
INTRODUCTION
World Health Organization (WHO) grade III anaplastic astrocytoma, and grade IV glioblastoma multiforme (GBM) are defined as high-grade gliomas with poor survival outcome [1, 2]. Conventional therapeutic strategies consist of extensive resection, concurrent chemo-radiotherapy. However, high-grade gliomas still progressed and recurred despite of aggressive therapies [3]. Therefore, biomarker for distinguishing clinical outcome is urgently needed in high-grade gliomas to provide oncotarget for clinician to improve the clinical outcome [4, 5].
Telomere maintenance 2 (TELO2) was first found in Saccharomyces cerevisiae to regulate the telomere length [6]. Later, TELO2 was found in Caenorhabditis elegans to control biological rhythms and life span and named CLK2 [7, 8]. TELO2 located in chromosome 16 and also called TEL2 [9]. TELO2 encodes protein for regulation of S-phase checkpoint protein in cell cycle, ataxia telangiectasia and Rad3-related (ATR) signaling pathway, and involves in DNA repair [10], and radiation sensitivity [11]. Studies showed TEL2/CLK2 orthologs increased the telomere length in Caenorhabditis elegans and the budding yeast Saccharomyces cerevisiae [12–15].
Overexpression of TELO2 accelerated cell cycle, induced cells hypersensitive to apoptosis caused by DNA replication block or oxidative stress, and gradually expanded telomere length in human liver adenocarcinoma cell line [9]. However, the correlation of TELO2 with the human gliomas is not yet elucidated. This study aims to clarify whether TELO2 expression correlated with survival prognosis and the WHO pathological grading. The Gene Expression Omnibus (GEO) profiles offer database to assess the gene expression of particular disease. Investigating TELO2 expression level in GEO dataset revealed higher expression and shorter survival outcome in high-grade gliomas than in low-grade gliomas. Validation of TELO2 using qRT-PCR, Western blot and immunohistochemical (IHC) staining supports the overexpression of TELO2 mRNA and protein level in high-grade gliomas, implying that TELO2 correlates with adverse outcome in high-grade gliomas.
RESULTS
Demographic data of human high-grade gliomas
The demographic data including gender, age, WHO pathologic grades, and survival times were shown in Table 1, and Table 2. There were total 100 patients contained in WHO grade IV (76), and grade III (24). After exclusion 20, and 3 missing data in WHO grade IV, and III (76, and 24 respectively), remaining 56 patients (18 female, 38 male) in grade IV population, and 21 patients (8 female, 13 male) in grade III population. The age of grade IV, and III patients were 48.52 ± 12.82, and 37.38 ± 9.86 years old, respectively (P = 0.001). The overall survival time in grade IV, and III was 106.73 ± 82.02, and 226.95 ± 161.12 weeks, respectively (P = 0.003) (Table 1). Moreover, as shown in Table 2, univariable analysis identified three significant predictors (WHO grade, TELO2 mRNA expression, and age). Multivariable analysis further confirmed WHO grade, and TELO2 mRNA expression were significant predictors (P = 0.0056, and 0.0141, respectively).
Table 1: Analyses of demographic data in human high-grade gliomas
Grade III |
Grade IV |
P value |
|
|---|---|---|---|
n=24 |
n=76 |
||
Gender |
0.968§ |
||
Male |
16(66.7%) |
51(67.1%) |
|
Female |
8(33.3%) |
25(32.9%) |
|
Age |
37.38±9.86a |
48.52±12.82b |
0.001* |
< 153.15 |
22(91.7%) |
52(68.4%) |
|
> 153.15 |
2(8.3%) |
24(31.6%) |
|
Survival time (weeks) |
226.95±161.12a |
106.73±82.02b |
0.003* |
TELO2: Telomere maintenance 2; *P<0.05, by Wilcoxon Test; §Chi-square analysis
a: There are three missing data; b: 20 missing data.
