
INTRODUCTION
Aging is defined as the progressive decline of biological function with an increasing risk for many late-life onset diseases, including cancer, diabetes, neurodegeneration and metabolic syndromes [1]. As a major elucidation that underlies aging, excess reactive oxygen species (ROS) and induced oxidative stress adversely cause inevitable damage to organisms and age-related pathological conditions. This comprises the principle of the well-known free radical theory of aging [2]. Mice administered with oral or subcutaneous D-galactose, a reducing sugar that generates advanced glycation end products in vivo [3], are extensively used to mimic the aging process. This oxidative damage-induced aging model is accompanied by high-level thiobarbituric acid reactive substances (TBARS), low superoxide dismutase (SOD) activity in various tissues [4, 5], telomere loss and compromised telomerase activities in the hippocampus [6].
Accumulating evidence has clarified the change of telomeres in human aging-related diseases and the aging process [7] while telomere attrition is acknowledged as a robust hallmark of aging [8]. Telomeres, which are characterized by repeated DNA sequences at the terminal end of eukaryotic chromosomes, are protective against the DNA damage response and are essential for genome stability [9]. Unfortunately, telomeres naturally shorten during cell division, which ultimately triggers replicative senescence [10]. The telomere length is canonically maintained by a ribonucleoprotein reverse transcriptase referred to as telomerase [9]. In humans, telomerase is only ubiquitously expressed during the initial weeks of embryogenesis, followed by down-regulation in most cell types. Therefore, the inactivated telomerase activity and telomere attrition function as a tumor-suppressing mechanism by preventing cells from dividing indefinitely [10].
Both p53-p21 and p16-pRb are major cellular pathways during the senescence process. The p16 expression, which markedly increases with aging in many tissues in rodents and humans, may be used as a biomarker of physiologic age [11]. Physiological p53 activity is beneficial for cancer prevention and aging protection, whereas excessive p53 activation is detrimental to healthy aging [12]. Increasing p16 and p53 levels are commonly induced in senescent cells and have been identified as consistent oncogene-induced senescence markers both in humans and mice [11, 13].
Nutrition is believed to promote healthy aging. In this context, n-3 polyunsaturated fatty acids (PUFA) are promising as an anti-aging dietary supplement. Representative n-3 PUFA bioactive compounds include docosahexaenoic acid (DHA) and eicosapentaenoic acid (EPA) which are abundant in fish oil. These primarily marine-derived fatty acids are able to ameliorate chronic diseases and many age-related diseases or impairments [14, 15]. Moreover, recent studies have shed light on the association between n-3 PUFA and senescence. For example, DHA prevents tumor necrosis factor-alpha (TNF-α)-induced senescence and dysfunction in endothelial cells [16], while concentrated fish oil extends the lifespan of lupus-prone short-lived (NZB×NZW)F1 mice [17]. However, the mechanisms responsible for n-3 PUFA counteracting senescence remain poorly understood. Moreover, several critical studies have demonstrated the association of n-3 PUFA with the delay of human telomere shortening. A 5-year follow-up study reported an inverse relationship between the baseline levels of whole blood n-3 PUFA and the rate of telomere shortening in 608 ambulatory outpatients with stable coronary artery disease [18]. Another work indicated that telomere shortening in elderly individuals with mild cognitive impairment or patients with chronic kidney disease may be attenuated with n-3 PUFA supplementation [19]. To our best knowledge, no animal experiment has been conducted to investigate the mechanisms of telomere protection by n-3 PUFA because the functionality of n-3 PUFA remains emerging in the anti-aging field, let alone its influence on telomere.
Here we systematically investigated the anti-aging effect of fish oil and long-chain PUFA monomers on D-galactose-induced mice in facets of in vivo redox-telomere-antioncogene axis. We subsequently emphasized the effect of PUFA on oxidative stress in aging mice and evidenced the PUFA protection of telomere and antioncogene homeostasis via comparing the functionality of n-3 and n-6 PUFA.
RESULTS
Body weights
There was no significant difference of body weights among all groups at the baseline levels. However, the weights of mice in the aging model group significantly decreased due to the aging outcome compared with those in the saline control group at the end of animal study (P < 0.05). Nevertheless, body weights were not significantly changed in all other groups of mice induced by D-galactose treatment (P > 0.05) except the moderate-dose No. 2 fish oil (200FO2) group (Supplementary Table S1).
PUFA improve the in vivo redox state
The effects of PUFA on the redox state were initially investigated to determine the primary anti-aging effects of PUFA in the facet of the free radical theory of aging. Various antioxidase activities in selected tissues were investigated because of their critical antioxidative defense capacities. Moreover, malonaldehyde is considered as an oxidative stress biomarker, which is a typical end product of PUFA and expressed as TBARS equivalents to indicate the lipid peroxidation state in biological membranes [20].
Compared with the aging model group, fish oil and DHA treatments in all groups except the high-dose No. 1 fish oil (400FO1) group significantly promoted hepatic SOD activities within the range of 18%-46% in a non-linear dose-response manner, whereas the arachidonic acid (AA) treatment enhanced activities in a dose-dependent manner (P < 0.05) (Figure 1A). FO1 at a moderate dose, FO2 at low and moderate doses and PUFA monomers at all doses were significantly superior to vitamin E for their promotion effects on hepatic SOD activities. Moreover, FO1 (100 and 200 mg/kg/d), FO2 (400 mg/kg/d), DHA (60 mg/kg/d) and AA (120 mg/kg/d) treatments improved hepatic catalase (CAT) activities by 65%, 56%, 28%, 26% and 31%, respectively (P < 0.05, Figure 1B). However, improvements of hepatic glutathione peroxidase (GSH-Px) activities were not identified in all groups (P > 0.05, Figure 1C). Similarly, previous work identified different improvements in these hepatic antioxidase activities [21], indicating antioxidase species may vary in the sensitivity to nutritional intervention of PUFA. Notably, fish oil had no significant effect on lipid peroxidation in the liver (P > 0.05, Supplementary Table S2), whereas PUFA monomers at high and moderate doses adversely increased the hepatic TBARS level (P < 0.05, Supplementary Table S2). This result indicates that PUFA monomers may exhibit a high oxidative potential due to their multiple conjugated double bonds.
Expectedly, the mice in aging model group exhibited significantly minimized cardiac SOD activities compared with the control group (P < 0.05). Fish oil and PUFA monomers promoted cardiac SOD activity within the range of 19%-58% in the aging mice (P < 0.05, Supplementary Figure S1). Interestingly, only FO1 at a moderate dose significantly reduced cardiac lipid peroxidation, whereas AA monomer at high and low doses increased the TBARS levels (P < 0.05, Supplementary Table S2), which indicates a side effect of AA monomer supplementation.
