
Introduction
Coronary heart disease (CHD), is a major cause of morbidity and mortality worldwide [1] and one of the most common chronic inflammatory diseases, characterized by remodeling and narrowing of the blood vessels (coronary arteries) that supply oxygen and blood to the heart [2]. CHD is a complex disease influenced by both environmental and genetic factors [3, 4]. Epidemiological studies have identified many risk factors for CHD, including age, gender, smoking, obesity, diabetes, hyperlipidemia, hypertension, lack of exercise, and dietary factors. Twin and family studies have demonstrated that a significant proportion (40-50%) of susceptibility to CHD is inherited [5]. Although atherosclerosis is one of the main pathophysiological mechanisms of CHD [2], the responsible molecular and genetic determinants remain largely unidentified. Recently, both genome-wide association studies (GWAS) and candidate gene studies have reported that CDKN2BAS (cyclin-dependent kinase inhibitor 2B antisense RNA) is a risk gene for CHD susceptibility [6-8].
CDKN2BAS encodes an antisense non-coding RNA, and is located near the CDKN2A-CDKN2B gene. The precise function of CDKN2BAS is unclear, but it regulates the expression of neighboring protein-coding genes, like CDKN2A, CDKN2B, and MTAP, that enhance the progression of atherosclerosis by influencing vascular remodeling, thrombogenesis, and plaque stability [9, 10]. Therefore, CDKN2BAS expression plays a pivotal role in the development of CHD by altering the dynamics of vascular cell proliferation. In addition, single nucleotide polymorphisms (SNPs) in CDKN2BAS are associated with the risk of multiple diseases, including comprising CHD [7, 9, 11], myocardial infarction [7], type 2 diabetes [12], ischemic stroke [13], and periodontitis [14].
While many studies have demonstrated that polymorphisms in CDKN2BASare associated with the risk of CHD, few studies have focused on the effects of these alterations on susceptibility to CHD in the Han Chinese population. Therefore, we performed a case-control study to investigate the associations between these SNPs and the risk of CHD in the Han Chinese men and women.
Results
Basic information on the eight SNPs in CDKN2BAS, including chromosomal position, role, allele, minor allele frequency (MAF), and Hardy-Weinberg Equilibrium (HWE) test results are shown in Table 1. All SNPs were in HWE in the control groups (P > 0.05). We then compared the differences in frequency distributions of alleles between cases and controls using Chi-squared tests and found that the frequency of allele “G” of rs10757274 in CDKN2BAS was higher in cases, and it was associated with a 1.30-fold increased risk of CHD in men at a 5% level (OR = 1.30, 95% CI: 1.05-1.61, P = 0.018).
Table 1: Allele frequencies in cases and controls and odds ratio estimates for CHD
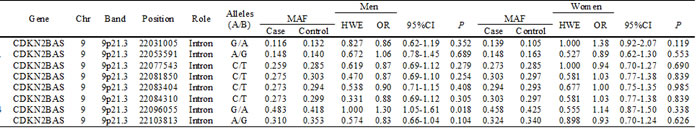
Chr: Chromosome; MAF: Minor allele frequency; HWE: Hardy-Weinberg equilibrium; OR: Odd ratio; 95% CI: 95% Confidence interval.
P values were calculated from Chi-square /Fisher's exact.
P <0.05 indicates statistical significance.
We further assessed the association between each SNP and CHD risk using unconditional logistic regression analysis after adjusting for age. The results of genetic model analyses (codominant, dominant, recessive, log-additive) are presented in Table 2. We identified that rs10757274 was associated with increased CHD risk in men under a codominant model (P = 0.042); a recessive model (GG vs AA+AG: OR = 1.70, 95% CI: 1.10-2.62, P = 0.016) and a log-additive model (OR = 1.34, 95% CI: 1.05-1.71, P = 0.019). Interestingly, rs10757274 was also associated with increased CHD risk in women under both a codominant model (P = 0.004) and a dominant model (AG+GG vs AA: OR = 2.26, 95% CI: 1.28-3.99, P = 0.004). Allele “G” of rs7865618 was also associated with increased CHD risk under a codominant model (P = 0.021). A similar trend was observed for rs7865618, which was associated with a 6.79-fold increased risk of CHD in women under a recessive model (GG vs AA+GA: OR = 8.10, 95% CI: 1.74-37.68, P = 0.006).
