
INTRODUCTION
Cancer of the head and neck region is a frequent and worldwide health burden with 61.760 estimated new cases and 13.190 estimated deaths in the United States for 2016 alone [1]. More than 90% of these tumors are squamous cell carcinomas (HNSCC) occurring in the oral and nasal cavity, pharynx, and larynx [2]. Tobacco smoking and alcohol consumption are well known carcinogenic risk factors, however, during the past two decades human papilloma virus (HPV) infections have also been related to oropharyngeal carcinomas. HPV-related tumors are a distinct entity that is associated with an overall better prognosis [3–5], and an etiological link between infection with high risk HPV serotypes 16 and 18 and the development of oropharyngeal tumors is well documented [6]. HPV-related tumors show an activation of the PI3Kinase pathway through activating mutations as well as truncating mutations and deletions of TRAF3, whereas smoking-related tumors predominantly harbor inactivating TP53 and CDKN2A mutations. Genomic instability of HNSCC in general is demonstrated by a large number of copy number alterations [7].
Two-thirds of HNSCC patients are diagnosed at an advanced stage (Stage III to IVA/B) [8]. Treatment modalities e.g. surgery, radiotherapy, and chemotherapy are often combined in a multimodality approach resulting in satisfactory loco-regional disease control [9]. Five year survival rates range between 40-60% [10]. Up to 60% of patients develop loco-regional recurrence and up to 30% will have distant failure. Recurrent or metastatic cancer represents the onset of a highly aggressive disease with dismal survival rates [11, 12].
As for other tumors, biomarkers are urgently needed to identify patients that are at risk of recurrent disease and may therefore benefit from a more aggressive first line treatment or intensified surveillance. Current strategies rely solely on established clinico-pathological parameters, e.g. tumor-stage and lymph node involvement, to detect high-risk patients. Unfavorable molecular features are presently not considered to decide for a neoadjuvant therapy or to define follow-up examinations. With the advance of immunotherapeutic agents like the immune checkpoint modulators pembrolizumab and nivolumab, new therapeutic options are available for an effective treatment of HNSCC patients; even in the setting of metastatic disease [13, 14]. This necessitates the identification of high risk patients in order intensify surveillance and enable the early initiation of palliative treatment. Furthermore, adjuvant and neoadjuvant immunotherapy protocols, which are under investigation but not yet established, might be considered for patients who are at high risk of disease-related death.
In HNSCC, HPV status has been shown to be associated with better overall survival after platinum-based radiochemotherapy [15]. Independent of HPV status, a high expression of intratumoral CD8-positive T-lymphocytes can be employed as a prognostic maker for prolonged overall survival [16]. Comprehensive molecular profiling has revealed an abundance of molecular variations in HNSCC, but the majority of alterations lack prospective validation as biomarkers in large patient cohorts [17, 18].
A promising approach to identify biomarkers in any tumor is the investigation of gene methylation status and its correlation with clinical parameters. Epigenetic deregulation, indicated by aberrant hypo- and hypermethylation, is a frequent event in human cancer and plays a key role in carcinogenesis [19, 20]. Several attempts have been made to employ methylation status as a predictive or prognostic biomarker. MGMT promoter methylation predicts progression-free survival in patients with glioblastoma multiforme undergoing therapy with temozolomide. So far, MGMT methylation is the only routinely tested methylated biomarker in clinical practice [21, 22]. Recently, QIAGEN (Hilden, Germany) and Therawis (Munich, Germany) have embarked on a cooperation to market a PITX2 methylation assay for treatment stratification in breast carcinoma patients.
The homeobox gene PITX2 is located on chromosome 4q25 and encodes four isoforms (PITX2A, PITX2B, PITX2C, PITX2D) of bicoid transcription factors involved in the development of anterior structures [23]. Mutations in PITX2 are responsible for Axenfeld-Rieger syndrome Type I, a disorder affecting the development of teeth, eyes, and abdominal structures [24]. PITX2 gene expression is regulated by the Wnt pathway and interacts with the potent transcriptional activator and mediator of cell adhesion β-catenin as well as promoting cell growth via Cyclin A1 and D2 [25, 26]. Its contribution to tumorigenesis has not been fully elucidated and may depend on the tumor type according to its role as tissue specific transcription factor. PITX2 hypermethylation, among others, has been found in breast carcinoma and acute myeloid leukemia [27–31]. In prostate cancer, hypermethylation is associated with a significant risk of disease progression [32, 33]. In contrast, PITX2 overexpression suggestive of low methylation may result in tumor progression in ovarian and thyroid carcinomas [34]. In patients with non-small cell lung cancer (NSCLC), PITX2 DNA hypomethylation was associated with an increased risk of disease progression [35].
Additionally, non-coding RNAs (ncRNA) have been reported as regulators of gene expression. Even though these RNA strands – representing the vast majority of the transcriptome – are typically not translated into proteins they significantly contribute to cell biology and development regulation [36–39]. Dysregulation of these complex mechanisms can result in a variety of diseases from neurodegenerative disorders to cancer [40]. ncRNAs are divided into two categories: small ncRNAs, like the well-studied microRNAs, and long non-coding RNAs (lncRNA), that are more than 200 nucleotides in length [41]. Recently, PANCR, a lncRNA adjacent to the PITX2 gene, has been identified. It regulates expression of the splice variant PITX2c in cardiomyocytes [42]. So far, the methylation status of PANCR has not been described in any disease.
Given the promising, albeit conflicting, data about PITX2 methylation and risk of disease progression in cancer, the methylation status of PITX2 and PANCR and its potential function as a prognostic biomarker was investigated in a well annotated HNSCC cohort.
RESULTS
Design and validation of the PITX2 and PANCR methylation assay
The genomic location of both PITX2 and PANCR assays are shown in Figure 1. The PITX2 methylation quantitative methylation-specific real-time PCR (qMSP) assay used in the presented study has previously been described in two studies [43, 44]. Methylation analysis is by necessity a quantitative approach, since normal tissue can exhibit different levels of methylation. Thus, a highly accurate quantitative assay is required. The analytical performance of the PITX2 assay has been verified earlier [43, 44].
