
Introduction
Alzheimer’s disease (AD) is defined clinically by gradual decline in cognition and function as well as loss of memory and pathologically by extensive neuronal and synaptic loss, intracellular neurofibrillary tangles and extracellular amyloid-β (Aβ) peptides deposition [1, 2]. Genetic variations are widely considered to have vital roles in the progress of AD [3]. Mutations in β-amyloid precursor protein (APP), presenilin 1 (PSEN 1) and presenilin 2 (PSEN 2) genes contribute to the uncommon early-onset form of the disease (onset < 65 years; EOAD) [4]. As to the more common late-onset AD (onset > 65 years; LOAD), only the APOE ε4 has been consistently identified to be a risk factor of the disease [5]. In addition, some GWAS have identified 21 additional genetic susceptibility factors for LOAD including bridging integrator 1 (BIN1), inositol polyphosphate-5-phosphatase D (INPP5D), triggering receptor expressed on myeloid cell 2 (TREM2), NME8 (encoding NME/NM23 family member 8), the clusterin gene (CLU), protein tyrosine kinase 2β (PTK2B), desmoglein 2 (DSG2) and others [6]. However, the known LOAD genes mentioned above are not sufficient to account for the total genetic variance, indicating additional risk genes or locus remain to be discovered [7].
Among the known genetic factors, MEF2C gene is supposed to be connected with synaptic function that is altered in AD. Transcription factors MEF2 family (MEF2A-D) modulate the structural and synaptic plasticity underlying memory formation [8] and can regulate the synapse number in the hippocampus in which its activation restrains the development of dendritic spines, highlighting its important roles in memory and learning [9]. A recent work revealed that the overexpression of MEF2 function impeded the learning-induced increases in spine density and impaired memory formation via an Arc-mediated reduction in the surface expression of the GluA2-AMPA-type glutamate receptor [10]. In brain, MEF2C is highly expressed in the regions which are closely related with learning and memory such as dentate gyrus, frontal cortex, entorhinal cortex, and amygdala [11]. On the other hand, MEF2C may be involved in the inflammatory process altered in AD via the regulation of microglia proliferation [11]. In addition, MEF2 seems to play an important role in APP-mediated anti-apoptotic neuroprotection [12], and MEF2C is identified as a regulator of APP proteolytic process in which Amyloid-β (Aβ), one central factor to initiate AD pathogenesis, is produced [13]. Recently, a large meta-analysis has confirmed that the SNP rs190982 polymorphism within MEF2C on chromosome 5 acted as a protective factor for AD dementia (OR = 0.93, 95%CI = 0.90-0.95, P = 3.2×10-8) in the Caucasian population [14]. Similarly, the association between the MEF2C gene rs190982 polymorphism and AD was also confirmed in a large Spanish sample (OR = 0.885, 95%CI = 0.811-0.966) [15]. Furthermore, another two studies replicated the association in China Han. Regretfully, they failed to reveal any association of rs190982 polymorphism in MEF2C gene with AD susceptibility in our Han Chinese cohort [16, 17]. Here, we conducted a case-control study containing 2332 individuals to clarify whether the selected SNP on MEF2C gene has an association with LOAD in Northern Han Chinese population.
Results
We enrolled 2332 ethnic Northern Han Chinese subjects including 984 subjects (42.20%) with probable LOAD and 1348 healthy control subjects (57.80%). Table 1 lists the demographic and clinical characteristics of the recruited subjects. LOAD cases and control group were matched in terms of age (P = 0.242) and gender (P= 0.088). As expected, AD patients showed significantly lower MMSE scores (11.99±6.198) than controls (28.99±0.124; P < 0.001). The APOEε4 allele frequency was also significantly different between patients and control subjects, and was a risk factor of LOAD (OR = 2.409, 95%CI = 1.960-2.962, P < 0.001) (Table1).
Table 1: The characteristics of the study population
AD (n= 984) |
Controls (n= 1348) |
P |
OR(95%CI) |
||
Age, years; mean±SD |
75.15±6.079a |
75.46±6.463b |
0.242 |
||
Gender, n (%) |
0.088 |
||||
Male |
406 (41.3) |
604(44.8) |
|||
Female |
578 (58.7) |
744 (55.2) |
|||
MMSE score, mean±SD |
11.99±6.198 |
28.99±0.124 |
<0.001* |
||
APOE ε4 status, n (%) |
<0.001* |
2.409(1.960-2.962) |
|||
APOE ε4 (+) |
280 (28.5) |
191 (14.2) |
|||
APOE ε4 (-) |
704 (71.5) |
1157 (85.8) |
|||
Abbreviations: AD, Alzheimer's disease; n, number; OR, odds ratio; CI, confidence interval; SD, standard deviation; MMSE, Mini-Mental State Examination; APOE ε4 (+): subjects who contain 1 or 2 ε4 alleles; APOE ε4 (-): subjects who do not contain ε4 allele.
