL tyrosine kinase sequence. The CRISPR-Cas9 system was highly effective at inducing indels in their target sequences with all the sgRNAs used. Notably, Bcr-Abl sgRNA was the most efficient, involving up to 80% of the edited cells.
Since CRISPR/Cas9 nuclease can induce off-target cleavages, due to unspecific recognition of non-target sequences [40], the integrity of the main off-target sequences was tested. It is worth pointing out that no off-target sequences were observed in any of the edited pools of cells. These findings suggest that the CRISPR/Cas9 system is highly specific, as has been demonstrated by earlier whole-genome sequencing studies [41–43].
The effects of the constitutive BCR/ABL activity are pleiotropic and promote leukemogenesis by acquisition of tumor abilities. These abilities include the increase of cell survival [4], the avoidance of apoptosis [39, 44, 45] and the promotion of genomic instability by downregulation of DNA-repair mechanisms [46, 47]. In this cellular model, the expression of BCR-ABL oncoprotein induces IL-3-independent proliferation and survival, while Baf/3 cells require IL-3 to survive and proliferate (Figure 3B–3D). To study the oncogene-disruption effect in this cellular model, CRISPR-Cas9 edition assays were performed. The CRISPR-Cas9 system efficiently induced different mutations at the expected cleavage expected point, giving rise to a cellular pool with several altered BCR/ABL sequences. A significant increase in cell death was observed in Bcr-Abl-EP cells in the absence of IL- 3, compared with Boff-p210 parental cells or mock Boff-p210 cells. However, a massive cell death was not observed in Bcr-Abl-EP cells, indicating the presence of edited cells with survival capacity in the absence of IL-3. This finding was corroborated by the fact that Bcr-Abl-EP did not show any significant variations in the level of p210 expression as shown by western blot analysis. As confirmed by NGS, the CRISPR-Cas9 system induced several different indel mutations in our edited cell lines. Some of them were frame-shift mutations that resulted in truncated proteins, whereas others generated a gain/loss of amino acids that gave rise to a functional protein. These functional repaired p210 proteins could protect cells from apoptosis induced by IL-3 withdrawal, promoting the cell survival of several clones. This result is consistent with previous reports about the antiapoptotic effect of the BCR/ABL [44, 45]. To avoid this drawback, a single cell with a specific edited sequence of BCR/ABL was isolated from the edited pool. In particular, a specific single-cell clone with an 8-bp deletion at the cleavage point was selected for further experiments. The selected cell clone had an edited p210 sequence, which gave rise to a premature translation end. In fact, no BCR-ABL oncoprotein expression was observed by western blot. Interestingly, this single edited cell-derived clone showed the highest level of apoptosis (more than 97% of cells) when it grew in the absence of IL-3, compared with that observed in cells that did not express p210.
To further test the tumorigenic capability of the BCR-ABL-edited cells, the xenograft model was generated by injecting these edited cells into mice. In all cases, the pool of Boff-p210-edited cells gave rise to a subcutaneous tumor 24 days post-injection. These findings indicate that cells with functional BCR-ABL oncoproteins maintain their tumorigenic capabilities. In contrast, cell clones derived from a single-edited cell were unable to induce tumors in all cases. This result demonstrates that CRISPR-Cas9 could efficiently prevent the tumorigenic potential of the p210 oncoprotein.
Despite major improvements in therapeutic strategies in patients with CML in recent decades, as a consequence of the use of tyrosine kinase inhibitors, eradication of CML remains a challenge because of the emergence of drug resistance. In the context of stem cell transplantation [48], the use of CRISPR-Cas9 technology could be a promising new therapeutic option for CML patients who have developed tyrosine-kinase resistance. Bone marrow leukemic stem cells could be edited by CRISPR-Cas9 technology to truncate BCRL/ABL fusions, and truncated cells specifically selected before infusion. However, the fact that of sgRNAs can tolerate certain mismatches and that Cas9 is integrated and constantly produced represent the key limit of this technology. The use of integrase-deficient lentivirus vectors or Cas9-guide RNA ribonucleoprotein (RNP) complexes into primary cells, could represent a safety advantage in human therapy [49–51]. Further studies with human primary cells should be performed to test this hypothesis.