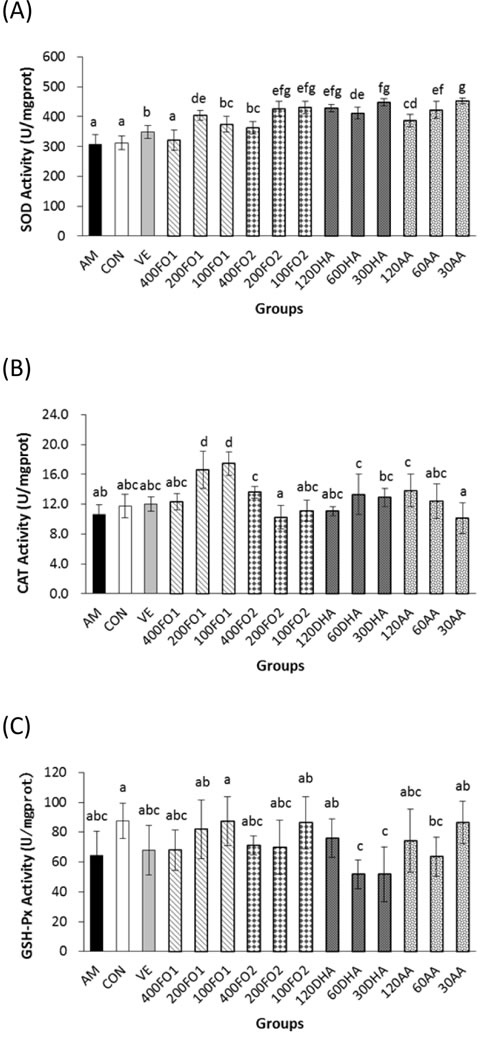
Figure 1: PUFA ameliorate hepatic oxidative stress. Both fish oil and PUFA monomers significantly promoted hepatic A. SOD and B. CAT activities (P < 0.05). However, the improvement in C. GSH-Px activities did not seem significant (P > 0.05). Each column with error bar indicates the data expressed as mean ± standard deviation (SD) (n = 5). Bars marked with different letters indicate significant discrepancies (P < 0.05). AM, aging model group; Con, saline control group; VE, vitamin E positive control; 400FO1, 200FO1, and 100FO1, FO1 (DHA/EPA ratio = 2.28) at high, moderate and low doses, respectively; 400FO2, 200FO2, and 100FO2, FO2 (DHA/EPA ratio = 0.66) at high, moderate and low doses, respectively; 120DHA, 60DHA, and 30DHA, DHA at high, moderate and low doses, respectively; 120AA, 60AA, and 30AA, AA at high, moderate and low doses, respectively.
Furthermore, monoamine oxidase (MAO) also contributes to the ROS production because it catalyzes the oxidative deamination of dietary amines, monoamine neurotransmitters, and hormones that generate hydrogen peroxide [22]. Moreover, the age-related increase in MAO activity is acknowledged to contribute to brain cellular degeneration [23]. Therefore, we assessed the activities of MAO and SOD in the brain. As shown in Supplementary Figure S2, the reduced SOD activities and increased MAO activities in the brains of mice in the aging model group were consistent with the mimetic aging process via injection with D-galactose (P < 0.05). FO1 at all doses and FO2 at high and moderate doses ameliorated the reduction in cerebral SOD activities within the range of 27%-34% and 31%-38%, respectively, whereas AA at a moderate dose only enhanced SOD activities by 5% compared with the aging model group (P < 0.05, Supplementary Figure S2A). In addition, both fish oil and PUFA monomers were effective in inhibiting cerebral MAO activities (Supplementary Figure S2B) and TBARS levels (Supplementary Table S2) within the range of 56%-90% and 16%-62%, respectively (P < 0.05). Notably, DHA and AA monomers exhibited opposite dose-response effects in the inhibition of cerebral MAO activities. Briefly, these data indicate that PUFA may be beneficial for cerebral oxidative stress and have a protective potential for age-related degeneration.
Considering the challenging facet of ROS as extremely reactive and short-lived molecules, F2-isoprostane, which represents the gold standard for oxidative stress status in vivo, was measured. This prostaglandin F2-like compound is formed in vivo via non-enzymatic free radical-catalyzed peroxidation of AA [24]. In addition, the serum GSH-Px activities are investigated to determine the antioxidative defense. The results demonstrated that supplemental PUFA substantially reduced the in vivo oxidative stress status, as evidenced by significant inhibition of the plasma F2-isoprostane levels within the range of 25%-79% (P < 0.05, Supplementary Figure S3A). However, only high-dose FO2 significantly enhanced the serum GSH-Px activities (P < 0.05, Supplementary Figure S3B).
Taken together, both fish oil and PUFA monomers exerted promising regulation of the in vivo redox state via the promotion of antioxidase (hepatic CAT and SOD in the liver, heart and brain) activities and the reduction of cerebral MAO activities, cerebral TBARS levels and the plasma F2-isoprostane levels (P < 0.05). However, supplemental PUFA monomers at high and moderate doses may have the side effect of an increased TBARS level (P < 0.05). In addition, the dose-response manner of redox state improvement depends on the observed index. The different DHA/EPA ratios in FO1 and FO2 produced a minimized discrepancy in the in vivo redox state improvement, particularly the dose-response effects on the hepatic CAT, and cerebral SOD and MAO activities.
n-3 rather than n-6 PUFA ameliorate telomere shortening
The effects of PUFA on telomere attrition were subsequently investigated to indicate an intrinsic aging process. Expectedly, the hepatic telomere length of mice in the aging model group was significantly shorter than the control group (P < 0.05, Figure 2A), which was in accordance with the expectation that telomere erosion is accelerated in the D-galactose-induced aging model. Compared with the aging model group, the samples in high-dose FO1, low- and moderate-dose FO2, and high- and low-dose DHA groups significantly inhibited hepatic telomere shortening within the range of 13%-25% (P < 0.05), whereas no promising effect was identified in the AA groups (P > 0.05, Figure 2A). Telomere attrition in the testes was not as obvious as in the liver; nevertheless, moderate-dose FO2 and low-dose DHA exhibited 25% and 27% inhibitory effects on the testicular telomere attrition, respectively (P < 0.05, Figure 2B).