Table 2: Genetic models analyses of the association between CDKN2BAS SNPs and CHD susceptibility in men and women
SNP-ID |
Model |
Genotype |
Men |
Women |
||||||
Case |
Control |
OR (95% CI) |
P-value |
Case |
Control |
OR (95% CI) |
P-value |
|||
rs7865618 |
Codominant |
A/A |
227 (78.3%) |
290 (75.3%) |
1 |
127 (77%) |
240 (80%) |
1 |
0.021 |
|
G/A |
59 (20.3%) |
88 (22.9%) |
0.80 (0.52-1.23) |
0.430 |
30 (18.2%) |
57 (19%) |
1.07 (0.57-1.98) |
|||
G/G |
4 (1.4%) |
7 (1.8%) |
0.54 (0.13-2.18) |
8 (4.8%) |
3 (1%) |
8.20 (1.76-38.35) |
||||
Dominant |
A/A |
227 (78.3%) |
290 (75.3%) |
1 |
127 (77%) |
240 (80%) |
1 |
|||
G/A-G/G |
63 (21.7%) |
95 (24.7%) |
0.78 (0.52-1.18) |
0.240 |
38 (23%) |
60 (20%) |
1.37 (0.77-2.45) |
0.290 |
||
Recessive |
A/A-G/A |
286 (98.6%) |
378 (98.2%) |
1 |
157 (95.2%) |
297 (99%) |
1 |
|||
G/G |
4 (1.4%) |
7 (1.8%) |
0.57 (0.14-2.28) |
0.420 |
8 (4.8%) |
3 (1%) |
8.10 (1.74-37.68) |
0.006 |
||
Log-additive |
--- |
--- |
--- |
0.78 (0.54-1.14) |
0.200 |
--- |
--- |
1.58 (0.97-2.58) |
0.068 |
|
rs11790231 |
Codominant |
G/G |
217 (74.6%) |
281 (73.6%) |
1 |
121 (73.3%) |
208 (69.3%) |
1 |
0.520 |
|
A/G |
62 (21.3%) |
95 (24.9%) |
0.99 (0.65-1.50) |
0.380 |
39 (23.6%) |
86 (28.7%) |
0.74 (0.42-1.29) |
|||
A/A |
12 (4.1%) |
6 (1.6%) |
2.15 (0.71-6.52) |
5 (3%) |
6 (2%) |
1.23 (0.28-5.36) |
||||
Dominant |
G/G |
217 (74.6%) |
281 (73.6%) |
1 |
121 (73.3%) |
208 (69.3%) |
1 |
|||
A/G-A/A |
74 (25.4%) |
101 (26.4%) |
1.07 (0.72-1.60) |
0.720 |
44 (26.7%) |
92 (30.7%) |
0.78 (0.45-1.33) |
0.350 |
||
Recessive |
G/G-A/G |
279 (95.9%) |
376 (98.4%) |
1 |
160 (97%) |
294 (98%) |
1 |
|||
A/A |
12 (4.1%) |
6 (1.6%) |
2.15 (0.71-6.51) |
0.160 |
5 (3%) |
6 (2%) |
1.33 (0.31-5.76) |
0.700 |
||
Log-additive |
--- |
--- |
--- |
1.14 (0.81-1.60) |
0.450 |
--- |
--- |
0.85 (0.53-1.35) |
0.490 |
|
rs1412832 |
Codominant |
T/T |
161 (55.5%) |
194 (50.5%) |
1 |
84 (50.9%) |
153 (51%) |
1 |
0.760 |
|
T/C |
108 (37.2%) |
161 (41.9%) |
0.76 (0.52-1.09) |
0.250 |
72 (43.6%) |
123 (41%) |
1.02 (0.62-1.69) |
|||
C/C |
21 (7.2%) |
29 (7.5%) |
0.70 (0.36-1.37) |
9 (5.5%) |
24 (8%) |
0.71 (0.27-1.90) |
||||
Dominant |
T/T |
161 (55.5%) |
194 (50.5%) |
1 |
84 (50.9%) |
153 (51%) |
1 |
|||
T/C-C/C |
129 (44.5%) |
190 (49.5%) |
0.75 (0.53-1.06) |
0.100 |
81 (49.1%) |
147 (49%) |
0.97 (0.60-1.57) |
0.890 |
||
Recessive |
T/T-T/C |
269 (92.8%) |
355 (92.5%) |
1 |
156 (94.5%) |
276 (92%) |
1 |
|||
C/C |
21 (7.2%) |
29 (7.5%) |
0.79 (0.42-1.51) |
0.480 |
9 (5.5%) |
24 (8%) |
0.70 (0.27-1.83) |
0.470 |
||
Log-additive |
--- |
--- |
--- |
0.80 (0.61-1.05) |
0.110 |
--- |
--- |
0.92 (0.63-1.36) |
0.680 |
|
rs6475606 |
Codominant |
T/T |
156 (53.6%) |
183 (47.7%) |
1 |
77 (46.7%) |
146 (48.7%) |
1 |
0.920 |
|
C/T |
110 (37.8%) |
169 (44%) |
0.73 (0.50-1.05) |
0.210 |
76 (46.1%) |
130 (43.3%) |
0.91 (0.55-1.50) |
|||
C/C |
25 (8.6%) |
32 (8.3%) |
0.76 (0.40-1.43) |
12 (7.3%) |
24 (8%) |
0.89 (0.36-2.23) |
||||
Dominant |
T/T |
156 (53.6%) |
183 (47.7%) |
1 |
77 (46.7%) |
146 (48.7%) |
1 |
|||
C/T-C/C |
135 (46.4%) |
201 (52.3%) |
0.73 (0.52-1.04) |
0.077 |
88 (53.3%) |
154 (51.3%) |
0.90 (0.56-1.47) |
0.680 |
||
Recessive |
T/T-C/T |
266 (91.4%) |
352 (91.7%) |
1 |
153 (92.7%) |
276 (92%) |
1 |
|||
C/C |
25 (8.6%) |
32 (8.3%) |
0.88 (0.48-1.61) |
0.670 |
12 (7.3%) |
24 (8%) |
0.94 (0.39-2.27) |
0.880 |
||
Log-additive |
--- |
--- |
--- |
0.81 (0.62-1.06) |
0.120 |
--- |
--- |
0.93 (0.63-1.36) |
0.700 |
|
rs1333040 |
Codominant |
T/T |
156 (53.6%) |
188 (49.1%) |
1 |
80 (48.5%) |
148 (49.3%) |
1 |
0.930 |
|
T/C |
111 (38.1%) |
165 (43.1%) |
0.78 (0.54-1.12) |
0.380 |
73 (44.2%) |
128 (42.7%) |
0.91 (0.55-1.52) |
|||
C/C |
24 (8.2%) |
30 (7.8%) |
0.81 (0.43-1.55) |
12 (7.3%) |
24 (8%) |
0.90 (0.36-2.24) |
||||
Dominant |
T/T |
156 (53.6%) |
188 (49.1%) |
1 |
80 (48.5%) |