Our study is the first step towards providing proof-of-principle for genome editing of the main genetic event in CML patients. In summary, this is the first report describing the use of CRISPR/Cas9 genome editing to completely truncate human BCR-ABL oncoprotein fusion in order to prevent its tumorigenic abilities in an in vitro as well as in an in vivo CML model.
MATERIALS AND METHODS
Cell lines and culture conditions
Boff-p210 is a murine interleukin 3 (IL3) -independent cell line derived from the hematopoietic cell line Baf/3 [38] that expresses BCR/ABL as a tetracycline-regulated transgene (BCR/ABL expression is constitutive in the absence of tetracycline or doxycycline) [52–54]. Boff-p210 was maintained in Dulbecco’s Modified Eagle’s Medium (DMEM) (Life Technologies) supplemented with 10% fetal bovine serum (FBS) and 1% of penicillin/streptomycin (Life Technologies) in 5% CO2 at 37°C. IL-3-dependent Baf/3 cells, used as a parental cell line, were grown in the same medium supplemented with 10% WEHI-3-conditioned medium as a source of IL-3.
HEK 293T cells were used for lentivirus production. They were maintained in RPMI 1640 (Life Technologies) supplemented with 10% FBS and 1% penicillin/streptomycin (Life Technologies).
Lentiviral constructs, sgRNA design and sgRNA cloning
The constitutive Cas9 expression vector (plasmid LentiCas9-Blast, a gift from Feng Zhang, Addgene plasmid # 52962) [55] was used to generate the Boff-p210 cell line with stable Cas9 expression.
pLKO5.sgRNA.EFS.GFP (Addgene plasmid # 57822) [56] containing the coding sequence of GFP and a cloning site for sgRNA sequence was digested with Esp3I (BsmBI) (NEB). To clone the sgRNAs into the pLKO5 vector, two complementary oligos for each sgRNA were designed including two 4bp-overhang sequences (Table 3). The sgRNAs sequences were designed with the Broad Institute CRISPR designs software (http://www.broadinstitute.org/rnai/public/analysis-tools/sgrna-design). One sgRNA (Bcr-Abl sgRNA) was designed to specifically target the fusion junction sequence. To control for variability in targeting efficiency and potential off-targets effects, two sgRNAs were designed to target the ABL tyrosine kinase sequence (Tk-Abl1 sgRNA and Tk-Abl2 sgRNA) (Figure 1B). An sgRNA designed not to target the genome was used as a negative control (mock). The two complementary oligos were denatured at 95°C for 5 min, ramp cooled to 25°C over a period of 45 min to allow annealing, and finally ligated with the linearized pLKO5. Competent cells were transformed with 2 μl of the ligated plasmid, and single colonies were expanded before to plasmid extraction using a QIAprep Spin Maxiprep kit (Qiagen). The correct insertion of the sgRNA sequences was confirmed by using Sanger sequencing.
Table 3: Oligos designed for each sgRNA
Forward |
Reverse |
|
|---|---|---|
Bcr-Abl |
CACCGAGAGTTCAAAAGCCCTTCAG |
AAACCTGAAGGGCTTTTGAACTCTC |
TK-Abl 1 |
CACCGAGCCGCTCGTTGGAACTCCA |
AAACTGGAGTTCCAACGAGCGGCTC |
TK-Abl 2 |
CACCGGTCTGAGTGAAGCCGCTCGT |
AAACACGAGCGGCTTCACTCAGACC |
mock |
CACCGACGGAGGCTAAGCGTCGCAA |
AAACTTGCGACGCTTAGCCTCCGTC |
Cas9 activity was tested using a previously reported system based on the pXPR-011 plasmid (a gift from John Doench and David Root; Addgene plasmid #59702) which delivers GFP and sgRNA targeting GFP [29].
