
INTRODUCTION
Neuroblastoma (NB), a high-grade malignancy with poor prognosis, originates from the primitive neural crest and is the most common extra-cranial solid tumor in children. More than 70% of NB patients have metastatic disease at diagnosis, which is classified as high- risk NB. Though current multidisciplinary treatment is available, the 5-year progression-free survival of high-risk NB patients is only 36%–56% [1–2]. Therefore, new therapies are needed to improve treatment outcomes in NB patients. The early metastasis suggests that NB cells are highly competent. Gaining a thorough understanding of the regulatory mechanism underlying NB migration and invasion would help identify new therapeutic targets.
Classic human leukocyte antigen-I (HLA-Ia) molecules are reduced or absent on the surface of tumor cells, whereas non-classic HLA (HLA-Ib) molecules, such as HLA-G and HLA-E, are highly expressed on the surface of tumor cells. HLA-E is the main ligand for the inhibitory receptor CD94/NKG2A on the surface of natural killer (NK) and cytotoxic T lymphocyte (CTL) cells. In the major mechanism underlying tumor immune escape, HLA-E binds to the leading peptides of HLA-A, -B, -C, and -G, activating the CD94/NKG2A receptor and delivering an inhibitory signal to NK and CTL cells [3]. Studies have found that HLA-E is highly expressed in a variety of tumors [4] and closely associated with certain biological characteristics of tumors [5]. However, HLA-E expression in NB tissue and its clinical significance is yet to be determined.
In addition to immune escape, HLA-E may promote tumor growth via other mechanisms related to tumor stem cell differentiation [6], tumor-associated macrophage activity [7], and functional activation of the vascular endothelium [8]. In a cohort of laryngeal carcinoma patients, HLA-E may be a biomarker of tumor invasiveness [9], but the definitive mechanism is unknown. Hepatic cells infected with hepatitis virus have been shown to express a high level of HLA-E, which inhibits NK-cell cytotoxicity in abnormal hepatic cells and stimulates NK cells to release cytokines, including interleukin (IL)-10 and transforming growth factor (TGF)-β1 [10]. In breast cancer and lung adenocarcinoma, IL-10 and TGF-β1 have been confirmed to promote tumor growth through pathways including tumor migration and invasion [11–12]. NB is an aggressive tumor. Therefore, we hypothesize that HLA-E is highly expressed in NB tissue, inhibits NK cells, and promotes the release of IL-10 and TGF-β1 to affect tumor growth.
RESULTS
Expression of HLA-E in NB was correlated to disease stage and N-MYC gene status
Eighty-four cases of NB were assessed for HLA-E expression. Overall, 70.2% (59/84) of the tumors stained positive for HLA-E (Figure 1A). HLA-E expression was significantly associated with disease stage. The percentage of HLA-E-positive tumors in patients with stage II, III, and IV disease was 16.7%, 44.4%, and 80.0%, respectively (P < 0.01). Thirty-two of the children underwent N-MYC oncogene testing; 10 were positive for N-MYC amplification. The HLA-E expression rates were 90.0% and 50.0% respectively in patients with or without N-MYC gene amplification (P < 0.05).
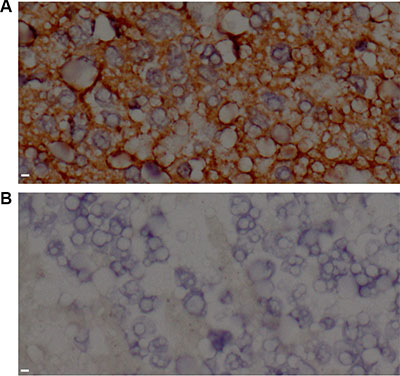
Figure 1: HLA-E expression in tissue from neuroblastoma patients. (A) Tumor tissue stained positive for HLA-E in a patient with stage IV disease. (B) Tumor tissue stained negative for HLA-E in a patient with stage II disease. All images were taken at 400 × magnification. Scale bar is 10 μm.
HLA-E expressed in NB cells inhibited NK-cell cytotoxicity and stimulated cytokine release
Cells from the bone marrow of one patient with high HLA-E expression in the tumor tissue and confirmed metastatic disease in the bone marrow were cultured to produce a cell line named NB-Ehigh in this study. NB-Ehigh cells stably expressed high levels of HLA-E (Figure 2). HLA-E expression in NB-Ehigh cells was down-regulated by siRNA transfection, creating NB-Elow cells. The blank control, non-specific control, and NB-Ehigh groups expressed HLA-E at high levels at 48 hours, whereas most NB-Elow cells expressed little or no HLA-E expression. Western blot analyses showed that NB-Elow cells expressed significantly less HLA-E protein than NB-Ehigh and control cells (Figure 2).