Table 2: Statistical analyses of the association between demographic data and survival time using Cox proportional hazard model
Univariable analysis Crude-HR (95% CI) |
P value |
Multivariable analysis |
P value |
|
|---|---|---|---|---|
WHO grade |
||||
III |
1.000 |
1.000 |
||
IV |
3.223 (1.735 to 5.987) |
0.0002* |
2.518 (1.310 to 4.840) |
0.0056* |
TELO2 mRNA expression |
||||
< 153.15 |
1.000 |
1.000 |
||
> 153.15 |
2.312 (1.350 to 3.959) |
0.0023* |
2.051 (1.156 to 3.640) |
0.0141* |
Gender |
||||
Male |
1.000 |
1.000 |
||
Female |
0.761 (0.468 to 1.235) |
0.2689 |
0.736 (0.443 to 1.223) |
0.2367 |
Age (per 20 years) |
1.587 (1.087 to 2.317) |
0.0169* |
1.226 (0.832 to 1.809) |
0.3032 |
WHO: World Health Organization; TELO2: Telomere maintenance 2; CI: confidence interval; *P<0.05 with statistical significance
Overexpression of TELO2 mRNA expression correlates with poor survival in high-grade gliomas
To investigate the role of TELO2 in human glioma, we first analyzed the correlation of TELO2 expression with overall survival of human high-grade gliomas. As shown in Figure 1, the Kaplan-Meier survival curve of 77 patients with high-grade gliomas revealed that patients with low TELO2 mRNA expression levels (n = 56) had longer overall survival than those with high TELO2 mRNA expression levels (n = 21) (P = 0.0017, by log-rank test; 95% CI: 0.1824-0.6735, Ratio 0.3505). The cut-off value was set at 153.15. The median survival interval in the high- and low-TELO2 expressions level was 59 weeks and 113 weeks, respectively. These data demonstrate that high TELO2 mRNA expression correlates with shorter survival in human high-grade gliomas, supporting the hypothesis of TELO2 belongs to oncogene.
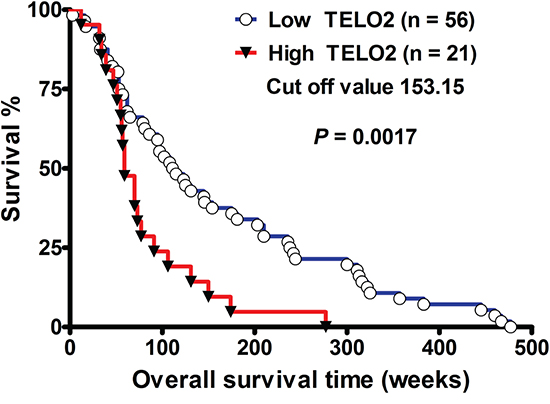
Figure 1: TELO2 mRNA overexpression decreases survival in human high-grade gliomas. The Kaplan-Meier survival curve showed shorter survival in those with high TELO2 (> 153.15) (n=21) compared to those with low TELO2 (< 153.15) (n=56) expression levels (median survival, 113 vs. 59 weeks, P=0.0017, by log-rank test, hazard ratio: 0.3505, 95% CI: 0.1824-0.6735).
Human high-grade gliomas increase TELO2 mRNA expression
Next, we analyzed the correlation of TELO2 mRNA expression and pathological grading. As shown in Figure 2, TELO2 mRNA expression level was statistically greater in WHO grade IV (n = 81) then in non-tumor controls (n = 23) (P = 2.85 x 10-9). Moreover, the TELO2 mRNA level was also significantly higher in WHO grade III (n = 19) than in non-tumor controls (P = 0.017, P adjusted by Bonferroni method). Consistently, these data further support the hypothesis of TELO2 belonging to oncogene in human high-grade gliomas.
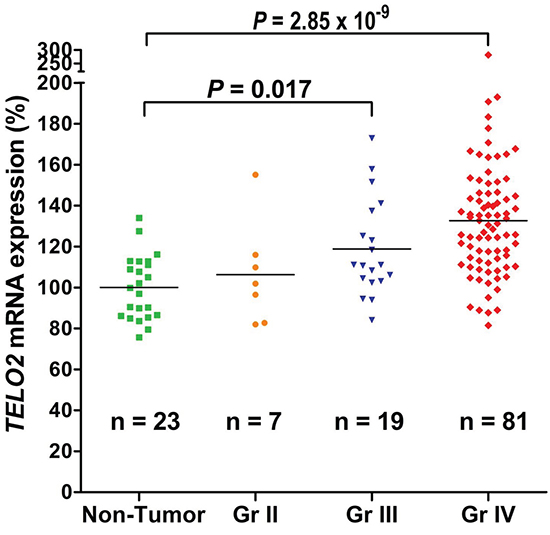
Figure 2: Human high-grade gliomas increase TELO2 mRNA expression. The scattered plots showed the TELO2 mRNA gene expression in WHO grade II, III, IV, and non-tumor control. Increased TELO2 mRNA levels significantly correlated with WHO grades of human high-grade gliomas. The Y-axis was TELO2 mRNA expression (%). The original mean value of non-tumor group was 292.7. The non-tumor group represented the baseline (100%). The P value was adjusted using Bonferroni method between each group.
Validation the level of TELO2 mRNA expression in human glioma cell lines
Since two dry lab datasets of quantitative gene study supporting the role of TELO2 is oncogene in human high-grade gliomas. We further validate the level of TELO2 mRNA expression in human normal brain and three glioma cell lines, LN229, GBM8401, and U118MG through wet lab approach using q-RT PCR. As shown in Figure 3, the mRNA expression of TELO2 was significantly higher in human LN229 glioma cell line than in normal brain (P < 0.05). Moreover, TELO2 was also statistically higher in human GBM8401 and U118MG glioma cell line than in normal brain (P < 0.01, P < 0.005, respectively). These data further consolidate the high expression levels of TELO2 mRNA in human gliomas.