Lentiviral production and cell transduction
Lentiviral particles were produced by transient transfection of HEK 293T cells using Lipofectamine 2000® (Life Technologies). Viral constructs were co-transfected with pMD2.G (Addgene plasmid 12259) and psPAX2 (Addgene plasmid 12260) (both kindly provided by Didier Trono, EPFL, Lausanne, Switzerland). Lentiviral particles were collected at 24 and 48 h, and, finally, were concentrated using Lenti-X concentrator® (Clontech). Cells were infected in 24-well plate format, with each well containing 2 × 105 cells, cultured in media supplemented with 4 μg/mL polybrene.
Cas9 activity assay system
To obtain stable Boff-p210 cells expressing Cas9, lentiviral particles containing LentiCas9-Blast, plasmid-expressing Cas9 and blasticidine resistance (a gift from Feng Zhang, Addgene plasmid # 52962) [55], were transduced into Boff-p210 cells and selected by blasticidine (10 μg/μl), over ten days.
Cas9 activity was assessed using the Cas9-activity vector as previously reported [29]. The Cas9-expressing Boff-p210 cell line and its corresponding parental Boff-p210 cell line were transduced with plasmid pXPR-011, encoding both GFP and sgRNA against GFP (a gift from John Doench and David Root; Addgene plasmid # 59702) [29]. 48 h post-infection, 2 μg/mL puromycin was added and cells were selected for three days. GFP-positive cells from both cell lines were analyzed by BD FACScalibur ten days post-infection.
Sequencing of sgRNA targets sites
Genomic DNA was extracted using the QIAamp DNA Micro Kit (Qiagen) following the manufacturer’s protocol. To amplify the region of the BCR-ABL fusion, PCR was performed using the following primers: forward 5′- TCGTGTGTGAAACTCCAGACTGTC – 3′ and reverse 5′- TTGGGCTTCACACCATTCCCC – 3′. PCR products were purified using a High Pure PCR Product Purification Kit (Roche) and were sequenced by the Sanger method using each forward and reverse PCR primers.
The editing efficiency of the sgRNAs and the potential induced mutations were assessed using Tracking of Indels by Decomposition (TIDE) software (https://tide-calculator.nki.nl; Netherlands Cancer Institute), which only required two Sanger sequencing runs from wild-type cells and mutated cells.
To identify specifically the different generated mutations, total DNA of Cas9-edited cells were PCR-amplified, subcloned and transformed in bacteria. DNA from single clones were extracted with a QIAprep Spin Miniprep Kit (Qiagen) and sequenced by Sanger sequencing.
In parallel, Next Generation Sequencing (NGS) technology was employed with the same Sanger primers with the corresponding adapters added, to read each edited sequence individually. The amplicon libraries were sequenced on the GS Junior platform (454 Life Sciences, Roche, Branford, CT, USA) [57].
Western blot
BCR-ABL protein expression was assessed by SDS-PAGE and western blotting using a mouse anti-ABL antibody (1:1000; SC-23, Santa Cruz). Horseradish peroxidase-conjugated α-mouse antibody (1:10000; NA931V, GE Healthcare) was used as a secondary antibody. Antibodies were detected using ECLTM Western Blotting Detection Reagents (RPN2209, GE Healthcare).
Flow cytometry analysis and cell sorting for Cas9-mediated editing and for isolating single edited cell-derived clone
72 hours after infection by lentivirus-expressing sgRNAs with GFP of Boff-p210 Cas9 cell lines, GFP-positive cells were selected by fluorescence-activated cell sorting (FACS) using FACS Aria (BD Biosciences), establishing the cell lines known as Bcr-Abl-EP, Tk-Abl1-EP and Tk-Abl2-EP. Single-cells were seeded in 96-well plate by FACS, establishing four clones with frame-shift mutations that disrupt the BCR/ABL sequence. Clone 1 was chosen to establish the cell line known as Bcr-Abl-SC.