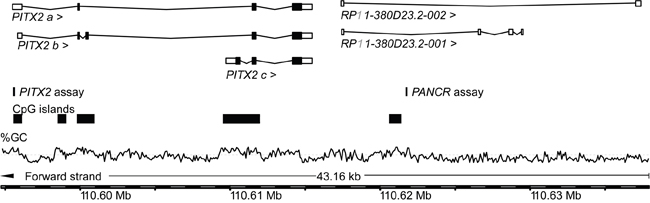
Figure 1: Genomic locations of the PITX2 and PANCR qPCR assays. The information was taken from Ensembl Homo sapiens version 82.38 (GRCh38.p3). The GC content [%] is shown with the dashed line indicating 50% GC.
FFPE samples from 399 patients with HNSCC were analyzed for PITX2 and PANCR methylation. Valid quantification was obtained from 385 (PITX2) and 394 (PANCR) samples. The distribution of DNA methylation in 5% increments for PITX2 (Figure 2A) and PANCR (Figure 2B) is depicted in histograms. The majority of samples for PITX2 showed DNA methylation between 0% and 10% (median: 4.9%), whereas PANCR was highly methylated with a median of 75.2%. Mean methylation amounted to 18.8% for PITX2 and 70.8% for PANCR.
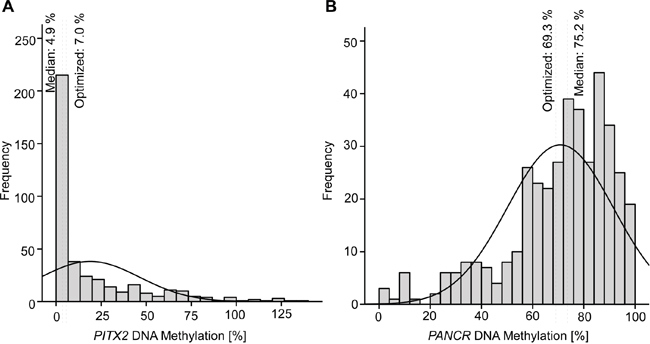
Figure 2: Histogram showing the distribution of PITX2 A. and PANCR B. values in tumor samples from HNSCC patients. DNA methylation is depicted in 5% increments on the x-axis. Methylation cut-offs used to dichotomize into low and high DNA methylation was achieved by using median values (Cut-offs for PITX2: median = 4.9%, optimized = 7.0%; and PANCR: median = 75.2%, optimized = 69.3%).
The comparison of PITX2 and PANCR DNA methylation in tumor tissue and normal adjacent tissue (NAT) revealed a significantly higher frequency of hypermethylation in cancer tissue (p < 0.001, Figure 3A, 3B).
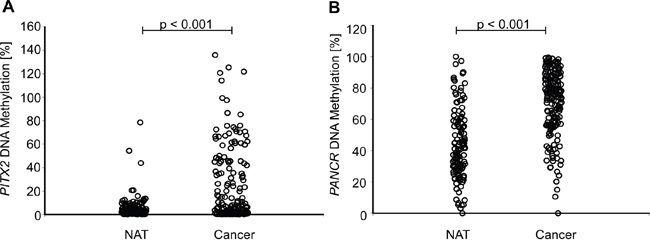
Figure 3: Comparison of PITX2 A. and PANCR B. methylation in tumor and normal adjacent tissue (NAT). HNSCC tumor tissue showed a significantly higher methylation of PITX2 and PANCR compared to the surrounding tissue.
PITX2 and PANCR hypermethylation is associated with clinicopathologic parameters
Patients characteristics were obtained for the majority of patients and were in concordance with reported demographic and clinical data of HNSCC patients [45]. A detailed description of the cohort can be found in Table 1.
Table 1: Clinicopathological data and their association/correlation with PITX2 and PANCR methylation in the cohort comprised of surgical specimens from 399 HNSCC patients
Characteristic |
No. (%) of Patients |
Median PITX2 Methylation (%) |
p-value |
Median PANCR Methylation (%) |
p-value |
|---|---|---|---|---|---|
All Patients |
399 (100) |
4.9 |
75.2 |
||
Sex |
0.24 |
0.65 |
|||
Female |
93 (23.3) |
4.1 |
74.0 |
||
Male |
306 (76.7) |
5.6 |
75.6 |
||
Age (Years) |
0.75 |
0.10 |
|||
Mean |
62.4 |
||||
Median |
62 |
||||
n ≤ Median |
168 (42.1) |
4.3 |
75.2 |
||
n > Median |
155 (38.8) |
5.0 |
75.4 |
||
Unknown |
76 (19.0) |
||||
Smoking Status |
0.048* |
0.17 |
|||
Non-Smoker |
29 (7.3) |
13.6 |
70.7 |
||
Smoker |
230 (57.6) |
4.5 |
76.0 |
||
Unknown |
140 (35.1) |
||||
Pack Years |
0.012* |
0.14 |
|||
(< 40) |
122 (30.6) |
12.6 |
72.3 |
||
(> 40) |
72 (18.0) |
4.5 |
75.2 |
||
Unknown |
205 (51.4) |
||||
Alcohol Consumption |
0.34 |
0.10 |
|||
Never |
71 (17.8) |
10.4 |
69.2 |
||
Occasionally |
40 (10.0) |
4.0 |
74.2 |
||
Abuse |
70 (17.5) |
2.2 |
78.4 |
||
Unknown |
218 (54.6) |
||||
Tumor Site |
0.002* |
0.003* |
|||
Oral Cavity |
68 (17.0) |
2.5 |
74.1 |
||
Oropharynx |
159 (39.8) |
6.7 |
79.1 |
||
Hypopharynx |
34 (8.5) |
1.9 |
76.1 |
||
Larynx |
117 (29.3) |
8.9 |
71.7 |
||
Unknown |
21 (5.3) |
||||
pT |
0.75 |
0.24 |
|||
T1/T2 |
199 (49.9) |
5.5 |
72.3 |
||
T3/T4 |
141 (35.4) |
4.7 |
76.3 |
||
Unknown |
59 (14.8) |
||||
pN |
0.22 |
<0.001* |
|||
pN neg |
169 (42.4) |
5.3 |
72.5 |
||
pN pos |
203 (50.9) |
4.3 |
78.8 |
||
Unknown |
27 (6.8) |
||||
Vascular Invasion (V) |
|||||
V neg |
202 (50.6) |
4.9 |
0.95 |
76.0 |
0.90 |
V pos |
24 (6.0) |
4.9 |
75.6 |
||
Unknown |
173 (43.4) |
||||
p16 |
<0.001* |
0.004* |
|||
Negative |
241 (60.4) |
3.0 |
73.8 |
||
Positive |
60 (15.0) |
39.4 |
81.3 |
||
Unknown |
98 (24.6) |
||||
Grade |
0.62 |
0.18 |
|||
1 |
7 (1.8) |
4.6 |
74.2 |
||
2 |
199 (49.9) |
4.9 |
74.4 |
||
3 |
107 (26.8) |
6.3 |
77.4 |