Briefly, n-3 rather than n-6 PUFA supplements effectively ameliorate telomere shortening in the format of an n-3 PUFA-enriched fish oil or monomer as exemplified in the telomere length analysis of the liver and testes (P < 0.05). Moreover, the telomere protected by n-3 PUFA does not appear in a linear dose-response manner.
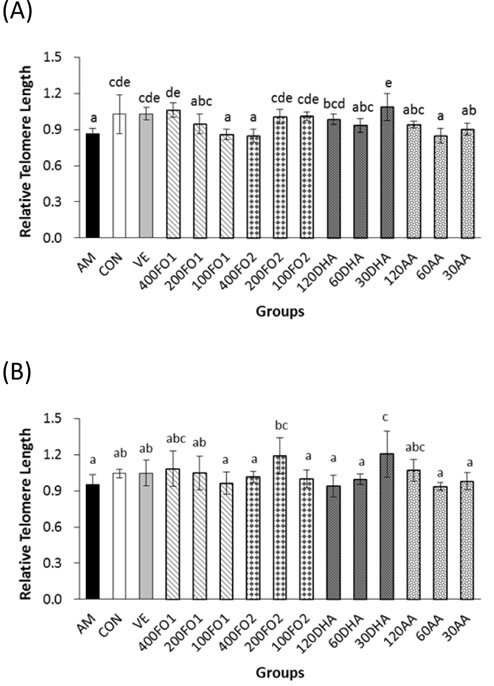
Figure 2: n-3 rather than n-6 PUFA inhibit excessive telomere length loss in A. livers and B. testes of aging mice. Each column with error bar indicates the data expressed as mean ± SD (n = 5). Bars marked with different letters indicate significant discrepancies (P < 0.05). The instructions about the group labels were the same as indicated in Figure 1.
Both n-3 and n-6 PUFA inactivate testicular telomerase through the inhibition of c-Myc-mediated TERT expression
To further explain the telomere protection in the n-3 PUFA-supplemental groups, the testicular telomerase activities were measured. Representative images of the testes in the experimental groups were shown in Supplementary Figure S4. Surprisingly, the mice that exhibited shorter telomere in the aging model group demonstrated substantially higher telomerase activities compared with the control group (P < 0.05, Figure 3A). It has been reported that telomere length homeostasis required limiting telomerase levels [25]. Therefore, the explanation for ectopic telomerase expression in aging mice is likely attributed to the response induced by acute telomere erosion. Moreover, high-dose fish oil, all-dose DHA and high- and low-dose AA significantly repressed telomerase activities within the ranges of 40%-71%, 45%-61% and 52%-54%, respectively (P < 0.05, Figure 3A). Collectively, FO2 at a moderate dose exerted a telomere-protective effect, whereas a high dose was required to suppress telomerase activity. The low-dose DHA monomer was capable of protecting telomere erosion, whereas all-dose DHA monomer was effective on inactivating telomerase activity. Moreover, AA at all doses had no influence on the telomere length, but AA at high and low doses significantly inhibited telomerase activity. There was no significant difference of telomerase inhibition between n-3 and n-6 PUFA (P > 0.05).
To further confirm telomerase inhibition by PUFA, we subsequently analyzed the expression of TERT and its transcription factor, c-Myc, in the testes. TERT, which may be regulated both transcriptionally and epigenetically, is a catalytic subunit of telomerase and its expression is acknowledged as the rate-limiting factor for telomerase catalytic activity [26]. For example, c-Myc directly mediates TERT transcriptional activation because the TERT promoter contains numerous c-Myc-binding sites [27]. Figure 3B indicates that no significant change in the TERT mRNA expression was identified in the aging mice (P > 0.05). However, both FO2 and DHA at moderate and low doses and AA at high dose significantly reduced the TERT protein expression within the ranges of 25%-45%, 26%-36% and 34%, respectively (P < 0.05, Figure 3C). The discrepancy between the mRNA and protein expression of TERT remains enigmatic. Nevertheless, a similar inconsistency has been reported with regard to other genes [28]. One potential explanation is that complicated biological processes, including transcriptional or post-transcriptional splicing, translational modifications, and other processes, may alter the relative quantities of mRNA and protein to various degrees [28]. Furthermore, the protein expression of c-Myc was also significantly reduced within the range of 30%-60% following the intervention of moderate- and low-dose FO1, all-dose FO2, low-dose DHA, and all-dose AA (P < 0.05, Figure 3D). We further used immunofluorescence assay to demonstrate the above reduction effect of PUFA on the protein expression of TERT and c-Myc (Figure 4). Overall, telomerase inactivation by PUFA in aging mice may be explained by the c-Myc-mediated TERT pathway.
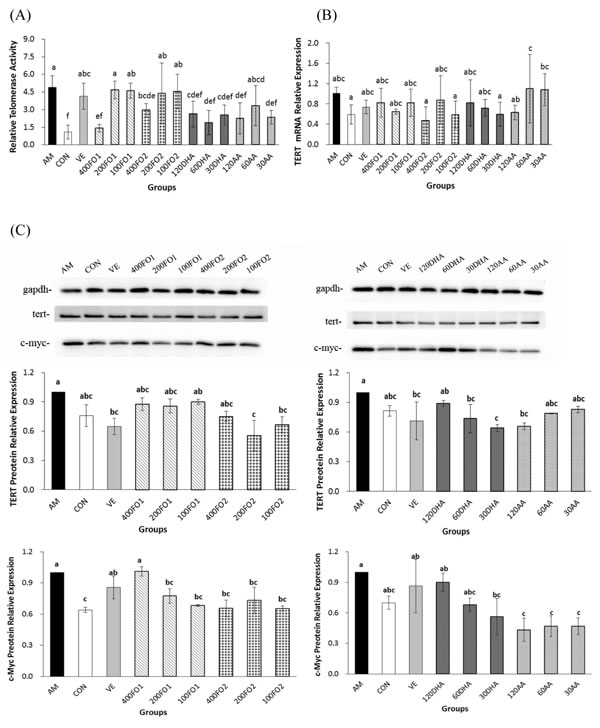
Figure 3: PUFA repress testicular telomerase activities through the reduction of c-Myc-mediated TERT expression in aging mice. A. The effect of PUFA on telomerase activities (n = 5). B. The effect of PUFA on the mRNA expression of testicular TERT (n = 5). C. The effect of PUFA on the protein expression of testicular TERT and c-Myc (n = 3). The TERT and c-Myc primary antibody recognized a single protein band with molecular weights of approximately 126 kDa and 57 kDa, respectively. Each column with error bar indicates the data expressed as mean ± SD. Bars marked with different letters indicate significant discrepancies (P < 0.05). gapdh, glyceraldehyde-3-phosphate dehydrogenase. The instructions about the group labels were the same as indicated in Figure 1.