*P < 0.05, significant values.
a Mean age at onset.
b Mean age at examination.
Genotype distributions of the rs190982 was in Hardy-Weinberg equilibrium (HWE) in controls (P > 0.05) but was not in the HWE (P = 0.02) in LOAD. The genotype and allele frequencies of rs190982 in AD patients and controls in the total sample and after stratification for APOE ε4 allele were summarized in Table 2. There is no statistical significance between the frequency of the minor allele G in LOAD and that in controls (16.06% versus 16.25%). No significant difference was found in allele frequency between LOAD patients and controls (OR = 0.986, 95% CI = 0.842-1.155, P = 0.862). Identically, the genotype distribution did not differ significantly between this two groups (P = 0.861). Furthermore, multivariate analysis also failed to demonstrate any significant differences between LOAD and controls after adjustment for age, gender, and the APOE ε4 allele carrier status as shown in Table 3 (Dominant, OR = 1.001, 95% CI = 0.835-1.200, P = 0.993; Recessive, OR = 0.737, 95% CI = 0.388-1.397, P = 0.349; and Additive, OR = 0.980, 95% CI = 0.830-1.156, P = 0.810). Our sample size had over 90% power to detect the moderate difference in allele and genotype distributions between LOAD cases and controls at a 0.05 significance level.
Table 2: Allele and genotype frequencies of the SNP rs190982 in total subjects and stratified by ApoEε4 status respectively.
rs190982 |
Total samples |
ApoE ε4 carriers |
ApoE ε4 non-carriers |
||||||
AD (n%) |
Control (n%) |
P |
AD (n%) |
Control (n%) |
P |
AD (n%) |
Control (n%) |
P |
|
A/A |
684(69.51) |
936(69.44) |
0.861 |
194 (69.28) |
135 (70.68) |
0.858 |
490 (69.60) |
801 (69.23) |
0.445 |
A/G |
284(28.86) |
386(28.64) |
78 (27.86) |
52 (27.23) |
206 (29.26) |
334 (28.87) |
|||
G/G |
16(1.63) |
26(1.93) |
8 (2.86) |
4 (2.09) |
8 (1.14) |
22 (1.90) |
|||
A |
1652(83.94) |
2258(83.75) |
0.862 |
466 (83.21) |
322 (84.29) |
0.660 |
1186 ( 84.23) |
1936 (83.67) |
0.647 |
G |
316(16.06) |
438 (16.25) |
94 (16.79) |
60 (15.71) |
222 (15.77) |
378 (16.33) |
|||
Abbreviations: AD, Alzheimer’s disease; ApoE ε4 carriers, subjects who contain 1 or 2 ε4 alleles; ApoE ε4 non-carriers, subjects who do not contain ε4 allele.
We then explored the effects of the interaction between rs190982 polymorphisms and APOE genotype on the risk of LOAD, and no interaction between rs190982 and APOE genotype was observed (Table 3). To further investigate whether the presence of the APOE ε4 allele significantly influenced the effect of MEF2C on LOAD, we stratified the data according to APOE ε4 status. Finally, there were still no differences in the genotypic or allelic distributions between the two groups (Table 2), and rs190982 did not influence the risk for LOAD adjusting for age and gender in APOE ε4 allele carriers or non-carriers (Table 3).
Table 3: Logistic regression analysis of rs190982 polymorphisms.
Model |
Total samplea |
ApoE ε4 carriersb |
ApoE ε4 non-carriersb |
||||
OR(95%CI) |
P |
P for APOE interaction |
OR(95%CI) |
P |
OR(95%CI) |
P |
|
rs190982 |
|||||||
Dom |
1.001(0.835-1.200) |
0.993 |
0.734 |
1.062(0.710-1.588) |
0.771 |
0.986(0.804-1.209) |
0.890 |
Rec |
0.737(0.388-1.397) |
0.349 |
0.208 |
1.441(0.425-4.881) |
0.557 |
0.551(0.243-1.248) |
0.153 |
Add |
0.980(0.830-1.156) |
0.810 |
0.534 |
1.082(0.758-1.544) |
0.663 |
0.954(0.790-1.151) |
0.622 |
Abbreviations: ApoE ε4 carrier, subjects who contain 1 or 2 ε4 alleles; ApoE ε4 non-carriers, subjects who do not contain ε4 allele; OR, odds ratios; CI, confidence interval; Dom, dominant model; Rec, recessive model; Add, additive model.
a Adjusted for age, gender and APOE ε4 status,
b Adjusted for age and gender.
