
INTRODUCTION
Extensive research from in vitro, in vivo and clinical studies has now established that Toll-like receptors (TLRs) are important in the development of carcinogenesis and tumor progression of a variety of cancers. Indeed, heightened expression of TLRs is evidenced in multiple cancer cell lines, and in tumor cells and tissues obtained from patients [1–3]. Furthermore, activation of TLRs on cancer cells promotes chronic inflammation which stimulates cancer cell proliferation, migration, tumor angiogenesis and creates a tumor microenvironment which impairs the anti-tumor function of the immune system allowing tumors to develop and survive [4].
Much like TLRs, the Wnt family proteins have also been implicated in carcinogenesis - in both the initiation and maintenance of cancer [5–7]. In vitro studies with various cancer cell lines and tumor specimens from patients have revealed elevated expression of Wnt proteins (e.g., Wnt5a) and deregulated Wnt signal transduction pathways [5, 8–11]. The elevated expression of Wnt proteins (e.g., Wnt5a) in tumor samples correlates with advanced stages and poor prognosis in solid tumors as well as hematological malignancies [5, 8–11].
The TLR-mediated pro-inflammatory cytokine, IL-6, is another integral molecular effector of initiation and growth of malignant tumors. This pro-inflammatory cytokine is implicated in a milieu of cancer types because of its propensity to drive the activation of the oncogenic transcription factor signal transducer and activator of transcription 3 (STAT3). By virtue of interacting with other transcription factors (e.g., NF-B), STAT3 is known to mediate crosstalk between tumor cells and inflammatory cells within the tumor microenvironment and promote the development and progression of multiple types of human cancers [12–17]. The expression of STAT3 and presence of its active form (i.e., phosphorylated form) is increased in tumor tissues from patients and this pronounced expression indicates a poor prognosis in a variety of cancers [18].
Although much fundamental, translational and clinical research has been conducted on TLRs, Wnts and IL-6/STAT3, the majority of the investigations have studied these cardinal mediators of oncogenesis in “isolation”. Only recently has there has been an increasing interest in “co-associating” and tying these aforementioned oncologic mediators together. An IL-6/STAT3/Wnt5a signaling loop has been described by different groups [19, 20]. Our group demonstrated that IL-6, a TLR signaling product, can activate STAT3 with resulting overexpression of Wnt5a in human papillary thyroid carcinoma, melanoma and pancreatic cancer [1, 2]. Importantly, a novel inhibitor of pathologic TLR3 signaling, phenylmethimazole (C10), has the ability to block IL-6 production, as well as decrease viability/growth and migration in these same cancers [2, 20]. We hypothesized that the effect of C10 on viability/growth and migration of these cancers was related to its suppressive effect on TLR3 signaling which led to the inhibition of TLR-mediated STAT3 and Wnt5a signaling [20]. Moreover, in these reports we showed that C10 also inhibited tumor growth in mouse xenograft models of human pancreatic cancer and malignant melanoma thus indicating the potential of using a TLR inhibitor for the treatment of TLR-expressing cancers [2]. In this present study, we sought to extend our earlier observations of TLR3 and Wnt5a association and their role in tumor viability/growth and migration in human breast cancer. We also evaluated the ability of C10 to synergistically inhibit tumor cell viability/growth in combination with tamoxifen, the current standard of care therapy for breast cancer.
RESULTS
C10 significantly decreases high basal TLR3 and Wnt5a expression in human breast cancer cells
Previous studies in our laboratory demonstrated that C10 decreased high constitutive TLR3 and Wnt5a expression in human malignant melanoma, papillary thyroid, and pancreatic cancer cell lines [1, 2]. In these studies we sought to evaluate the effect of the same TLR signaling inhibitor, C10, on human MCF-7 breast cancer cells that exhibit high constitutive levels of TLR3 and Wnt5a RNA. As was the case in our previous studies, we observed significant decreases in TLR3 RNA levels following C10 treatment in MCF-7 cells (Figure 1A). Similarly, MCF-7 cells also exhibited significant decreases in Wnt5a levels after C10 treatment (Figure 1B).