||
Unknown |
86 (21.6) |
||||
Surgical Margin |
0.53 |
0.22 |
|||
Negative |
272 (68.2) |
4.8 |
75.0 |
||
Positive |
44 (11.1) |
5.8 |
76.8 |
||
Unknown |
83 (20.8) |
||||
Second Tumor |
0.81 |
0.38 |
|||
Yes |
48 (12.0) |
4.3 |
74.5 |
||
No |
201 (50.4) |
4.6 |
77.0 |
||
Unknown |
150 (37.6) |
Mann-Whitney U test for sex, smoking status, pT, p16, second tumor, surgical margin, grade; One-Way ANOVA for alcohol consumption, tumor site; Spearman’s correlation for age, pack years, pN
*significant feature
Methylation of PITX2 and PANCR was tested as a dichotomized variable with clinico-pathological parameters (p16 expression, nodal status (N), distant metastases (M), vascular invasion (V), and age). A significant correlation was found between p16 expression and PITX2 and PANCR methylation (p< 0.001 and p = 0.004, respectively). Also, the tumor site correlated with PITX2 methylation (p<0.001). Interestingly, methylation of PANCR correlated with lymph node status (p<0.001). No further correlations were observed.
Hypermethylation of PITX2 and hypomethylation of PANCR is prognostic for overall survival in HNSCC
Cox proportional hazard model analysis was performed with methylation as a continuous variable (Table 2). The quantitative level of PITX2 was a significant predictor for overall survival (HR = 0.99, 95%CI: 0.98-1.00, p = 0.005). Low methylation levels were associated with a higher risk of death. High PANCR methylation, on the other hand, was associated with a significant increase of death (HR = 1.01, 95%CI: 1.00-1.02, p = 0.025). Optimized cut-offs for dichotomizing the data in hyper- and hypomethylated tumor samples were achieved by using a publicly available cut-off finder [46]. The optimization has been carried out with regard to low p-values of the log-rank test. The optimized cut-off was 7.0% for PITX2 and 69.3% for PANCR, respectively (Figure 2). Calculating the Cox proportional hazard model with dichotomized data for both parameters showed that methylation of both loci added significant information about risk of death. PITX2 hypermethylation resulted in a significantly reduced risk (HR = 0.51, 95%CI: 0.35-0.74, p<0.001), while PANCR hypermethylation was associated with a higher risk of death (HR = 1.64, 95%CI: 1.12-2.39, p=0.010).
Table 2: Univariate and multivariate Cox proportional hazards model analyses of overall survival including p16 expression, N-category, T-category, vascular invasion (V), tumor site, PITX2 hypermethylation, and PANCR hypermethylation. For dichotomization of PITX2 and PANCR DNA methylation, the optimized cut-off was used
Variable |
Univariate |
Multivariate |
||
|---|---|---|---|---|
HR (95% CI) |
p-value |
HR (95% CI) |
p-value |
|
PITX2 methylation (continuous variable) |
0.99 (0.98-1.00) |
0.005* |
0.98 (0.96-1.00) |
0.001* |
PANCR methylation (continuous variable) |
1.01 (1.00-1.02) |
0.025* |
1.03 (1.01-1.05) |
0.010* |
p16 expression (neg. reference) |
0.40 (0.20-0.81) |
0.010* |
0.78 (0.30-2.06) |
0.61 |
N-category |
1.28 (1.07-1.53) |
0.008* |
1.12 (0.81-1.56) |
0.49 |
T-category |
1.14 (0.96-1.36) |
0.14 |
1.63 (1.20-2.20) |
0.002* |
Vascular invasion (V) |
3.51 (1.90-6.47) |
<0.001* |
8.35 (3.70-18.9) |
<0.001* |
Tumor site |
0.86 (0.73-1.01) |
0.073 |
0.66 (0.48-0.90) |
0.009* |
*significant feature
Multivariate analysis performed with continous variables clearly revealed significant independent information for methylation in both loci, even outperforming p16 expression (Table 2). Methylation analysis continues to be significant when other powerful prognostic parameters like vascular invasion are included in the analysis.
Dichotomized data revealed a significantly prolonged overall survival for patients with PITX2 hypermethylated tumors, as demonstrated in a Kaplan-Meier survival analysis (p=0.002) (Figure 4B). PANCR low methylation was significantly associated with prolonged overall survival (p = 0.009) (Figure 4C).
