
INTRODUCTION
Hepatocellular carcinoma (HCC), a leading cause of cancer-related death worldwide, shows high metastasis and recurrence rates and chemoresistance [1,2]. Risk factors for HCC include aflatoxin, cirrhosis and hepatitis, chronic hepatitis B virus (HBV) and hepatitis C virus (HCV) infections, alcoholic liver diseases, and nonalcoholic fatty liver diseases [3]. Among these risk factors, chronic inflammation induced by HBV or HCV accounts for the majority of liver cancer development [4,5]. The causal roles of HBV and HCV infections in HCC tumorigenesis have been demonstrated as the efficient reduction of HCC development by eradicating these viruses [6]. However, distinct pathophysiologic mechanisms underlying HBV- and HCV-related hepatocarcinogenesis have been proposed [7].
Hepatitis B virus X protein (HBx), a critical antigen involved in HBV-associated liver diseases, determines an unique profile of gene expression in the host liver cells and contributes to HCC formation. As a multifunctional regulator, HBx modulates several cellular processes such as oxidative stress, DNA repair, signal transduction pathways, transcriptional regulations, protein degradation, cell cycle progression, apoptosis, and genetic stability by direct or indirect interaction with host factors [8,9]. Several reports have demonstrated that HBx promotes HCC cell proliferation by downregulation of p16 protein expression and upregulation of cyclin D1 protein expression via activation of the MEK/ERK and PI3K/Akt signaling pathways [10–13]. HBx also promotes tumor metastasis by inducing cyclooxygenase-2 and matrix metalloproteinase-1, 2, 3, and 9 expressions, and repressing E-cadherin and fibronectin type III domain containing 3B (FNDC3B) expressions [1,14–18]. Moreover, downregulation of p53 and p21, inhibition of caspase-3 activation, and upregulation of anti-apoptotic protein survivin and multi-drug resistance proteins were reported to mediated the chemoresistance of HBx-expressing hepatocellular carcinoma cells [19–22]. Disruption of HBx protein expression can effectively inhibit tumor growth and enhance chemotherapy-induced apoptosis in hepatocellular carcinoma cells [23–25]. Notably, microRNAs have been recognized as important regulators in HBV-related HCC progression via targeting gene expressions [26,27].
Recently, our study demonstrated that maspin, a mammary serine protease inhibitor, was specifically reduced by HBx protein in HBV-associated HCC patients, [28]. The downregulation of maspin promoted cell motility and rendered resistance to anoikis and chemotherapy in HCC cells. Furthermore, the increased microRNA-7, -103, -107, and -21 in response to HBx overexpression was demonstrated to directly target maspin mRNA, and the levels of microRNA-7/21/107 were correlated to poor prognosis in HBV-associated HCC patients. However, the molecular mechanisms underlying these HBx-induced microRNA expressions remain to be elucidated.
Nuclear factor κB (NF-κB) pathway controls many important pathological processes including inflammation, immunity, cell proliferation, differentiation, survival, as well as cancer progression [29]. Aberrant NF-κB activation in response to various stimuli can promote cancer invasion, metastasis, and chemoresistance in liver cancer [30,31]. Activation of NF-κB signaling pathway by HBx has been well documented [8,32]. In addition, our previous study also demonstrated that HBx induced the nuclear localization of IKKα, a upstream kinase for NF-κB activation, in an Akt phosphorylation-dependent manner to promote the migration and invasion of HCC cells via phosphorylating histone H3 at Ser10 [33]. Phosphorylation of Histone H3 Ser10 on maspin promoter by RANKL-activated nuclear IKKα was proposed to directly repress maspin transcription through subsequent DNA methylation [34]. However, this histone posttranslational modification was widely reported to enhance transcription of most genes involved in chromosome decondensation and cell-cycle progression during mitosis and meiosis as well as the NF-κB-targeted gene expressions during inflammation [35–37]. The closed proximity to other modifiable residues on the histone H3 tail leads to the cross-talk of serine 10 phosphorylation with the transcription-activating acetylation at lysine 9 and lysine 14 [38]. Thus, maspin suppression by nuclear IKKα may involve an indirect regulation through inducing gene expression of intermediate suppressors such as microRNAs rather than DNA methylation merely.
In the present study, we found an inverse correlation between phosphorylated nuclear IKKα and maspin protein expression in HBV-associated HCC patients. The activity and nuclear translocation of IKKα but not IKKβ was crucial for HBx-mediated maspin downregulation and chemoresistance in HCC cells. Furthermore, nuclear IKKα-induced microRNA-7, -21, -103, and -107 expressions relying on histone H3 Ser10 phosphorylation to disrupt maspin mRNA stability and translation. These results provide new insights into the molecular mechanisms of maspin suppression in response to HBx, and revealed nuclear IKKα as a prognostic biomarker and a potential therapeutic target to improve the clinical outcome of HBV-associated HCC patients.
RESULTS
Nuclear IKKα significantly correlates with low levels of maspin expression in HBV-associated HCC patients
Our previous study has demonstrated that HBx-mediated maspin suppression contributed to HBV-induced HCC progression [28]. We also demonstrated that HBx induced nuclear IKKα translocation through Akt-dependent Thr-23 phosphorylation to promote motility of hepatocarcinoma cells [33]. Furthermore, cytokine-activated nuclear IKKα has been reported to repress maspin to promote metastasis of prostate cancer [34]. Therefore, the correlation between nuclear IKKα and maspin suppression in HBV-associated HCC tumors was first examined. The phosphorylation of IKKα at Thr-23, which was recognized as a marker for nuclear localization, was elevated and predominantly localized in the nucleus, and was inversely correlated with maspin expression in HBV-associated HCC tumors (Figure 1A and 1B, respectively), supporting the involvement of nuclear IKKα in maspin suppression. Additionally, the clinical association of IKKα T23 phosphorylation and maspin expression with the status of HBV-associated HCC tumors was also analyzed. In the comparison to the normal tissues, IKKα T23 phosphorylation is up-regulated and maspin expression is downregulated in the stage III but not in stage I and II HCC tumor tissues (Figure 1C).
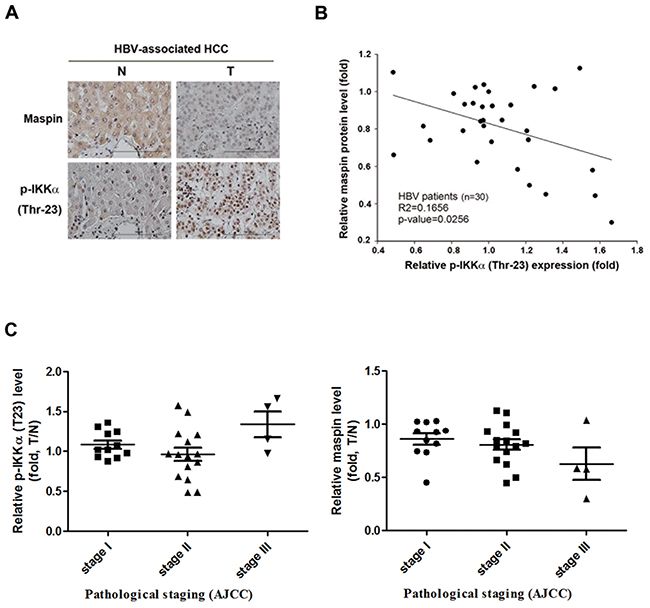
Figure 1: Inverse correlation between phospho-IKKα and maspin expression in HBV-associated HCC patients. A. Representative immunohistochemical staining of maspin (top) and phospho-IKKα (Thr-23) (bottom) in HBV-associated HCC tumor liver tissues (T) and adjacent normal liver tissues (N) was shown. Scale bar: 100 μm. B. and C. Total lysates from HBV-associated HCC tumor liver tissues were prepared and subjected to Western blot with anti- phospho-IKKα (Thr-23), maspin, and ERK antibodies. The coefficient of determination (r2) between IKK phosphorylation and maspin expression levels was analyzed by simple regression with normalization to ERK protein level (n=30). The clinical association of p-IKKα and maspin levels with the stages of HBV-associated HCC was further analyzed by a Student’s t-test.