148 (49.3%) |
1 |
|||
T/C-C/C |
135 (46.4%) |
195 (50.9%) |
0.78 (0.55-1.11) |
0.170 |
85 (51.5%) |
152 (50.7%) |
0.91 (0.56-1.48) |
0.710 |
||
Recessive |
T/T-T/C |
267 (91.8%) |
353 (92.2%) |
1 |
153 (92.7%) |
276 (92%) |
1 |
|||
C/C |
24 (8.2%) |
30 (7.8%) |
0.91 (0.49-1.69) |
0.760 |
12 (7.3%) |
24 (8%) |
0.94 (0.39-2.27) |
0.880 |
||
Log-additive |
--- |
--- |
--- |
0.85 (0.65-1.11) |
0.230 |
--- |
--- |
0.93 (0.64-1.37) |
0.720 |
|
rs1537370 |
Codominant |
T/T |
156 (53.6%) |
185 (48%) |
1 |
77 (46.7%) |
146 (48.7%) |
1 |
0.920 |
|
C/T |
111 (38.1%) |
170 (44.2%) |
0.73 (0.51-1.06) |
0.240 |
76 (46.1%) |
130 (43.3%) |
0.91 (0.55-1.50) |
|||
C/C |
24 (8.2%) |
30 (7.8%) |
0.79 (0.41-1.51) |
12 (7.3%) |
24 (8%) |
0.89 (0.36-2.23) |
||||
Dominant |
T/T |
156 (53.6%) |
185 (48%) |
1 |
77 (46.7%) |
146 (48.7%) |
1 |
|||
C/T-C/C |
135 (46.4%) |
200 (52%) |
0.74 (0.53-1.05) |
0.093 |
88 (53.3%) |
154 (51.3%) |
0.90 (0.56-1.47) |
0.680 |
||
Recessive |
T/T-C/T |
267 (91.8%) |
355 (92.2%) |
1 |
153 (92.7%) |
276 (92%) |
1 |
|||
C/C |
24 (8.2%) |
30 (7.8%) |
0.91 (0.49-1.70) |
0.760 |
12 (7.3%) |
24 (8%) |
0.94 (0.39-2.27) |
0.880 |
||
Log-additive |
--- |
--- |
--- |
0.82 (0.62-1.07) |
0.150 |
--- |
--- |
0.93 (0.63-1.36) |
0.700 |
|
rs10757274 |
Codominant |
A/A |
82 (28.2%) |
130 (33.8%) |
1 |
39 (23.6%) |
102 (34%) |
1 |
0.004 |
|
A/G |
137 (47.1%) |
188 (48.8%) |
1.16 (0.78-1.72) |
0.042 |
101 (61.2%) |
141 (47%) |
2.60 (1.44-4.71) |
|||
G/G |
72 (24.7%) |
67 (17.4%) |
1.86 (1.13-3.05) |
25 (15.2%) |
57 (19%) |
1.52 (0.72-3.23) |
||||
Dominant |
A/A |
82 (28.2%) |
130 (33.8%) |
1 |
39 (23.6%) |
102 (34%) |
1 |
|||
A/G-G/G |
209 (71.8%) |
255 (66.2%) |
1.33 (0.91-1.93) |
0.130 |
126 (76.4%) |
198 (66%) |
2.26 (1.28-3.99) |
0.004 |
||
Recessive |
A/A-A/G |
219 (75.3%) |
318 (82.6%) |
1 |
140 (84.8%) |
243 (81%) |
1 |
|||
G/G |
72 (24.7%) |
67 (17.4%) |
1.70 (1.10-2.62) |
0.016 |
25 (15.2%) |
57 (19%) |
0.81 (0.43-1.51) |
0.500 |
||
Log-additive |
--- |
--- |
--- |
1.34 (1.05-1.71) |
0.019 |
--- |
--- |
1.31 (0.92-1.87) |
0.130 |
|
rs1333042 |
Codominant |
G/G |
139 (47.9%) |
162 (42.6%) |
1 |
74 (44.9%) |
130 (43.3%) |
1 |
0.810 |
|
G/A |
122 (42.1%) |
168 (44.2%) |
0.88 (0.61-1.27) |
0.120 |
75 (45.5%) |
136 (45.3%) |
0.87 (0.52-1.45) |
|||
A/A |
29 (10%) |
50 (13.2%) |
0.55 (0.31-0.97) |
16 (9.7%) |
34 (11.3%) |
0.81 (0.36-1.84) |
||||
Dominant |
G/G |
139 (47.9%) |
162 (42.6%) |
1 |
74 (44.9%) |
130 (43.3%) |
1 |
|||
G/A-A/A |
151 (52.1%) |
218 (57.4%) |
0.79 (0.56-1.12) |
0.190 |
91 (55.1%) |
170 (56.7%) |
0.86 (0.53-1.39) |
0.530 |
||
Recessive |
G/G-G/A |
261 (90%) |
330 (86.8%) |
1 |
149 (90.3%) |
266 (88.7%) |
1 |
|||
A/A |
29 (10%) |
50 (13.2%) |
0.59 (0.34-1.01) |
0.050 |
16 (9.7%) |
34 (11.3%) |
0.87 (0.40-1.90) |
0.730 |
||
Log-additive |
--- |
--- |
--- |
0.78 (0.61-1.01) |
0.058 |
--- |
--- |
0.89 (0.62-1.28) |
0.530 |
|
OR: Odd ratio; 95% CI: 95% Confidence.
P values were calculated from the Wald test.
P < 0.05 indicates statistical significance.
Linkage Disequilibrium (LD) analysis demonstrated that two SNPs (rs10757274 and rs1333042) in CDKN2BAS showed strong linkage (Figure 1). Furthermore, the haplotype AA (rs10757274 and rs1333042) was significantly associated with a decreased risk of CHD in men (OR = 0.72, 95% CI: 0.55 - 0.95, P = 0.022; Table 3).
Table 3: Haplotype frequencies and association with the risk of CHD in men
Chr |
Gene |
SNPs |
Haplotype |
F_A |
F_U |
OR (95% CI) |
P-value |
9 |
CDKN2BAS |
rs10757274|rs1333042 |
GG |
0.481 |
0.415 |
1 |
--- |
AA |
0.309 |
0.350 |
0.72 (0.55 - 0.95) |
0.022 |
|||
AG |
0.208 |
0.232 |
0.75 (0.54 - 1.04) |
0.083 |
F_A: Frequency in cases; F_U: Frequency in controls; OR: Odd ratio; 95% CI: 95% Confidence.
P values were calculated using the Wald test.
P < 0.05 indicates statistical significance.