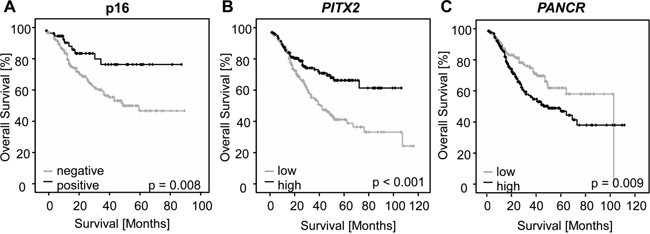
Figure 4: Kaplan-Meier analysis of overall survival in HNSCC patients. A. Patients (n=284) stratified by p16 expression, a surrogate biomarker for HPV infection. Concordant with published data, patients with HPV-associated (p16-positive) tumors in this study experienced significantly prolonged overall survival. B and C. Kaplan-Meier analysis of overall survival in HNSCC patients (n = 373) stratified according to the PITX2 (A) and PANCR (B) DNA methylation levels in tumors. Patients with high PITX2 methylation status survived significantly longer than those with low methylation status. Patients with low PANCR methylation status survived significantly longer than those with high methylation status.
PITX2 and PANCR methylation is associated with p16 expression
As a surrogate marker of HPV infection, p16 expression was evaluated in 301 primary tumors in a two-tiered manner as either absent or present. A total of 241 tumors (80.1%) were negative, while 60 cases (19.9%) were positive. In concordance with published literature, patients whose tumors presented with p16 expression had a significantly longer survival (p = 0.002, Figure 4A) [47]. Additionally, Cox proportional hazard analysis confirmed that a positive p16 status significantly reduced the risk of death (Hazard Ratio (HR) = 0.40, 95%CI: 0.20-0.81, p = 0.01). High PITX2 methylation was associated with p16 expression and patients with both p16 positive and PITX2 hypermethylated HNSCC had a significantly longer survival (p = 0.001). No difference in survival was found when stratifying the data for PANCR and p16 status.
DISCUSSION
HNSCC represents a large health burden worldwide. With regard to survival, only little improvement has been achieved over the last decades [48]. Despite the integration of targeted therapies into the therapeutic portfolio, surgery and radiotherapy are still the mainstay of therapy. A significant number of patients will experience local or distant relapse and some of these may have benefitted from a more aggressive therapy upfront. Tissue-based prognostic biomarkers can be of assistance in stratifying treatment and surveillance intensity. Currently, adjuvant treatment is established only for patients with classical adverse risk factors like pT3/T4 tumors or lymph node metastases [49]. A molecular biomarker could help identifying high-risk patients that would benefit from an adjuvant therapy even without harboring classical risk factors. On the other hand, for patients with high-risk tumors an advantageous molecular profile might prompt omission of adjuvant therapy. However, the molecular mechanisms are incompletely understood therefore the possibility that PITX2 and PANCR status might add to radiosensitivity cannot be ruled out.
A subgroup of patients with both classical and molecular adverse risk factors may benefit from intensifying treatment by adding immunotherapeutic agents upfront. The immune checkpoint inhibitor pembrolizumab has recently been approved by the Food and Drug Administration (FDA) for the treatment of patients with recurrent or metastatic disease after chemotherapy failure. Pembrolizumab is currently being investigated in combination with radiation and chemotherapy and could be part of an adjuvant or neoadjuvant regime in the future (ClinicalTrials.gov Identifiers: NCT02641093, NCT02777385). Promising results in terms of overall survival have also been demonstrated for the PD-1 antibody nivolumab [50].
The findings of the present study show that PITX2 and PANCR methylation status is associated with overall survival. HNSCC patients with tumors harboring a low PITX2 methylation status have a significantly reduced overall survival. Considering the important contribution of PITX2 to cell cycle regulation, low methylation may result in augmented transcriptional activity of PITX2 and subsequent tumor growth. Consequently, high methylation levels may lead to PITX2 silencing and thus, inhibition of its downstream target effects.
PITX2 methylation has been investigated in other tumors as well. In contrast to the findings in the present study hypermethylation in breast and prostate cancer is associated with poor outcome [28-30, 32, 33]. In prostate cancer, PITX2 is an upstream regulator of a key transcription factor, the androgen receptor (AR). Hypermethylation of the PITX2 promoter in postate cancer cell lines might result in dysregulation and activation of this pathway, in particular in advanced disease [51]. In NSCLC, particular the squamous cell subtype however, hypermethylation is associated with an improved overall survival [43]. Both HNSCC and NSCLC are smoking-related, hormone-independent tumors harboring a high frequency of alterations in the TP53 pathway [7, 52]. Further research is needed to elucidate the interaction between PITX2, PANCR, and the TP53 pathway.
Interestingly, a significant correlation between p16 expression and tumor site and high methylation was demonstrated, underscoring that tumorigenesis in HPV-positive tumors does not predominantly involve the Wnt/μ-Catenin pathway. In contrast a high PITX2 protein expression, suggestive of a low or unmethylated PITX2 gene, is found in colorectal cancer where the WNT/β-Catenin is predominantly activated due to APC mutations [53]. The same has been demonstrated in other tumors with an aberrant WNT/ß-catenin pathway like esophageal squamous cell cancer [54, 55].
P16 expression has been shown to be prognostic for overall survival in patients with oropharyngeal squamous cell carcinomas [47]. Therefore, it would be interesting to evaluate whether PITX2 or PANCR methylation add additional prognostic information beyond that of p16 status. However, since the number of oropharyngeal tumors in this cohort is too low this question could not be addressed.
Long noncoding RNA has been identified as an important regulator of gene expression both during and after transcription. Dysregulation of lncRNA is implicated in the development of many cancer types [56]. The exact regulatory mechanism, however, is not very well understood and even less is known about epigenetic regulation of lncRNA. The recently described PITX2 adjacent non-coding RNA (PANCR) has so far never been investigated in human cancer and this is the first study to analyze PANCR methylation at all. In our study, improved survival was associated with low PANCR methylation, however, the transcriptional consequence of PANCR methylation is unclear. Further investigation including transcriptomic data is needed to define whether PANCR plays a role in HNSCC development and progression. High PANCR methylation furthermore augmented the risk of death. It is well established that methylation in the transcriptional start region of a gene may lead to transcriptional repression. Methylation of CpG island in the gene body, on the other hand, can be positively associated with gene expression [57]. Therefore, it is intriguing to hypothesize that the low methylation of PANCR occurs at an intragenic CpG island and is associated with a high transcriptional activity of the whole locus, including PTIX2.