Nuclear IKKα but not IKKβ mediated HBx-dependent maspin suppression and chemoresistance in HCC cells
Since the IKK-NF-κB signaling pathway plays an important role in the development of HCC, the regulatory function of IKKα and IKKβ, the essential kinases controlling canonical and noncanonical NF-κB signaling, in maspin expression were further examined. Overexpression of IKKα but not IKKβ downregulated maspin protein expression as well as the mRNA level in Hep3B cells (Figure 2A and 2B). Furthermore, the maspin suppression was abolished by mutation of the IKKα nuclear localization signal (NLS) (Figure 2C). Our previous study has demonstrated that HBx suppressed maspin expression and enhanced chemoresistance [28]. The role of IKKα in HBx-mediated maspin suppression was further examined by silencing of IKKα with shRNA. Indeed, knockdown of IKKα prevented HBx-induced maspin suppression in transient (Figure 2D) and stable (Figure 2E) HBx transfectants of Hep3B cells. To further verify the critical role of IKKα in HBx-mediated chemoresistance, IKK inhibitor VII was utilized and the cytotoxicity of Hep3Bx cells was determined by MTT assay. We found that IKK inhibitor VII significantly increased the sensitivity of Hep3Bx cells to doxorubicin (Figure 2F). Additionally, the effects of IKK inhibitor VII on IKKα activity and nuclear translocation as well as maspin expression were also addressed. As shown in Figure 2G, the expression of, phospho-IKKα/β (Ser177/176), phospho-IKKα (Thr-23), and phospho-p65 (Ser536) were downregulated whereas the expression of IκBα was upregulated by IKK inhibitor VII. Accompanied with the IKKα activity inhibition, the nuclear translocation of IKKα was decreased (Figure 2H) and the expression of maspin was significantly restored in Hep3Bx cells (Figure 2G). These data support the essential role of nuclear IKKα in HBx-induced maspin downregulation and chemoresistance.
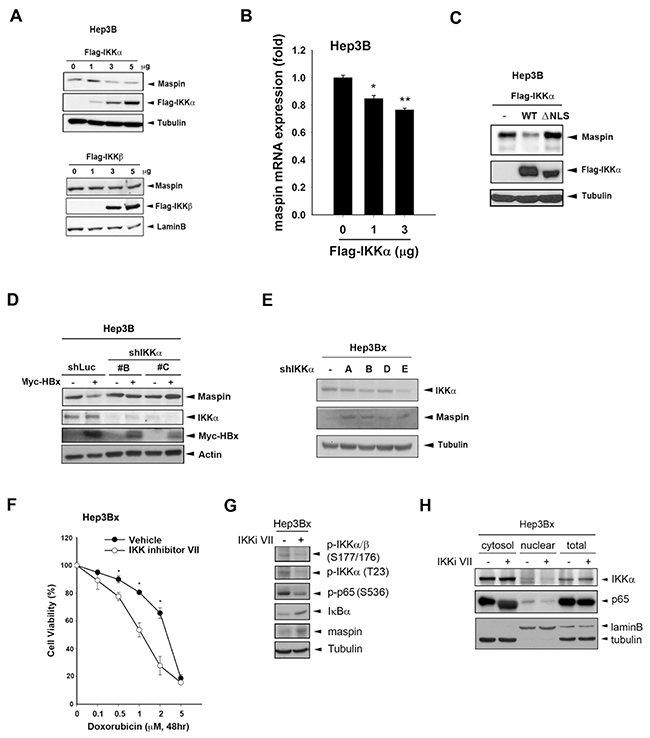
Figure 2: Nuclear IKKα mediated HBx-dependent maspin suppression and chemoresistance. A-D. Total lysates from Hep3B cells transfected with IKKα and its NLS deletion mutant IKKβ,myc-HBx along with IKKα shRNA were subjected to Western blot analysis with indicated antibodies. E. Hep3Bx cells were transfected with IKKα shRNA for 3 days, and then total lysates were prepared and subjected to Western blot analysis. F. Hep3Bx cells treated with doxorubicin in the presence or absence of IKK inhibitor VII for 24 hours were subjected to MTT assays to determine the cell viability (n=3). G. and H. After treatment with IKKα inhibitor VII for 48 hours, total, nuclear, and cytosolic lysates from Hep3Bx cells were prepared and subjected to Western blot analysis. The difference was calculated by a Student’s t-test (*: p<0.05; **: p<0.01)
Nuclear IKKα downregulats maspin expression through disrupting its mRNA stability
Although cytokine-activated nuclear IKKα has been suggested to suppress maspin transcription in prostate cancer cells [34], Overexpression of IKKα wild-type but not its T23A mutant significantly decreased maspin mRNA stability in the presence of actinomycin D (Figure 3A), indicating that a post-transcriptional regulation might be involved in nuclear IKKα-mediated maspin suppression. To further address whether nuclear IKKα mediates maspin suppression via affecting maspin-3′UTR activity, IKKα and maspin-3′UTR luciferase were co-transfected into HEK-293 cells. Overexpression of IKKα suppressed the luciferase activity dose-dependently (Figure 3B, left). However, there was no suppressive effect of IKKα on maspin promoter activity, suggesting that the post-transcriptional regulation is more critical for nuclear IKKα-mediated maspin suppression (Figure 3B, right). Furthermore, IKKα T23A mutation, but not the nuclear-prone IKKα T23E and IKKα NES mutations, abolished IKKα-mediated maspin 3′UTR suppression (Figure 3C). Additionally, silence of IKKα reversed HBx-mediated suppression of maspin 3′UTR activity, which was found to be mediated by microRNAs induction in our previous study (Figure 3D), suggesting the suppression of maspin mRNA stability by nuclear IKKα through induction of microRNAs.
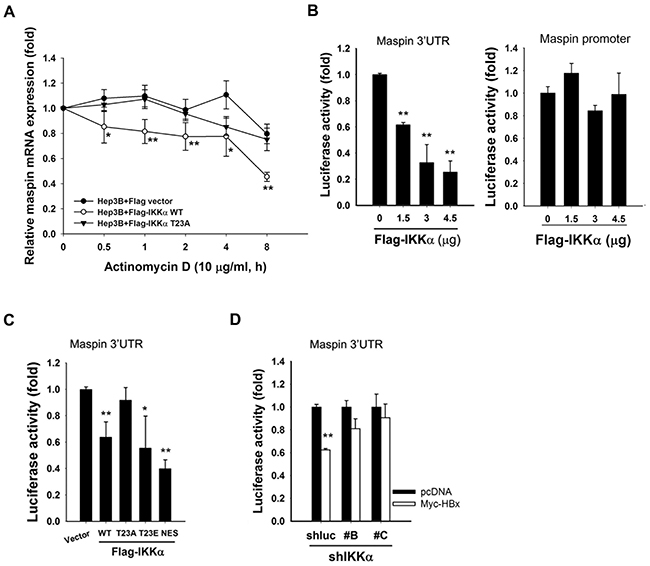
Figure 3: Nuclear IKKα disrupted maspin mRNA stability to mediate HBx-dependent maspin suppression. A. Hep3B cells were transfected with Flag-IKKα wild-type and T23A mutant for 3 days and then harvested total RNA at the indicated time points after actinomycin D (10 μ0g/ml) treatment to examine the stability of maspin mRNA by RT-qPCR (n=5). B-D. HEK-293 cells were transfected with maspin 3′UTR or promoter-luciferase gene, Flag-IKKα WT and its mutants, or IKKα shRNA with myc-HBx expression for 24 hours and then were subjected to luciferase activity assays (n=3). The difference was calculated by a Student’s t-test (*: p<0.05; **: p<0.01).