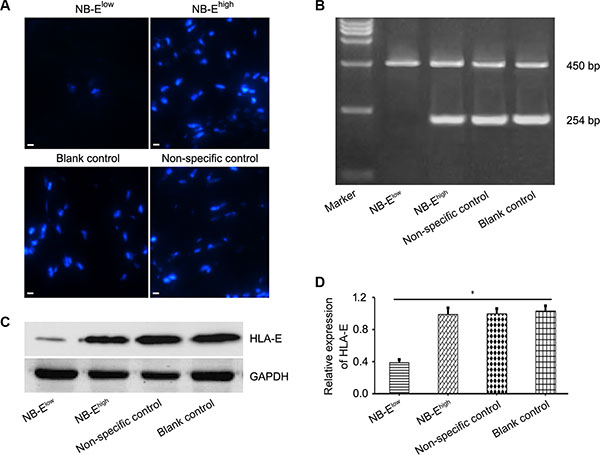
Figure 2: HLA-E expression in neuroblastoma cells. (A) Immunofluorescence microscopy of NB cells with HLA-E expression in the NB-Elow, NB-Ehigh, and control groups after 48 h of siRNA transfection (× 200). (B–C) Real-time quantitative PCR and Western blot confirmed that HLA-E was down-regulated in NB-Elow cells. (D) Relative expression of HLA-E, expressed as IODHLA-E/IODGAPDH, was lower in the NB-Elow group than in the NB-Ehigh and control groups. GADPH: endogenous control; IOD: average optical density. The data are representative of five independent experiments. Scale bar is 10 μm. *P < 0.01, two-tailed t test.
NK-cell cytotoxicity was significantly enhanced in the NB-Elow group compared to the NB-Ehigh group (Figure 3A). Down-regulation of HLA-E expression in NB cells enhanced NK-cell cytotoxicity toward NB cells. Supernatant was collected after NB cells were co-cultured with NK cells. Sandwich ELISA was used to measure IL-10 and TGF-β in the supernatant. IL-10 and TGF-β levels were higher in the NB-Ehigh group than in the NB-Elow group (Figure 3B).
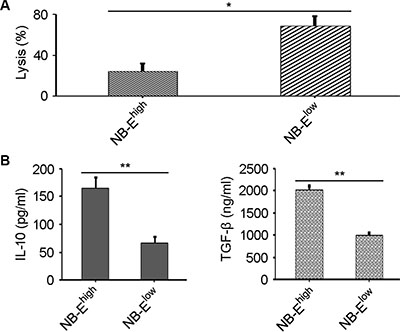
Figure 3: Effect of HLA-E expression on NK-cell cytotoxicity and cytokine release. (A) Lysis percentage of target cells in the NB-Elow and NB-Ehigh groups. The data are representative of five independent experiments. *P < 0.01, two-tailed t test. (B) Higher IL-10 and TGF-β levels are secreted by NK cells when inhibited by tumor cells in the NB-Ehigh group compared to the NB-Elow group. The data are representative of five independent experiments. **P < 0.001, two-tailed t test.
HLA-E activated the migration and invasion of NB cells in vitro
To study the impact of HLA-E expression on the migration and invasion of NB cells in vitro, the scratch-wound assay and Transwell invasion assay were adopted. In the scratch-wound assay, the width of the scratch was smaller in the NB-Ehigh group than in the NB-Elow group (Figure 4A–4B). The Transwell invasion assay showed more film-crossing cells in the NB-Ehigh group than in the NB-Elow group (Figure 4C–4D). When NB-Ehigh cells and NB-Elow cells were cultured with NB/NK supernatant, the rate of migration and invasion in the NB-Ehigh group increased, but no significant change was observed in the NB-Elow group (Figure 4). When anti-IL-10 and anti-TGF-β mAbs were added to the NB-Ehigh cells cultured with NB/NK supernatant, the width of the scratch and the number of film-crossing cells were similar to those of the NB-Ehigh group without treatment (Figure 4).
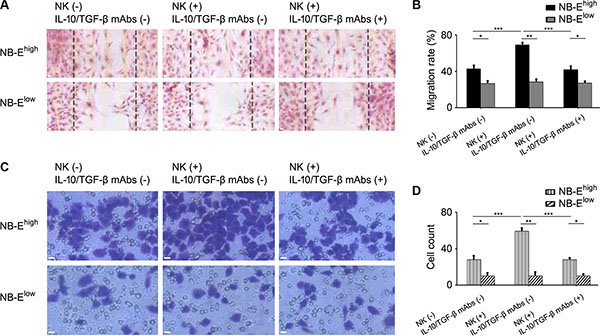
Figure 4: Effect of HLA-E expression on the migration and invasion of NB cells. (A) Representative migration of NB cells with or without NK/NB supernatant and with or without anti-IL-10/TGF-β mAbs. (B) Migration rates of NB cells in the NB-Elow and NB-Ehigh groups with or without NK/NB supernatant and with or without anti-IL-10/TGF-β mAbs. The data are representative of five independent experiments. *P < 0.05, ** P < 0.001, *** P < 0.01. (C) Representative invasion of NB cells with or without NK/NB supernatant and with or without anti-IL-10/TGF-β mAbs. (D) Invasion rates of NB cells in the NB-Elow and NB-Ehigh groups with or without NK/NB supernatant and with or without anti-IL-10/TGF-β mAbs. The data are representative of five independent experiments. Scale bar is 10 μm. *P < 0.05, ** P < 0.001, *** P < 0.01.