DHA inhibits testicular antioncogene p16 and p53 expression
Furthermore, effects of PUFA on the critical molecular pathways of cellular senescence in the testes of aging mice were investigated. The results indicated that all-dose DHA significantly inhibited the testicular p16 protein expression and low-dose DHA repressed the p53 protein expression (P < 0.05, Figure 5A and 5B), whereas fish oil did not significantly alter the p16 and p53 expression. The senescence-associated β-galactosidase (SA-β-gal) staining of whole-mount testes and testicular cryosections only indicated a mild discrepancy in the PUFA intervention groups (Supplementary Figure S5). However, DHA remained effective in the inhibition of SA-β-gal activities in the testicular cryosections of aging mice (Figure 5C).
Taken together, DHA rather than AA significantly inactivated the p16 and p53 expressions at their certain intervention levels. Combined with previous findings, we conclude that n-3 rather than n-6 PUFA may ameliorate the senescence process via the redox-telomere-antioncogene axis.
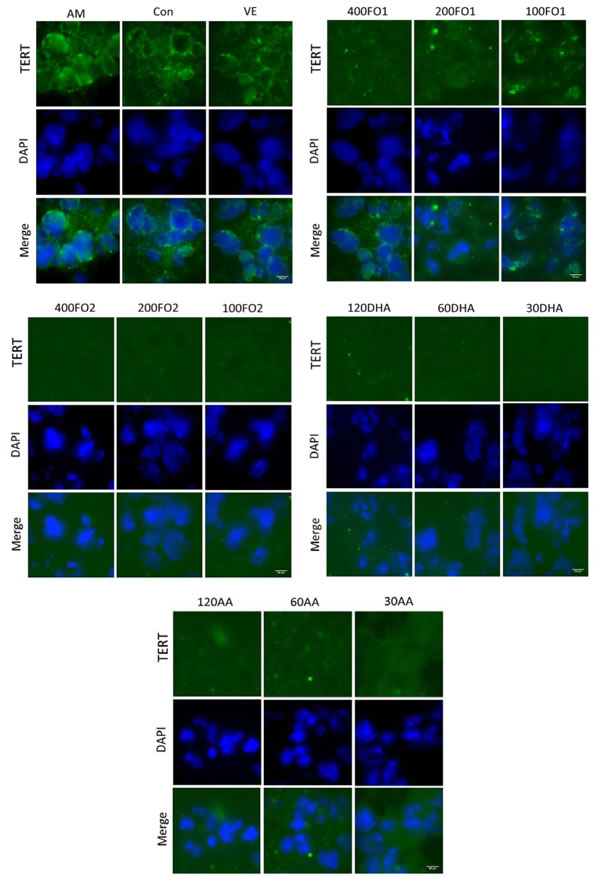
Figure 4: PUFA reduce the testicular TERT protein expression confirmed by the immunofluorescence of histological testicular sections. DAPI, 4’,6-diamidino-2- phenylindole. Scale bar, 50 μm.
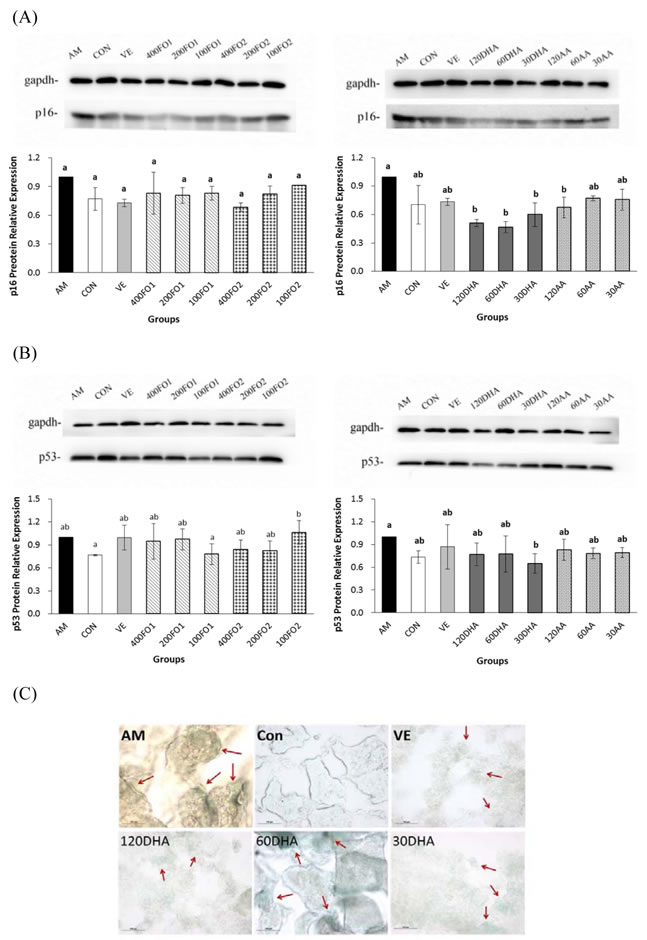
Figure 5: DHA inhibits testicular p16 and p53 protein expression and SA-β-gal. A. The effect of PUFA on the protein expression of testicular p16. B. The effect of PUFA on the protein expression of testicular p53. The p16 and p53 primary antibodies recognized a single protein band with molecular weights of approximately 16 kDa and 53 kDa, respectively. C. Representative fields of in situ SA-β-gal staining in testicular cryosections. Arrows indicate foci with high SA-β-gal activities. Each column with error bar indicates the data expressed as mean ± SD (n = 3). Bars marked with different letters indicate significant discrepancies (P < 0.05). The instructions about the group labels were the same as indicated in Figure 1.