Nuclear IKKα induces miR-7, -103, -107, and -21 to mediate HBx-dependent maspin suppression
Our previous study has demonstrated that microRNA-7/21/103/107 mediated HBx-induced maspin suppression. To further verify the involvement of these microRNAs in nuclear IKKα-mediated maspin downregulation, the expression levels of these microRNAs in response to IKKα overexpression and IKKα inhibition were analyzed. Indeed, overexpression of wild-type IKKα (Figures 4A) but not its T23A nor NLS deletion mutants (Figure 4B) increased the expressions of miR-7, -103, -107, and -21 in Hep3B cells. Moreover, inhibition of IKKα by IKK inhibitor VII and by different IKKα shRNAs significantly decreased these miRNA expressions in Hep3Bx cells (Figures 4C and 4D, respectively). Furthermore, the inhibitory effect of IKKα on maspin 3′UTR-luciferase activity was attenuated by individual or combinatory mutations on maspin 3′UTR targeted sites for miR-7, miR-107, or miR-21 (Figure 4E). The inhibition of maspin 3′UTR activity by IKKα was also partially reversed by the inhibitors against miR-7, -103, or -107 alone and was greatly reversed by the combination of miR-7 and -107 inhibitors (Figure 4F). These data suggested that nuclear IKKα can upregulate miR-7, -103, -107, and -21 expressions to suppress maspin expression in Hep3Bx cells.
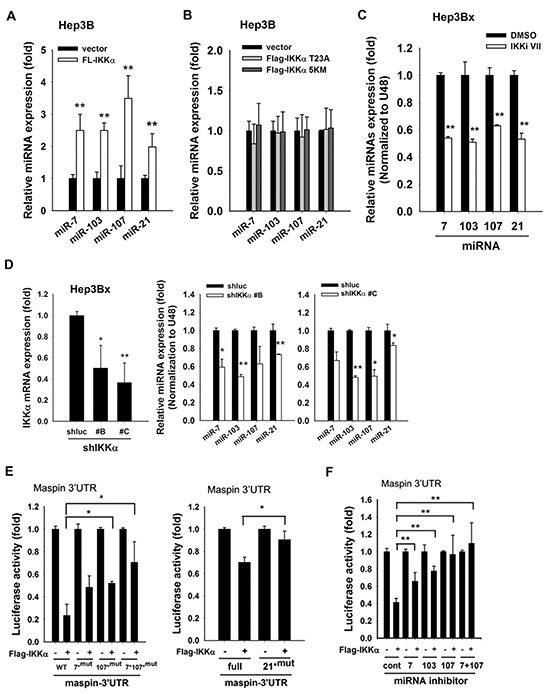
Figure 4: IKKα is involved in HBx-induced maspin suppression and maspin-targeting miRNA expressions. A. and B. Total RNA extracted from Hep3B cells transfected with or without Flag-IKKα or mutants (T23A and 5KM) were subjected to RT-qPCR to examine the expression levels of indicated microRNAs (n=3). C. and D. The total RNA extracted from IKKαi VII-treated or IKKα-knockdown Hep3Bx cells were analyzed by RT-qPCR. The expression levels of IKKα and maspin-targeting miRNAs were normalized to GAPDH and U48, respectively (n=3). E. HEK-293 cells were transfected with maspin 3′UTR-luciferase gene or its mutants, and Flag-IKKα for 24 hours and then were subjected to luciferase activity assays (n = 3). F. HEK-293 cells were co-transfected with maspin 3′UTR-luciferase gene, Flag-tagged IKKα, and miRNA inhibitors for 48 hours and then were subjected to luciferase activity assays (n = 4). The difference was calculated by a Student’s t-test (*: p<0.05; **: p<0.01).
Nuclear IKKα coordinates the transcriptional activity of NF-κB to mediate microRNA-7/21/103/107 expressions in HBx-expressing HCC cells
Our previous study has demonstrated that nuclear IKKα enhanced NFκB-mediated gene transcription by tipping CBP binding preference [39]. Overexpression of IKKα increased the active phosphorylation of p65 in Hep3B cells, and NFκB inhibitor PDTC downregulated miR-7, -103, -107, and -21, thereby restoring maspin expression in Hep3Bx cells (Figure 5A and 5B, respectively). These results suggest that HBx transcriptionally induced these microRNAs in a nuclear IKKα/NFκB-dependent manner to suppress maspin expression. The putative binding element for NF-κB is commonly observed on the promoter regions of these microRNAs’ host genes. The upstream regions and putative transcription starting site (TSS) of these microRNA precursors genes were predicted by using three independent database, TRANSFAC [40], JASPAR [41], and TFBIND [42] and were illustrated in Figure 6A, suggesting that HBx could elevate these microRNAs via epigenetically upregulating their host genes in a nuclear IKKα-dependent manner. Indeed, the gene expressions of their respective host genes including pituitary gland specific factor 1 (PGSF1) for miR-7, pantothenate kinase 2 (PANK2) for miR-103, and pantothenate kinase 1 (PANK1) for miR-107 were also elevated in Hep3Bx cells but suppressed by the silence of IKKα (Figure 6B and 6C). Moreover, accumulating evidence has indicated that the phosphorylation of histone H3 at serine 10 is necessary and sufficient for transcriptional activation, and IKKα/NFκB signaling is one of the cell-signaling cascades leading to this event [38,43]. Therefore, we performed chromatin immunoprecipitation (ChIP) assays to compare the level of phospho-H3 Ser10 on the promoters of these microRNAs in Hep3B and Hep3Bx cells. Consistently, the level of phospho-H3 Ser10 on the promoters of these microRNAs were much higher in Hep3Bx cells than Hep3B cells, indicating the chromatin fiber were more accessible to facilitate the binding of transcription factors in Hep3Bx cells (Figure 6D). To further demonstrate NFκB is the major transcription factor participated in the transcription of these miRNAs, the promoter binding ability of NFκB was further examined in Hep3B and Hep3Bx cells. As shown in Figure 6E, the binding ability of p65 on these miRNA promoters was significantly enhanced in Hep3Bx cells than Hep3B cells, strengthening the crucial role of NFκB in HBx-induced transcription of these miRNAs. These data indicated that nuclear IKKα might be responsible for phosphorylating histone H3 at Ser10 to facilitate the expressions of these NF-κB-dependent microRNA, resulting in maspin downregulation.
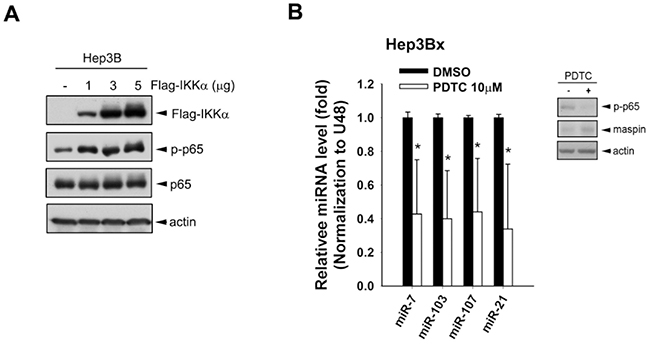
Figure 5: NF-κB activated by IKKα overexpression was involved in HBx-induced miRNA expressions. A. Total lysates prepared from Hep3B cells transfected with increasing amount of IKKα were subjected to Western blot analysis with indicated antibodies. B. The total RNA and protein lysates extracted from Hep3Bx cells treated with NFκB inhibitor PDTC, were analyzed by RT-qPCR and Western blot analysis. The expression levels of maspin-targeting miRNAs were normalized to U48 (n=4).