HLA-E promoted NB growth and stimulated cytokine release in vivo
To determine the role of HLA-E expressed in NB cells in tumor growth and cytokine release in vivo, a tumor model was establish in nude mice. On day 30 of the growth of the grafted tumor, the NB-Ehigh group had a heavier tumor weight (Figure 5A–5B) and larger tumor volume (Figure 5C–5D) than the NB-Elow group. The number of metastatic cells in the bone marrow was greater in the NB-Ehigh group than in the NB-Elow group (Figure 6). The expression levels of IL-10 and TGF-β in plasma and tumor tissue were higher in the NB-Ehigh group than in the NB-Elow group (Table 1).
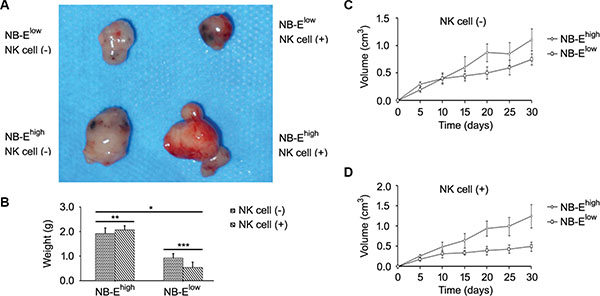
Figure 5: Effect of HLA-E expression on tumor growth. (A) Representative sizes of the neuroblastoma graft in nude mice. (B–D) Tumor weight and tumor volume in the NB-Ehigh group and NB-Elow group. NK cell infusion decreased tumor weight and volume in the NB-Elow group but had no effect on tumor weight and volume in the NB-Ehigh group. *P < 0.01, **P > 0.05, ***P < 0.05, two-tailed t test.
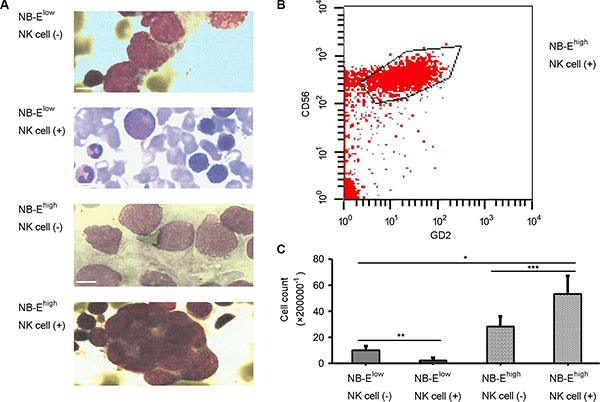
Figure 6: Metastatic neuroblastoma in bone marrow. Morphology (A) and flow cytology (B–C) examination confirmed that the number of metastatic cells in the bone marrow was greater in the NB-Ehigh group than in the NB-Elow group (P < 0.01). NK cell infusion decreased the number of metastatic cells in the bone marrow of mice grafted with NB-Elow cells, but increased the number of metastatic cells in the bone marrow of mice grafted with NB-Ehigh cells (P < 0.05). Scale bar is 10 μm.
Table 1: HLA-E-induced expression of IL-10 and TGF-β in vivo
Treatment |
Group |
Peripheral blood |
Tumor tissue |
||
|---|---|---|---|---|---|
IL-10 (pg/ml) |
TGF-β (ng/ml) |
IL-10 |
TGF-β |
||
No NK infusion |
NB-Ehigh* |
93.64 ± 15.06 |
18.9 ± 5.1 |
++ |
++ |
NB-Elow |
56.07 ± 14.58 |
12.5 ± 3.5 |
+ |
+ |
|
NK infusion |
NB-Ehigh** |
240.42 ± 23.71 |
49.3 ± 5.9 |
+++ |
+++ |
NB-Elow*** |
63.84 ± 17.13 |
14.9 ± 4.1 |
+ |
+ |
|
*all P < 0.05 vs. NB-Elow, ** all P < 0.001 vs. NB-Ehigh without NK infusion, *** all P > 0.05 vs. NB-Elow without NK infusion. + indicates < 10% of positive cells; ++ indicates 10–50% of positive cells; and +++ indicates >50% of positive cells.
NK cell infusion could not inhibit tumor growth in NB with high HLA-E expression
The effect of NK cell infusion on tumor growth in NB was tested in vivo. NK cell infusion significantly increased the expression of IL-10 and TGF-β in plasma and tumor tissue in the NB-Ehigh group, but not in the NB-Elow group (Table 1). In mice grafted with NB-Elow cells, NK cell infusion decreased the tumor volume, tumor weight and number of metastatic cells in the bone marrow (Figure 5 and Figure 6). However, in mice grafted with NB-Ehigh cells, NK cell infusion had no effect on the local tumor weight (Figure 5A–5B) and tumor volume (Figure 5C–5D), though the number of metastatic cells in the bone marrow was increased (Figure 6).
DISCUSSION
HLA-E expression is elevated in a number of tumors. Preclinical studies have suggested that tumor cells with high HLA-E expression inhibit NK-cell or CTL cytotoxicity, but blocking the binding between HLA-E and CD94/NKG2A receptor restores NK-cell and CTL cytotoxicity [13]. Similarly, clinical studies have shown that patients with colorectal cancer [14], breast cancer [15], or glioblastoma [16] and high HLA-E expression in the tumor tissue have poorer prognoses. However, contradictory evidence indicates that patients who have cervical [17] or renal carcinoma [18] and high HLA-E expression have good prognoses. Even in the same cancer type, such as colorectal cancer, there are contradictory opinions [19, 14]. Therefore, the effect of HLA-E on tumor prognosis is controversial.