Interestingly, when compared with the surrounding normal tissue, hypermethylation of both gene loci was significantly more frequent in the tumor tissue. Tumorigenesis in HNSCC has long been established to be subject to a field cancerization, i.e. long-term exposure to noxa results in accumulation of genetic damage in a given anatomic location [58]. In contrast, other alterations like overexpression of the epidermal growth factor receptor (EGFR) are present in tumor-adjacent, morphologically normal mucosa [59].
In general, recurrent disease is diagnosed by radiological studies and/or biopsy. Patients may not qualify for further tumor resection but instead undergo radiotherapy and/or chemotherapy. Thus, tissue for ancillary analysis in addition to clinically ordered tests (immunohistochemistry, whole exome sequencing or targeted gene panels) is very limited. In this study, the often scant tumor tissue of recurrent disease was therefore not tested. However, a stepwise analysis of normal mucosa of high risk/no risk patients, dysplastic epithelium, carcinoma in situ, invasive squamous cell cancer, lymph node and distant metastases is needed to fully understand the role of PITX2 and PANCR methylation in development and progression of HNSCC.
In brief, this study that shows that PITX2 and PANCR methylation status are strong predictors of overall survival in HNSCC patients. Tissue-based measurement of both PITX2 and PANCR provides an objective result that can support clinicians in making treatment and surveillance decisions. A transfer of the assay to biopsy samples may allow for decision making prior to surgery or definite radiochemotherapy. However, implementing PITX2 and PANCR methylation status as a biomarker into everyday clinical practice is not yet feasible. A prospective validation in an independent patient cohort is needed to address the overall benefit of these two tissue-based biomarkers.
MATERIALS AND METHODS
Patients and ethics statement
The present study was approved by the Institutional Review Board at the University Hospital of Bonn which waved the need for written informed consent. In total, 399 patients with localized or locally advanced HNSCC treated with curative intent at the University Hospital Bonn were included. Tumors presenting with non-squamous histology were not included in the cohort. Overall survival was calculated as time between primary therapy and death. Clinical and pathological parameters like smoking habits and p16 immunohistochemistry, as surrogate marker for HPV infection, were obtained for part of the cohort [60].
DNA extraction and bisulfite conversion
Samples were obtained from 399 HNSCC patients. For each patient, 2 μm sections from formalin-fixed and paraffin-embedded (FFPE) tissues were stained with hematoxylin and eosin. A board certified pathologist annotated the tumor area and 1-5 cores (0.6 mm in diameter) were punched from each tumor and collected in a 1.5 ml tube (Eppendorf, Hamburg, Germany). Sample lysis, bisulfite conversion, and DNA purification was conducted using the innuCONVERT Bisulfite All-in-one Kit (Analytik Jena, Jena, Germany) [61].
Quantitative real-time polymerase chain reaction (PCR) assays
The PCR assays were performed using an AB 7500 Fast Real-Time PCR System (Life Technologies Corporation, Carlsbad, CA, USA).
PITX2 Duplex-PCR
The PITX2 DNA methylation was quantified by a methylation-specific real-time PCR as described previously [43, 44]. The thresholds and baselines for analysis were set as follows: 0.015 (threshold PITX2), 0.01 (threshold PITX2-Reference), 3-20 (baseline). The methylation was calculated as follows by the ΔΔCT method as previously published [43].
PANCR assay
The PANCR DNA methylation was analyzed by a quantitative methylation (QM) real-time PCR assay. The PANCR-assay was performed in a 20-μl reaction containing 0.4 μM of each primer, 0.3 μM of each detection probe, 0.25 mM each dNTP, 1 x PCR buffer [35 mM Tris (pH 8.4), 6 mM MgCl2, 50 mM KCl, 4% Glycerol] and 3 U FastStart Taq polymerase (Roche, Mannheim, Germany), 0.006 μl ROX solution (prepared as previously described [43]) and 50 ng bisulfite-converted sample DNA (according to UV quantification). The following temperature profile was used: 15 min at 95°C of initial denaturation, followed by 50 cycles with 60 sec at 56°C (100% ramp rate) and 15 sec at 95°C (75% ramp rate). The thresholds and baselines for analysis were set as follows: 0.01 (threshold, PANCR-P-M), 0.01 (threshold PANCR-P-U), 3-20 (baseline). The methylation was calculated as follows: ΔCT = ΔCTPANCR-P-U – ΔCTPANCR-P-M, ΔΔCT = ΔCTsample – ΔCTcalibrator, mPANCR = 100% / (1+2^(ΔΔCT)).
Statistical analysis
The statistical analyses were performed using SPSS, version 20 (SPSS Inc., Chicago, IL, USA). The relationship between input DNA methylation and measured DNA methylation was assessed by linear regression. Correlation with clinicopathological parameters was evaluated using the Spearman’s rho correlation coefficient. Comparisons were performed using the Mann-Whitney U and One-way ANOVA test. Overall survival analyses were conducted by Kaplan-Meier and the hazard ratio was evaluated using Cox proportional hazard model analysis. P-values refer to the log-rank test. P-values lower than 0.05 were considered significant.
CONFLICTS OF INTEREST
Dimo Dietrich has been an employee and is a stockholder of Epigenomics AG. Dimo Dietrich is inventor and co-inventor and owns patents on methylation biomarkers and related technologies which are commercially exploited by Epigenomics AG. Dimo Dietrich receives inventor’s compensation from Epigenomics AG. Dimo Dietrich is a consultant for AJ Innuscreen GmbH (Berlin, Germany), a 100% daughter company of Analytik Jena AG (Jena, Germany), and receives royalties from product sales. Dimo Dietrich is a consultant and receives or received compensation from Therawis GmbH (Munich, Germany), Oncgnostics GmbH (Jena, Germany), MDxHealth, Inc. (Irvine, CA, USA), Epigenomics AG (Berlin, Germany) and R-Biopharm AG (Darmstadt, Germany).