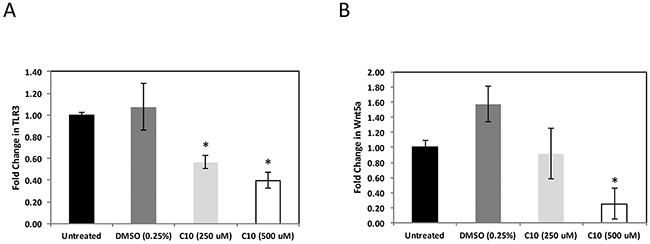
Figure 1: C10 significantly decreases high TLR3 and Wnt5a expression in MCF-7 cells. MCF-7 human breast cancer cells were grown to approximately 100% confluency. Subsequently, cells were treated with 250-500 μM C10 or vehicle control (0.25% DMSO). Following 24 hr. incubation, cells were harvested, total RNA was collected and gene-specific qRT-PCR was performed for A. TLR3 and B. Wnt5a. *Indicates significant difference from untreated and DMSO groups, p < 0.05. No significant differences between untreated and DMSO groups. One-way ANOVA with Tukey-Kramer post-hoc analysis. Results are representative of n = 3 separate experiments.
C10 significantly decreases IL-6 protein levels and subsequent STAT3Y705 phosphorylation
IL-6 is a pro-inflammatory cytokine regulated by TLR3 and has been shown to be important for the growth and migration of cancer cells [21–23]. It has also been shown to be an activator of a key oncogene, STAT3, known to mediate uncontrolled cell growth in many types of human cancers [24]. STAT3 is phosphorylated on multiple residues for activation; phosphorylation of both its tyrosine 705 (pSTAT3Y705) and serine 727 (pSTAT3S727) residues are necessary for maximal transcriptional activity [25–28]. Based on our previous findings that C10 significantly reduced IL-6 protein levels and STAT3 phosphorylated via TLR3 signaling in other cancers, we also evaluated these in MCF-7 breast cancer cells. IL-6 protein levels were analyzed via ELISA. We observed significantly reduced (> 90%) IL-6 protein levels following C10 treatment of MCF-7 cells (Figure 2A). Additionally, we observed a significant decrease in IL-6-induced phosphorylated STAT3Y705 in MCF-7 (Figure 2B). Of note, C10 did not affect STAT3S727 phosphorylation thus reflecting individual variations of STAT3 phosphorylation among different cancers (data not shown). These data, along with results shown in Figure 1, indicate that the constitutive levels of TLR3 are abnormally high in MCF-7 cells and that downstream signaling (Wnt5a, IL-6 and STAT3) can be inhibited by C10.
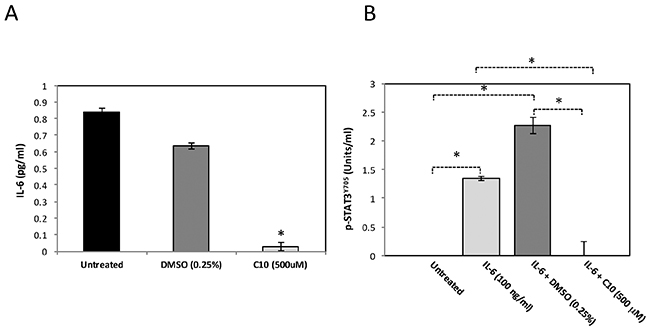
Figure 2: C10 significantly inhibits IL-6 levels and STAT3 phosphorylation in MCF-7 cells. A. MCF-7 human breast cancer cells were grown to approximately 100% confluency. Cells were then treated with 500 μM C10 or vehicle control (0.25% DMSO). Following a 24 hr. incubation, IL-6 levels were analyzed using ELISA. *Indicates significant difference from untreated and DMSO groups, p < 0.0000001. One-way ANOVA with Tukey-Kramer post-hoc analysis. Results presented are representative of n = 3 separate experiments. B. MCF-7 human breast cancer cells were treated with IL-6 for 10 min. to induce STAT3 phosphorylation. In certain cases, MCF-7 cells were pre-treated with vehicle control (0.25% DMSO) or C10 (500 μM) for 1 hr prior to IL-6 stimulation. Subsequently, nuclear protein was extracted and pSTAT3Y705 levels were analyzed using ELISA. *Indicates p < 0.0000001. One-way ANOVA with Tukey-Kramer post-hoc analysis. Results presented are representative of n = 3 separate experiments.