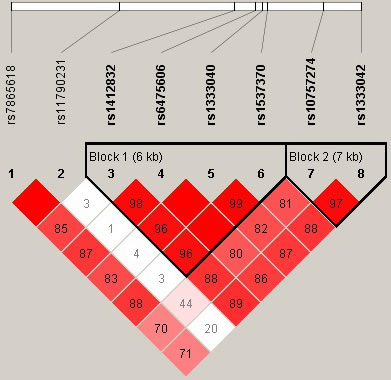
Figure 1: Haplotype block map for the eight SNPs in CDKN2BAS in men. Block 1 includes rs1412832, rs6475606, rs1333040 and rs1537370; Block 2 includes rs10757274 and rs1333042. The LD between two SNPs is standardized D’.
Additionally, we also investigated the associations between CDKN2BAS polymorphisms and CHD-related biochemical indicators, including triglyceride (TG), total cholesterol (TC), high-density lipoprotein cholesterol (HDL-C), low density lipoprotein cholesterol (LDL-C), creatinine (CRE), neutrophil (NEU), platelet distribution width (PDW) as shown in Table 4. The levels of HLD-C with rs6475606 (P = 0.022) and rs1333040 (P = 0.045) in men, and rs1412832 (P = 0.017) in women, were different between groups. Levels of CRE with rs7865618 (P = 0.035) and rs1412832 (P = 0.015) in men, and rs11790231 and rs6475606 (P = 0.033) in women, were also different between groups. In addition, the levels of NEU with three SNPs (rs7865618: P = 0.037, rs11790231: P = 0.018 and rs6475606: P = 0.027) and the levels PDW with four SNPs (rs7865618: P = 0.029, rs1333040: P = 0.047, rs1537370: P = 0.020 and rs10757274: P = 0.047) were different between groups, but only in women.
Table 4: Lipid levels according to genotype in men and women
Sex |
SNP-ID |
Genotype (N) |
TG |
TC |
HDL-C |
LDL-C |
CRE |
NEU |
PDW |
|||||||
Mean ± SD |
P |
Mean ± SD |
P |
Mean ± SD |
P |
Mean ± SD |
P |
Mean ± SD |
P |
Mean ± SD |
P |
Mean ± SD |
P |
|||
Men |
rs7865618 |
CC(137) |
1.70±0.11 |
3.90±0.09 |
1.20±0.02 |
1.84±0.08 |
86.82±1.75 |
5.24±0.27 |
14.54±0.25 |
|||||||
TT(19) |
1.74±0.27 |
0.524 |
3.95±0.43 |
0.607 |
1.15±0.07 |
0.549 |
1.87±0.24 |
0.927 |
85.63±3.18 |
0.035 |
5.18±0.69 |
0.699 |
14.16±0.59 |
0.496 |
||
CT(119) |
1.93±0.19 |
4.04±0.11 |
1.09±0.02 |
1.88±0.07 |
80.58±1.72 |
5.56±0.29 |
14.11±0.27 |
|||||||||
rs11790231 |
GG(4) |
1.54±0.46 |
3.54±0.77 |
1.00±0.01 |
1.75±0.52 |
74.63±9.56 |
4.67±0.63 |
13.03±1.11 |
||||||||
AA(217) |
1.77±0.12 |
0.752 |
3.96±0.08 |
0.745 |
1.10±0.02 |
0.644 |
1.87±0.06 |
0.953 |
84.50±1.38 |
0.546 |
5.22±0.20 |
0.210 |
14.22±0.20 |
0.277 |
||
GA(53) |
1.95±0.20 |
4.00±0.16 |
1.08±0.04 |
1.84±0.09 |
82.90±2.22 |
6.02±0.52 |
14.82±0.39 |
|||||||||
rs1412832 |
GG(203) |
1.79±0.12 |
3.95±0.08 |
1.10±0.02 |
1.86±0.06 |
83.94±1.33 |
5.57±0.23 |
14.41±0.20 |
||||||||
AA(12) |
1.46±0.15 |
0.659 |
3.80±0.26 |
0.803 |
0.99±0.06 |
0.281 |
1.88±0.20 |
0.994 |
99.43±11.89 |
0.015 |
5.42±0.81 |
0.161 |
12.67±0.74 |
0.186 |
||
AG(60) |
1.92±0.17 |
4.03±0.17 |
1.10±0.03 |
1.86±0.11 |
81.30±1.59 |
4.68±0.33 |
14.28±0.36 |
|||||||||
rs6475606 |
CC(20) |
1.76±0.24 |
3.64±0.25 |
0.95±0.04 |
1.72±0.13 |
79.66±4.68 |
6.02±1.05 |
14.00±0.61 |
||||||||
TC(101) |
1.78±0.15 |
0.979 |
4.05±0.13 |
0.371 |
1.12±0.03 |
0.022 |
1.91±0.08 |
0.651 |
84.30±2.00 |
0.613 |
5.76±0.30 |
0.131 |
14.11±0.28 |
0.519 |
||
TT(153) |
1.82±0.15 |
3.95±0.09 |
1.10±0.02 |
1.85±0.08 |
84.36±1.52 |
5.05±0.24 |
14.50±0.24 |
|||||||||
rs1333040 |
CC(24) |
1.77±0.20 |
3.78±0.22 |
0.98±0.05 |
1.79±0.13 |
86.96±6.80 |
5.97±0.90 |
13.51±0.59 |
||||||||
TT(147) |
1.79±0.15 |
0.992 |
3.94±0.09 |
0.638 |
1.11±0.02 |
0.045 |
1.86±0.08 |
0.907 |
84.74±1.55 |
0.501 |
5.12±0.25 |
0.320 |
14.61±0.25 |
0.148 |
||
CT(104) |
1.81±0.15 |
4.02±0.13 |
1.11±0.03 |
1.88±0.08 |
82.46±1.63 |
5.59±0.29 |
14.09±0.26 |
|||||||||
rs1537370 |
CC(23) |
1.64±0.16 |
3.68±0.21 |
0.98±0.05 |
1.79±0.13 |
86.34±7.09 |
6.05±0.93 |
13.43±0.62 |
||||||||
TC(105) |
1.84±0.15 |
0.867 |
4.04±0.13 |
0.410 |
1.11±0.03 |
0.055 |
1.88±0.08 |
0.915 |
82.62±1.62 |
0.592 |
5.57±0.29 |
0.299 |
14.10±0.25 |
0.134 |
||
TT(147) |
1.79±0.15 |
3.94±0.09 |
1.11±0.02 |
1.86±0.08 |
84.74±1.55 |
5.12±0.25 |
14.61±0.25 |
|||||||||
rs10757274 |
CC(23) |
1.64±0.16 |
3.68±0.21 |
0.98±0.05 |
1.79±0.13 |
86.34±7.09 |
6.05±0.93 |
13.43±0.62 |
||||||||
TT(147) |
1.79±0.15 |
0.867 |
3.94±0.09 |
0.410 |
1.11±0.02 |
0.055 |
1.86±0.08 |
0.915 |
84.74±1.55 |
0.592 |
5.12±0.25 |
0.299 |
14.61±0.25 |
0.134 |
||
CT(105) |
1.84±0.15 |
4.04±0.13 |
1.11±0.03 |
1.88±0.08 |
82.62±1.62 |
5.57±0.29 |
14.10±0.25 |