DISCUSSION
Consistent with previous findings that fish oil and PUFA are capable of ameliorating oxidative stress in various tissues [29, 30], we demonstrated that both fish oil and PUFA monomers also effectively ameliorate D-galactose-induced oxidative stress in vivo by promoting antioxidase activities (mainly hepatic CAT and hepatic, cardiac and cerebral SOD activities) and reducing the plasma F2-isoprostane levels. In addition, the beneficial protection of PUFA against cerebral oxidative stress evidenced by reduced cerebral TBARS levels and MAO activities is consistent with previous work [31]. However, long-chain PUFA are susceptible to oxidation, and a high dietary intake of n-3 PUFA may trigger additional oxidative stress without adequate antioxidative protection. For example, the plasma TBARS levels were significantly increased in the fish oil group compared with the placebo control during pregnancy [32]. Supplemental fish oil increased the TBARS levels in autoimmune-prone NZB/W female mice fed ad libitum and injected with cyclophosphamide compared with corn oil [15]. In our present work, supplemental PUFA monomers at high and moderate doses promoted hepatic TBARS levels, whereas AA at high and low doses also increased cardiac oxidative stress. PUFA monomers without the presence of antioxidants are more susceptible to oxidation in free fatty acid forms than fish oil in vivo, which ultimately produces an increased TBARS level. Previous studies have demonstrated that n-3 PUFA were superior to n-6 PUFA in the enhancement of antioxidase activities [21, 33]. Interestingly, our findings indicated that DHA may promote antioxidase activities to the same extent as AA because DHA has more double bonds than AA and is more susceptible to oxidation. Therefore, the oxidative propensity of PUFA, as well as its functionality should be simultaneously taken into consideration.
With respect to the telomere theory of aging, several recent studies have provided epidemiological evidence that n-3 PUFA may prevent excessive telomere attrition [18, 19, 34]. Similarly, we demonstrated that only n-3 PUFA was effective in inhibiting hepatic and testicular telomere shortening in aging mice. In addition, telomere shortening depends on the balance between oxidative stress and antioxidative defense because telomeres are triple-G-containing structures, which are susceptible to damage by oxidative stress [35]. Therefore, our results revealed that the protective effect of n-3 PUFA on telomere may be partly attributed to the amelioration of redox state. Nevertheless, both n-3 and n-6 PUFA significantly reduced telomerase activities independent of the telomere length. This finding is probably associated with the two-edged-sword property of telomerase. Telomerase is canonically responsible for telomere length maintenance, whereas its activation may favor tumorigenesis [36]. Telomere dysfunction is believed to drive the early stages of cancer development, and subsequent telomerase activation appears to be critical for malignant progression [37]. Moreover, telomerase activity and telomere length have been acknowledged as markers for distinguishing prostate cancer from normal and benign prostate tissues [38]. In this context, both n-3 and n-6 PUFA are beneficial for testicular telomere hemostasis and protective from prostate cancer via the inhibition of ectopic telomerase expression in the aging model group. An in vitro study has demonstrated that EPA and DHA suppressed the telomerase activity and TERT mRNA levels in a time- and dose-dependent manner [39]. Thus, telomerase acts as a critical molecular target in the growth and survival of cancer cells modulated by n-3 PUFA [40]. Similarly, our results demonstrated that long-chain PUFA were effective in inactivating telomerase likely through the inhibition of c-Myc-mediated TERT expression.
Oxidative stress has been demonstrated to be a potent inducer of p53 [2], which also acts in a context-dependent manner. To respond low levels of oxidative stress, p53 exhibits antioxidant activities to ensure cell survival. In contrast, to respond severe oxidative stress, p53 exhibits prooxidant activities and leads to cell death [41]. Therefore, it is not difficult to interpret the controversial data that n-3 PUFA may inhibit [42, 43] or induce [44, 45] p53 expression in both in vivo and in vitro experiments. However, none of these studies have addressed the effects of n-3 PUFA on p53 expression in an anti-aging view. Similarly, p16 also has a balance in cancer and senescence [44, 46]. Many noxious stimuli, including ROS, ionizing radiation, and UV light, have been demonstrated to induce p16 expression in vitro and in vivo [47]. The p16 expression substantially increases in nearly all rodent tissues with advancing age. Thus, it is acknowledged as a robust biomarker of aging [11]. Precise expression of p16 is essential for tissue homeostasis, maintaining a coordinated balance between tumor suppression and aging. Although anti-tumor drug development teams are in favor of activating the antioncogene p16, aging biologist groups aim to block the accumulation of p16-positive cells. As a result, the key to longevity likely depends on the hands of both groups, as a careful balance of p16 expression is required to stave off cancer and prevent aging [48]. Such balance is also applicable when n-3 PUFA are used to manage antioncogenes. A recent study indicated the anti-aging potential of n-3 PUFA that the overexpression of Fat1, an endogenously synthesizing n-3 PUFA model, in Boer goat fetal fibroblasts reduced the mRNA expression of p16 and p53 [49]. Nevertheless, fish oil mediates apoptosis of tumor cells by promoting the protein expression of p53 [50]. In accordance with this study, our results demonstrated the inhibition of both p16 and p53 protein expression by DHA, which contributes to the hypothesis that n-3 PUFA may slow down the process of cell and organ senescence.
Additionally, our present work demonstrated that both n-3 and n-6 PUFA inactivated telomerase, thereby exerting their anti-cancer potential. However, DHA significantly inhibited p16 and p53, which appears vague in the setting of tumor inhibition. In vivo studies have demonstrated that the inhibition of tumor suppressors (such as p16 and p53) and up-regulation of telomerase activity and TERT expression may promote tumorigenesis, including prostate cancer [51, 52], whereas the opposite features underline senescence [53]. Therefore, both n-3 and n-6 PUFA may inhibit tumorigenesis via the suppression of oxidative stress-induced telomerase activation, whereas n-3 PUFA exerted an anti-aging effect via the protection of telomere attrition and down-regulation of tumor suppressors (Supplementary Figure S6).
The reason for DHA rather than fish oil effectively inhibiting the protein expression of p16 and p53 may be ascribed to the multi-ingredient mixture of fish oil including DHA, EPA, monounsaturated fat and other compounds. In addition to n-3 PUFA compositions, ethyl esters and triglycerides are two main forms of commercial fish oil available in the market [54]. The apparent bioavailability is considered to be the lowest for the ethyl ester form and the highest for the free fatty acid form [55]. Thus, the anti-aging effect of PUFA in different conjugation forms requires further investigations.
In conclusion, we systematically investigated the in vivo anti-aging effect of PUFA from the facets of oxidative stress amelioration and telomere protection. Long-chain PUFA effectively ameliorated the in vivo redox state in aging mice via promoting SOD activity and reducing cerebral MAO activity, cerebral TBARS levels and plasma F2-isoprostane levels. However, supplemental PUFA monomers at high and moderate doses should be carefully administered in cases of increased oxidative stress. In addition, both n-3 and n-6 PUFA significantly inhibited telomerase activities due to the reduction of c-Myc-mediated TERT expression, which indicates the tumor-inhibitive potential of long-chain PUFA. However, only n-3 PUFA protected telomere from progressive attrition, and DHA was superior in the inhibition of tumor suppressors, p16 and p53, which indicates that n-3 PUFA may be served as a potent anti-aging supplement.