C10 inhibits migration and viability/growth of MCF-7 cells and synergistically enhances the therapeutic effect of tamoxifen in a dose-dependent manner
In addition to its effect on constitutive TLR3, Wnt5a levels and subsequent IL-6 production and STAT3 levels/activation, we then evaluated the ability of C10 to inhibit the migration and viability/growth of MCF-7 breast cancer cells. In a scratch assay, cell migration to cover a scratched area on the plate surface containing cells is measured; retention of a visible scratch line as in Figure 3A is evidence of inhibition of cell migration [5]. Treatment of MCF-7 cells with C10 led to significant inhibition of motility/migration as measured using scratch assays at 0, 24 and 48 hours (p < 0.05) (Figure 3A & 3B).
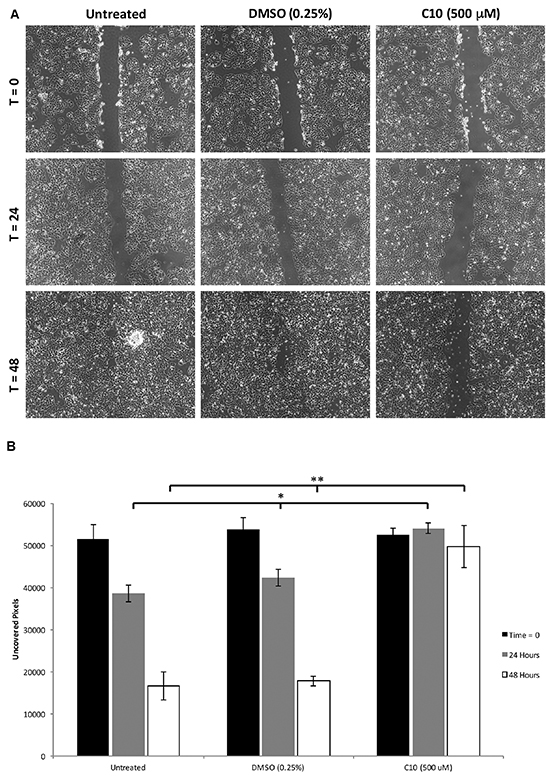
Figure 3: C10 significantly inhibits MCF-7 migration. MCF-7 cells were grown to confluency and migration was analyzed using a scratch assay. After the scratch was performed, cells were treated with 500 μM C10 or vehicle control (0.25% DMSO) for 0, 24 and 48 hours. A. Representative images of the scratch assay depicting inhibition of migration of MCF7 cells after C10 treatment. B. Quantification of cells between each treatment group at 0, 24 and 48 hours. C10 significantly (*p < 0.05) inhibits cell migration when compared to DMSO control and untreated groups. Results presented are representative of at least n = 3 separate experiments.
Tamoxifen is the current standard of care therapy for all stages of breast cancer; however, resistance to the drug has been a major concern [29]. The development of drugs that can be used in combination with tamoxifen may help to increase its effectiveness in treating breast cancer. Since C10 decreased tumor cell viability/growth of other types of human cancer cells, we then sought to determine if C10 could work alone or synergistically in combination with tamoxifen to further inhibit tumor cell viability/growth. As shown in Figure 4, treatment with C10 resulted in significantly reduced MCF-7 cell viability/growth. Moreover, C10 in combination with tamoxifen resulted in a more robust inhibitory effect of tamoxifen on cell viability/growth of MCF-7 cells (Figure 4). These results indicate that the combination of a pathologic TLR3 inhibitor (e.g. C10) and tamoxifen could potentially be of interest for treating breast cancer.