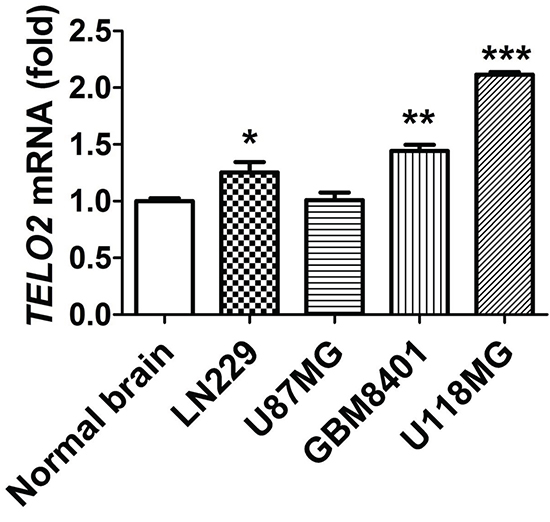
Figure 3: Validation of TELO2 mRNA expression in glioma cell lines and normal brain tissue. qRT-PCR was conducted to examine TELO2 mRNA expression and the quantitative results were shown. The relative expressions were normalized with normal brain. Bars, mean±SEM, *P < 0.05, **P < 0.01, ***P < 0.005 showed significant differences. Data are representative of three independent experiments.
TELO2 hub protein and the protein-protein interactions
The protein-protein interaction (PPI) network of TELO2-regulated oncogenesis was created using Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database. The network showed TELO2 is a hub protein regulating the signaling pathway among the mammalian target of rapamycin (mTOR), TELO2 interacting protein 1 (TTI1), ataxia telangiectasia and Rad3-related (ATR), ataxia telangiectasia-mutated (ATM), and RuvB-like 1 (RUVBL1) (Figure 4A). Moreover, the Ingenuity pathway analysis (IPA) predicts that TELO2 plays the transcriptional regulation role for ATM, mTOR, ATR and RUVBL1 to translocate into nucleus in signaling pathways (Figure 4B). Finally, the protein production of TELO2 and mTOR in human normal brain and four glioma cell lines, LN229, U87MG, GBM8401, and U118MG through wet lab approach using Western blotting (Figure 5A). The protein level of TELO2 significantly increased correlates with the mTOR expression in human glioma cell lines including U118MG, GBM8401, U87MG, and LN229 as compared with normal brain tissue (Figure 5B and 5C). Taken together, these studies demonstrated the expression of TELO2 correlated with mTOR expression in human gliomas.
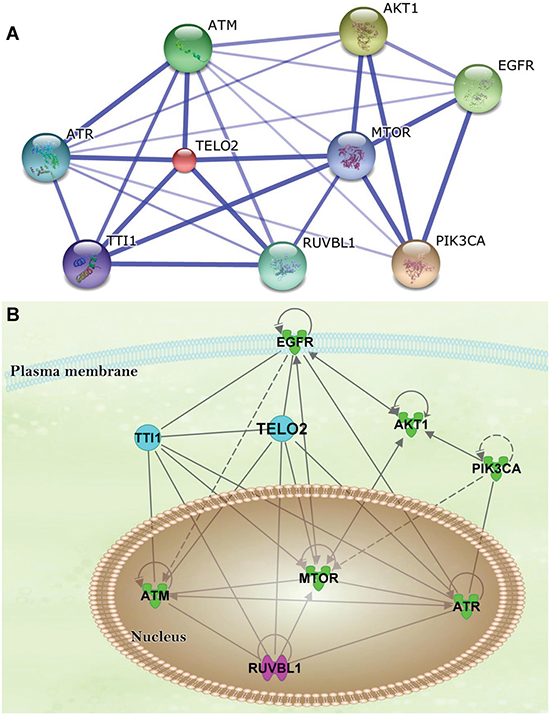
Figure 4: The protein-protein interaction (PPI) network and ingenuity pathway analysis (IPA). A. The PPI network established by the STRING database. TELO2 is controlling hub; and B. The IPA predicted the mTOR-regulated AKT1, EGFR, and PIK3CA signaling pathways transcriptionally regulated through TELO2.