MATERIALS AND METHODS
Animals and experimental design
One hundred twenty Institute of Cancer Research (ICR) male mice (8 weeks old, weighed 30-40 g), purchased from the Laboratory Animal Research Center of Zhejiang Chinese Medical University, were housed at (20±1) °C on a cycle of 12-h light/12-h dark. All experimental protocols were approved by the Ethical Committee of the College of Biosystems Engineering and Food Science at Zhejiang University in China. Mice were randomly assigned to 15 groups (8 animals per group): the negative saline control (Group Con), the D-galactose-induced aging model (Group AM), the vitamin E positive control (Group VE), and groups of FO1 (Groups 400FO1, 200FO1, and 100FO1), FO2 (Groups 400FO2, 200FO2, and 100FO2), DHA (Groups 120DHA, 60DHA, and 30DHA) and AA (Groups 120AA, 60AA, and 30AA) with high, moderate and low administration doses. The mice in the negative control group were intraperitoneally injected with vehicle (saline) throughout the two-month experiment, whereas the mice in all other groups were injected with D-galactose (Aladdin-Reagent Co. Ltd., Shanghai, China) solution at a dose of 80 mg/kg/d to induce senescence. The mice in the Con and AM groups were administered 100 mg/kg/d of corn oil, whereas the mice in the VE group received 100 mg/kg/d of vitamin E. To observe the possible dose-response anti-aging effect of PUFA, the mice in FO1 and FO2 groups received 400, 200 and 100 mg/kg/d of fish oil via oral gavage, while the mice in DHA and AA groups were orally administered with 120, 60 and 30 mg/kg/d of DHA and AA, respectively. Both the intraperitoneal injection and oral gavage were performed daily. The mice in each group were weighed weekly to adjust the dose of the D-galactose and treatment samples.
Determination of antioxidase activities
The hepatic, cerebral and cardiac SOD activities, hepatic CAT activity, hepatic and serum GSH-Px activities, and cerebral MAO activity were determined using kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the manufacturer’s instructions.
Determination of TBARS levels
The TBARS in the liver, heart and brain were determined using the well-established thiobarbituric acid test with a kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Determination of F2-isoprostane levels
The plasma F2-isoprostane levels were measured using an enzyme immunoassay kit (Cayman Chemical, Ann Arbor, MI, USA).
Telomere length analysis
The relative telomere length in the liver and testicle, expressed as a ratio of the telomere repeat copy number to single-copy gene copy number (T/S), was measured via quantitative real-time polymerase chain reaction (PCR) as previously described, while 36B4 (acidic ribosomal phosphoprotein PO) is acknowledged as the single-copy gene [56]. The sequences and final concentrations of the primers (Invitrogen Life Sciences, Carlsbad, CA) were as follows: Tel F, 5’-CGGTTTGTTTGGGTTTGGGTTTGGGTTTGGGTT TGGGTT-3’, 300 nM; Tel R, 5’-GGCTTGCCT TACCCTTACCCTTACCCTTACCCTTACCCT-3’, 900 nM; 36B4 F, 5’-ACT GGT CTA GGA CCC GAG AAG-3’, 300 nM; and 36B4 R, 5’-TCA ATG GTG CCT CTG GAG ATT-3’, 500 nM.
Telomerase activity assay
The testicular telomerase activities in all groups were measured using a TeloTAGGG telomerase PCR ELISA kit (Roche applied science, Indianapolis, IN, USA).
Real-time quantitative reverse transcription PCR
RNA was isolated using TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA) from frozen testicle samples and reverse transcribed into cDNA using a RT reagent kit with gDNA Eraser (Takara Bio Inc., Otsu, Japan) according to the manufacturer’s instructions. The sequences and final concentrations of the primers for mTERT and β-actin (as the internal control) were as follows: β-actin F, 5’-TGACATCCGTAAAGA-3’, 400 nM; β-actin R, 5’-CAGCTCAGTAACAGTCC-3’, 400 nM; mTERT F, 5’-ATGGCGTTCCTGAGTATG-3’, 400 nM; mTERT R, 5’-TTCAACCGCAAGACCGACAG-3’, 400 nM. Similarly, the ΔΔCt method (mTERT /β-actin = 2−ΔΔCt) was used to analyze the results.
Whole-mount and in situ SA-β-gal staining
Whole-mount testes SA-β-gal staining and in situ SA-β-gal staining in testicular cryosections were processed with a Senescence β-Galactosidase Staining Kit (Cell Signaling Technology, Beverly, USA) with modifications.
Western blotting analysis and immunofluorescence
Standard Western blotting and immunofluorescence procedures were conducted with modifications. The primary antibodies for detecting protein expression included rabbit polyclonal anti-β-actin (as the internal control; E021070; EarthOx, LLC, San Francisco, CA), rabbit polyclonal anti-TERT (sc-7212; Santa Cruz Bio., Santa Cruz, CA), rabbit monoclonal anti-c-Myc (#5605; Cell Signaling Technology, Beverly, MA), mouse monoclonal anti-p53 (ab26; Abcam, Cambridge, MA) and mouse monoclonal anti-p16 (SAB5300498, Sigma-Aldrich, St. Louis, MO). Secondary antibodies purchased from EarthOx, LLC (San Francisco, CA) and the anti-goat Alexa Fluor 488-conjugated secondary antibody (Invitrogen Life Technologies, Carlsbad, CA) were used in the Western blotting analysis and immunofluorescence, respectively.
Statistical analysis
Statistical analysis was conducted with SPSS version 16.0 software, and all data were presented as the mean ± SD. Multiple comparisons among different groups were conducted with a one-way analysis of variance (ANOVA) followed by Duncan’s test. Data were considered statistically significant at P < 0.05.
Detailed procedures of the above experiments were shown in Supplementary Materials and Methods.
ACKNOWLEDGMENTS
We gratefully thank the financial support from the National Natural Science Foundation of China (Grant No. 31401659 and 31201307) and the Zhejiang Provincial Natural Science Foundation of China (Grant No. LY14C200008).
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
REFRENCES
1. Sanchez JR, Milton SL, Corbit KC, Buffenstein R. Multifactorial processes to slowing the biological clock: Insights from a comparative approach. Exp Gerontol. 2015; 71:27-37.