GRANT SUPPORT
The study has been funded by the University Hospital of Bonn.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2016. CA Cancer J Clin. 2016; 66:7-30.
2. IARC. (2005). Pathology and Genetics of Head and Neck Tumours.
3. Mashberg A, Boffetta P, Winkelman R, Garfinkel L. Tobacco smoking, alcohol drinking, and cancer of the oral cavity and oropharynx among U.S. veterans. Cancer. 1993; 72:1369-1375.
4. Vokes EE, Agrawal N, Seiwert TY. HPV-Associated Head and Neck Cancer. Journal of the National Cancer Institute. 2015; 107:djv344.
5. zur Hausen H. Human papillomaviruses and their possible role in squamous cell carcinomas. Current topics in microbiology and immunology. 1977; 78:1-30.
6. Gillison ML, Koch WM, Capone RB, Spafford M, Westra WH, Wu L, Zahurak ML, Daniel RW, Viglione M, Symer DE, Shah KV, Sidransky D. Evidence for a causal association between human papillomavirus and a subset of head and neck cancers. Journal of the National Cancer Institute. 2000; 92:709-720.
7. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature. 2015; 517:576-582.
8. Argiris A, Karamouzis MV, Raben D, Ferris RL. Head and neck cancer. Lancet. 2008; 371:1695-1709.
9. Whang SN, Filippova M, Duerksen-Hughes P. Recent Progress in Therapeutic Treatments and Screening Strategies for the Prevention and Treatment of HPV-Associated Head and Neck Cancer. Viruses. 2015; 7:5040-5065.
10. Olthof NC, Straetmans JM, Snoeck R, Ramaekers FC, Kremer B, Speel EJ. Next-generation treatment strategies for human papillomavirus-related head and neck squamous cell carcinoma: where do we go? Rev Med Virol. 2012; 22:88-105.
11. De Felice F, Thomas C, Barrington S, Pathmanathan A, Lei M, Urbano TG. Analysis of loco-regional failures in head and neck cancer after radical radiation therapy. Oral oncology. 2015; 51:1051-1055.
12. Sacco AG, Cohen EE. Current Treatment Options for Recurrent or Metastatic Head and Neck Squamous Cell Carcinoma. Journal of clinical oncology. 2015; 33:3305-3313.
13. Gentzler R, Hall R, Kunk PR, Gaughan E, Dillon P, Slingluff CL, Jr., Rahma OE. Beyond melanoma: inhibiting the PD-1/PD-L1 pathway in solid tumors. Immunotherapy. 2016; 8:583-600.
14. Tanguy Y, Seiwert RIH, Gupta S, Mehra R, Tahara M, Berger R, Lee S-H, Burtness B, Le DT, Heath K, Blum A, Dolled-Filhart M, Emancipator K, et al. Antitumor activity and safety of pembrolizumab in patients (pts) with advanced squamous cell carcinoma of the head and neck (SCCHN): Preliminary results from KEYNOTE-012 expansion cohort. J Clin Oncol 33, 2015. 2015.
15. Lohaus F, Linge A, Tinhofer I, Budach V, Gkika E, Stuschke M, Balermpas P, Rodel C, Avlar M, Grosu AL, Abdollahi A, Debus J, Bayer C, et al. HPV16 DNA status is a strong prognosticator of loco-regional control after postoperative radiochemotherapy of locally advanced oropharyngeal carcinoma: results from a multicentre explorative study of the German Cancer Consortium Radiation Oncology Group (DKTK-ROG). Radiotherapy and oncology. 2014; 113:317-323.
16. Balermpas P, Rodel F, Rodel C, Krause M, Linge A, Lohaus F, Baumann M, Tinhofer I, Budach V, Gkika E, Stuschke M, Avlar M, Grosu AL, et al. CD8+ tumour-infiltrating lymphocytes in relation to HPV status and clinical outcome in patients with head and neck cancer after postoperative chemoradiotherapy: A multicentre study of the German cancer consortium radiation oncology group (DKTK-ROG). International journal of cancer. 2016; 138:171-181.
17. Reddy RB, Bhat AR, James BL, Govindan SV, Mathew R, Dr R, Hedne N, Illiayaraja J, Kekatpure V, Khora SS, Hicks W, Tata P, Kuriakose MA, Suresh A. Meta-Analyses of Microarray Datasets Identifies ANO1 and FADD as Prognostic Markers of Head and Neck Cancer. PloS one. 2016; 11:e0147409.
18. Chau NG, Li YY, Jo VY, Rabinowits G, Lorch JH, Tishler RB, Margalit DN, Schoenfeld JD, Annino DJ, Goguen L, Thomas T, Becker H, Haddad T, et al. Incorporation of next-generation sequencing into routine clinical care to direct treatment of head and neck squamous cell carcinoma. Clinical cancer research. 2016.
19. Rauch TA, Zhong X, Wu X, Wang M, Kernstine KH, Wang Z, Riggs AD, Pfeifer GP. High-resolution mapping of DNA hypermethylation and hypomethylation in lung cancer. Proceedings of the National Academy of Sciences of the United States of America. 2008; 105:252-257.
20. Ting AH, McGarvey KM, Baylin SB. The cancer epigenome--components and functional correlates. Genes & development. 2006; 20:3215-3231.
21. Bienkowski M, Berghoff AS, Marosi C, Wohrer A, Heinzl H, Hainfellner JA, Preusser M. Clinical Neuropathology practice guide 5-2015: MGMT methylation pyrosequencing in glioblastoma: unresolved issues and open questions. Clinical neuropathology. 2015; 34:250-257.
22. Brigliadori G, Foca F, Dall'Agata M, Rengucci C, Melegari E, Cerasoli S, Amadori D, Calistri D, Faedi M. Defining the cutoff value of MGMT gene promoter methylation and its predictive capacity in glioblastoma. Journal of neuro-oncology. 2016.