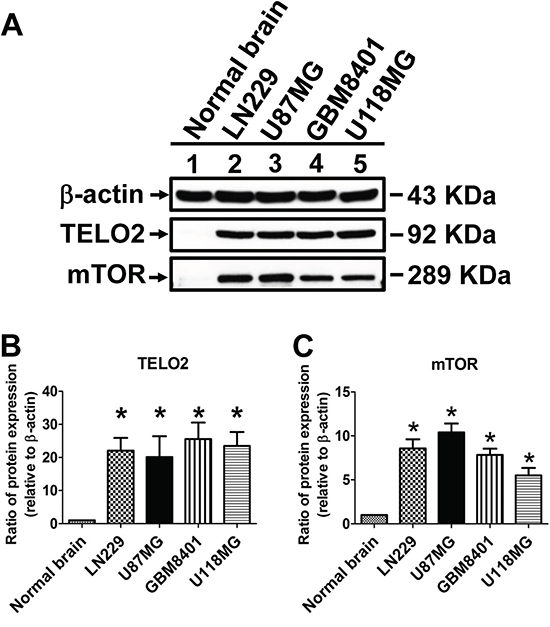
Figure 5: Validation and statistical analyses of TELO2 and mTOR protein levels. A. Protein lysates of glioma cell lines, including LN229, U87MG, GBM8401 and U118MG, were applied to SDS-PAGE and Western blotting. The statistical analyses of TELO2 and mTOR protein levels were presented in B and C. β-actin served as a loading control. Bars, mean±SEM, *P < 0.05 showed significant differences in comparison with normal brain control.
TELO2 protein production overexpresses in human high-grade gliomas
To investigate the TELO2 protein production in human gliomas tissues and normal brain tissues, IHC staining of human tissue microarray was conducted (Figure 6A to 6J). The TELO2 immunostain score was significant higher in high-grade (WHO grade III, and IV) gliomas than in low-grade (WHO grade I, and II) gliomas or normal brain (P = 2.19x10-3, 7.53x10-6, respectively). Moreover, TELO2 immunostain score was significant higher in low-grade gliomas than in normal brain (P = 1.54x10-4, P adjusted by Bonferroni method, Figure 6K), supporting TELO2 protein overexpressed in high-grade gliomas in comparison with normal brain tissue control.
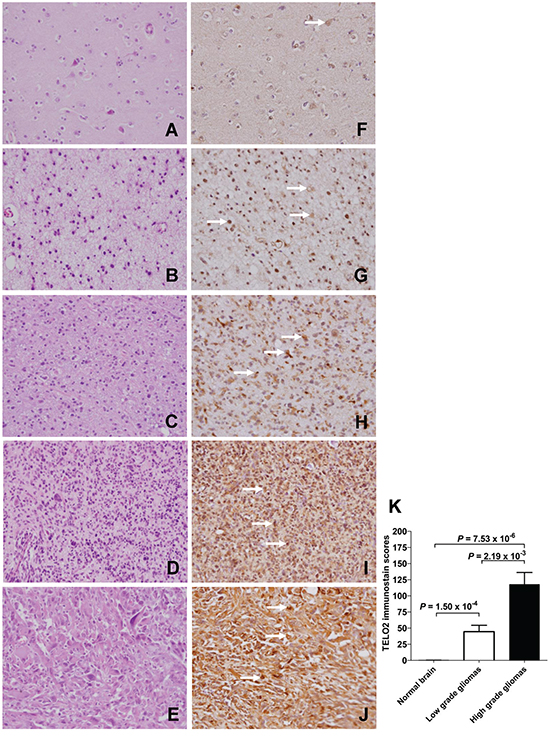
Figure 6: Overexpression of TELO2 in high-grade gliomas compared to low-grade gliomas and non-tumor control. Representative hematoxylin and eosin staining of A. normal brain, B. pilocytic astrocytomas, C. diffuse astrocytomas, D. anaplastic astrocytomas, and E. glioblastomas multiforme. Representative immunohistochemical staining of TELO2 in F. normal brain, G. pilocytic astrocytomas, H. diffuse astrocytomas, I. anaplastic astrocytomas, and J. glioblastomas multiforme. K. Statistical analysis showed the TELO2 immunostain score was significantly higher in high-grade (WHO grade III, and IV) gliomas than in in low-grade (WHO grade I, and II) gliomas, and normal brain (P = 2.19x10-3, 7.53x10-6, respectively), Moreover, the TELO2 immunostain score was significantly higher in low-grade gliomas than in normal brain control (P = 1.54x10-4, P adjusted by Bonferroni method). The significant expression of TELO2 in glioma cell was labeled by arrow marks. Original magnification x 400.
DISCUSSION
This study showed that high TELO2 mRNA expression predicts poor survival outcome in comparison with low TELO2 mRNA expression. Moreover, TELO2 expression was significantly greater in patients with high-grade gliomas than in low-grade gliomas and in those with non-tumor brain controls. Validation using qRT-PCR, Western blotting, and IHC staining further confirmed that TELO2 overexpresses in high-grade gliomas. To our understanding, this is the first study to launch the correlation of TELO2 biomarker with WHO pathological grading in human gliomas, and survival outcome, supporting TELO2 belongs to oncogene.