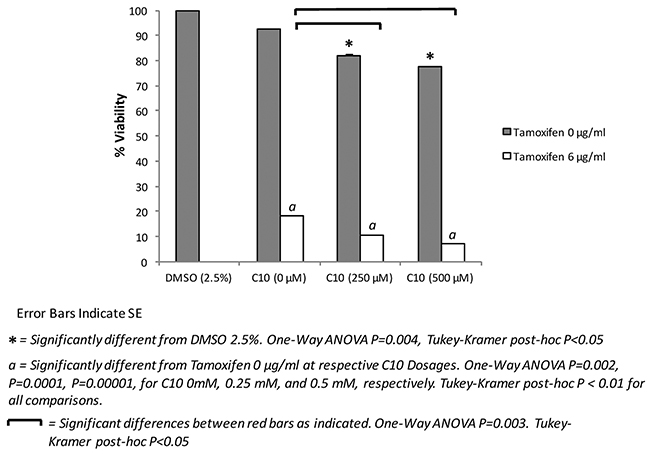
Figure 4: Combination of tamoxifen and C10 significantly inhibits MCF-7 cell viability/growth over Tamoxifen or C10 alone. MCF-7 human breast cancer cells were plated onto a 96-well plate, for 72 hrs and then were treated with C10 (0, 250, or 500 μM), vehicle control (0.25% DMSO), tamoxifen (6 μg/ml) or a combination thereof. A MTT-based cell viability assay was then performed to assess cellular viability/growth at 72 hours. C10 further decreases cellular viability/growth in combination with tamoxifen in a dose-dependent manner. Error bars indicate SE. * indicates significant difference from DMSO 2.5%. One-Way ANOVA P=0.004, Tukey-Kramer post-hoc P<0.05. a indicates significantly different from Tamoxifen 0 µg/ml at respective C10 Dosages. One-Way ANOVA P=0.002, P=0.0001, P=0.00001, for C10 0mM, 0.25 mM, and 0.5 mM, respectively. Tukey-Kramer post-hoc P < 0.01 for all comparisons. Bars indicate significant differences as indicated. One-Way ANOVA P=0.003. Tukey-Kramer post-hoc P<0.05. Results presented are representative of at least n = 3 separate experiments.
DISCUSSION
A significant number of women across different geographies are afflicted with breast cancer. Estrogen plays a critical role in pathology of the vast majority of these breast cancers. Accordingly, patients are treated with anti-estrogen therapies (e.g., tamoxifen). However, these anti-estrogen therapies have their limitations: (i) some patients have inadequate response to these therapies and (ii) patients can develop resistance to these therapies – eventually relapsing. Hence, there remains a significant unmet medical need to develop better and novel therapeutics (monotherapy or combination therapy) for this common disease.
Recent studies have highlighted the important pathologic role of TLR3 and Wnt5a in progression, sustenance and metastasis of breast cancer [30, 31]. Our group has identified phenylmethimazole (C10), an inhibitor of pathologic TLR3 signaling/expression, and shown that C10, by virtue of inhibiting the TLR3/Wnt5a pathway is significantly efficacious in in-vitro and in-vivo models of different cancers and auto-immune diseases [1, 2, 32, 33]. Accordingly, in this study, we sought to further investigate the effects of C10 on human breast tumor cells alone and in combination with standard of care therapy, tamoxifen. Our results demonstrate that C10 inhibits TLR3 and Wnt5a mRNA expression in MCF-7 breast cancer cells. Subsequently, C10 significantly reduced IL-6 protein levels as well as downstream STAT3Y705 phosphorylation. Functionally, the inhibition of this signaling pathway translates into the inhibition of viability/growth and migration of the MCF-7 breast cancer cells. These observations are in-line with our previous studies examining the inhibition of TLR3 signaling with C10 in various other TLR-expressing cancers and the importance of TLR3 signaling modulation as a potential therapeutic target for breast cancer [1, 2, 34].
Although this current study, as well as previously published data, revealed viability/growth inhibition of various tumor cell lines with a TLR signaling inhibitor, we furthered these experiments to investigate the potental combinational use of a TLR signaling inhibitior with a currently FDA-approved drug used to treat breast cancer, tamoxifen. Strikingly, we observed that C10 in combination with tamoxifen synergistically enhanced the cytotoxic potential of tamoxifen by more than 50% when compared to tamoxifen therapy alone. These data suggest that a TLR3 inhibitor alongside tamoxifen, and potentially other existing ER therapies, may be a potential novel therapeutic approach to enhance current standard of care therapies. Future studies need to be conducted in vivo to determine if this combination could be applicable in the clinic.