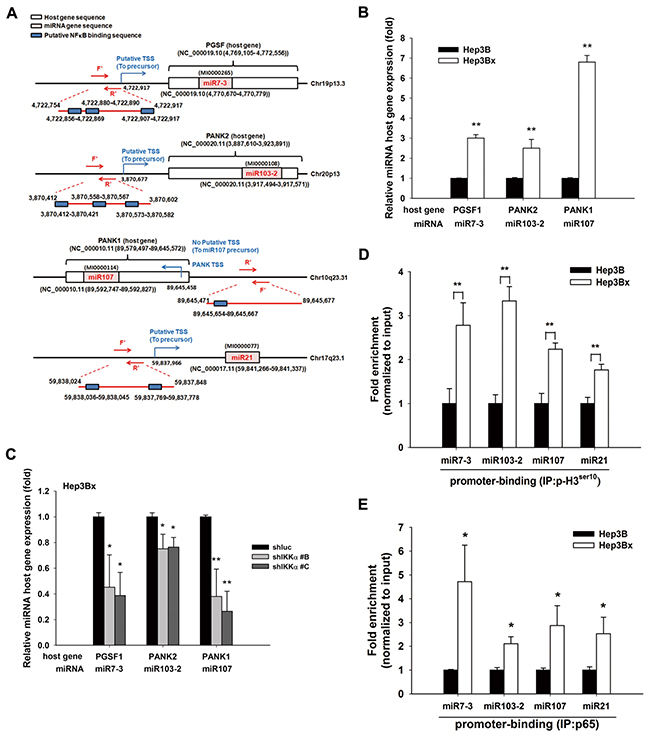
Figure 6: IKKα-activated NF-κB pathway transcriptional regulated HBx-mediated miRNAs and their host gene expression. A. The diagram illustrates the gene locations of miRNA-7, -103, -107, and -21 and their predicted promoter regions. B. and C. The total RNA extracted from Hep3B, Hep3Bx, and IKKα-knockdown Hep3Bx cells was analyzed by RT-qPCR. The expression levels of miRNA host genes were normalized to GAPDH (n=4). D. and E. Total lysate from Hep3B and Hep3Bx cells were subjected to ChIP assays with anti-H3 phospho-Ser10 and anti-p65 antibodies, followed by RT-qPCR with specific primers for indicated gene promoters. The difference was calculated by a Student’s t-test. *p<0.05; **p<0.01.
DISCUSSION
It is well-accepted that constitutive NF-κB signaling activation promotes cancer development by increasing cell proliferation, angiogenesis, metastasis, and resistance to apoptotic stimuli [44]. It has been known that activation of NF-κB signal by both chronic hepatitis virus HBV and HCV infections contributes to the development of HCC [45]. The IKKα kinase complex is the master regulator for NF-κB activation [46,47]. High expression levels of IKKα and IKKβ have found to be necessary for the malignant properties of liver cancer [30]. However, the role of IKKs on virus infection-mediate gene regulation and tumor progression need to be further investigated. Previously, we demonstrated that HBx activates Akt to phosphorylate IKKα at T23 residue, thus promoting IKKα nuclear localization. Furthermore, we also revealed the unique role of HBx protein in maspin suppression through microRNA induction to promote HBV-associated HCC tumor progression [28]. In the present study, the mechanism underlying HBx-dependent microRNA inductions was further uncovered. Nuclear activated IKKα transcriptionally up-regulates microRNA-7, -21, -103, and -107 expressions to target maspin 3′UTR, contributing to HBx-mediated maspin suppression and subsequent chemoresistance.
Maspin has been characterized as a class II tumor suppressor based on its ability to inhibit cell invasion and angiogenesis and to promote apoptosis [48–50], suggesting maspin as a potential therapeutic target in multiple cancer types [51]. Several lines of evidence indicate the involvement of IKKα in regulating maspin expression. Downregulation of IKKα increased maspin expression to inhibit metastasis and promote apoptosis in HCC cells [30]. In addition, cytokine-activated nuclear IKKα repressed maspin transcription to control prostate cancer metastasis [34]. However, the detailed regulatory mechanism of IKKα-mediated maspin suppression remains unclear. Restoration of maspin expression by 5-aza-dc/TSA can synergistically enhance myocardin–induced apoptosis in MCF-7 cells, suggesting the involvement of promoter hypermethylation and histone hypoacetylation in maspin gene silence [52–54]. Although nuclear cytokine-activated IKKα was reported to repress maspin through DNA methylation in prostate cancer metastasis, the DNA demethylation drug 5-aza-2′-deoxycytidine didn’t restore the maspin expression in Hep3Bx cell (our unpublished data). Additionally, the maspin-3′UTR activity and maspin mRNA stability were significantly disrupted by nuclear IKKα, indicating that nuclear IKKα mainly suppressed maspin expression through post-transcriptional regulation in HCC. Our study provided a clear mechanism that HBx activated IKKα to nuclear translocation, and then phosphorylated H3 at Ser-10 to facilitate NFκB-mediated miRNA transcriptions to target maspin mRNA. Suppression of maspin by HBx-IKKα-NFκB-miRNA axis played a crucial role in HCC progression and chemoresistance. Blockade of nuclear IKKα function by silencing IKKα and inhibiting its nuclear translocation significantly retarded these microRNAs and their host gene expressions, and restored maspin protein expression and re-sensitized HBx-expressing HCC cells to doxorubicin. These data raised the possibility that combination treatment with IKKα specific inhibitor to restore maspin expression may improve the chemotherapeutic responses in HBV-associated HCC patients.
Recent studies have shown that microRNAs play essential roles in tumorigenesis, metastasis, and chemoresistance through the post-transcriptional regulation of tumor associated-genes [55–57]. MicroRNA-7 has been proposed controversially to function as an oncogene or a tumor-suppressor in different cancer types. Upregulation of microRNA-7 was observed in renal cell carcinoma and plays an important role in migration, cell proliferation, and apoptosis [58]; however, microRNA-7 inhibited metastasis and invasion through targeting focal adhesion kinase in cervical cancer [59]. In our previous study, induction of microRNA-7 by trichostatin A (TSA) suppressed the off-target effect of lapatinib on EGFR up-regulation, thus overcome the metastatic ability of HER2-negative breast cancer cells [60–62]. Although HBx-induced microRNA-7 downregulated EGFR expression to render HCC cells a slow-growth behavior [63], the expression of microRNA-7 was demonstrated regulated by nuclear IKKα and served as a oncogene to promote metastasis and chemoresistance through suppressing maspin expression in this study. Additionally, resveratrol increased the expressions of tumor suppressors, PDCD4 and maspin, to reduce prostate cancer growth and metastasis by inhibiting the Akt/microRNA-21 pathway [64]. These studies are consistent with our finding that HBx-activated nuclear IKKα transcriptionally upregulated maspin-targeting microRNA-7/-21/-103/-107 expression in HCC tumor progression. MicroRNA-103/107 have been identified as a invasive predictor of tumor relapse and overall survival for triple-negative breast cancer patients [65]; additionally, microRNA-103/-107 also modulated multiple drug resistance in human gastric carcinoma by downregulating caveolin-1 [66]. In contrast, microRNA-103/107 overexpression was considered as a possible chemosensitizer to promote genomic instability [67]. Interestingly, in high metastatic breast cancer cells, overexpression of microRNA-103/107 targeted and degraded NF-κB-interacting LncRNA, NKILA, which can interact with NF-κB/IκB to prevent overactivation of NF-κB pathway [68]. According to these studies, microRNA-103 and 107, which were transcriptionally upregulated by HBx-activated nuclear IKKα, might also negatively regulate the NKILA expression, thus provided positive feedback for enforcing NF-κB activation to suppress maspin expression and promote tumor progression in HCC cells.