Off-target sequence analysis
To investigate off-target cleavage potentially generated by Cas9 in Bcr-Abl-EP and Bcr-Abl-SC cells, the top five off-target sites (obtained from the crispr.mit.edu website) were analyzed by PCR and Sanger sequencing, just before to start the functional and xenograft experiments. The primers used to amplify the top five predicted off-targets regions are described in Table 2. PCR products were purified using a High Pure PCR Product Purification Kit (Roche) and were sequenced using each forward and reverse PCR primers.
Apoptosis and cell cycle analysis.
Apoptosis was measured by flow cytometry with an annexin V-Dy634 apoptosis detection kit (ANXVVKDY, Immunostep) following the manufacturer’s instructions. Briefly, 5 × 105 cells were collected and washed twice in PBS, and labeled with annexin V-DY-634 and non-vital dye propidium iodide (PI), allowing the discrimination of living-intact cells (annexin-negative, PI-negative), early apoptotic cells (annexin-positive, PI-negative) and late apoptotic or necrotic cells (annexin-positive, PI-positive). In parallel, cell distribution in the cell cycle phase was also analyzed by measuring DNA content (PI labeling after cell permeabilization).
Mouse xenograft tumorigenesis
18 four- to five-week-old female CB.17 SCID mice (Charles River, Barcelona, Spain) were used (six mice per group). Tumor xenografts were induced by subcutaneous injection of cell suspensions containing 8 × 106 cells in 0.2 ml of cellular medium into the mouse flank.
Cells were counted using a Neubauer chamber (VWR) and cellular viability monitored by trypan blue staining (Sigma). This study followed the Spanish and European Union guidelines for animal experimentation (RD 1201/05, RD 53/2013 and 86/609/CEE, respectively). The study received prior approval from the Bioethics Committee of our institution.
In the first mouse group, Boff-p210 Cas9 mock sgRNA cells were injected in the left flank and Bcr-Abl-EP cells in the right flank; in the second group, Boff-p210Cas9 mock sgRNA cells were injected in the left flank and Bcr-Abl-SC cells in the right flank; and, finally, in the third group (controls), Boff-p210 cells were injected in the left flank and Baf/3 in the right flank. Tumor diameters were measured every 2–3 days with a caliper. Tumor volume was calculated as described elsewhere [58] by the formula a2bπ/6 (a and b being, respectively, the smallest and the biggest diameters). Mice were sacrificed by anesthesia overdose 24 days after cell injection, upon which the tumors were collected and weighted.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 6 Software (GraphPad Software). Differences in annexin V labeling were tested using one-way ANOVA and Tukey’s multiple comparisons test. Differences in tumor volume over time were tested by two-way ANOVA, and Tukey’s multiple comparisons test. Differences in median tumor masses were tested by the Kruskal-Wallis and Mann-Whitney U tests, and Dunn’s multiple comparisons test. Statistical significance at values of p < 0.05 (**) and p < 0.001 (***) was noted.
Abbreviations
CML: Chronic myeloid leukemia; CRISPR: Clustered regularly interspaced short palindromic repeats; sgRNA: Single-guide RNA; EP: Edited pool; SC: Single edited cell-derived.
Authors’ contributions
IGT and MHS designed the CRISPR system and the cell genome editing experiments. IGT and JLO carried out the xenograft experiments. VAP and MQ cloned the sgRNAs and produced the viral plasmids. RB and JMHR helped interpret the results and write the manuscript. CG provided the cell lines and helped interpret the results and write the manuscript. MSM designed all experiments and interpreted the results. IGT and MSM wrote the paper in conjunction with the other authors.