TELO2 as an essential orphan protein involved in many function including DNA repair, telomere maintenance [13], the biological clock, signal transduction pathways, and DNA damage checkpoints [11, 16–18]. Treatment of TELO2 (also called CLK2) small interference RNA significantly decreased growth rate and arrested the cell cycle reversibly of human hepatic adenocarcinoma cell. In contrast, overexpression of TELO2 increased the growth rate of SK-HEP-1 cells through the mechanisms of shortened cell cycle length, and gradually expanded telomere length in human SK-HEP-1 liver adenocarcinoma cell line [9], suggesting the role of TELO2 for glioblastoma growth. Both genomic and proteomic investigations showed that there is virtually common activation of the PI3K/Akt/ mTOR signaling pathway in human glioblastomas [1, 19–21]. PI3K signaling increases glioblastomas formation and tumor progression in mouse-engineered models, launching PI3K and its effecter mTOR as convincing targets. mTOR incorporates oncogenic signaling from some growth factor receptors via PI3K, to modulate nutrient status and cellular energy, to stimulate downstream targets that promote tumor growth and suppress glioma cell invasion [22, 23]. The standard chemotherapy for GBM is temozolamide (TMZ) [24], an alkylation agent that leads to DNA damage. However, the therapeutic effect of TMZ will decrease, and the drug resistance occurs due to the activation of PI3K/Akt/mTOR signaling pathway. Tel2 and Tti1 stabilization correlated with the elevated expression levels of all phosphatidylinositol 3-kinase related protein kinase (PIKK) family. This effect was most obvious in mTOR [25]. Consistently, our data further showed higher expression of TELO2 correlated with the poor survival outcome in high-grade gliomas, supporting the oncogenic role of TELO2.
We applied STRING to identify the PPI [26]. STRING platform found some important proteins were linked by hub protein TELO2, and mTOR. PPI predicts the TELO2 protein is involved in transcriptional regulation of the mTOR, RUVBL1, ATR, and ATM in signaling pathways. Moreover, mTOR also plays a hub protein of TELO2, ATM, AKT1, EGFR, PIK3CA, and RUVBL1 in signaling pathways. TELO2 is required for the stable expression of all six members of the PIKK family, including ATM, ATR, DNA-dependent protein kinase catalytic subunit (DNA-PKcs), and mTOR [27]. Importantly, mTOR inhibition related to feedback activation of PI3K/Akt signaling and regulation of other crucial cellular signals in human glioblastoma to escape from the effect of mTOR inhibitor rapamycin [20]. TELO2, TTI1 and TTI2 (Tel two-interacting protein 2) compose a 2-MDA complex called the Triple T complex. DNA damages response (DDR) is critical for maintain genomic stability, which is a complex signal transduction network including activated transcription and DNA repair, and coordinated cell cycle transitions. ATM and ATR are central regulators of the DDR. The triple T complex was noted required for DNA damage single, and stabilize the ATM and ATR [10]. In addition, depletion of TELO2 would decrease the mTOR protein level but not the mRNA level through the binding of R2TP Complex to TELO2 which phosphorylated by casein kinase 2 (CK2) [27, 28]. These studies suggested that the TELO2 gene could be the novel therapeutic targets for high-grade gliomas. Further study of downstream targets (pS6k, 4EBP1, pp70S6k) in signaling pathways activated in glioma cell lines would be another one interesting study in the future. The main theme of this study is to clarify the novel role of TELO2 expression in clinical significance of high-grade glioma patients. Further statistical analysis of patient clinical samples in serial histological sections would explore a novel discovery between mTOR and TELO2 in future study.
The limitation of this study included difficulty in collecting massive amounts of human brain gliomas, grade I low-grade glioma samples are rare, and paired non-tumor control for validation of protein level or mRNA expression. Alternatively, large-scale analyses of total 280 sheets of data from two GEO profiles were conducted to determine the role of TELO2 as a pathological grade and survival outcome marker. Since the gene expression profilings were quite different between tissue and cell lines, therefore, low grade compared with high grade glioma cell lines would be a better experimental design. Alternatively, the normal cell line could be as a reference.
In conclusion, human high-grade gliomas increase TELO2 mRNA expression. Overexpression of TELO2 mRNA expression correlates with shorter survival outcome. Validation of gene and protein levels, using qRT-PCR, Western blotting, and IHC staining, consistently supports that TELO2 is an oncotarget in human gliomas.