Taken together, our study demonstrated that HBx transcriptionally enhanced the levels of microRNA-7, -103, -107, and -21 in a nuclear IKKα/NF-κB- manner. These elevated miRNAs directly targeted and suppressed maspin expression to promote HCC tumor progression and were strongly associated with the poor survival of HBV-related HCC patients (Figure 7). This study not only provides the molecular insight into the nuclear IKKα-mediated maspin suppression in response to HBx, but also highlights the possibility of IKKα-targeting therapy in the treatment of HCC patients.
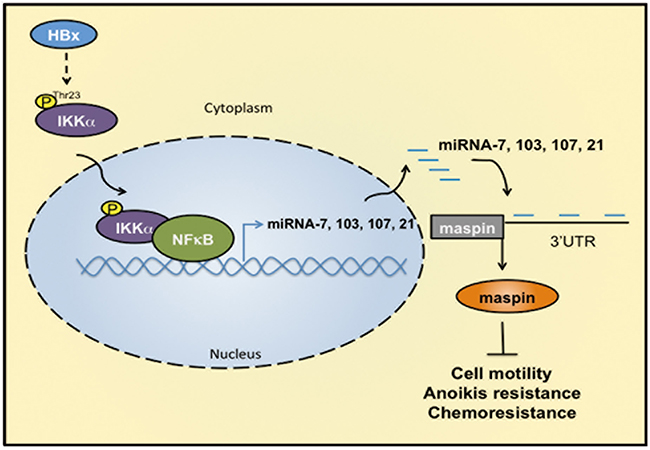
Figure 7: IKKα/NFκB-dependent microRNAs contribute to HBx-mediated hepatocellular tumor progression through suppression of maspin. HBx induces IKKα phosphorylation at Thr-23, leading to its nuclear translocation. Nuclear IKKα cooperated with NFκB to induce miRNA-7, -103, -107, and -21 transcriptions for maspin downregulation by targeting its 3′UTR. Suppression of maspin leads to the motility, anoikis resistance, and chemoresistance of HCC cells.
MATERIALS AND METHODS
Cell culture
HEK-293, Hep3B, and Hep3Bx cell lines were cultured in Dulbecco’s modified Eagle’s medium/F12 medium supplemented with 10% fetal bovine serum.
Plasmids, antibodies, and chemicals
Maspin-3′UTR was constructed in to pMIR-REPORT™ Luciferase plasmid (Ambion, Austin, TX, USA) [28]. Mutations of indicated sites in maspin-3′UTR and Flag-IKKα were generated using a Quickchange site-directed mutagenesis kit (Stratagene, La Jolla, CA, USA) according to the manufacturer’s instructions. All of the above plasmids were confirmed by DNA sequencing. We purchased antibodies against IKKα, phospho-IKKα Thr23, IKKβ, and maspin from Santa Cruz (Santa Cruz, CA, USA), antibody against Flag-tag from Sigma-Aldrich (St. Louis, MO, USA), and antibody against phospho-IKKα Thr-23 (for IHC) from Abnova, The validated miRNA inhibitors were purchased from Dharmacon (Lafayette, CO, USA). The validated shRNA for negative control and IKKα were purchased from National RNAi Core Facility at Academia Sinica (Taipei, Taiwan). Tripure isolation reagent for RNA isolation was purchased from Roche (Indianapolis, IN, USA). The MMLV First-Strand cDNA Synthesis kit and Universal probelibrary Probe#21 was purchased from Roche (Indianapolis, IN, USA). The KAPA SYBR® FAST Master Mix (2X) and KAPA Probe FAST Universal qPCR Kit were purchased from Kapa biosystem (Woburn, MA). The chemotherapeutic drugs, doxorubicin hydrochloride was purchased from Sigma-Aldrich (St. Louis, MO, USA). Luciferase assay system was purchased from Promega (Madison, WI, USA). Actinomycin D was purchased from Sigma-Aldrich (St. Louis, MO).
Clinical specimens
HCC tissue sections and specimens were purchased from Taiwan Liver Cancer Network, Zhunan, Taiwan and provided from National Cheng Kung University Hospital, Tainan, Taiwan. Informed consents were signed by patients with approval by the Institutional Review Board, China Medical University Hospital, Taichung, Taiwan (DMR101-IRB1-119) and by the Institutional Review Board of the Human Investigation Committee of College of Medicine, National Cheng Kung University Tainan, Taiwan (B-ER-102-210). The clinical sample information met REMARK (REporting recommendations for tumor MARKer prognostic studies) guideline was shown in our previous study [28]
Immunohistochemical Staining (IHC)
Five-micron thick paraffin-embedded tissue sections were deparaffinized and rehydrated. After antigen retrieval, the tissue sections were treated with Peroxidase Block, Protein block, and subsequently incubated with rabbit monoclonal anti-human maspin and anti-phospho-IKKα (Thr23) antibodies (100 dilution, Santa Cruz and Abnova, respectively) at 4°C for overnight. After washing to remove unbound primary antibody, sections were treated with a NovoLink Polymer anti-mouse/rabbit IgG-Poly-HRP according to manufacturer’s instructions (NovoLinkTM polymer detection system, Leica) for 30 minutes. Tissue sections were incubated in the chromogenic peroxidase substrate, diaminobenzidine (DAB), for 30 second or 5 minutes, and subsequently counterstain with Hematoxylin for nucleus staining. The specificity of labeling by this procedure was verified by negative control reactions using buffer to replace the primary antibody and isotype-specific IgG.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide (MTT) cell viability assay
In vitro cell viability was measured using an MTT colorimetric assay. Hep3Bx cells (1×104 cells/well) were seed in 96-well plate. After IKK inhibitor VII treatment, the culture medium was removed and 1μg/ml MTT solution (Sigma, St. Louis, MO, USA) was added to incubate for 3 hours. Finally, DMSO was added to lyse the cells and the absorbance at OD550 wavelength was detected by ELISA reader.
Reporter gene luciferase assay
As described previously [69], cells with 60–80% of confluence were transfected with maspin-3′UTR luciferase plasmids and maspin promoter plasmid along with or without miRNA inhibitors, myc-HBx, or IKKα expression vectors. After 48hrs of transfection, cell lysates were harvested and subjected to luciferase assay system. Luciferase activity was normalized to β-gal activity.
Preparation and infection of shRNA-IKKα expressing lentivirus
Briefly, 2 μg pCMV-dR8.91, 200 μg pMD2.G, and 2 μg pLKO-shLuciferase, or pLKO-shIKKα were cotransfected into HEK293T cells using Lipofectamine 2000. The supernatants containing infectious lentivirus were collected after 1 day of transfection. For lentivirus infection, cells (2×105) were infected with lentivirus at a multiplicity of infection (MOI). After 5 days infection, cells were harvested for protein or RNA extraction.
Quantitative real-time PCR (RT-qPCR)
Total RNA was extracted by using Tripure isolation reagent according previous report [70]. One μg of RNA was subjected to reverse transcription with the MMLV First-Strand cDNA Synthesis kit. After reverse transcription, the qPCR analysis of maspin, IKKα, PGSF1, PANK2, and PANK1 mRNA expressions was performed on ABI 7500 system (Applied Biosystems, Foster, CA) by using KAPA SYBR® FAST Master Mix (2X) and was normalized to GAPDH expression. The qPCR analysis of miR-7, -103, -107, and -21 expression was performed on LightCycler 480 System (Roche, Indianapolis, IN, USA) by using KAPA Probe FAST Universal qPCR Kit and was normalized to U48 expression. Specific primers used in real-time PCR were listed in Supplementary Table.