Finally, we performed a meta-analysis about the relationship between rs190982 and LOAD and detected that rs190982 had a protective role in LOAD (OR = 0.93, 95%CI = 0.90-0.95) with no heterogeneity (I2 = 0.0%) in the 82507 individuals without ethnic stratification. Then, rs190982 reduced the risk for LOAD in meta-analysis of 78418 Caucasian individuals (OR = 0.92, 95%CI = 0.90-0.95); however, when we combined our data with results from other two studies in China, MEF2C (rs190982) was not indicated to be related to LOAD risk (OR = 1.03, 95%CI = 0.90-1.18) (Figure 1). In addition, there is no publish bias in our meta-analysis and the funnel plot is listed (Figure 2).
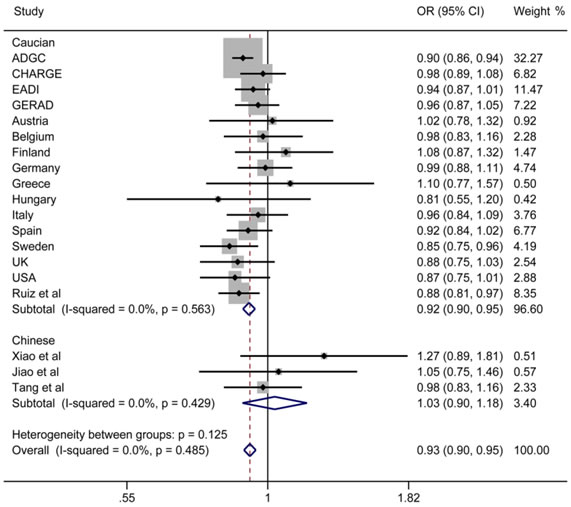
Figure 1: Forest plots for rs190982 in LOAD and healthy controls in 82507 individuals.
Figure 2: Begg' funnel plot with pseudo 95% confidence limits.
Discussion
Myocyte enhancer factor 2 (MEF2) family is a member of the transcription factors, which is vital for linking external stimuli to protein production. MEF2C is more widely expressed and modulates the development and differentiation of many tissues as well as other various transcriptional events [18]. Gene transcription and protein synthesis are also critical for the long-term memory formation. Individual members of the MEF2 family can differentially regulate the expression of brain-derived neurotrophic factor (BDNF), which performs essential roles in modulating synaptic growth and function [19]. MEF2 is also a regulator of the activity-regulated cytoskeleton-associated (ARC) which is in the controller of memories. As such, MEF2 remains a key player in memory processes [20]. In addition, MEF2 decreases spine growth that is critical for the cognition and memory formation and several forms of intellectual disability have been strongly related with the changed spine densities in both animal models and human patients [8, 21]. Moreover, MEF2 also participates in time-dependent reorganization and context memory consolidation [8]. Interestingly, MEF2 and an amount of MEF2 target genes have been implied in several neuropsychiatric and cognitive disorders, which indicate that at least some of the cognitive deficits may be ascribe to the dysfunction of MEF2.
Our study did not detect any association of SNP rs190982 polymorphism within MEF2C with the development of LOAD in Han Chinese population, even after statistical adjustment for age, gender and APOE ε4 status in the total sample and stratification for APOE ε4 status. Similarly, a research with the same aim conducted in China Han also did not declare any significant association between rs190982 and LOAD (P = 0.196, OR = 1.27, 95%CI = 0.89-1.81) [16]. Subsequently, Jiao et al. [17] draw the same conclusion that MEF2C gene rs190982 polymorphism had no relation with LOAD as well (P = 0.798, OR = 1.047, 95%CI = 0.747-1.461). In contrast to our results, Lambert et al. [14] identified rs190982 as a protective factor to the development of LOAD in Caucasian patients in a large-scale GWAS (OR = 0.93, 95%CI = 0.90-0.95). In addition, Ruiz et al. [15] also detected that rs190982 could decrease the risk of AD through multiple test in a large Spanish sample (OR = 0.885, 95%CI = 0.81 1-0.966).