MATERIALS AND METHODS
Analysis of human gliomas datasets from GEO profile
This study was approved by the institutional review board (TSGHIRB No: B-102-10) of Tri-Service General Hospital in Taipei, Taiwan, ROC. The genomic databases obtained from the GEO profile were analyzed by previous methodology [29, 30, 32]. Briefly, 100 sheets of de-linked data (GDS1815 / 209528_s_at / TELO2) on TELO2 mRNA expression, age, gender, overall survival time, and pathologic grading of primary high-grade glioma were gathered from http://www.ncbi.nlm.nih.gov/geo/tools/profileGraph.cgi?ID=GDS1815:209528_s_at. Twenty-three sheets of data without thorough information on survival period and age were excluded so that 77 sheet were collected in the statistical analyses.
The analysis of TELO2 gene expression and pathological grading obtained from additional database GDS1962 / 34260_s_at / TELO2 including 180 sheets from 81 patients with grade IV glioma, 19 with grade III glioma, seven with grade II glioma, 23 without tumor (non-tumor control) searching from http://www.ncbi.nlm.nih.gov/geo/tools/profileGraph.cgi?ID=GDS1962:34260_at.
RNA isolation and cDNA synthesis
Total RNA of glioma cell lines were extracted using the EasyPure Total RNA reagent (Bioman, Taiwan, ROC) according to the manufacturer’s protocol. Briefly, cells were harvested and lysed in EasyPure Total RNA reagent. After the addition of chloroform followed by centrifugation, the aqueous phase were transferred into a new tube for RNA recovery by precipitation with isopropyl alcohol. cDNA were prepared using oligo dT, and MMLV Reverse transcriptase (Epicentre Biotechnologies, USA). The normal brain cDNA was purchased from Origene Technologies (Rockville, MD, USA).
Quantitative real-time PCR (qRT-PCR)
qRT-PCR was carried out in a 20 μl volume using IQ2 fast qPCR system with ROX (Bio-genesis Technology Inc., Taipei, Taiwan) in illumine ECO™ Real-Time PCR system. The relative gene expression was analyzed using the 2-ΔΔCt method against GAPDH expression. PCR primers for each gene were obtained from PrimerBank (http://pga.mgh.harvard.edu/primerbank/links.html). The primer pairs of TELO2 and GAPDH were summarized in Table 3.
Table 3: Sequences of primers used in polymerase chain reaction
Gene |
Oligonucleotide |
Sequence (5’-3’) |
Product Size (bp) |
|---|---|---|---|
TELO2 |
Forward |
CCCGCAGAGATCGTGGATG |
96 bp |
Reverse |
CATGTCGTAGGGGACAAACTC |
||
GAPDH |
Forward |
GCACCGTCAAGGCTGAGAAC |
142 bp |
Reverse |
ATGGTGGTGAAGACGCCAGT |
Cell lysate preparation and western blotting
Cells were lysed by RIPA buffer (100 mM Tris-HCl, 150 mM NaCl, 0.1% SDS, and 1% Triton-X-100) and the cell lysates were harvested by centrifugation at 15,000 rpm for 10 min to remove the debris. The normal brain lysate were purchased from Origene Technologies. Thirty-microgram cell lysates from each group were applied to 10% sodium dodecyl sulfate polyacrylamide gels electrophoresis and proteins were transferred onto polyvinyl difluoride membranes (Millipore, MA, USA). The membrane was blocked with 5% skim milk in TBST for 1 h at room temperature. The antibodies used include anti-TELO2 (Cat. No. ab122722, Abcam, Cambridge, UK), mTOR antibody (cat. No. 2972; Cell Signaling Technology, Beverly, Boston, MA, USA), and β-actin (Santa Cruz Biotechnology, Inc.). Band detection was conducted by enhanced chemi-luminescence and X-ray film (GE Healthcare, Piscataway, NJ, USA).
Protein-protein interactions network analysis
Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database (http://string-db.org) was applied for analysis of predicted and known PPI [26]. PPI of TELO2 was performed according to the method of our previous publication [29].
Immunohistochemical staining of human gliomas specimen
IHC staining of commercially available tissue microarray (BS17016; Biomax, Rochester, NY, USA) was performed according to previous protocol [29–31], incubated with a polyclonal rabbit anti-human TELO2 antibody (Cat. No. ab122722, Abcam, Cambridge, UK) (1:100 diluted in phosphate buffered saline (PBS) for 1 h at room temperature, washed 3 times (each for 5 min in PBS), incubated with biotin-labeled secondary immunoglobulin (1:100, DAKO, Glostrup, Denmark) for 1 h at room temperature, washed 3 times, and treated with 3-amino-9-ethylcarbazole substrate chromogen (DAKO) at room temperature to visualize peroxidase activity [32]. Sections of human urinary bladder cancer tissue (known to stain positive for TELO2) were used as positive control according to the datasheet of TELO2 antibody (ab122722).