MATERIALS AND METHODS
Cells
Human breast cancer cell line MCF-7 was purchased from ATCC (Manassas, VA). Cells were grown in RPMI 1640 supplemented with 2 g/L sodium bicarbonate, 1.4 mmol/L sodium pyruvate, 0.14 mmol/L nonessential amino acids, and 10% fetal bovine serum (pH 7.2). Phenylmethimazole (C10 [1, 2, 32, 33]) was prepared as 200 mM stock solution in DMSO and diluted in cell culture media.
Real-Time PCR
Total RNA was isolated (RNeasy Kit, Qiagen, Valencia, CA, USA) and treated with DNase (RNase-Free DNAse Kit, Qiagen). cDNA was synthesized using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Carlsbad, CA, USA). Pre-amplification of cDNA was done using the TaqMan PreAmp Master Mix Kit (Applied Biosystems, Carlsbad, CA, USA). Multiplex real-time PCR was performed using human TLR3 and Wnt5a Gene Expression Assays and GAPDH Gene Expression Assay was used as the internal control (Applied Biosystems). Non-preamplified cDNA was used for Wnt5a real-time PCR, whereas pre-amplified cDNA was used to amplify TLR3 by real-time PCR. Changes in gene expression were calculated using the ΔΔCt method.
ELISAs
Human recombinant IL-6 and pSTAT3Y705 ELISA kits were from Life Technologies (Grand Island, NY). For the measurement of IL-6, cells were treated for 24 hours with 500 μM C10 or DMSO (0.25%) for 24 hours. Supernatants were collected and IL-6 levels were evaluated using the human IL-6 ELISA kit as per the manufacturer’s instructions. For detection of pSTAT3Y705, cells were pre-treated with 500 μM C10 or DMSO (0.25%) for 1 hour and then stimulated with 100 ng/mL human recombinant IL-6 for ten minutes. Nuclear proteins were then collected using the NE-PER kit from ThermoFisher Scientific (Waltham, MA) as per the manufacturer’s instructions. Nuclear protein was then used to determine pSTAT3Y705 levels via the pSTAT3Y705 ELISA kit as per the manufacturer’s protocol.
Cell viability/growth
MCF-7 cells were evenly seeded and grown on sterile 96-well plates. Cells were then treated with C10 (0, 250, or 500 μM), DMSO (0.25%), or tamoxifen (6 μg/mL; Sigma Aldrich) alone or in combination for 72 hours. Cell viability/growth was then quantified using the MTT-based In Vitro Toxicology Assay Kit from Sigma-Aldrich (St. Louis, MO).
Cell migration
Scratch assays were performed as previously described [1, 2]. Briefly, MCF-7 cells were grown to confluency and a scratch was made in the confluent layer of cells using a sterile pipette tip. In some instances the cells were treated with C10 (500 μM) or 0.25% DMSO. Scratch assays were documented after 24 and 48 hrs using digital photography. ImageJ (NIH) was used to analyze 3 separate images of each treatment group to quantify cellular migration.
Statistics
All experiments were replicated at least three times on different groups of cells. All data are expressed as mean ± SD. Statistical significance was evaluated using a one-way ANOVA, and statistical significance for comparison of means between different groups was calculated using the Tukey-Kramer multiple comparison post hoc analysis using NCSS software. The differences were considered significant at p values < 0.05 as indicated in the figure legends.
ACKNOWLEDGMENTS
ALS designed the experiments, performed the experiments, analyzed and interpreted data, and wrote and edited the manuscript.
ED performed the experiments.
RM and ND designed the experiments, performed the experiments, analyzed and interpreted data.
KDM designed the experiments, performed the experiments, analyzed and interpreted data, and wrote and edited the manuscript.
CONFLICTS OF INTEREST
The authors declare no conflict of interest.
REFERENCES
1. McCall KD, Harii N, Lewis CJ, Malgor R, Kim WB, Saji M, Kohn AD, Moon RT, Kohn LD. High basal levels of functional toll-like receptor 3 (TLR3) and noncanonical Wnt5a are expressed in papillary thyroid cancer and are coordinately decreased by phenylmethimazole together with cell proliferation and migration. Endocrinology. 2007; 148:4226-4237.