Chromatin immunoprecipitation assay
Cells were cross-linked with 1.42% formaldehyde for 15 min and quenched unreacted formaldehyde with 0.125 M Glycine solution for 5 min. Cells were scraped in 1 ml of cold PBS, centrifuged, and lysed in 1mL of IP buffer (150mM NaCl, 50mM Tris-HCl, pH 7.5, 5mM EDTA, 0.5% Nonidet P-40, and 1% Triton X-100) containing protease inhibitors (1mM phenylmethylsulfonyl fluoride, 1μM eupeptin and 1μM aprotinin). The nuclear pellet was resuspended in IP buffer and sonicated to shear chromatin. The sonicated lysates were immunoprecipited with antibodies against phospho-H3ser10 followed by the pull-down with protein A/G-Sepharose (Thermo). The immunoprecipitated DNA and input DNA were extracted by incubating with 100 μl of 10% Chelex (Bio-Rad), boiling to reverse the cross-link, and centrifuging to remove Chelex slurry. RT-qPCR was performed with the purified DNA using specific primers as shown in Supplementary Table.
Transient transfection
Cells at 60% confluence were transfected with indicated plasmids or microRNA inhibitors using Nanofectin (PAA, Pasching, Austria), Lipofetamine 2000 (Invitrogen, Boston, USA), or DharmaFECT (Thermo Scientific Dharmacon, Lafayette, CO, USA) as described previously [71]. Nanofectin and Lipofectamine 2000 were used in plasmid DNA transfection, and the DharmaFECT was used in microRNA inhibitor transfection. After transfection for 48 h, cells were subjected to total lysate preparation, total RNA extraction, or luciferase assays.
Protein extraction and immunoblot
For total cell lysates, cells were washed with ice-cold PBS one time and lysed in RIPA buffer (20 mM Tris-HCl, pH7.4, 150 mM NaCl, 1% NP-40, 1% sodium deoxycholate, 1 mM EDTA and 1 mM EGTA) containing protease inhibitors and phosphatase inhibitors cocktails (Roche, Indianapolis, IN, USA). Proteins were separated by SDS-PAGE, transferred to PVDF membrane, and blotted with indicated antibodies.
Statistical analysis
The difference in relative gene expression between tumor and normal tissues was calculated by a two-tailed Student’s t test. Coefficient analyses were performed for the correlation between gene expressions. The percentage of cumulative survival was determined by Kaplan-Meier survival test. The univariate and multivariate analyses were used in Cox proportional hazards models. All these statistical analyses were performed using Sigma Plot 10.0. A p-value < 0.05 was defined as statistically significant.
Abbreviations
Maspin,Mammary serine protease inhibitor;HCC, hepatocellular carcinoma; IKKα kinase alpha; miR, microRNA; HBV, hepatitis virus B; HBx, HBV X protein; NF-κB, nuclear factor kappa B; UTR, untranslational region; NLS, nuclear localization signal; NES, nuclear export signal; shRNA, small-hairpin RNA; CBP, CREB-binding protein; PGSF, pituitary gland specific factor; PANK, pantothenate kinase.
ACKNOWLEDGMENTS
We thank Taiwan Liver Cancer Network (TLCN), which is supported by grants from National Science Council, and National Health Research Institutes in Taiwan and National Cheng Kung University Hospital in Taiwan for providing the HCC tissue sections, specimens and related anonymous clinical data.
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
FINANCIAL SUPPORT
This work was supported by grants from the National Science Council of Taiwan (MOST 103-2320-B-039-052, MOST-103-2320-B-039-001, MOST 104-2911-I-002-303, MOST102-2320-B-214-008-MY3, and MOST 103-2320-B-214-005), the National Health Research Institutes of Taiwan (NHRI-EX104-10329BI), Ministry of Health and Welfare, China Medical University Hospital Cancer Research Center of Excellence (MOHW103-TD-B-111-03 and MOHW103-TDU-212-114002), and China Medical University Hospital (DMR104070).
Authors’ contributions
Study concept and design: LCL, WCH; acquisition of data: WSC, CJY, and JYC; analysis and interpretation of data: WSC, CJY and CYH; drafting of the manuscript: WCH, YJC, and CCC; critical revision of the manuscript for important intellectual content: LCL, WCH; statistical analysis: WSC, SHL; obtained funding: LCL, WCH; study supervision: WCH.
REFERENCES
1. Lara-Pezzi E, Gomez-Gaviro MV, Galvez BG, Mira E, Iniguez MA, Fresno M, Martinez AC, Arroyo AG and Lopez-Cabrera M. The hepatitis B virus X protein promotes tumor cell invasion by inducing membrane-type matrix metalloproteinase-1 and cyclooxygenase-2 expression. The Journal of clinical investigation. 2002; 110:1831-1838.
2. Lee JO, Lee KW, Oh DY, Kim JH, Im SA, Kim TY and Bang YJ. Combination chemotherapy with capecitabine and cisplatin for patients with metastatic hepatocellular carcinoma. Annals of oncology. 2009; 20:1402-1407.
3. Anwar WA, Khaled HM, Amra HA, El-Nezami H and Loffredo CA. Changing pattern of hepatocellular carcinoma (HCC) and its risk factors in Egypt: possibilities for prevention. Mutation research. 2008; 659:176-184.
4. Parkin DM, Bray F, Ferlay J and Pisani P. Estimating the world cancer burden: Globocan 2000. International Journal of Cancer. 2001; 94:153-156.
5. Bartosch B. Hepatitis B and C viruses and hepatocellular carcinoma. Viruses. 2010; 2:1504-1509.
6. Fink SA and Jacobson IM. Managing patients with hepatitisB-related or hepatitisC-related decompensated cirrhosis. Nature reviews Gastroenterology & hepatology. 2011; 8:285-295.
7. McGlynn KA and London WT. Epidemiology and natural history of hepatocellular carcinoma. Best practice & research Clinical gastroenterology. 2005; 19:3-23.
8. Ali A, Abdel-Hafiz H, Suhail M, Al-Mars A, Zakaria MK, Fatima K, Ahmad S, Azhar E, Chaudhary A and Qadri I. Hepatitis B virus, HBx mutants and their role in hepatocellular carcinoma. World journal of gastroenterology. 2014; 20:10238-10248.
9. Huang WC, Hsieh YL, Hung CM, Chien PH, Chien YF, Chen LC, Tu CY, Chen CH, Hsu SC, Lin YM and Chen YJ. BCRP/ABCG2 inhibition sensitizes hepatocellular carcinoma cells to sorafenib. PloS one. 2013; 8:e83627.
10. Mai L, Yang L, Kuang JY, Zhu JY, Kang YH, Zhang FC, Xie QF and Gao ZL. [Hepatitis B virus X promotes HepG2 cell cycle progression and growth via downregulation expression of p16 protein]. [Article in Chinese]. Zhonghua gan zang bing za zhi. 2013; 21:614-618.
11. Wang HY, Yang SL, Liang HF and Li CH. HBx protein promotes oval cell proliferation by up-regulation of cyclin D1 via activation of the MEK/ERK and PI3K/Akt pathways. International journal of molecular sciences. 2014; 15:3507-3518.
12. Khattar E, Mukherji A and Kumar V. Akt augments the oncogenic potential of the HBx protein of hepatitis B virus by phosphorylation. The FEBS journal. 2012; 279:1220-1230.
13. Chen X, Zhang L, Zheng S, Zhang T, Li M, Zhang X, Zeng Z, McCrae MA, Zhao J, Zhuang H and Lu F. Hepatitis B Virus X Protein Stabilizes Cyclin D1 and Increases Cyclin D1 Nuclear Accumulation through ERK-Mediated Inactivation of GSK-3beta. Cancer prevention research. 2015; 8:455-463.