2. Gambino V, De Michele G, Venezia O, Migliaccio P, Dall’Olio V, Bernard L, Minardi SP, Fazia MAD, Bartoli D, Servillo G, Alcalay M, Luzi L, et al. Oxidative stress activates a specific p53 transcriptional response that regulates cellular senescence and aging. Aging Cell. 2013; 12:435-445.
3. Li YJ, Han Z, Ge L, Zhou CJ, Zhao YF, Wang DH, Ren J, Niu XX, Liang CG. C-phycocyanin protects against low fertility by inhibiting reactive oxygen species in aging mice. Oncotarget. 2016; 7:17393-17409. doi: 10.18632/oncotarget.8165.
4. Budni J, Pacheco R, Da Silva S, Garcez ML, Mina F, Bellettini-Santos T, de Medeiros J, Voss BC, Steckert AV, Valvassori SDS, Quevedo J. Oral administration of d-galactose induces cognitive impairments and oxidative damage in rats. Behav Brain Res. 2016; 302:35-43.
5. Ho SC, Liu JH, Wu RY. Establishment of the mimetic aging effect in mice caused by D-galactose. Biogerontology. 2003; 4:15-18.
6. Zhu J, Mu X, Zeng J, Xu C, Liu J, Zhang M, Li C, Chen J, Li T and Wang Y. Ginsenoside Rg1 prevents cognitive impairment and hippocampus senescence in a rat model of D-galactose-induced aging. PLoS One. 2014; 9:e101291.
7. Blackburn EH, Epel ES, Lin J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science. 2015; 350:1193-1198.
8. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013; 153:1194-1217.
9. Aubert G. (2014). Chapter Four - Telomere Dynamics and Aging. In: Rodrigo TC, ed. Progress in Molecular Biology and Translational Science. pp. 89-111.
10. Suram A, Herbig U. The replicometer is broken: telomeres activate cellular senescence in response to genotoxic stresses. Aging Cell. 2014; 13:780-786.
11. Bernardes de Jesus B, Blasco MA. Assessing cell and organ senescence biomarkers. Circ Res. 2012; 111:97-109.
12. Rufini A, Tucci P, Celardo I, Melino G. Senescence and aging: The critical roles of p53. Oncogene. 2013; 32:5129-5143.
13. Park H, Kim C, Jeong J, Park M, Kim KS. GDF15 contributes to radiation-induced senescence through the ROS-mediated p16 pathway in human endothelial cells. Oncotarget. 2016; 7:9634-9644. doi: 10.18632/oncotarget.7457.
14. Ubeda N, Achon M, Varela-Moreiras G. Omega 3 fatty acids in the elderly. Br J Nutr. 2012; 107:S137-S151.
15. Bhattacharya A, Rahman M, Sun D, Fernandes G. Effect of fish oil on bone mineral density in aging C57BL/6 female mice. J Nutr Biochem. 2007; 18:372-379.
16. Yamagata K, Suzuki S, Tagami M. Docosahexaenoic acid prevented tumor necrosis factor alpha-induced endothelial dysfunction and senescence. Prostaglandins Leukot Essent Fatty Acids. 2016; 104:11-18.
17. Halade GV, Williams PJ, Veigas JM, Barnes JL, Fernandes G. Concentrated fish oil (Lovaza®) extends lifespan and attenuates kidney disease in lupus-prone short-lived (NZBxNZW) F1 mice. Exp Biol Med. 2013; 238:610-622.
18. Farzaneh-Far R, Lin J, Epel ES, Harris WS, Blackburn EH, Whooley MA. Association of marine omega-3 fatty acid levels with telomeric aging in patients with coronary heart disease. JAMA. 2010; 303:250-257.
19. O’Callaghan N, Parletta N, Milte CM, Benassi-Evans B, Fenech M, Howe PRC. Telomere shortening in elderly individuals with mild cognitive impairment may be attenuated with ω-3 fatty acid supplementation: a randomized controlled pilot study. Nutrition. 2014; 30:489-491.
20. Yin H, Xu L, Porter NA. Free radical lipid peroxidation: mechanisms and analysis. Chem Rev. 2011; 111:5944-5972.
21. Venkatraman JT, Chandrasekar B, Kim JD, Fernandes G. Effects of n-3 and n-6 fatty acids on the activities and expression of hepatic antioxidant enzymes in autoimmune-prone NZBxNZW F1 mice. Lipids. 1994; 29:561-568.
22. Bortolato M, Chen K, Shih JC. Monoamine oxidase inactivation: From pathophysiology to therapeutics. Adv Drug Deliv Rev. 2008; 60:1527-1533.
23. Mallajosyula JK, Kaur D, Chinta SJ, Rajagopalan S, Rane A, Nicholls DG, Di Monte DA, Macarthur H, Andersen JK. MAO-B elevation in mouse brain astrocytes results in Parkinson’s pathology. Plos One. 2008; 3:e1616.
24. Montuschi P, Barnes PJ, Roberts LJ. Isoprostanes: markers and mediators of oxidative stress. FASEB J. 2004; 18:1791-1800.
25. Cristofari G, Lingner J. Telomere length homeostasis requires that telomerase levels are limiting. EMBO J. 2006; 25:565-574.
26. Yokoyama Y, Takahashi Y, Shinohara A, Wan X, Takahashi S, Niwa K, Tamaya T. The 5’-end of hTERT mRNA is a good target for hammerhead ribozyme to suppress telomerase activity. Biochem Biophys Res Commun. 2000; 273:316-321.
27. Wu K, Grandori C, Amacker M, Simon-Vermot N, Polack A, Lingner J, Dalla-Favera R. Direct activation of TERT transcription by c-MYC. Nat Genet. 1999; 21:220-224.
28. Guo Y, Xiao P, Lei S, Deng F, Xiao GG, Liu Y, Chen X, Li L, Wu S, Chen Y, Jiang H, Tan L, et al. How is mRNA expression predictive for protein expression? A correlation study on human circulating monocytes. Acta Biochim Biophys Sin. 2008; 40:426-436.
29. Dasilva G, Pazos M, Garcia-Egido E, Manuel Gallardo J, Rodriguez I, Cela R, Medina I. Healthy effect of different proportions of marine omega-3 PUFAs EPA and DHA supplementation in Wistar rats: Lipidomic biomarkers of oxidative stress and inflammation. J Nutr Biochem. 2015; 26:1385-1392.
30. Fan C, Zirpoli H, Qi K. n-3 fatty acids modulate adipose tissue inflammation and oxidative stress. Curr Opin Clin Nutr. 2013; 16:124-132.