Statistical analysis
The methodology of statistical analysis has been defined proceeding [17, 30]. Two datasets (GDS1815 / 209528_s_at / TELO2 and GDS1962 / 34260_s_at / TELO2) gather from the GEO profiles were analyzed. The single tail test was used to calculate the TELO2 expression level in four groups of WHO pathologic grades gilomas. The Bonferroni method was used to adjust the P value to avoid the potentiality of type I error in multi-groups analyses. Overall survival was analyzed by The Kaplan-Meier method. Cohorts of low-versus high-TELO2 gene expressions were evaluated in WHO grade IV combined with grade III human glioma groups. Chi-square test was applied to analyze survival parameters in patients from high-grade gilomas. The cut-off value of TELO2 expression is decided based on conditional inference tree via ‘party’ package with R language (R 3.1.2 software). Conditional inference tree is a ‘decision tree’-based method. It can help us to classify patients and reduce the entropy of data. GraphPad Prism 5 software was used to draw figures. The Y-axis was presented as TELO2 mRNA expression (%). The TELO2 mRNA expression of each WHO grade glioma showed in percentage to non-tumor group (baseline 100%). The P value less than 0.05 reached statistical significance.
ACKNOWLEDGMENTS
This study was supported in part by grants from Health and Welfare surcharge of tobacco to the Ministry of Health and Welfare (MOHW104-TDU-B-211-124-005, MOHW105-TDU-B-211-134001), Ministry of Science and Technology (MOST 102-2628-B-016 -002 -MY2, MOST 103-2314-B-016 -027-MY3, MOST 103-2911-I-016 -501, and MOST104-2911-I-016 -501), and Tri-Service General Hospital (TSGH-C104-008-S04, and TSGH-C105-079), Ministry of National Defense-Medical Affairs Bureau (103-M071, MAB-104-047), Taipei, Taiwan, R.O.C.
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Jan HJ, Lee CC, Shih YL, Hueng DY, Ma HI, Lai JH, Wei HW, Lee HM. Osteopontin regulates human glioma cell invasiveness and tumor growth in mice. Neuro Oncol. 2010; 12:58-70.
2. Rachet B, Mitry E, Quinn MJ, Cooper N, Coleman MP. Survival from brain tumours in England and Wales up to 2001. Br J Cancer. 2008; 99:S98-101.
3. Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn U, Curschmann J, Janzer RC, Ludwin SK, Gorlia T, Allgeier A, Lacombe D, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005; 352:987-996.
4. Roversi G, Pfundt R, Moroni RF, Magnani I, van Reijmersdal S, Pollo B, Straatman H, Larizza L, Schoenmakers EF. Identification of novel genomic markers related to progression to glioblastoma through genomic profiling of 25 primary glioma cell lines. Oncogene. 2006; 25:1571-1583.
5. Zhao M, Xu H, Zhang B, Hong B, Yan W, Zhang J. Impact of nuclear factor erythroid-derived 2-like 2 and p62/sequestosome expression on prognosis of patients with gliomas. Hum Pathol. 2015; 46:843-849. doi: 10.1016/j.humpath.2015.02.009.
6. Lustig AJ, Petes TD. Identification of yeast mutants with altered telomere structure. Proc Natl Acad Sci U S A. 1986; 83:1398-1402.
7. Hekimi S, Boutis P, Lakowski B. Viable maternal-effect mutations that affect the development of the nematode Caenorhabditis elegans. Genetics. 1995; 141:1351-1364.
8. Lakowski B, Hekimi S. Determination of life-span in Caenorhabditis elegans by four clock genes. Science. 1996; 272:1010-1013.
9. Jiang N, Benard CY, Kebir H, Shoubridge EA, Hekimi S. Human CLK2 links cell cycle progression, apoptosis, and telomere length regulation. The Journal of biological chemistry. 2003; 278:21678-21684.
10. Hurov KE, Cotta-Ramusino C, Elledge SJ. A genetic screen identifies the Triple T complex required for DNA damage signaling and ATM and ATR stability. Genes Dev. 2010; 24:1939-1950.
11. Ahmed S, Alpi A, Hengartner MO, Gartner A. C. elegans RAD-5/CLK-2 defines a new DNA damage checkpoint protein. Curr Biol. 2001; 11:1934-1944.
12. Benard C, McCright B, Zhang Y, Felkai S, Lakowski B, Hekimi S. The C. elegans maternal-effect gene clk-2 is essential for embryonic development, encodes a protein homologous to yeast Tel2p and affects telomere length. Development. 2001; 128:4045-4055.
13. Kota RS, Runge KW. Tel2p, a regulator of yeast telomeric length in vivo, binds to single-stranded telomeric DNA in vitro. Chromosoma. 1999; 108:278-290.