|||||||||
rs1333042 |
GG(69) |
1.74±0.15 |
3.95±0.13 |
1.15±0.03 |
1.94±0.15 |
84.05±1.95 |
4.85±0.36 |
14.49±0.35 |
||||||||
AA(78) |
1.69±0.16 |
0.650 |
3.88±0.15 |
0.766 |
1.07±0.03 |
0.098 |
1.81±0.09 |
0.644 |
82.23±2.22 |
0.591 |
5.71±0.40 |
0.258 |
14.24±0.31 |
0.856 |
||
AG(128) |
1.90±0.17 |
4.01±0.10 |
1.08±0.02 |
1.84±0.06 |
85.10±1.84 |
5.43±0.26 |
14.28±0.26 |
|||||||||
Women |
rs7865618 |
CC(78) |
1.88±0.17 |
4.33±0.11 |
1.16±0.03 |
1.96±0.07 |
72.77±1.89 |
4.01±0.24 |
13.77±0.36 |
|||||||
TT(14) |
2.09±0.42 |
0.320 |
4.78±0.30 |
0.213 |
1.24±0.07 |
0.264 |
2.24±0.23 |
0.383 |
70.62±4.72 |
0.849 |
5.94±1.02 |
0.037 |
16.28±0.82 |
0.029 |
||
CT(62) |
1.62±0.09 |
4.23±0.14 |
1.22±0.03 |
2.05±0.10 |
70.82±3.47 |
4.32±0.34 |
13.97±0.41 |
|||||||||
rs11790231 |
GG(7) |
1.41±0.13 |
4.49±0.28 |
1.12±0.10 |
2.15±0.12 |
89.85±25.84 |
6.83±0.87 |
14.08±1.01 |
||||||||
AA(120) |
1.72±0.12 |
0.119 |
4.24±0.10 |
0.163 |
1.19±0.02 |
0.759 |
1.99±0.07 |
0.615 |
71.83±1.53 |
0.033 |
4.14±0.24 |
0.018 |
14.09±0.31 |
0.986 |
||
GA(28) |
2.22±0.26 |
4.66±0.22 |
1.20±0.05 |
2.12±0.14 |
66.94±1.65 |
4.28±0.38 |
13.98±0.57 |
|||||||||
rs1412832 |
GG(113) |
1.84±0.13 |
4.35±0.11 |
1.16±0.02 |
2.02±0.07 |
71.43±2.13 |
4.44±0.25 |
13.69±0.31 |
||||||||
AA(5) |
1.59±0.13 |
0.769 |
5.26±0.42 |
0.082 |
1.39±0.12 |
0.017 |
2.70±0.35 |
0.079 |
67.38±4.81 |
0.788 |
4.27±0.99 |
0.490 |
14.90±1.40 |
0.051 |
||
AG(37) |
1.69±0.15 |
4.15±0.13 |
1.26±0.04 |
1.94±0.11 |
73.57±3.07 |
3.86±0.35 |
15.13±0.49 |
|||||||||
rs6475606 |
CC(8) |
2.22±0.44 |
5.00±0.21 |
1.27±0.08 |
2.45±0.21 |
89.42±22.61 |
6.44±0.98 |
14.23±0.92 |
||||||||
TC(68) |
1.91±0.20 |
0.291 |
4.33±0.13 |
0.179 |
1.19±0.03 |
0.681 |
1.99±0.08 |
0.212 |
69.15±1.62 |
0.033 |
3.97±0.28 |
0.027 |
13.38±0.37 |
0.065 |
||
TT(79) |
1.65±0.09 |
4.26±0.12 |
1.19±0.03 |
2.01±0.08 |
72.22±1.96 |
4.35±0.30 |
14.63±0.38 |
|||||||||
rs1333040 |
CC(11) |
1.99±0.34 |
4.82±0.19 |
1.23±0.07 |
2.33±0.17 |
82.56±17.23 |
5.93±0.82 |
14.18±0.80 |
||||||||
TT(73) |
1.62±0.09 |
0.276 |
4.25±0.13 |
0.248 |
1.19±0.03 |
0.838 |
2.00±0.09 |
0.326 |
72.20±2.13 |
0.172 |
4.28±0.31 |
0.068 |
14.73±0.40 |
0.047 |
||
CT(71) |
1.94±0.20 |
4.34±0.12 |
1.18±0.03 |
1.19±0.08 |
69.74±1.50 |
4.05±0.28 |
13.39±0.36 |
|||||||||
rs1537370 |
CC(11) |
1.99±0.34 |
4.82±0.19 |
1.23±0.07 |
2.33±0.17 |
82.56±17.23 |
5.93±0.82 |
14.18±0.80 |
||||||||
TC(69) |
1.95±0.20 |
0.263 |
4.36±0.12 |
0.221 |
1.19±0.03 |
0.850 |
2.00±0.08 |
0.327 |
69.40±1.55 |
0.152 |
3.96±0.27 |
0.050 |
13.27±0.36 |
0.020 |
||
TT(75) |
1.62±0.09 |
4.23±0.13 |
1.19±0.03 |
2.00±0.09 |
72.42±2.06 |
4.36±0.31 |
14.78±0.39 |
|||||||||
rs10757274 |
CC(11) |
1.99±0.34 |
4.82±0.19 |
1.23±0.07 |
2.33±0.17 |
82.56±17.23 |
5.93±0.82 |
14.18±0.80 |
||||||||
TT(73) |
1.62±0.09 |
0.276 |
4.25±0.13 |
0.248 |
1.19±0.03 |
0.838 |
2.00±0.09 |
0.326 |
72.20±2.13 |
0.172 |
4.28±0.31 |
0.068 |
14.73±0.40 |
0.047 |
||
CT(71) |
1.94±0.20 |
4.34±0.12 |
1.18±0.03 |
1.19±0.08 |
69.74±1.50 |
4.05±0.28 |
13.39±0.36 |
|||||||||
rs1333042 |
GG(24) |
1.62±0.18 |
3.95±0.23 |
1.24±0.06 |
1.83±0.16 |
70.55±3.08 |
4.13±0.55 |
15.27±0.67 |
||||||||
AA(36) |
1.99±0.21 |
0.528 |
4.52±0.14 |
0.116 |
1.17±0.03 |
0.554 |
2.16±0.11 |
0.223 |
74.91±5.50 |
0.608 |
4.68±0.44 |
0.584 |
13.45±0.60 |
0.113 |
||
AG(95) |
1.77±0.14 |
4.36±0.11 |
1.19±0.03 |
2.02±0.07 |
70.93±1.70 |
4.20±0.26 |
14.03±0.31 |
TG: Triglyceride; TC: Total cholesterol; HDL-C: High-density lipoprotein cholesterol; LDL-C: Low density lipoprotein cholesterol; CRE: Creatinine; NEU: Neutrophil;
PDW: Platelet distribution width
DISCUSSION
The goal of our case-control study was to explore the associations of eight polymorphisms in CDKN2BAS with the risk of CHD in a Han Chinese population. We found that rs10757274 was associated with an increased risk of CHD both in men and women. However, rs7865618 was correlated with an increased risk of CHD only in women. In addition, we also found that the haplotype “AA” (rs10757274 and rs1333042) of CDKN2BAS was associated with a decreased CHD risk in men.