A recent study showed that NB patients with high serum soluble HLA-E (sHLA-E) have a good prognosis. However, that study did not determine whether sHLA-E is released by tumor cells or the immune cells involved in anti-tumor immunity [20]. Our study seemed to have the opposite result. We demonstrated that HLA-E was expressed in the tumor tissues of 70.2% of the NB patients. This high expression of HLA-E was associated with tumor stage and N-myc gene status, which definitively correlated with prognosis. This finding suggested a role of HLA-E in NB growth.
To determine the effect of HLA-E on the cytolytic activity of NK cells, the interaction between NK cells and NB cells with high or low HLA-E expression was observed. siRNA was used to down-regulate HLA-E expression in the NB-Ehigh cells obtained by primary culture to produce the NB-Elow cells as confirmed by immunofluorescence assay, Western blot, and quantitative PCR. The cytotoxicity assay showed that NB-Ehigh cells significantly inhibited the cytotoxicity of NK cells towards tumor cells in vitro. When HLA-E expression was down-regulated, the cytotoxicity of NK cells was restored as reported in a previous study [3].
HLA-E has been shown to be highly expressed in hepatic cells infected with the hepatitis C virus, inhibit the activity of NK cells, and stimulate NK cells to secrete massive amounts of IL-10 and TGF-β by triggering the NKG2A receptor, modifying the functions of DC cells and CD4+ T cells [10]. Similarly, in this study, high IL-10 and TGF-β levels were detected in the supernatant from NK cells co-cultured with NB-Ehigh cells, but not NB-Elow cells. This indicates that the release of these cytokines correlated with high HLA-E expression. Recently, TGF-β was found to be related to a pathway important for promoting NB invasion [21] and may be a potential treatment target in NB [22]. IL-10 has also been shown to promote tumor growth via various pathways [23, 12].
Both the scratch-wound assay and Transwell invasion assay showed that NB-Ehigh cells were more capable of migration and invasion than NB-Elow cells. The supernatant taken after the co-culture of NB-Ehigh cells and NK cells enhanced the ability of NB-Ehigh cells to migrate and invade tissue. This effect was reversed by the anti-IL-10/TGF-β mAbs, which confirmed that the HLA-E-mediated release of IL-10 and TGF-β promotes tumor infiltration.
In the xenograft experiment in mice, tumors with high HLA-E expression grew rapidly and expressed high levels of IL-10 and TGF-β in plasma and tumor tissue and were associated with metastatic disease in bone marrow. The underlying mechanism of HLA-E-mediated promotion of tumor progression was studied recently at the molecular level [24]. Moreover, we observed that NK cell infusion had an inhibitory effect on NB-Elow tumor growth in mice but no effect on local NB-Ehigh tumor growth. Increased IL-10 and TGF-β levels were detected in plasma and tumor tissue from NB-Ehigh mice, as well as the increased metastatic diseases in the bone marrow. Previous studies had proved tumor cell could avoid NK cell attack by direct and indirect mechanismins. Direct mechanisms consisted of shedding soluble ligands for NK cell-activating receptors, upregulation of HLA molecules and release of inhibitory cytokines. Indirect mechanisms consisted of activation of inhibitory regulatory T cells, DC killing, and phagocyte-derived inhibitory cytokines [25]. We speculated HLA-E induced expression of IL-10 and TGF-β by NK cells via these indirect mechanisms in the tumor-NK cell interaction in vivo.
Some preclinical studies have shown that NK cell infusion has good efficacy as an immunotherapy for NB [26]. However, NK cells express a complicated range of receptors, such as inhibitory and activating receptors, and their binding to different ligands may affect the efficacy of immunotherapy in NB [27]. Due to the high-affinity of HLA-E for inhibitory CD94/NKG2A receptor and a negative regulatory feedback mechanism in the expression of inhibitory CD94/NKG2A and activating CD94/NKG2C receptors, HLA-E definitively inhibited NK-cell activity [28]. Thus the effect of tumor HLA-E expression on efficacy must be considered when using NK cells as immunotherapy in NB. The present study proved that NK cell infusion does not inhibit the growth of grafted NB with high HLA-E expression in nude mice.
This study did not investigate another HLA-Ib molecule, HLA-G, which is also highly expressed in a number of tumors, including NB [29]. HLA-G also inhibits NK-cell activity and assists tumors in immune escape. However, HLA-G cannot bind directly to CD94/NKG2A; it must first form a complex with HLA-E via its leading peptide and then activates the CD94/NKG2A receptor to deliver an inhibitory signal to NK cells [30]. A recent study using comparative proteomics analyzed 363 proteins expressed in glioma cells and confirmed that the inhibition of NK-cell cytotoxicity is mediated by the HLA-E expressed in tumor cells [31]. Therefore, HLA-G would not have affected the results of the present study.
In conclusion, our study found that high HLA-E expression in NB inhibits NK-cell cytolysis both in vitro and in vivo. High HLA-E expression also induces NK cells to secrete IL-10 and TGF-β, which affects the promotion of tumor migration and invasion. Because the down-regulation of HLA-E may inhibit tumor growth, HLA-E may be a new treatment target in NB and is worthy of further investigation.