2. Schwartz AL, Malgor R, Dickerson E, Weeraratna AT, Slominski A, Wortsman J, Harii N, Kohn AD, Moon RT, Schwartz FL, Goetz DJ, Kohn LD, McCall KD. Phenylmethimazole decreases Toll-like receptor 3 and noncanonical Wnt5a expression in pancreatic cancer and melanoma together with tumor cell growth and migration. Clin Cancer Res. 2009; 15:4114-4122.
3. Sato Y, Goto Y, Narita N, Hoon DS. Cancer Cells Expressing Toll-like Receptors and the Tumor Microenvironment. Cancer Microenviron. 2009; 2 Suppl 1:205-214.
4. Chen K, Huang J, Gong W, Iribarren P, Dunlop NM, Wang JM. Toll-like receptors in inflammation, infection and cancer. Int Immunopharmacol. 2007; 7:1271-1285.
5. Weeraratna AT, Jiang Y, Hostetter G, Rosenblatt K, Duray P, Bittner M, Trent JM. Wnt5a signaling directly affects cell motility and invasion of metastatic melanoma. Cancer Cell. 2002; 1:279-288.
6. Kurayoshi M, Oue N, Yamamoto H, Kishida M, Inoue A, Asahara T, Yasui W, Kikuchi A. Expression of Wnt-5a is correlated with aggressiveness of gastric cancer by stimulating cell migration and invasion. Cancer Res. 2006; 66:10439-10448.
7. McDonald SL, Silver A. The opposing roles of Wnt-5a in cancer. Br J Cancer. 2009; 101:209-214.
8. Iozzo RV, Eichstetter I, Danielson KG. Aberrant expression of the growth factor Wnt-5A in human malignancy. Cancer Res. 1995; 55:3495-3499.
9. Lejeune S, Huguet EL, Hamby A, Poulsom R, Harris AL. Wnt5a cloning, expression, and up-regulation in human primary breast cancers. Clin Cancer Res. 1995; 1:215-222.
10. Saitoh T, Mine T, Katoh M. Frequent up-regulation of WNT5A mRNA in primary gastric cancer. Int J Mol Med. 2002; 9:515-519.
11. Huang CL, Liu D, Nakano J, Ishikawa S, Kontani K, Yokomise H, Ueno M. Wnt5a expression is associated with the tumor proliferation and the stromal vascular endothelial growth factor--an expression in non-small-cell lung cancer. J Clin Oncol. 2005; 23:8765-8773.
12. Greten FR, Eckmann L, Greten TF, Park JM, Li ZW, Egan LJ, Kagnoff MF, Karin M. IKKbeta links inflammation and tumorigenesis in a mouse model of colitis-associated cancer. Cell. 2004; 118:285-296.
13. Bromberg JF, Wrzeszczynska MH, Devgan G, Zhao Y, Pestell RG, Albanese C, Darnell JE Jr. Stat3 as an oncogene. Cell. 1999; 98:295-303.
14. Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer. 2009; 9:798-809.
15. Yu H, Jove R. The STATs of cancer--new molecular targets come of age. Nat Rev Cancer. 2004; 4:97-105.
16. Lin L, Hutzen B, Zuo M, Ball S, Deangelis S, Foust E, Pandit B, Ihnat MA, Shenoy SS, Kulp S, Li PK, Li C, Fuchs J, Lin J. Novel STAT3 phosphorylation inhibitors exhibit potent growth-suppressive activity in pancreatic and breast cancer cells. Cancer Res. 70:2445-2454.
17. Grivennikov SI, Karin M. Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 21:11-19.
18. Azare J, Doane A, Leslie K, Chang Q, Berishaj M, Nnoli J, Mark K, Al-Ahmadie H, Gerald W, Hassimi M, Viale A, Stracke M, Lyden D, Bromberg J. Stat3 mediates expression of autotaxin in breast cancer. PLoS One. 2011; 6:e27851.
19. Katoh M. STAT3-induced WNT5A signaling loop in embryonic stem cells, adult normal tissues, chronic persistent inflammation, rheumatoid arthritis and cancer (Review). Int J Mol Med. 2007; 19:273-278.