Previous studies have demonstrated that CDKN2BAS transcript levels show a strong correlation with the severity of atherosclerosis. In addition, the modulation of CDKN2BAS expression influences CHD susceptibility. The function of CDKN2BAS is unknown, but it regulates the expression of CDKN2A and CDKN2B, which encode cyclin-dependent kinase inhibitors 2A and 2B, indicating a regulatory role for CDKN2BAS in cellular proliferation. Polymorphisms at the 9p21 region may induce higher expression of the CDKN2BAS transcript, thereby inhibiting the expression of CDKN2A and CDKN2B.
Several studies have demonstrated a strong association of rs10757274 with CHD in Pakistani [9], Caucasian [15], and South-West Iranian [16] population. Our results are consistent with these previous findings d. However, one previous study found no association of rs10757274 with CHD in a Han Chinese population (Shenzhen) [17].The differences between our study and this study are likely due to differences in the two study populations. How rs10757274 affects the risk of CHD is unclear, but this SNP may regulate the expression of EU741058 and p16INK4a, which modulate the risk of developing CHD [18]. Some studies have also suggested that rs10757274impairs the mechanical properties of the arterial wall and thus influences vascular diseases [19].
It was previously shown that rs1333042 is associated with the risk of CHD in the Han Chinese [20, 21] and Saudi populations [7], which is inconsistent with our findings. The differences in these studies may be explained by ethnic differences, environment, or lifestyle that also affected the development of CHD. We also observed that rs7865618 was associated with CHD risk in women. However, this SNP has previously only been found to interact with other SNPs to affect the development of CHD [6]. In future studies, we will verify our results using a larger sample size.
We didn’t observe any association between rs1333040 and the risk of CHD, which is consistent with the studies by Cao et al. [21] and Golabgir Khademi et al. [22]. This SNP was significantly associated with the risk of CHD in a Northern Indian population [23] and in African American women [24]. Genetic variation and differences in life styles among populations probably explain the population disparities in the association of this SNP and susceptibility to CHD. Furthermore, recent studies have reported that the levels of TG, TC, HDL-C, LDL-C, CRE, NEU and PDW effectively predict the risk of CHD [25-28]. Interestingly, we also demonstrated that SNPs in CDKN2BAS were correlated with the levels of these biochemical indicators and differed between cases and controls.
Several limitations should be acknowledged in the present study. First, the sample size was relatively small and the participants were limited to Chinese ethnicity. Second, there were differences in some clinical characteristics between the patients and controls. Although several confounders have been adjusted for the statistical analyses, we could not completely eliminate the potential influences of these factors on the results. Finally, the biological mechanism of genetic variants in CDKN2BAS was not investigated in this study. It will be important to follow up and validate our findings with larger sample sizes.
In conclusion, our results suggest that, in a Chinese Han population, rs10757274 in CDKN2BAS is associated with the risk of CHD both in men and women, rs7865618 is correlated with an increased risk of CHD only in women, and the haplotype AA (rs10757274 and rs1333042) of CDKN2BAS is associated with a decreased CHD risk in men. Thus, these SNPs could have clinical importance as pre-diagnostic markers. Further study is required to determine the functional effects of these SNPs and validate these findings in larger populations.
Materials and Methods
Ethics statement
Written informed consent was obtained from all study participants before the interview. This study protocol was approved by the Ethical Committee of the Yanan University Affiliated Hospital and the First Affiliated Hospital of Xi’an Jiaotong University, and complied with the World Medical Association Declaration of Helsinki.