MATERIALS AND METHODS
Patients and specimens
All human samples were donated freely, and informed consent was obtained from all children/guardians. Ethical approval was obtained from the Institutional Review Board of Sun Yat-Sen University, Guangzhou, China. Tumor tissues were collected from 84 children (55 boys, 29 girls; median age 4 years, range 2–10 years) with newly diagnosed NB at Sun Yat-Sen University Cancer Center between June 2004 and June 2014. All tissues were obtained after successful clinical diagnosis. The primary tumor sites were the retroperitoneal region in 52 cases, mediastinum in 18 cases, and others in 14 cases. According to the international NB staging system, 6 cases were stage II, 28 cases stage III, and 50 stage IV disease. Thirty-two of the children underwent N-MYC oncogene testing; 10 were positive for N-MYC amplification. Other risk factors, including international NB pathological classification and DNA ploidy, were unavailable at our center and not analyzed in this retrospective cohort. At the time of tissue collection, all patients with stage III and stage IV disease had received reduction chemotherapy, whereas patients with stage II disease were chemotherapy-naive. Due to the long recruitment period and non-uniformity in the treatment protocol, the relationship between HLA-E expression and treatment outcome was not analyzed.
Immunohistochemical analysis
Conventional formalin-fixed paraffin-embedded tumor tissue was cut into 4-μm sections and stained using the immunohistochemical streptavidin-peroxidase (SP) method [32]. The anti-HLA-E monoclonal antibody 3H2679 (Santa Cruz, USA) was used. HLA-E expression was evaluated as described previously [5].
Primary culture
Cells from the bone marrow of one patient with high HLA-E expression in the tumor tissue and confirmed metastatic disease in the bone marrow were obtained at diagnosis. The cells were suspended in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal calf serum, 1% non-essential amino acids, and 1% penicillin/streptomycin and distributed into 25 cm3 cell culture flasks. Primary cultures were incubated at 37°C with 5% CO2 in air. After collecting the tumor cells, high HLA-E expression on the cell surface was confirmed by Western blotting and polymerase chain reaction (PCR). These cells were referred to as NB-Ehigh in this study.
Cellular siRNA transfection
A siRNA sequence was designed based on the mRNA sequence of HLA-E (NM_005516) [33]. The target region was 525-547: ATCTCCGAGCAAAAGTCAAATGA. The sense primer was 5′-CUCCGAGCAAAAGUCA AAUGA-3′ and the anti-sense primer 5′-AUUUGACUUU UGCUCGGAGAU-3′. NB-Ehigh cells were seeded in 24-well plates (5 × 104 cells/well) and cultured for 18 h before transfection. Transfection was performed using 50 pmol siRNA and 2.5 μl Lipofectamine 2000 at a total volume of 0.5 ml using a previously reported method [34]. The goal was to obtain a cell line with low HLA-E expression, referred to as NB-Elow. PBS and non-specific siRNA were used to establish a blank control and non-specific control, respectively. The effect of silencing the HLA-E gene was detected by immunofluorescence, Western blotting, and real-time quantitative PCR as reported previously [35–37].
NK-cell cytotoxicity
A 2 ml venous blood sample was taken from healthy individuals, mixed with lymphocyte separation medium, and centrifuged (2000 r·min−1 × 20 min) to obtain peripheral mononuclear cells. The magnetic bead system coated with anti-CD56 monoclonal antibody was used to select NK cells as effector cells. NB cells (i.e., target cells) were added to 96-well plates at a density of 2.5 × 104 cells/well and cultured for 14 hours. The cultured NB cells were combined with the effector cells at an effector/target ratio of 100:1 and cultured for 20 hours. The activity of adherent target cells in the wells was measured using the methyl thiazolyl tetrazolium (MTT) method. The rate of NK-cell cytotoxicity was calculated as reported previously [38].
Enzyme-linked immunosorbent assay (ELISA)
NB cells at a density of 1 × 106 cells/well were cultured with NK cells at an effector/target ratio of 100:1. The supernatant was collected and tested for IL-10 and TGF-β using an ELISA kit (Phoenix Pharmaceuticals Inc., US). The supernatant from NK cells cultured with NB-Ehigh and NB-Elow cells was termed NB/NK supernatant 1 and NB/NK supernatant 2, respectively, and stored for further usage in scratch-wound assay and Transwell invasion assay.
Scratch-wound assay
The migration capability of NB cells with different HLA-E expression was determined by the scratch-wound assay as previously described [39]. To investigate the impact of IL-10 and TGF-β secreted by NK cells on NB cell migration, NB-Ehigh and NB-Elow cells were cultured with NB/NK supernatant 1 and NB/NK supernatant 2, respectively, and the scratch-wound assay repeated. Anti-IL-10 and anti-TGF-β monoclonal antibodies (mAbs) (Santa Cruz, USA) were added (5 μg/ml each) to the NB cells cultured with NB/NK supernatant to determine whether the promotion of NB cell migration by NB/NK supernatant was impaired.