20. McCall KD, Harii N, Lewis CJ, Malgor R, Kim WB, Saji M, Kohn AD, Moon RT, Kohn LD. High Basal Levels of Functional Toll-Like Receptor 3 (TLR3) and Non-Cannonical Wnt5a Are Expressed in Papillary Thyroid Cancer (PTC) and Are Coordinately Decreased by Phenylmethimazole Together with Cell Proliferation and Migration. Endocrinology. 2007; 148:4226-4237.
21. Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol. 2003; 21:335-376.
22. Lu H, Ouyang W, Huang C. Inflammation, a key event in cancer development. Mol Cancer Res. 2006; 4:221-233.
23. Garcia M, Fernandez-Garcia NI, Rivas V, Carretero M, Escamez MJ, Gonzalez-Martin A, Medrano EE, Volpert O, Jorcano JL, Jimenez B, Larcher F, Del Rio M. Inhibition of xenografted human melanoma growth and prevention of metastasis development by dual antiangiogenic/antitumor activities of pigment epithelium-derived factor. Cancer Res. 2004; 64:5632-5642.
24. Calo V, Migliavacca M, Bazan V, Macaluso M, Buscemi M, Gebbia N, Russo A. STAT proteins: from normal control of cellular events to tumorigenesis. J Cell Physiol. 2003; 197:157-168.
25. Wen Z, Zhong Z, Darnell JE Jr. Maximal activation of transcription by Stat1 and Stat3 requires both tyrosine and serine phosphorylation. Cell. 1995; 82:241-250.
26. Gartsbein M, Alt A, Hashimoto K, Nakajima K, Kuroki T, Tennenbaum T. The role of protein kinase C {delta} activation and STAT3 Ser727 phosphorylation in insulin-induced keratinocyte proliferation. J Cell Sci. 2006; 119:470-481.
27. Barboza JA, Wang S, Schaefer TS. Generation and characterization of a constitutively active Stat3 protein. Mol Biol Rep. 2004; 31:13-21.
28. Yeh YT, Ou-Yang F, Chen IF, Yang SF, Wang YY, Chuang HY, Su JH, Hou MF, Yuan SS. STAT3 ser727 phosphorylation and its association with negative estrogen receptor status in breast infiltrating ductal carcinoma. Int J Cancer. 2006; 118:2943-2947.
29. Gakhar G, Hua DH, Nguyen TA. Combinational treatment of gap junctional activator and tamoxifen in breast cancer cells. Anticancer Drugs. 2010; 21:77-88.
30. Bhattacharya D, Yusuf N. Expression of toll-like receptors on breast tumors: taking a toll on tumor microenvironment. Int J Breast Cancer. 2012; 2012:716564.
31. MacMillan CD, Leong HS, Dales DW, Robertson AE, Lewis JD, Chambers AF, Tuck AB. Stage of breast cancer progression influences cellular response to activation of the WNT/planar cell polarity pathway. Sci Rep. 2014; 4:6315.
32. Benavides U, Gonzalez-Murguiondo M, Harii N, Lewis CJ, Schwartz AL, Giuliani C, Napolitano G, Dagia NM, Malgor R, McCall KD, Kohn LD. Phenylmethimazole inhibits production of proinflammatory mediators and is protective in an experimental model of endotoxic shock*. Crit Care Med. 2012; 40:886-894.
33. McCall KD, Holliday D, Dickerson E, Wallace B, Schwartz AL, Schwartz C, Lewis CJ, Kohn LD, Schwartz FL. Phenylmethimazole blocks palmitate-mediated induction of inflammatory cytokine pathways in 3T3L1 adipocytes and RAW 264.7 macrophages. J Endocrinol. 2010; 207:343-353.
34. Salaun B, Zitvogel L, Asselin-Paturel C, Morel Y, Chemin K, Dubois C, Massacrier C, Conforti R, Chenard MP, Sabourin JC, Goubar A, Lebecque S, Pierres M, Rimoldi D, Romero P, Andre F. TLR3 as a biomarker for the therapeutic efficacy of double-stranded RNA in breast cancer. Cancer Res. 2011; 71:1607-1614.