Study participants
The study included 676 men (291 CHD cases with a mean age of 60 years and 385 healthy controls with a mean age of 48 years) and 465 women (165 CHD cases with a mean age of 64 years and 300 healthy controls with a mean age of 50 years). All CHD cases were recruited from the Cardiovascular Internal Medicine Department of Yanan University Affiliated Hospital between February 2014 and April 2015. The 685 healthy controls were randomly selected from physical examination center of the same hospital during the same period. The inclusion and exclusion criteria for participants were as follows: First, all subjects were of the ethnic Han origin and not related to each other. Second, all participants diagnosis we based on standardized electrocardiogram, echocardiography, blood tests and coronary angiography and judged by two or three independent cardiologists. Third, all individuals were excluded from the study if they had other cardiac diseases (congenital heart disease, cardiomyopathy, or rheumatic heart disease), diabetes, hypertension, or severe liver or kidney disease.s, Patients who had previously received angioplasty, intravenous thrombolysis, coronary artery stents, or coronary artery bypass surgery were also excluded. Basic characteristics of all enrolled controls were collected with a standard epidemiological questionnaires conducted by well-trained interviewers. The cases information was collected through consultation with treating physicians or from medical chart review. Peripheral venous blood (5 ml) was collected from each participant using vacutainer tubes containing ethylene diamine tetra-acetic acid (EDTA) and then stored at -80°C. A clinical examination at which a blood sample was drawn for routine analysis of blood levels, biochemical tests, coagulation function, and genetic analyses.
SNP selection and genotyping
The eight SNPs in CDKN2BAS (rs7865618, rs1179023, rs1412832, rs6475606, rs1333040, rs1537370, rs10757274 and rs1333042) were selected from previous reports for their association with CHD [9, 21, 23, 29, 30]. The minor allele frequency of each SNP was > 5% in the HapMap of the Chinese Han Beijing (CHB) population. Genomic DNA was extracted from whole blood using the GoldMag-Mini Whole Blood Genomic DNA Purification Kit according to the manufacturer’s protocol (GoldMag. Co. Ltd., Xi’an, China). DNA concentration and purity were evaluated using a spectrophotometer (NanoDrop 2000; Thermo Fisher Scientific, Waltham, MA, USA). Polymerase chain reaction (PCR) and extension primers for the SNPs were designed using the Sequenom MassARRAY Assay Design 3.0 software (Sequenom, San Diego, CaliforniaCA, USA). Genotyping was performed using the Sequenom MassARRAY platform (Sequenom, San Diego, CA, USA) according to the standard instructions. Sequenom Typer 4.0 software was used for data management and analyses.
Statistical analysis
SPSS 19.0 (SPSS Inc., Chicago, IL, USA) and Microsoft Excel (Microsoft Corp., Redmond, WA, USA) were used for statistical analyses. Genotypic frequencies in controls (men and women) were tested for departure from HWE using a Fisher’s exact test. The allelic frequencies were compared between cases and controls by Chi-squared test/Fisher’s exact test, and the relative risk was estimated by odd ratios (ORs) and 95% confidence intervals (CIs). The genetic model analyses (codominant, dominant, recessive, log-additive) were applied using PLINK software to assess the significance of SNPs. ORs and 95% CIs were calculated using unconditional logistic regression analysis with adjustment for age. The p values were calculated with the Wald test. Haploview software (version 4.2) was used for analyses of the pairwise linkage disequilibrium (LD), haplotype structure and genetic association at polymorphism loci. All p values were two-sided, and p < 0.05 is considered statistically significant.
Acknowledgments
We thank all authors for their contributions and support. We are grateful to all participants in the study who provided blood samples. We would also like to thank the hospital staff who contributed to data collection for this study.
Conflicts of interest
The authors declare that there are no conflicts of interest.
Grant support
This work is supported by the National Natural Science Foundations (No. 81200098) and the Key Science and Technology Program of Shaanxi Province (No. 2014K11-03-04-06).
References
1. Lopez AD, Mathers CD, Ezzati M, Jamison DT and Murray CJ. Global and regional burden of disease and risk factors, 2001: systematic analysis of population health data. Lancet. 2006; 367(9524):1747-1757.
2. Sayols-Baixeras S, Lluis-Ganella C, Lucas G and Elosua R. Pathogenesis of coronary artery disease: focus on genetic risk factors and identification of genetic variants. The application of clinical genetics. 2014; 7:15-32.
3. Topol EJ, Smith J, Plow EF and Wang QK. Genetic susceptibility to myocardial infarction and coronary artery disease. Human molecular genetics. 2006; 15 Spec No 2:R117-123.
4. Ozaki K and Tanaka T. Molecular genetics of coronary artery disease. Journal of human genetics. 2016; 61(1):71-77.
5. Marenberg ME, Risch N, Berkman LF, Floderus B and de Faire U. Genetic susceptibility to death from coronary heart disease in a study of twins. The New England journal of medicine. 1994; 330(15):1041-1046.
6. Samani NJ, Erdmann J, Hall AS, Hengstenberg C, Mangino M, Mayer B, Dixon RJ, Meitinger T, Braund P, Wichmann HE, Barrett JH, Konig IR, Stevens SE, et al. Genomewide association analysis of coronary artery disease. The New England journal of medicine. 2007; 357(5):443-453.
7. AbdulAzeez S, Al-Nafie AN, Al-Shehri A, Borgio JF, Baranova EV, Al-Madan MS, Al-Ali RA, Al-Muhanna F, Al-Ali A, Al-Mansori M, Ibrahim MF, Asselbergs FW, Keating B, et al. Intronic Polymorphisms in the CDKN2B-AS1 Gene Are Strongly Associated with the Risk of Myocardial Infarction and Coronary Artery Disease in the Saudi Population. International journal of molecular sciences. 2016; 17(3).
8. Wellcome Trust Case Control C. Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls. Nature. 2007; 447(7145):661-678.
9. Nawaz SK, Noreen A, Rani A, Yousaf M and Arshad M. Association of the rs10757274 SNP with coronary artery disease in a small group of a Pakistani population. Anatolian journal of cardiology. 2015; 15(9):709-715.
10. Holdt LM, Sass K, Gabel G, Bergert H, Thiery J and Teupser D. Expression of Chr9p21 genes CDKN2B (p15(INK4b)), CDKN2A (p16(INK4a), p14(ARF)) and MTAP in human atherosclerotic plaque. Atherosclerosis. 2011; 214(2):264-270.