ACKNOWLEDGMENTS
The authors wish to express their sincere thanks to Dr. Martin-Zanca for the scientific advice and useful help. We offer our profound thanks to Dr. Catherine Wu and and Dr. Michaela Gruber, extendible to all members of Wu’s lab from Dana-Farber Cancer Institute, for all the support and their scientific advice in the experiments with CRISPR/Cas9 system. We are also grateful to Lucía Méndez, Irene Rodríguez, Sara González, Teresa Prieto, María Ángeles Ramos, Almudena Martín, Ana Díaz, Ana Simón, María del Pozo, Isabel M Isidro, Vanesa Gutiérrez, Sandra Pujante and Sandra Santos of the Centro de Investigación del Cáncer, Salamanca, Spain for their technical assistance
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
FUNDING
This work was supported in part by a grant from the Consejería de Educación, Junta de Castilla y León, Fondos FEDER (SA085U16 to JMHR and JCYL-EDU/346/2013 PhD scholarship to MHS), the ISCIII-FEDER Spanish Cancer Network (RD12/0036/0069) and the Fondo de Investigaciones Sanitarias (FIS) of the Spanish Ministry of Economy and Competitiveness and the European Regional Development Fund (ERDF) “Una manera de hacer Europa” (grant PI15/01471).
REFERENCES
1. Faderl S, Talpaz M, Estrov Z, O’Brien S, Kurzrock R, Kantarjian HM. The biology of chronic myeloid leukemia. The New England journal of medicine. 1999; 341:164–172.
2. Deininger MW, Goldman JM, Melo JV. The molecular biology of chronic myeloid leukemia. Blood. 2000; 96:3343–3356.
3. Kurzrock R, Kantarjian HM, Druker BJ, Talpaz M. Philadelphia chromosome-positive leukemias: from basic mechanisms to molecular therapeutics. Annals of internal medicine. 2003; 138:819–830.
4. Colicelli J. ABL tyrosine kinases: evolution of function, regulation, and specificity. Sci Signal. 2010; 3:re6.
5. Hughes TP, Saglio G, Quintas-Cardama A, Mauro MJ, Kim DW, Lipton JH, Bradley-Garelik MB, Ukropec J, Hochhaus A. BCR-ABL1 mutation development during first-line treatment with dasatinib or imatinib for chronic myeloid leukemia in chronic phase. Leukemia. 2015; 29:1832–1838.
6. Gorre ME, Mohammed M, Ellwood K, Hsu N, Paquette R, Rao PN, Sawyers CL. Clinical resistance to STI-571 cancer therapy caused by BCR-ABL gene mutation or amplification. Science. 2001; 293:876–880.
7. O’Hare T, Eide CA, Deininger MW. Bcr-Abl kinase domain mutations, drug resistance, and the road to a cure for chronic myeloid leukemia. Blood. 2007; 110:2242–2249.
8. Bhamidipati PK, Kantarjian H, Cortes J, Cornelison AM, Jabbour E. Management of imatinib-resistant patients with chronic myeloid leukemia. Ther Adv Hematol. 2013; 4:103–117.
9. Jabbour E, Lipton JH. A critical review of trials of first-line BCR-ABL inhibitor treatment in patients with newly diagnosed chronic myeloid leukemia in chronic phase. Clin Lymphoma Myeloma Leuk. 2013; 13:646–656.
10. Gaj T, Mercer AC, Sirk SJ, Smith HL, Barbas CF 3rd. A comprehensive approach to zinc-finger recombinase customization enables genomic targeting in human cells. Nucleic Acids Res. 2013; 41:3937–3946.
11. Mercer AC, Gaj T, Fuller RP, Barbas CF 3rd. Chimeric TALE recombinases with programmable DNA sequence specificity. Nucleic Acids Res. 2012; 40:11163–11172.
12. Gaj T, Gersbach CA, Barbas CF 3rd. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013; 31:397–405.
13. Nemudryi AA, Valetdinova KR, Medvedev SP, Zakian SM. TALEN and CRISPR/Cas Genome Editing Systems: Tools of Discovery. Acta Naturae. 2014; 6:19–40.
14. Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, Hsu PD, Wu X, Jiang W, Marraffini LA, Zhang F. Multiplex genome engineering using CRISPR/Cas systems. Science. 2013; 339:819–823.