23. Quentien MH, Barlier A, Franc JL, Pellegrini I, Brue T, Enjalbert A. Pituitary transcription factors: from congenital deficiencies to gene therapy. Journal of neuroendocrinology. 2006; 18:633-642.
24. Lin CR, Kioussi C, O'Connell S, Briata P, Szeto D, Liu F, Izpisua-Belmonte JC, Rosenfeld MG. Pitx2 regulates lung asymmetry, cardiac positioning and pituitary and tooth morphogenesis. Nature. 1999; 401:279-282.
25. Kioussi C, Briata P, Baek SH, Rose DW, Hamblet NS, Herman T, Ohgi KA, Lin C, Gleiberman A, Wang J, Brault V, Ruiz-Lozano P, Nguyen HD, et al. Identification of a Wnt/Dvl/beta-Catenin —> Pitx2 pathway mediating cell-type-specific proliferation during development. Cell. 2002; 111:673-685.
26. Liu Y, Huang Y, Zhu GZ. Cyclin A1 is a transcriptional target of PITX2 and overexpressed in papillary thyroid carcinoma. Molecular and cellular biochemistry. 2013; 384:221-227.
27. Toyota M, Kopecky KJ, Toyota MO, Jair KW, Willman CL, Issa JP. Methylation profiling in acute myeloid leukemia. Blood. 2001; 97:2823-2829.
28. Nimmrich I, Sieuwerts AM, Meijer-van Gelder ME, Schwope I, Bolt-de Vries J, Harbeck N, Koenig T, Hartmann O, Kluth A, Dietrich D, Magdolen V, Portengen H, Look MP, et al. DNA hypermethylation of PITX2 is a marker of poor prognosis in untreated lymph node-negative hormone receptor-positive breast cancer patients. Breast cancer research and treatment. 2008; 111:429-437.
29. Maier S, Nimmrich I, Koenig T, Eppenberger-Castori S, Bohlmann I, Paradiso A, Spyratos F, Thomssen C, Mueller V, Nahrig J, Schittulli F, Kates R, Lesche R, et al. DNA-methylation of the homeodomain transcription factor PITX2 reliably predicts risk of distant disease recurrence in tamoxifen-treated, node-negative breast cancer patients—Technical and clinical validation in a multi-centre setting in collaboration with the European Organisation for Research and Treatment of Cancer (EORTC) PathoBiology group. Eur J Cancer. 2007; 43:1679-1686.
30. Harbeck N, Nimmrich I, Hartmann A, Ross JS, Cufer T, Grutzmann R, Kristiansen G, Paradiso A, Hartmann O, Margossian A, Martens J, Schwope I, Lukas A, et al. Multicenter study using paraffin-embedded tumor tissue testing PITX2 DNA methylation as a marker for outcome prediction in tamoxifen-treated, node-negative breast cancer patients. Journal of clinical oncology. 2008; 26:5036-5042.
31. Hartmann O, Spyratos F, Harbeck N, Dietrich D, Fassbender A, Schmitt M, Eppenberger-Castori S, Vuaroqueaux V, Lerebours F, Welzel K, Maier S, Plum A, Niemann S, Foekens JA, Lesche R, Martens JW. DNA methylation markers predict outcome in node-positive, estrogen receptor-positive breast cancer with adjuvant anthracycline-based chemotherapy. Clinical cancer research. 2009; 15:315-323.
32. Weiss G, Cottrell S, Distler J, Schatz P, Kristiansen G, Ittmann M, Haefliger C, Lesche R, Hartmann A, Corman J, Wheeler T. DNA methylation of the PITX2 gene promoter region is a strong independent prognostic marker of biochemical recurrence in patients with prostate cancer after radical prostatectomy. The Journal of urology. 2009; 181:1678-1685.
33. Banez LL, Sun L, van Leenders GJ, Wheeler TM, Bangma CH, Freedland SJ, Ittmann MM, Lark AL, Madden JF, Hartman A, Weiss G, Castanos-Velez E. Multicenter clinical validation of PITX2 methylation as a prostate specific antigen recurrence predictor in patients with post-radical prostatectomy prostate cancer. The Journal of urology. 2010; 184:149-156.
34. Fung FK, Chan DW, Liu VW, Leung TH, Cheung AN, Ngan HY. Increased expression of PITX2 transcription factor contributes to ovarian cancer progression. PloS one. 2012; 7:e37076.
35. Weller M, Tabatabai G, Kastner B, Felsberg J, Steinbach JP, Wick A, Schnell O, Hau P, Herrlinger U, Sabel MC, Wirsching HG, Ketter R, Bahr O, et al. MGMT Promoter Methylation Is a Strong Prognostic Biomarker for Benefit from Dose-Intensified Temozolomide Rechallenge in Progressive Glioblastoma: The DIRECTOR Trial. Clinical cancer research. 2015; 21:2057-2064.
36. Costa FF. Non-coding RNAs: Meet thy masters. BioEssays: news and reviews in molecular, cellular and developmental biology. 2010; 32:599-608.
37. Maia BM, Rocha RM, Calin GA. Clinical significance of the interaction between non-coding RNAs and the epigenetics machinery: challenges and opportunities in oncology. Epigenetics. 2014; 9:75-80.
38. Ragusa M, Barbagallo C, Statello L, Condorelli AG, Battaglia R, Tamburello L, Barbagallo D, Di Pietro C, Purrello M. Non-coding landscapes of colorectal cancer. World journal of gastroenterology. 2015; 21:11709-11739.
39. Amaral PP, Dinger ME, Mercer TR, Mattick JS. The eukaryotic genome as an RNA machine. Science. 2008; 319:1787-1789.
40. Taft RJ, Pang KC, Mercer TR, Dinger M, Mattick JS. Non-coding RNAs: regulators of disease. The Journal of pathology. 2010; 220:126-139.