Transwell invasion assay
The invation capability of NB cells with different HLA-E expression was determined by the Transwell invasion assay as previously described [40]. To investigate the impact of IL-10 and TGF-β secreted by NK cells on NB cell invasion, NB-Ehigh and NB-Elow cells were cultured with NB/NK supernatant 1 and NB/NK supernatant 2, respectively, and the scratch-wound assay repeated. Furthermore, anti-IL-10 and anti-TGF-β mAbs (5 μg/ml each) were added to the NB cells cultured with NB/NK supernatant to determine whether the promotion of NB cell invasion by NB/NK supernatant was impaired.
Animal model
NB cells were inoculated in the right armpits of Balb/c nu/nu nude mice (Laboratory Animal Center, Southern Medical University, Guangzhou, China) at a density of 1 × 106 cells/200 μl to establish the NB model. On day 30 after inoculation, the mice were sacrificed and the tumor tissue obtained and weighed. ELISA was used to measure IL-10 and TGF-β in the peripheral serum from the nude mice. Tumor tissue was obtained and semiquantitative immunohistochemistry performed to measure the expression of IL-10 and TGF-β as described previously [41]. In tumor tissue, reactions were grouped by intensity, quantity, and type of staining (intensity: absent, weak, medium, strong; quantity: no positive tumor cells, < 10% of cells, 10–50% of cells, > 50% of cells).
NK cell infusion
The NB xenografts in nude mice were established and grouped as described previously. NK cells from healthy volunteers were stimulated overnight with a combination of 10 ng/ml IL-15 and 40 IU/ml IL-2. A total of 5 ×105 activated NK cells were infused in each of the mice via the caudal vein weekly over a total of 4 weeks. On day 30 after inoculation, the mice were sacrificed and the tumor tissue obtained and weighed. ELISA was used to measure the peripheral serum levels of IL-10 and TGF-β in the nude mice. Immunohistochemistry was used to determine the expression profile of IL-10 and TGF-β in the grafted tumor tissue.
Bone marrow cytology and flow cytology
To quantitatively evaluate the NB metastatic disease in nude mice on day 30, bone marrow, which is one of the most commonly involved sites in NB, was taken from the left femur for examination by morphology and flow cytology. FCM (FACScaliburTM, Becton-Dickinson, US) analysis was performed, as previously described [42], on the bone marrow specimens using the antibodies of anti-human ganglioside D2 (GD2) monoclonal antibody, phycoerythrin (PE) labeled anti-neural cell adhesion molecule monoclonal antibody (CD56) and peridinin chlorophyll protein (PerCP) labeled CD45 monoclonal antibody.
ACKNOWLEDGMENTS AND FUNDING
This work was supported by a grant from the Science and Technology Planning Project of Guangdong Province, China (2013B021800069 and 2012B031800460).
CONFLICTS OF INTEREST
No potential conflicts of interest were disclosed.
REFERENCES
1. Cheung NK, Cheung IY, Kushner BH, Ostrovnaya I, Chamberlain E, Kramer K, Modak S. Murine anti-GD2 monoclonal antibody 3F8 combined with granulocyte-macrophage colony-stimulating factor and 13-cis-retinoic acid in high-risk patients with stage 4 neuroblastoma in first remission. J Clin Oncol. 2012; 30:3264–70.
2. Kreissman SG, Seeger RC, Matthay KK, London WB, Sposto R, Grupp SA, Haas-Kogan DA, Laquaglia MP, Yu AL, Diller L, Buxton A, Park JR, Cohn SL, et al. Purged versus non-purged peripheral blood stem-cell transplantation for high-risk neuroblastoma (COG A3973): a randomised phase 3 trial. Lancet Oncol. 2013; 14:999–1008.
3. Petrie EJ, Clements CS, Lin J, Sullivan LC, Johnson D, Huyton T, Heroux A, Hoare HL, Beddoe T, Reid HH, Wilce MC, Brooks AG, Rossjohn J. CD94-NKG2A recognition of human leukocyte antigen (HLA)-E bound to an HLA class I leader sequence. J Exp Med. 2008; 205:725–35.
4. Palmisano GL, Contardi E, Morabito A, Gargaglione V, Ferrara GB, Pistillo MP. HLA-E surface expression is independent of the availability of HLA class I signal sequence-derived peptides in human tumor cell lines. Hum Immunol. 2005; 66:1–12.
5. Zhen ZJ, Ling JY, Cai Y, Luo WB, He YJ. Impact of HLA-E gene polymorphism on HLA-E expression in tumor cells and prognosis in patients with stage III colorectal cancer. Med Oncol. 2013; 30:482–490.
6. Wolpert F, Roth P, Lamszus K, Tabatabai G, Weller M, Eisele G. HLA-E contributes to an immune-inhibitory phenotype of glioblastoma stem-like cells. Journal of Neuroimmunology. 2012; 250:27–34.
7. Kren L, Muckova K, Lzicarova E, Sova M, Vybihal V, Svoboda T, Fadrus P, Smrcka M, Slaby O, Lakomy R, Vanhara P, Krenova Z, Michalek J. Production of immune-modulatory nonclassical molecules HLA-G, HLA-E by tumor infiltrating ameboid microglia/macrophages in glioblastomas: a role in innate immunity? Journal of Neuroimmunology. 2010; 220:131–35.
8. Coupel S, Moreau A, Hamidou M, Horejsi V, Soulillou JP, Charreau B. Expression and release of soluble HLA-E is an immunoregulatory feature of endothelial cell activation. Blood. 2007; 109:2806–14.