15. Hsu PD, Lander ES, Zhang F. Development and applications of CRISPR-Cas9 for genome engineering. Cell. 2014; 157:1262–1278.
16. Niu J, Zhang B, Chen H. Applications of TALENs and CRISPR/Cas9 in human cells and their potentials for gene therapy. Mol Biotechnol. 2014; 56:681–688.
17. Konermann S, Brigham MD, Trevino AE, Joung J, Abudayyeh OO, Barcena C, Hsu PD, Habib N, Gootenberg JS, Nishimasu H, Nureki O, Zhang F. Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature. 2015; 517:583–588.
18. Shalem O, Sanjana NE, Hartenian E, Shi X, Scott DA, Mikkelsen TS, Heckl D, Ebert BL, Root DE, Doench JG, Zhang F. Genome-scale CRISPR-Cas9 knockout screening in human cells. Science. 2014; 343:84–87.
19. Schwank G, Koo BK, Sasselli V, Dekkers JF, Heo I, Demircan T, Sasaki N, Boymans S, Cuppen E, van der Ent CK, Nieuwenhuis EE, Beekman JM, Clevers H. Functional repair of CFTR by CRISPR/Cas9 in intestinal stem cell organoids of cystic fibrosis patients. Cell Stem Cell. 2013; 13:653–658.
20. Valletta S, Dolatshad H, Bartenstein M, Yip BH, Bello E, Gordon S, Yu Y, Shaw J, Roy S, Scifo L, Schuh A, Pellagatti A, Fulga TA, et al. ASXL1 mutation correction by CRISPR/Cas9 restores gene function in leukemia cells and increases survival in mouse xenografts. Oncotarget. 2015; 6:44061–44071. doi: 10.18632/oncotarget.6392.
21. Hoban MD, Lumaquin D, Kuo CY, Romero Z, Long J, Ho M, Young CS, Mojadidi M, Fitz-Gibbon S, Cooper AR, Lill GR, Urbinati F, Campo-Fernandez B, et al. CRISPR/Cas9-Mediated Correction of the Sickle Mutation in Human CD34+ cells. Mol Ther. 2016; 24:1561–1569.
22. Guan Y, Ma Y, Li Q, Sun Z, Ma L, Wu L, Wang L, Zeng L, Shao Y, Chen Y, Ma N, Lu W, Hu K, et al. CRISPR/Cas9-mediated somatic correction of a novel coagulator factor IX gene mutation ameliorates hemophilia in mouse. EMBO Mol Med. 2016; 8:477–488.
23. Torres R, Martin MC, Garcia A, Cigudosa JC, Ramirez JC, Rodriguez-Perales S. Engineering human tumour-associated chromosomal translocations with the RNA-guided CRISPR-Cas9 system. Nat Commun. 2014; 5:3964.
24. Vanden Bempt M, Demeyer S, Mentens N, Geerdens E, De Bock CE, Wlodarska I, Cools J. Generation of the Fip1l1-Pdgfra fusion gene using CRISPR/Cas genome editing. Leukemia. 2016.
25. Maddalo D, Manchado E, Concepcion CP, Bonetti C, Vidigal JA, Han YC, Ogrodowski P, Crippa A, Rekhtman N, de Stanchina E, Lowe SW, Ventura A. In vivo engineering of oncogenic chromosomal rearrangements with the CRISPR/Cas9 system. Nature. 2014; 516:423–427.
26. Zhang H, McCarty N. CRISPR-Cas9 technology and its application in haematological disorders. Br J Haematol. 2016; 175:208–225.
27. Gutierrez-Berzal J, Castellano E, Martin-Encabo S, Gutierrez-Cianca N, Hernandez JM, Santos E, Guerrero C. Characterization of p87C3G, a novel, truncated C3G isoform that is overexpressed in chronic myeloid leukemia and interacts with Bcr-Abl. Experimental cell research. 2006; 312:938–948.