14. Chung TW, Lee YC and Kim CH. Hepatitis B viral HBx induces matrix metalloproteinase-9 gene expression through activation of ERK and PI-3K/AKT pathways: involvement of invasive potential. FASEB journal. 2004; 18:1123-1125.
15. Lee JO, Kwun HJ, Jung JK, Choi KH, Min DS and Jang KL. Hepatitis B virus X protein represses E-cadherin expression via activation of DNA methyltransferase 1. Oncogene. 2005; 24:6617-6625.
16. Zhang X, Liu S, Hu T, Liu S, He Y and Sun S. Up-regulated microRNA-143 transcribed by nuclear factor kappa B enhances hepatocarcinoma metastasis by repressing fibronectin expression. Hepatology. 2009; 50:490-499.
17. Li C, Sun Y, Geng L and Fan F. [HBx silencing by shRNA down-regulates MMP-2 expression in human hepatocellular carcinoma cell HepG2.2.15]. [Article in Chinese]. Xi bao yu fen zi mian yi xue za zhi. 2014; 30:371-374.
18. Yu FL, Liu HJ, Lee JW, Liao MH and Shih WL. Hepatitis B virus X protein promotes cell migration by inducing matrix metalloproteinase-3. Journal of hepatology. 2005; 42:520-527.
19. Kapoor NR, Ahuja R, Shukla SK and Kumar V. The HBx protein of hepatitis B virus confers resistance against nucleolar stress and anti-cancer drug-induced p53 expression. FEBS letters. 2013; 587:1287-1292.
20. Gottlob K, Fulco M, Levrero M and Graessmann A. The hepatitis B virus HBx protein inhibits caspase 3 activity. The Journal of biological chemistry. 1998; 273:33347-33353.
21. Kuo TC and Chao CC. Hepatitis B virus X protein prevents apoptosis of hepatocellular carcinoma cells by upregulating SATB1 and HURP expression. Biochemical pharmacology. 2010; 80:1093-1102.
22. Zhu H, Luo SF, Wang J, Li X, Wang H, Pu WY, Zhang H and Zhuang ZX. Effect of environmental factors on chemoresistance of HepG2 cells by regulating hypoxia-inducible factor-1alpha. Chinese medical journal. 2012; 125:1095-1103.
23. Cheng AS, Wong N, Tse AM, Chan KY, Chan KK, Sung JJ and Chan HL. RNA interference targeting HBx suppresses tumor growth and enhances cisplatin chemosensitivity in human hepatocellular carcinoma. Cancer letters. 2007; 253:43-52.
24. He Y, Sun HQ, He XE, Wang WL and Lei JH. Knockdown of HBx by RNAi inhibits proliferation and enhances chemotherapy-induced apoptosis in hepatocellular carcinoma cells. Medical oncology. 2010; 27:1227-1233.
25. Kim HY, Jung HU, Yoo SH, Yoo KS, Cheong J, Park BS, Yun I and Yoo YH. Sorafenib overcomes the chemoresistance in HBx-expressing hepatocellular carcinoma cells through down-regulation of HBx protein stability and suppresses HBV gene expression. Cancer letters. 2014; 355:61-69.
26. Tan YL and Chen WN. MicroRNAs as therapeutic strategy for hepatitis B virus-associated hepatocellular carcinoma: current status and future prospects. World journal of gastroenterology. 2014; 20:5973-5986.
27. Lamontagne J, Steel LF and Bouchard MJ. Hepatitis B virus and microRNAs: Complex interactions affecting hepatitis B virus replication and hepatitis B virus-associated diseases. World journal of gastroenterology. 2015; 21:7375-7399.
28. Chen WS, Yen CJ, Chen YJ, Chen JY, Wang LY, Chiu SJ, Shih WL, Ho CY, Wei TT, Pan HL, Chien PH, Hung MC, Chen CC and Huang WC. miRNA-7/21/107 contribute to HBx-induced hepatocellular carcinoma progression through suppression of maspin. Oncotarget. 2015; 6:25962-25974. doi: 10.18632/oncotarget.4504.
29. Ruland J. Return to homeostasis: downregulation of NF-kappaB responses. Nature immunology. 2011; 12:709-714.
30. Jiang R, Xia Y, Li J, Deng L, Zhao L, Shi J, Wang X and Sun B. High expression levels of IKKalpha and IKKbeta are necessary for the malignant properties of liver cancer. International journal of cancer. 2010; 126:1263-1274.
31. Marquardt JU, Gomez-Quiroz L, Arreguin Camacho LO, Pinna F, Lee YH, Kitade M, Dominguez MP, Castven D, Breuhahn K, Conner EA, Galle PR, Andersen JB, Factor VM and Thorgeirsson SS. Curcumin effectively inhibits oncogenic NF-kB signaling and restrains stemness features in liver cancer. Journal of hepatology. 2015.
32. Kew MC. Hepatitis B virus x protein in the pathogenesis of hepatitis B virus-induced hepatocellular carcinoma. Journal of gastroenterology and hepatology. 2011; 26 Suppl 1:144-152.
33. Huang WC, Chen WS, Chen YJ, Wang LY, Hsu SC, Chen CC and Hung MC. Hepatitis B virus X protein induces IKKalpha nuclear translocation via Akt-dependent phosphorylation to promote the motility of hepatocarcinoma cells. Journal of cellular physiology. 2012; 227:1446-1454.
34. Luo JL, Tan W, Ricono JM, Korchynskyi O, Zhang M, Gonias SL, Cheresh DA and Karin M. Nuclear cytokine-activated IKKalpha controls prostate cancer metastasis by repressing Maspin. Nature. 2007; 446:690-694.
35. Yamamoto Y, Verma UN, Prajapati S, Kwak YT and Gaynor RB. Histone H3 phosphorylation by IKK-alpha is critical for cytokine-induced gene expression. Nature. 2003; 423:655-659.
36. Yoshida K, Ozaki T, Furuya K, Nakanishi M, Kikuchi H, Yamamoto H, Ono S, Koda T, Omura K and Nakagawara A. ATM-dependent nuclear accumulation of IKK-alpha plays an important role in the regulation of p73-mediated apoptosis in response to cisplatin. Oncogene. 2008; 27:1183-1188.
37. Fernandez-Majada V, Aguilera C, Villanueva A, Vilardell F, Robert-Moreno A, Aytes A, Real FX, Capella G, Mayo MW, Espinosa L and Bigas A. Nuclear IKK activity leads to dysregulated notch-dependent gene expression in colorectal cancer. Proceedings of the National Academy of Sciences of the United States of America. 2007; 104:276-281.
38. Nowak SJ and Corces VG. Phosphorylation of histone H3: a balancing act between chromosome condensation and transcriptional activation. Trends in genetics. 2004; 20:214-220.
39. Huang WC, Ju TK, Hung MC and Chen CC. Phosphorylation of CBP by IKKalpha promotes cell growth by switching the binding preference of CBP from p53 to NF-kappaB. Molecular cell. 2007; 26:75-87.
40. Burden HE and Weng Z. Identification of conserved structural features at sequentially degenerate locations in transcription factor binding sites. Genome informatics International Conference on Genome Informatics. 2005; 16:49-58.
41. Sandelin A, Alkema W, Engstrom P, Wasserman WW and Lenhard B. JASPAR: an open-access database for eukaryotic transcription factor binding profiles. Nucleic acids research. 2004; 32:D91-94.
42. Tsunoda T and Takagi T. Estimating transcription factor bindability on DNA. Bioinformatics. 1999; 15:622-630.
43. Perez-Cadahia B, Drobic B and Davie JR. H3 phosphorylation: dual role in mitosis and interphase. Biochemistry and cell biology. 2009; 87:695-709.