14. Lim CS, Mian IS, Dernburg AF, Campisi J. C. elegans clk-2, a gene that limits life span, encodes a telomere length regulator similar to yeast telomere binding protein Tel2p. Curr Biol. 2001; 11:1706-1710.
15. Runge KW, Zakian VA. TEL2, an essential gene required for telomere length regulation and telomere position effect in Saccharomyces cerevisiae. Mol Cell Biol. 1996; 16:3094-3105.
16. Collis SJ, Barber LJ, Clark AJ, Martin JS, Ward JD, Boulton SJ. HCLK2 is essential for the mammalian S-phase checkpoint and impacts on Chk1 stability. Nature cell biology. 2007; 9:391-401.
17. Rendtlew Danielsen JM, Larsen DH, Schou KB, Freire R, Falck J, Bartek J, Lukas J. HCLK2 is required for activity of the DNA damage response kinase ATR. The Journal of biological chemistry. 2009; 284:4140-4147.
18. Shikata M, Ishikawa F, Kanoh J. Tel2 is required for activation of the Mrc1-mediated replication checkpoint. The Journal of biological chemistry. 2007; 282:5346-5355.
19. Comprehensive genomic characterization defines human glioblastoma genes and core pathways. Nature. 2008; 455:1061-1068.
20. Fan QW, Weiss WA. Targeting the RTK-PI3K-mTOR axis in malignant glioma: overcoming resistance. Current topics in microbiology and immunology. 2010; 347:279-296.
21. Li X, Wu C, Chen N, Gu H, Yen A, Cao L, Wang E, Wang L. PI3K/Akt/mTOR signaling pathway and targeted therapy for glioblastoma. Oncotarget. 2016; doi: 10.18632/oncotarget.7961.
22. Jiang Y, Zhang Q, Bao J, Du C, Wang J, Tong Q, Liu C. Schisandrin B suppresses glioma cell metastasis mediated by inhibition of mTOR/MMP-9 signal pathway. Biomedicine & pharmacotherapy. 2015; 74:77-82.
23. Tanaka K, Sasayama T, Kohmura E. Targeting glutaminase and mTOR. Oncotarget. 2015; 6:26544-26545. doi: 10.18632/oncotarget.5263.
24. Messaoudi K, Clavreul A, Lagarce F. Toward an effective strategy in glioblastoma treatment. Part I: resistance mechanisms and strategies to overcome resistance of glioblastoma to temozolomide. Drug discovery today. 2015; 20:899-905.
25. Fernandez-Saiz V, Targosz BS, Lemeer S, Eichner R, Langer C, Bullinger L, Reiter C, Slotta-Huspenina J, Schroeder S, Knorn AM, Kurutz J, Peschel C, Pagano M, Kuster B, Bassermann F. SCFFbxo9 and CK2 direct the cellular response to growth factor withdrawal via Tel2/Tti1 degradation and promote survival in multiple myeloma. Nature cell biology. 2013; 15:72-81.
26. Szklarczyk D, Franceschini A, Wyder S, Forslund K, Heller D, Huerta-Cepas J, Simonovic M, Roth A, Santos A, Tsafou KP, Kuhn M, Bork P, Jensen LJ, von Mering C. STRING v10: protein-protein interaction networks, integrated over the tree of life. Nucleic acids research. 2015; 43:D447-452.
27. Takai H, Wang RC, Takai KK, Yang H, de Lange T. Tel2 regulates the stability of PI3K-related protein kinases. Cell. 2007; 131:1248-1259.
28. Horejsi Z, Takai H, Adelman CA, Collis SJ, Flynn H, Maslen S, Skehel JM, de Lange T, Boulton SJ. CK2 phospho-dependent binding of R2TP complex to TEL2 is essential for mTOR and SMG1 stability. Molecular cell. 2010; 39:839-850.
29. Hueng DY, Tsai WC, Chiou HY, Feng SW, Lin C, Li YF, Huang LC, Lin MH. DDX3X Biomarker Correlates with Poor Survival in Human Gliomas. International journal of molecular sciences. 2015; 16:15578-15591.
30. Hueng DY, Lin GJ, Huang SH, Liu LW, Ju DT, Chen YW, Sytwu HK, Chang C, Huang SM, Yeh YS, Lee HM, Ma HI. Inhibition of Nodal suppresses angiogenesis and growth of human gliomas. Journal of neuro-oncology. 2011; 104:21-31.
31. Tsai WC, Hueng DY, Lin CK. Nuclear overexpression of urocortin discriminates primary brain tumors from reactive gliosis. APMIS. 2015; 123:465-472.
32. Tsai WC, Chen Y, Huang LC, Lee HS, Ma HI, Huang SM, Sytwu HK, Hueng DY. EMMPRIN expression positively correlates with WHO grades of astrocytomas and meningiomas. Journal of neuro-oncology. 2013; 114:281-290.