28. Palacios R, Henson G, Steinmetz M, McKearn JP. Interleukin-3 supports growth of mouse pre-B-cell clones in vitro. Nature. 1984; 309:126–131.
29. Doench JG, Hartenian E, Graham DB, Tothova Z, Hegde M, Smith I, Sullender M, Ebert BL, Xavier RJ, Root DE. Rational design of highly active sgRNAs for CRISPR-Cas9-mediated gene inactivation. Nat Biotechnol. 2014; 32:1262–1267.
30. Chylinski K, Le Rhun A, Charpentier E. The tracrRNA and Cas9 families of type II CRISPR-Cas immunity systems. RNA Biol. 2013; 10:726–737.
31. Mojica FJ, Diez-Villasenor C, Garcia-Martinez J, Almendros C. Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology. 2009; 155:733–740.
32. Deveau H, Barrangou R, Garneau JE, Labonte J, Fremaux C, Boyaval P, Romero DA, Horvath P, Moineau S. Phage response to CRISPR-encoded resistance in Streptococcus thermophilus. J Bacteriol. 2008; 190:1390–1400.
33. Dever DP, Bak RO, Reinisch A, Camarena J, Washington G, Nicolas CE, Pavel-Dinu M, Saxena N, Wilkens AB, Mantri S, Uchida N, Hendel A, Narla A, et al. CRISPR/Cas9 beta-globin gene targeting in human haematopoietic stem cells. Nature. 2016; 539:384–389.
34. Nowell PC. The minute chromosome (Phl) in chronic granulocytic leukemia. Blut. 1962; 8:65–66.
35. Rowley JD. Letter: A new consistent chromosomal abnormality in chronic myelogenous leukaemia identified by quinacrine fluorescence and Giemsa staining. Nature. 1973; 243:290–293.
36. Melo JV. The diversity of BCR-ABL fusion proteins and their relationship to leukemia phenotype. Blood. 1996; 88:2375–2384.
37. Ben-Neriah Y, Daley GQ, Mes-Masson AM, Witte ON, Baltimore D. The chronic myelogenous leukemia-specific P210 protein is the product of the bcr/abl hybrid gene. Science. 1986; 233:212–214.
38. Palacios R, Steinmetz M. Il-3-dependent mouse clones that express B-220 surface antigen, contain Ig genes in germ-line configuration, and generate B lymphocytes in vivo. Cell. 1985; 41:727–734.
39. Bedi A, Zehnbauer BA, Barber JP, Sharkis SJ, Jones RJ. Inhibition of apoptosis by BCR-ABL in chronic myeloid leukemia. Blood. 1994; 83:2038–2044.
40. Yee JK. Off-target effects of engineered nucleases. FEBS J. 2016; 283:3239–3248.
41. Smith C, Gore A, Yan W, Abalde-Atristain L, Li Z, He C, Wang Y, Brodsky RA, Zhang K, Cheng L, Ye Z. Whole-genome sequencing analysis reveals high specificity of CRISPR/Cas9 and TALEN-based genome editing in human iPSCs. Cell Stem Cell. 2014; 15:12–13.
42. Suzuki K, Yu C, Qu J, Li M, Yao X, Yuan T, Goebl A, Tang S, Ren R, Aizawa E, Zhang F, Xu X, Soligalla RD, et al. Targeted gene correction minimally impacts whole-genome mutational load in human-disease-specific induced pluripotent stem cell clones. Cell Stem Cell. 2014; 15:31–36.
43. Veres A, Gosis BS, Ding Q, Collins R, Ragavendran A, Brand H, Erdin S, Cowan CA, Talkowski ME, Musunuru K. Low incidence of off-target mutations in individual CRISPR-Cas9 and TALEN targeted human stem cell clones detected by whole-genome sequencing. Cell Stem Cell. 2014; 15:27–30.
44. Sanchez-Garcia I, Martin-Zanca D. Regulation of Bcl-2 gene expression by BCR-ABL is mediated by Ras. J Mol Biol. 1997; 267:225–228.