Several factors may be responsible for these discrepancies in these researches. The key one of these factors is genetic heterogeneity in various ethnic populations, such as the difference of minor allele, different minor allele frequency (MAF) and the complexity of the underlying genetic architecture [22]. The MAF of rs190982 on MEF2C is lower in Han Chinese populations than that in Caucasians (0.16% vs 0.42%) from the SNP web (http://www.ncbi.nlm.nih.gov/projects/SNP/snp_ref.cgi?rs=190982). Moreover, genotype distribution of the rs190982 was not in HWE in LOAD group, which may influence the results. In addition, MEF2C rs190982 polymorphism may have adverse effects on the development of AD that likely attribute to difference in sample sizes (Caucasian 74 046 vs. Chinese 4089) and unclear specific gene-gene or gene-environment interactions [23]. Furthermore, statistical deviation may have resulted from some other causes including various clinic characteristics of the samples, demographic variables, coexistence of some other unrealized neuropsychiatric change, experiment methods and statistical analyses [24]. Therefore, meta-analysis was performed in the total sample of 82507 individuals, Han Chinese and Caucasian to avoid these possible complex reasons and further explore the associations. The results revealed that rs190982 within MEF2C had a positive association with LOAD mainly in Caucasian residents. Although our study failed to replicate any association of the examined SNP with LOAD in the Chinese subgroup, we could not claim that other SNPs of MEF2C was not associated with LOAD. Additionally, the renewed genetic sequencing about MEF2C may be helpful to find new loci that related to LOAD in the near future.
In conclusion, our outcomes of meta-analysis did not detect any association of MEF2C gene rs190982 polymorphism with the risk of LOAD in the North Han Chinese. Further genetic analyses of this locus and the functional regions surrounding MEF2C are required to better elucidate the biochemical mechanisms and the interactions with AD. Additionally, larger independent replications both in Han Chinese and other ethnic groups are needed to elucidate the potential role of the locus at MEF2C in LOAD pathogenesis because of the genetic variations.
Materials and methods
Subjects
In our study, there are 2332 unrelated northern Han Chinese residents in origin from Shandong Province, comprising 984 sporadic LOAD patients and 1348 healthy controls. We screened the qualified AD patients in the Department of Neurology at Qingdao Municipal Hospital and some other 3A-level hospitals. All patients were undergone neuropsychological examination and structural neuroimaging (magnetic resonance imaging and/or brain computed tomography). At least two neurologists together defined a clinical diagnosis of AD based on the criteria of the National Institute of Neurological and Communicative Disorders and Stroke-Alzheimer’s Disease and Related Disorders Association (NINCDS/ADRDA) [25]. Never did their first-degree relatives have dementia in their family history. We obtained the information of patients such as age at onset and family history from caregivers. Healthy controls that were matched with patients for age and gender were recruited from the Health Examination Center of each collaborating hospital. Physicians and neurologists confirmed that control subjects were healthy and neurologically normal by following medical history, general examination, laboratory examination, and Mini Mental State Examination (score > 28). Subjects without significant illness such as type 2 diabetes mellitus (T2DM), heart disease, autoimmune disease, stroke, infectious disease, cancer, glaucoma, and atherosclerosis were considered as healthy and neurologically normal in our study [26]. Demographic details of the sample set are exhibited in Table 1. This study was performed after the informed consent of all subjects or legal guardians and with support from the Ethical Committee of Qingdao Municipal Hospital [27].
Genotyping
We used the Wizard genomic DNA purification kit (#A1125; Promega, USA) to extract genomic DNA from peripheral blood leukocytes of LOAD patients and healthy individuals in accordance with manufacturer’s protocol. MEF2C polymorphism (rs190982) was genotyped with the method of SNPscan™ kit. This is a patented SNP genotyping technology of Genesky Biotechnologies Inc., based on double ligation and multiplex fluorescent polymerase chain reaction (PCR). Genotyping of APOE (rs429358 and rs7412) polymorphisms was performed by the developed multiplex ligase detection reaction (iMLDR) via TaqMan genotyping assays on an ABI Prism 377 Sequence Detection System (Applied Biosystems), approved by the Shanghai Genesky Biotechnology Company. GeneMapper Software v4.1 (Applied Biosystems, USA) was used to analyze raw data and the ligation-PCR method was used to determine genotypes for each locus.
Statistical analysis
Statistical analysis was carried out with the use of SPSS16 software. Hardy-Weinberg equilibrium (HWE) test was performed before the association analysis. Differences in the characteristics of the study population between AD patients and controls were evaluated using Student’s t-test or the chi-square test. Genotypes and alleles were compared using the chi-square test or the Fisher’s exact test. Differences between cases and controls after stratification for APOE ε4 status were also examined by the chi-square test. We also performed logistic regression to further analyze the relation between the polymorphism and the LOAD risk adjusting for age, gender, and APOE ε4 status under various genetic models that were defined as 1 (GG+ AG) versus 0 (AA) for dominant, 1 (GG) versus 0 (AG + AA) for recessive, and 0 (AA) versus 1 (AG) versus 2 (GG) for additive. In addition, we also evaluated the effect of the interaction between rs190982 and APOE ε4 on the risk for LOAD in logistic regression models. The P value, odds ratios (ORs) and 95% confidence intervals (CIs) were computed to estimate the association between SNPs and AD. P < 0.05 was defined as the statistical significance level. Estimation of the statistical power was assessed with STPLAN 4.3 software.