11. Huang Y, Ye H, Hong Q, Xu X, Jiang D, Xu L, Dai D, Sun J, Gao X and Duan S. Association of CDKN2BAS polymorphism rs4977574 with coronary heart disease: a case-control study and a meta-analysis. International journal of molecular sciences. 2014; 15(10):17478-17492.
12. Wang MH, Li J, Yeung VS, Zee BC, Yu RH, Ho S and Waye MM. Four pairs of gene-gene interactions associated with increased risk for type 2 diabetes (CDKN2BAS-KCNJ11), obesity (SLC2A9-IGF2BP2, FTO-APOA5), and hypertension (MC4R-IGF2BP2) in Chinese women. Meta gene. 2014; 2:384-391.
13. Heckman MG, Soto-Ortolaza AI, Diehl NN, Rayaprolu S, Brott TG, Wszolek ZK, Meschia JF and Ross OA. Genetic variants associated with myocardial infarction in the PSMA6 gene and Chr9p21 are also associated with ischaemic stroke. European journal of neurology. 2013; 20(2):300-308.
14. Schaefer AS, Richter GM, Dommisch H, Reinartz M, Nothnagel M, Noack B, Laine ML, Folwaczny M, Groessner-Schreiber B, Loos BG, Jepsen S and Schreiber S. CDKN2BAS is associated with periodontitis in different European populations and is activated by bacterial infection. Journal of medical genetics. 2011; 48(1):38-47.
15. McPherson R, Pertsemlidis A, Kavaslar N, Stewart A, Roberts R, Cox DR, Hinds DA, Pennacchio LA, Tybjaerg-Hansen A, Folsom AR, Boerwinkle E, Hobbs HH and Cohen JC. A common allele on chromosome 9 associated with coronary heart disease. Science. 2007; 316(5830):1488-1491.
16. Foroughmand AM, Nikkhah E, Galehdari H and Jadbabaee MH. Association Study between Coronary Artery Disease and rs1333049 and rs10757274 Polymorphisms at 9p21 Locus in South-West Iran. Cell journal. 2015; 17(1):89-98.
17. Liu YH, Zhou YW, Yang JA, Tu ZG, Ji SY, Huang ZY and Zhou ZJ. Gene polymorphisms associated with susceptibility to coronary artery disease in Han Chinese people. Genetics and molecular research : GMR. 2014; 13(2):2619-2627.
18. Dehghan A, van Hoek M, Sijbrands EJ, Oostra BA, Hofman A, van Duijn CM and Witteman JC. Lack of association of two common polymorphisms on 9p21 with risk of coronary heart disease and myocardial infarction; results from a prospective cohort study. BMC medicine. 2008; 6:30.
19. Bjorck HM, Lanne T, Alehagen U, Persson K, Rundkvist L, Hamsten A, Dahlstrom U and Eriksson P. Association of genetic variation on chromosome 9p21.3 and arterial stiffness. Journal of internal medicine. 2009; 265(3):373-381.
20. Gui L, Wu F, Han X, Dai X, Qiu G, Li J, Wang J, Zhang X, Wu T and He M. A multilocus genetic risk score predicts coronary heart disease risk in a Chinese Han population. Atherosclerosis. 2014; 237(2):480-485.
21. Cao XL, Yin RX, Huang F, Wu JZ and Chen WX. Chromosome 9p21 and ABCA1 Genetic Variants and Their Interactions on Coronary Heart Disease and Ischemic Stroke in a Chinese Han Population. International journal of molecular sciences. 2016; 17(4).
22. Golabgir Khademi K, Foroughmand AM, Galehdari H, Yazdankhah S, Pourmahdi Borujeni M, Shahbazi Z and Dinarvand P. Association Study of rs1333040 and rs1004638 Polymorphisms in the 9p21 Locus with Coronary Artery Disease in Southwest of Iran. Iranian biomedical journal. 2016; 20(2):122-127.
23. Kumar J, Yumnam S, Basu T, Ghosh A, Garg G, Karthikeyan G and Sengupta S. Association of polymorphisms in 9p21 region with CAD in North Indian population: replication of SNPs identified through GWAS. Clinical genetics. 2011; 79(6):588-593.
24. Beckie TM, Groer MW and Beckstead JW. The relationship between polymorphisms on chromosome 9p21 and age of onset of coronary heart disease in black and white women. Genetic testing and molecular biomarkers. 2011; 15(6):435-442.
25. Sharrett AR, Ballantyne CM, Coady SA, Heiss G, Sorlie PD, Catellier D, Patsch W and Atherosclerosis Risk in Communities Study G. Coronary heart disease prediction from lipoprotein cholesterol levels, triglycerides, lipoprotein(a), apolipoproteins A-I and B, and HDL density subfractions: The Atherosclerosis Risk in Communities (ARIC) Study. Circulation. 2001; 104(10):1108-1113.
26. Zhu R, Liu C, Tang H, Zeng Q, Wang X, Zhu Z, Liu Y, Mao X and Zhong Y. Serum Galectin-9 Levels Are Associated with Coronary Artery Disease in Chinese Individuals. Mediators of inflammation. 2015; 2015:457167.
27. Papa A, Emdin M, Passino C, Michelassi C, Battaglia D and Cocci F. Predictive value of elevated neutrophil-lymphocyte ratio on cardiac mortality in patients with stable coronary artery disease. Clinica chimica acta; international journal of clinical chemistry. 2008; 395(1-2):27-31.
28. Ravindran R and Krishnan LK. Increased platelet cholesterol and decreased percentage volume of platelets as a secondary risk factor for coronary artery disease. Pathophysiology of haemostasis and thrombosis. 2007; 36(1):45-51.
29. Baudhuin LM. Genetics of coronary artery disease: focus on genome-wide association studies. American journal of translational research. 2009; 1(3):221-234.
30. Erdmann J, Willenborg C, Nahrstaedt J, Preuss M, Konig IR, Baumert J, Linsel-Nitschke P, Gieger C, Tennstedt S, Belcredi P, Aherrahrou Z, Klopp N, Loley C, et al. Genome-wide association study identifies a new locus for coronary artery disease on chromosome 10p11.23. European heart journal. 2011; 32(2):158-168.