41. St Laurent G, Wahlestedt C, Kapranov P. The Landscape of long noncoding RNA classification. Trends in genetics: TIG. 2015; 31:239-251.
42. Gore-Panter SR, Hsu J, Barnard J, Moravec CS, Van Wagoner DR, Chung MK, Smith JD. PANCR, the PITX2 Adjacent Noncoding RNA, Is Expressed in Human Left Atria and Regulates PITX2c Expression. Circulation Arrhythmia and electrophysiology. 2016; 9.
43. Dietrich D, Hasinger O, Liebenberg V, Field JK, Kristiansen G, Soltermann A. DNA methylation of the homeobox genes PITX2 and SHOX2 predicts outcome in non-small-cell lung cancer patients. Diagn Mol Pathol. 2012; 21:93-104.
44. Dietrich D, Hasinger O, Banez LL, Sun L, van Leenders GJ, Wheeler TM, Bangma CH, Wernert N, Perner S, Freedland SJ, Corman JM, Ittmann MM, Lark AL, et al. Development and clinical validation of a real-time PCR assay for PITX2 DNA methylation to predict prostate-specific antigen recurrence in prostate cancer patients following radical prostatectomy. The Journal of molecular diagnostics: JMD. 2013; 15:270-279.
45. Curado MP, Hashibe M. Recent changes in the epidemiology of head and neck cancer. Current opinion in oncology. 2009; 21:194-200.
46. Budczies J, Klauschen F, Sinn BV, Gyorffy B, Schmitt WD, Darb-Esfahani S, Denkert C. Cutoff Finder: a comprehensive and straightforward Web application enabling rapid biomarker cutoff optimization. PloS one. 2012; 7:e51862.
47. Salazar CR, Anayannis N, Smith RV, Wang Y, Haigentz M, Jr., Garg M, Schiff BA, Kawachi N, Elman J, Belbin TJ, Prystowsky MB, Burk RD, Schlecht NF. Combined P16 and human papillomavirus testing predicts head and neck cancer survival. Int J Cancer. 2014; 135:2404-2412.
48. Sun W, Califano JA. Sequencing the head and neck cancer genome: implications for therapy. Annals of the New York Academy of Sciences. 2014; 1333:33-42.
49. Expert Panel on Radiation O-H, Neck, Salama JK, Saba N, Quon H, Garg MK, Lawson J, McDonald MW, Ridge JA, Smith RV, Yeung AR, Yom SS, Beitler JJ. ACR appropriateness criteria(R) adjuvant therapy for resected squamous cell carcinoma of the head and neck. Oral oncology. 2011; 47:554-559.
50. Busch CJ, Laban S, Knecht R, Hoffmann TK. [Immunotherapeutic studies of head and neck tumors: Highlights of the 2016 ASCO Annual Meeting]. HNO. 2016.
51. Schayek H, Bentov I, Jacob-Hirsch J, Yeung C, Khanna C, Helman LJ, Plymate SR, Werner H. Global methylation analysis identifies PITX2 as an upstream regulator of the androgen receptor and IGF-I receptor genes in prostate cancer. Hormone and metabolic research. 2012; 44:511-519.
52. Comprehensive genomic characterization of squamous cell lung cancers. Nature. 2012; 489:519-525.
53. Hirose H, Ishii H, Mimori K, Tanaka F, Takemasa I, Mizushima T, Ikeda M, Yamamoto H, Sekimoto M, Doki Y, Mori M. The significance of PITX2 overexpression in human colorectal cancer. Ann Surg Oncol. 2011; 18:3005-3012.
54. Zhang JX, Tong ZT, Yang L, Wang F, Chai HP, Zhang F, Xie MR, Zhang AL, Wu LM, Hong H, Yin L, Wang H, Wang HY, Zhao Y. PITX2: a promising predictive biomarker of patients' prognosis and chemoradioresistance in esophageal squamous cell carcinoma. International journal of cancer. 2013; 132:2567-2577.
55. Zhang JX, Chen ZH, Xu Y, Chen JW, Weng HW, Yun M, Zheng ZS, Chen C, Wu BL, Li EM, Fu JH, Ye S, Xie D. Downregulation of MicroRNA-644a Promotes Esophageal Squamous Cell Carcinoma Aggressiveness and Stem-cell-like Phenotype via Dysregulation of PITX2. Clinical cancer research. 2016.
56. Yan K, Arfat Y, Li D, Zhao F, Chen Z, Yin C, Sun Y, Hu L, Yang T, Qian A. Structure Prediction: New Insights into Decrypting Long Noncoding RNAs. International journal of molecular sciences. 2016; 17.
57. Kulis M, Queiros AC, Beekman R, Martin-Subero JI. Intragenic DNA methylation in transcriptional regulation, normal differentiation and cancer. Biochimica et biophysica acta. 2013; 1829:1161-1174.
58. Angadi PV, Savitha JK, Rao SS, Sivaranjini Y. Oral field cancerization: current evidence and future perspectives. Oral and maxillofacial surgery. 2012; 16:171-180.
59. van Oijen MG, Rijksen G, ten Broek FW, Slootweg PJ. Increased expression of epidermal growth factor receptor in normal epithelium adjacent to head and neck carcinomas independent of tobacco and alcohol abuse. Oral diseases. 1998; 4:4-8.
60. Merlano MC, Denaro N, Vivenza D, Monteverde M, Russi E, Lerda W, Comino A, Lo Nigro C. p16 cutoff in head and neck squamous cell carcinoma: correlation between tumor and patient characteristics and outcome. The International journal of biological markers. 2015:0.
61. Holmes EE, Jung M, Meller S, Leisse A, Sailer V, Zech J, Mengdehl M, Garbe LA, Uhl B, Kristiansen G, Dietrich D. Performance evaluation of kits for bisulfite-conversion of DNA from tissues, cell lines, FFPE tissues, aspirates, lavages, effusions, plasma, serum, and urine. PloS one. 2014; 9:e93933.