45. Melo JV, Deininger MW. Biology of chronic myelogenous leukemia—signaling pathways of initiation and transformation. Hematol Oncol Clin North Am. 2004; 18:545–568, vii-viii.
46. Deutsch E, Dugray A, AbdulKarim B, Marangoni E, Maggiorella L, Vaganay S, M’Kacher R, Rasy SD, Eschwege F, Vainchenker W, Turhan AG, Bourhis J. BCR-ABL down-regulates the DNA repair protein DNA-PKcs. Blood. 2001; 97:2084–2090.
47. Koptyra M, Cramer K, Slupianek A, Richardson C, Skorski T. BCR/ABL promotes accumulation of chromosomal aberrations induced by oxidative and genotoxic stress. Leukemia. 2008; 22:1969–1972.
48. Radich J. Stem cell transplant for chronic myeloid leukemia in the imatinib era. Semin Hematol. 2010; 47:354–361.
49. Gori JL, Hsu PD, Maeder ML, Shen S, Welstead GG, Bumcrot D. Delivery and Specificity of CRISPR-Cas9 Genome Editing Technologies for Human Gene Therapy. Human gene therapy. 2015; 26:443–451.
50. Gundry MC, Brunetti L, Lin A, Mayle AE, Kitano A, Wagner D, Hsu JI, Hoegenauer KA, Rooney CM, Goodell MA, Nakada D. Highly Efficient Genome Editing of Murine and Human Hematopoietic Progenitor Cells by CRISPR/Cas9. Cell reports. 2016; 17:1453–1461.
51. Kim S, Kim D, Cho SW, Kim J, Kim JS. Highly efficient RNA-guided genome editing in human cells via delivery of purified Cas9 ribonucleoproteins. Genome research. 2014; 24:1012–1019.
52. Daley GQ, Baltimore D. Transformation of an interleukin 3-dependent hematopoietic cell line by the chronic myelogenous leukemia-specific P210bcr/abl protein. Proc Natl Acad Sci USA. 1988; 85:9312–9316.
53. Schultze N, Burki Y, Lang Y, Certa U, Bluethmann H. Efficient control of gene expression by single step integration of the tetracycline system in transgenic mice. Nat Biotechnol. 1996; 14:499–503.
54. Perez-Caro M, Gutierrez-Cianca N, Gonzalez-Herrero I, Lopez-Hernandez I, Flores T, Orfao A, Sanchez-Martin M, Gutierrez-Adan A, Pintado B, Sanchez-Garcia I. Sustained leukaemic phenotype after inactivation of BCR-ABLp190 in mice. Oncogene. 2007; 26:1702–1713.
55. Sanjana NE, Shalem O, Zhang F. Improved vectors and genome-wide libraries for CRISPR screening. Nat Methods. 2014; 11:783–784.
56. Heckl D, Kowalczyk MS, Yudovich D, Belizaire R, Puram RV, McConkey ME, Thielke A, Aster JC, Regev A, Ebert BL. Generation of mouse models of myeloid malignancy with combinatorial genetic lesions using CRISPR-Cas9 genome editing. Nat Biotechnol. 2014; 32:941–946.
57. Margulies M, Egholm M, Altman WE, Attiya S, Bader JS, Bemben LA, Berka J, Braverman MS, Chen YJ, Chen Z, Dewell SB, Du L, Fierro JM, et al. Genome sequencing in microfabricated high-density picolitre reactors. Nature. 2005; 437:376–380.
58. Ordonez JL, Amaral AT, Carcaboso AM, Herrero-Martin D, del Carmen Garcia-Macias M, Sevillano V, Alonso D, Pascual-Pasto G, San-Segundo L, Vila-Ubach M, Rodrigues T, Fraile S, Teodosio C, et al. The PARP inhibitor olaparib enhances the sensitivity of Ewing sarcoma to trabectedin. Oncotarget. 2015; 6:18875–18890. doi: 10.18632/oncotarget.4303.