9. Silva TG, Crispim JC, Miranda FA, Hassumi MK, de Mello JM, Simões RT, Souto F, Soares EG, Donadi EA, Soares CP. Expression of the nonclassical HLA-G and HLA-E molecules in laryngeal lesions as biomarkers of tumorinvasiveness. Histology and Histopathology. 2011; 26:1487–97.
10. Jinushi M, Takehara T, Tatsumi T, Kanto T, Miyagi T, Suzuki T, Kanazawa Y, Hiramatsu N, Hayashi N. Negative regulation of NK cell activities by inhibitory receptor CD94/NKG2A leads to altered NK cell-induced modulation of dendritic cell functions in chronic hepatitis C virus infection. J Immunol. 2004; 173:6072–81.
11. Singh R, Shankar BS, Sainis KB. TGF-β1-ROS-ATM-CREB signaling axis in macrophage mediated migration of human breast cancer MCF7 cells. Cell Signal. 2014; 26:1604–15.
12. Sung WW, Wang YC, Lin PL, Cheng YW, Chen CY, Wu TC, Lee H. IL-10 promotes tumor aggressiveness via upregulation of CIP2A transcription in lung adenocarcinoma. Clin Cancer Res. 2013; 19:4092–103.
13. Wischhusen J, Friese MA, Mittelbronn M, Meyermann R, Weller M. HLA-E protects glioma cells from NKG2D-mediated immune responses in vitro: implications for immune escape in vivo. Journal of Neuropathology and Experimental Neurology. 2005; 64:523–28.
14. Bossard C, Bézieau S, Matysiak-Budnik T, Volteau C, Laboisse CL, Jotereau F, Mosnier JF. HLA-E/β2 microglobulin overexpression in colorectal cancer is associated with recruitment of inhibitory immune cells and tumor progression. International Journal of Cancer. 2012; 131:855–63.
15. de Kruijf EM, Sajet A, van Nes JG, Natanov R, Putter H, Smit VT, Liefers GJ, van den Elsen PJ, van de Velde CJ, Kuppen PJ. HLA-E and HLA-G expression in classical HLA class I-negative tumors is of prognostic value for clinical outcome of early breast cancer patients. J Immunol. 2010; 185:7452–59.
16. Kren L, Slaby O, Muckova K, Lzicarova E, Sova M, Vybihal V, Svoboda T, Fadrus P, Lakomy R, Vanhara P, Krenova Z, Sterba J, Smrcka M, et al. Expression of immune-modulatory molecules HLA-G and HLA-E by tumor cells in glioblastomas: an unexpected prognostic significance? Neuropathology. 2011; 31:129–34.
17. Spaans VM, Peters AA, Fleuren GJ, Jordanova ES. HLA-E expression in cervical adenocarcinomas: association with improved long-term survival. J Transl Med. 2012; 10:184–94.
18. Kren L, Valkovsky I, Dolezel J, Capak I, Pacik D, Poprach A, Lakomy R, Redova M, Fabian P, Krenova Z, Slaby O. HLA-G and HLA-E specific mRNAs connote opposite prognostic significance in renal cell carcinoma. Diagn Pathol. 2012; 7:58–64.
19. Benevolo M, Mottolese M, Tremante E, Rollo F, Diodoro MG, Ercolani C, Sperduti I, Lo Monaco E, Cosimelli M, Giacomini P. High expression of HLA-E in colorectal carcinoma is associated with a favorable prognosis. J Transl Med. 2011; 9:184–93.
20. Morandi F, Cangemi G, Barco S, Amoroso L, Giuliano M, Gigliotti AR, Pistoia V, Corrias MV. Plasma levels of soluble HLA-E and HLA-F at diagnosis may predict overall survival of neuroblastoma patients. BioMed Res Int. 2013; doi: 10.1155/2013/956878.
21. Lynch J, Fay J, Meehan M, Bryan K, Watters KM, Murphy DM, Stallings RL. MiRNA-335 suppresses neuroblastoma cell invasiveness by direct targeting of multiple genes from the non-canonical TGF-β signalling pathway. Carcinogenesis. 2012; 33:976–85.
22. Xu Y, Sun J, Sheard MA, Tran HC, Wan Z, Liu WY, Asgharzadeh S, Sposto R, Wu HW, Seeger RC. Lenalidomide overcomes suppression of human natural killer cell anti-tumor functions by neuroblastoma microenvironment-associated IL-6 and TGFβ1. Cancer Immunology Immunotherapy. 2013; 62:1637–48.
23. Gupta M, Han JJ, Stenson M, Maurer M, Wellik L, Hu G, Ziesmer S, Dogan A, Witzig TE. Elevated serum IL-10 levels in diffuse large B-cell lymphoma: a mechanism of aberrant JAK2 activation. Blood. 2012; 119:2844–53.
24. Sasaki T, Ravindranath MH, Terasaki PI, Freitas MC, Kawakita S, Jucaud V. Gastric cancer progression may involve a shift in HLA-E profile from an intact heterodimer to β2-microglobulin-free monomer. International Journal of Cancer. 2014; 134:1558–70.