31. Saada HN, Said UZ, Mahdy EME, Elmezayen HE, Shedid SM. Fish oil omega-3 fatty acids reduce the severity of radiation-induced oxidative stress in the rat brain. Int J Radiat Biol. 2014; 90:1179-1183.
32. Franke C, Demmelmair H, Decsi T, Campoy C, Cruz M, Molina-Font JA, Mueller K, Koletzko B. Influence of fish oil or folate supplementation on the time course of plasma redox markers during pregnancy. Br J Nutr. 2010; 103:1648-1656.
33. Yessoufou A, Soulaimann N, Merzouk SA, Moutairou K, Ahissou H, Prost J, Simonin AM, Merzouk H, Hichami A, Khan NA. n-3 Fatty acids modulate antioxidant status in diabetic rats and their macrosomic offspring. Int J Obes. 2006; 30:739-750.
34. Kiecolt-Glaser JK, Epel ES, Belury MA, Andridge R, Lin J, Glaser R, Malarkey WB, Hwang BS, Blackburn E. Omega-3 fatty acids, oxidative stress, and leukocyte telomere length: a randomized controlled trial. Brain Behav Immun 2013; 28:16-24.
35. Von Zglinicki T. Oxidative stress shortens telomeres. Trends Biochem Sci. 2002; 27:339-344.
36. Blasco MA. Mice with bad ends: mouse models for the study of telomeres and telomerase in cancer and aging. EMBO J. 2005; 24:1095-1103.
37. Ding Z, Wu C, Jaskelioff M, Ivanova E, Kost-Alimova M, Protopopov A, Chu GC, Wang G, Lu X, Labrot ES, Hu J, Wang W, et al. Telomerase reactivation following telomere dysfunction yields murine prostate tumors with bone metastases. Cell. 2012; 148:896-907.
38. Sommerfeld HJ, Meeker AK, Piatyszek MA, Bova GS, Shay JW, Coffey DS. Telomerase activity: A prevalent marker of malignant human prostate tissue. Cancer Res. 1996; 56:218-222.
39. Eitsuka T, Nakagawa K, Miyazawa T. Dual mechanisms for telomerase inhibition in DLD-1 human colorectal adenocarcinoma cells by polyunsaturated fatty acids. Biofactors. 2004; 21:19-22.
40. Calviello G, Serini S, Palozza P. n-3 polyunsaturated fatty acids as signal transduction modulators and therapeutical agents in cancer. Curr Signal Transd Ther. 2006; 1:255-271.
41. Liu D, Xu Y. p53, oxidative stress, and aging. Antioxid Redox Signal. 2011; 15:1669-1678.
42. Jing K, Song K, Shin S, Kim N, Jeong S, Oh H, Park J, Seo K, Heo J, Han J, Park J, Han C, et al. Docosahexaenoic acid induces autophagy through p53/AMPK/mTOR signaling and promotes apoptosis in human cancer cells harboring wild-type p53. Autophagy. 2011; 7:1348-1358.
43. Manzhulo IV, Ogurtsova OS, Lamash NE, Latyshev NA, Kasyanov SP, Dyuizen IV. Analgetic effect of docosahexaenoic acid is mediated by modulating the microglia activity in the dorsal root ganglia in a rat model of neuropathic pain. Acta Histochem. 2015; 117:659-666.
44. Wan X, Fu X, Ababaikeli G. Docosahexaenoic acid induces growth suppression on epithelial ovarian cancer cells more effectively than eicosapentaenoic acid. Nutr Cancer. 2016; 68:320-327.
45. Perales S, Jose Alejandre M, Palomino Morales R, Torres C, Linares A. Fish oil supplementation reverses the effect of cholesterol on apoptotic gene expression in smooth muscle cells. Lipids Health Dis. 2010; 9:70.
46. McLaughlin-Drubin ME, Park D, Munger K. Tumor suppressor p16(INK4A) is necessary for survival of cervical carcinoma cell lines. Proc Natl Acad Sci U S A. 2013; 110:16175-16180.
47. Vurusaner B, Poli G, Basaga H. Tumor suppressor genes and ROS: complex networks of interactions. Free Radical Bio Med. 2012; 52:7-18.
48. LaPak KM, Burd CE. The molecular balancing act of p16INK4a in cancer and aging. Mol Cancer Res. 2014; 12:167-183.
49. Sun XH, Sun XF, Ma JM, Liu HQ, Min LJ, Pan QJ, Qin GQ, Shen W, Li L. Anti-senescence effect of Fat1 gene in goat somatic cells. Biotechnol Appl Biochem. 2014; 61:691-698.
50. Borghetti G, Yamaguchi AA, Aikawa J, Yamazaki RK, de Brito GA, Fernandes LC. Fish oil administration mediates apoptosis of Walker 256 tumor cells by modulation of p53, Bcl-2, caspase-7 and caspase-3 protein expression. Lipids Health Dis. 2015; 14:94.
51. Harada H, Nakagawa H, Oyama K, Takaoka M, Andl CD, Jacobmeier B, von Werder A, Enders GH, Opitz OG, Rustgi AK. Telomerase induces immortalization of human esophageal keratinocytes without p16(INK4a) inactivation. Mol Cancer Res. 2003; 1:729-738.
52. Fan C, He L, Kapoor A, Gillis A, Rybak AP, Cutz J, Tang D. Bmi1 promotes prostate tumorigenesis via inhibiting p16(INK4A) and p14(ARF) expression. Biochem Biophys Acta-Mol Basis Dis. 2008; 1782:642-648.
53. Bhatia B, Jiang M, Suraneni M, Patrawala L, Badeaux M, Schneider-Broussard R, Multani AS, Jeter CR, Calhoun-Davis T, Hu L, Hu J, Tsavachidis S, et al. Critical and distinct roles of p16 and telomerase in regulating the proliferative life span of normal human prostate epithelial progenitor cells. J Biol Chem. 2008; 283:27957-27972.
54. Tang X, Li Z, Xu J, Xue Y, Li J, Wang J, Yanagita T, Xue C, Wang Y. Short term effects of different omega-3 fatty acid formulation on lipid metabolism in mice fed high or low fat diet. Lipids Health Dis. 2012; 11:70.
55. Ghasemifard S, Turchini GM, Sinclair AJ. Omega-3 long chain fatty acid “bioavailability”: A review of evidence and methodological considerations. Prog Lipid Res. 2014; 56:92-108.
56. Cawthon RM. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002; 30:47.