44. Allavena P, Garlanda C, Borrello MG, Sica A and Mantovani A. Pathways connecting inflammation and cancer. Current opinion in genetics & development. 2008; 18:3-10.
45. Sun B and Karin M. NF-kappaB signaling, liver disease and hepatoprotective agents. Oncogene. 2008; 27:6228-6244.
46. Hacker H and Karin M. Regulation and function of IKK and IKK-related kinases. Science's STKE. 2006; 2006:re13.
47. Scheidereit C. IkappaB kinase complexes: gateways to NF-kappaB activation and transcription. Oncogene. 2006; 25:6685-6705.
48. Sheng S, Carey J, Seftor EA, Dias L, Hendrix MJ and Sager R. Maspin acts at the cell membrane to inhibit invasion and motility of mammary and prostatic cancer cells. Proceedings of the National Academy of Sciences of the United States of America. 1996; 93:11669-11674.
49. Zhang M, Volpert O, Shi YH and Bouck N. Maspin is an angiogenesis inhibitor. Nature medicine. 2000; 6:196-199.
50. Jiang N, Meng Y, Zhang S, Mensah-Osman E and Sheng S. Maspin sensitizes breast carcinoma cells to induced apoptosis. Oncogene. 2002; 21:4089-4098.
51. Bodenstine TM, Seftor RE, Khalkhali-Ellis Z, Seftor EA, Pemberton PA and Hendrix MJ. Maspin: molecular mechanisms and therapeutic implications. Cancer metastasis reviews. 2012; 31:529-551.
52. Maass N, Biallek M, Rosel F, Schem C, Ohike N, Zhang M, Jonat W and Nagasaki K. Hypermethylation and histone deacetylation lead to silencing of the maspin gene in human breast cancer. Biochemical and biophysical research communications. 2002; 297:125-128.
53. Futscher BW, Oshiro MM, Wozniak RJ, Holtan N, Hanigan CL, Duan H and Domann FE. Role for DNA methylation in the control of cell type specific maspin expression. Nature genetics. 2002; 31:175-179.
54. Liao XH, Li YQ, Wang N, Zheng L, Xing WJ, Zhao DW, Yan TB, Wang Y, Liu LY, Sun XG, Hu P, Zhou H and Zhang TC. Re-expression and epigenetic modification of maspin induced apoptosis in MCF-7 cells mediated by myocardin. Cellular signalling. 2014; 26:1335-1346.
55. Liu LN, Li DD, Xu HX, Zheng SG and Zhang XP. Role of microRNAs in hepatocellular carcinoma. Frontiers in bioscience. 2015; 20:1056-1067.
56. Srivastava SK, Arora S, Averett C, Singh S and Singh AP. Modulation of microRNAs by phytochemicals in cancer: underlying mechanisms and translational significance. BioMed research international. 2015; 2015:848710.
57. Neelakandan K, Babu P and Nair S. Emerging roles for modulation of microRNA signatures in cancer chemoprevention. Current cancer drug targets. 2012; 12:716-740.
58. Yu Z, Ni L, Chen D, Zhang Q, Su Z, Wang Y, Yu W, Wu X, Ye J, Yang S, Lai Y and Li X. Identification of miR-7 as an oncogene in renal cell carcinoma. Journal of molecular histology. 2013; 44:669-677.
59. Hao Z, Yang J, Wang C, Li Y, Zhang Y, Dong X, Zhou L, Liu J, Zhang Y and Qian J. MicroRNA-7 inhibits metastasis and invasion through targeting focal adhesion kinase in cervical cancer. International journal of clinical and experimental medicine. 2015; 8:480-487.
60. Tu CY, Chen CH, Hsia TC, Hsu MH, Wei YL, Yu MC, Chen WS, Hsu KW, Yeh MH, Liu LC, Chen YJ and Huang WC. Trichostatin A suppresses EGFR expression through induction of microRNA-7 in an HDAC-independent manner in lapatinib-treated cells. BioMed research international. 2014; 2014:168949.
61. Hsia TC, Tu CY, Chen YJ, Wei YL, Yu MC, Hsu SC, Tsai SL, Chen WS, Yeh MH, Yen CJ, Yu YL, Huang TC, Huang CY, Hung MC and Huang WC. Lapatinib-mediated cyclooxygenase-2 expression via epidermal growth factor receptor/HuR interaction enhances the aggressiveness of triple-negative breast cancer cells. Molecular pharmacology. 2013; 83:857-869.
62. Chen YJ, Yeh MH, Yu MC, Wei YL, Chen WS, Chen JY, Shih CY, Tu CY, Chen CH, Hsia TC, Chien PH, Liu SH, Yu YL and Huang WC. Lapatinib-induced NF-kappaB activation sensitizes triple-negative breast cancer cells to proteasome inhibitors. Breast cancer research. 2013; 15:R108.
63. Chen YJ, Chien PH, Chen WS, Chien YF, Hsu YY, Wang LY, Chen JY, Lin CW, Huang TC, Yu YL and Huang WC. Hepatitis B Virus-Encoded X Protein Downregulates EGFR Expression via Inducing MicroRNA-7 in Hepatocellular Carcinoma Cells. Evidence-based complementary and alternative medicine. 2013; 2013:682380.
64. Sheth S, Jajoo S, Kaur T, Mukherjea D, Sheehan K, Rybak LP and Ramkumar V. Resveratrol reduces prostate cancer growth and metastasis by inhibiting the Akt/MicroRNA-21 pathway. PloS one. 2012; 7:e51655.
65. Kleivi Sahlberg K, Bottai G, Naume B, Burwinkel B, Calin GA, Borresen-Dale AL and Santarpia L. A serum microRNA signature predicts tumor relapse and survival in triple-negative breast cancer patients. Clinical cancer research. 2015; 21:1207-1214.
66. Zhang Y, Qu X, Li C, Fan Y, Che X, Wang X, Cai Y, Hu X and Liu Y. miR-103/107 modulates multidrug resistance in human gastric carcinoma by downregulating Cav-1. Tumour biology. 2015; 36:2277-2285.
67. Huang JW, Wang Y, Dhillon KK, Calses P, Villegas E, Mitchell PS, Tewari M, Kemp CJ and Taniguchi T. Systematic screen identifies miRNAs that target RAD51 and RAD51D to enhance chemosensitivity. Molecular cancer research. 2013; 11:1564-1573.
68. Liu B, Sun L, Liu Q, Gong C, Yao Y, Lv X, Lin L, Yao H, Su F, Li D, Zeng M and Song E. A cytoplasmic NF-kappaB interacting long noncoding RNA blocks IkappaB phosphorylation and suppresses breast cancer metastasis. Cancer cell. 2015; 27:370-381.
69. Hsiao YC, Yeh MH, Chen YJ, Liu JF, Tang CH and Huang WC. Lapatinib increases motility of triple-negative breast cancer cells by decreasing miRNA-7 and inducing Raf-1/MAPK-dependent interleukin-6. Oncotarget. 2015; 6:37965-37978. doi: 10.18632/oncotarget.5700.
70. Chen JY, Chen YJ, Yen CJ, Chen WS and Huang WC. HBx sensitizes hepatocellular carcinoma cells to lapatinib by up-regulating ErbB3. Oncotarget. 2016; 7:473-89. doi: 10.18632/oncotarget.6337.
71. Huang WC, Hsu SC, Huang SJ, Chen YJ, Hsiao YC, Zhang W, Fidler IJ and Hung MC. Exogenous expression of human SGLT1 exhibits aggregations in sodium dodecyl sulfate polyacrylamide gel electrophoresis. American journal of translational research. 2013; 5:441-449.