25. Chester C, Fritsch K, Kohrt HE. Natural Killer Cell Immunomodulation: Targeting Activating, Inhibitory, and Co-stimulatory Receptor Signaling for Cancer Immunotherapy. Front Immunol. 2015; 6:601–9.
26. Rujkijyanont P, Chan WK, Eldridge PW, Lockey T, Holladay M, Rooney B, Davidoff AM, Leung W, Vong Q. Ex vivo activation of CD56(+) immune cells that eradicate neuroblastoma. Cancer Res. 2013; 73:2608–18.
27. Delgado DC, Hank JA, Kolesar J, Lorentzen D, Gan J, Seo S, Kim K, Shusterman S, Gillies SD, Reisfeld RA, Yang R, Gadbaw B, DeSantes KB, et al. Genotypes of NK cell KIR receptors, their ligands, and Fcγ receptors in the response of neuroblastoma patients to Hu14.18-IL2 immunotherapy. Cancer Res. 2010; 70:9554–61.
28. Sáez-Borderías A, Romo N, Magri G, Gumá M, Angulo A, López-Botet M. IL-12-dependent inducible expression of the CD94/NKG2A inhibitory receptor regulates CD94/NKG2C+ NK cell function.J Immunol. 2009; 182:829–36.
29. Morandi F, Scaruffi P, Gallo F, Stigliani S, Moretti S, Bonassi S, Gambini C, Mazzocco K, Fardin P, Haupt R, Arcamone G; Italian Cooperative Group for Neuroblastoma, Pistoia V, et al. Bone marrow-infiltrating human neuroblastoma cells express high levels of calprotectin and HLA-G proteins. Plos One. 2012; 7:e29922.
30. Brooks AG, Borrego F, Posch PE, Patamawenu A, Scorzelli CJ, Ulbrecht M, Weiss EH, Coligan JE. Specific recognition of HLA-E, but not classical, HLA class I molecules by soluble CD94/NKG2A and NK cells. J Immunol. 1999; 162:305–13.
31. He W, Kuang Y, Xing X, Simpson RJ, Huang H, Yang T, Chen J, Yang L, Liu E, He W, Gu J. Proteomic comparison of 3D and 2D glioma models reveals increased HLA-E expression in 3D models is associated with resistance to NK cell-mediated cytotoxicity. J Proteome Res. 2014; 13:2272–81.
32. Kunisada T, Moseley JM, Slavin JL, Martin TJ, Choong PF. Co-expression of parathyroid hormone-related protein (PTHrP) and PTH/PTHrP receptor in cartilaginous tumors: a marker for malignancy? Pathology. 2002; 34:133–7.
33. Ui-Tei K, Naito Y, Takahashi F, Haraguchi T, Ohki-Hamazaki H, Juni A, Ueda R, Saigo K. Guidelines for the selection of highly effective siRNA sequences for mammalian and chick RNA interference. Nucleic Acids Res. 2004; 32:936–48.
34. Lee S, Qiao J, Paul P, Chung DH. Integrin β1 is critical for gastrin-releasing peptide receptor-mediated neuroblastoma cell migration and invasion. Surgery. 2013; 154:369–75.
35. Forte P, Baumann BC, Weiss EH, Seebach JD. HLA-E expression on porcine cells: protection from human NK cytotoxicity depends on peptide loading. Am J Transplant. 2005; 5:2085–93.
36. Enqvist M, Nilsonne G, Hammarfjord O, Wallin RP, Björkström NK, Björnstedt M, Hjerpe A, Ljunggren HG, Dobra K, Malmberg KJ, Carlsten M. Selenite induces posttranscriptional blockade of HLA-E expression and sensitizes tumor cells to CD94/NKG2A-positive NK cells. J Immunol. 2011; 187:3546–54.
37. Wolpert F, Roth P, Lamszus K, Tabatabai G, Weller M, Eisele G. HLA-E contributes to an immune-inhibitory phenotype of glioblastoma stem-like cells. J Neuroimmunol. 2012; 250:27–34.
38. Moriai S, Takahara M, Ogino T, Nagato T, Kishibe K, Ishii H, Katayama A, Shimizu N, Harabuchi Y. Production of interferon-{gamma}-inducible protein-10 and its role as an autocrine invasion factor in nasal natural killer/T-cell lymphoma cells. Clin Cancer Res. 2009; 15:6771–79.
39. Yao J, Qian CJ, Ye B, Zhao ZQ, Wei J, Liang Y, Zhang X. Signal transducer and activator of transcription 3 signaling upregulates fascin via nuclear factor-κB in gastric cancer: Implications in cell invasion and migration. Oncol Lett. 2014; 7:902–908.
40. Jiang J, Liu LY. Zinc finger protein X-linked is overexpressed in colorectal cancer and is associated with poor prognosis. Oncol Lett. 2015; 10:810–814.
41. Demirović A, Cesarec S, Marušić Z, Tomas D, Milošević M, Hudolin T, Krušlin B. TGF-β1 expression in chromophobe renal cell carcinoma and renal oncocytoma. Eur J Histochem. 2014; 58:2265–79.
42. Komada Y, Zhang XL, Zhou YW, Inaba H, Deguchi T, Azuma E, Sakurai M. Flow cytometric analysis of peripheral blood and bone marrow for tumor cells in patients with neuroblastoma. Cancer. 1998; 82:591–599.