In order to identify all articles that explored the association of the MEF2C rs190982 polymorphism with AD, we used the key search terms including “MEF2C,” “rs190982,” “Alzheimer’s disease,” and “AD,” combined with Boolean operators as appropriate to conduct a systematic literature search in PubMed, EMBASE, and the Cochrane library for publications up to December 2015. As to the limitation of language, only English publications were qualified. Additional studies were acquired from the reference lists of relevant primary articles. Finally, we adopted fixed-effects inverse variance-weighted methods to combine our results with the data from meta-analysis of 74,046 individuals [14] and other reports on the associations between MEF2C (rs190982) and LOAD [15-17] . At the same time, we generated I2 to estimate the possible influence of study heterogeneity on our results. Stata V.12.0 was used to perform all these analyses.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (81471309, 81371406, 81571245, and 81501103), the Shandong Provincial Outstanding Medical Academic Professional Program, Qingdao Key Health Discipline Development Fund, Qingdao Outstanding Health Professional Development Fund, and Shandong Provincial Collaborative Innovation Center for Neurodegenerative Disorders.
conflicts of interest
The authors disclose no conflicts of interest.
References
1. Mattson MP. Pathways towards and away from Alzheimer’s disease. Nature. 2004; 430:631-639.
2. Selkoe DJ. Alzheimer’s disease results from the cerebral accumulation and cytotoxicity of amyloid beta-protein. J Alzheimers Dis. 2001; 3:75-80.
3. Gatz M, Reynolds CA, Fratiglioni L, Johansson B, Mortimer JA, Berg S, Fiske A and Pedersen NL. Role of genes and environments for explaining Alzheimer disease. Arch Gen Psychiatry. 2006; 63:168-174.
4. Bertram L, Lill CM and Tanzi RE. The genetics of Alzheimer disease: back to the future. Neuron. 2010; 68:270-281.
5. Bertram L and Tanzi RE. Thirty years of Alzheimer’s disease genetics: the implications of systematic meta-analyses. Nat Rev Neurosci. 2008; 9:768-778.
6. Wang X, Lopez OL, Sweet RA, Becker JT, DeKosky ST, Barmada MM, Demirci FY and Kamboh MI. Genetic determinants of disease progression in Alzheimer’s disease. J Alzheimers Dis. 2015; 43:649-655.
7. Bertram L. Alzheimer’s genetics in the GWAS era: a continuing story of ‘replications and refutations’. Curr Neurol Neurosci Rep. 2011; 11:246-253.
8. Rashid AJ, Cole CJ and Josselyn SA. Emerging roles for MEF2 transcription factors in memory. Genes Brain Behav. 2014; 13:118-125.
9. Flavell SW, Cowan CW, Kim TK, Greer PL, Lin Y, Paradis S, Griffith EC, Hu LS, Chen C and Greenberg ME. Activity-dependent regulation of MEF2 transcription factors suppresses excitatory synapse number. Science. 2006; 311:1008-1012.
10. Cole CJ, Mercaldo V, Restivo L, Yiu AP, Sekeres MJ, Han JH, Vetere G, Pekar T, Ross PJ, Neve RL, Frankland PW and Josselyn SA. MEF2 negatively regulates learning-induced structural plasticity and memory formation. Nat Neurosci. 2012; 15:1255-1264.
11. Zhang ZG, Li Y, Ng CT and Song YQ. Inflammation in Alzheimer’s Disease and Molecular Genetics: Recent Update. Arch Immunol Ther Exp (Warsz). 2015; 63:333-344.
12. Burton TR, Dibrov A, Kashour T and Amara FM. Anti-apoptotic wild-type Alzheimer amyloid precursor protein signaling involves the p38 mitogen-activated protein kinase/MEF2 pathway. Brain Res Mol Brain Res. 2002; 108:102-120.
13. Camargo LM, Zhang XD, Loerch P, Caceres RM, Marine SD, Uva P, Ferrer M, de Rinaldis E, Stone DJ, Majercak J, Ray WJ, Yi-An C, Shearman MS, et al. Pathway-based analysis of genome-wide siRNA screens reveals the regulatory landscape of APP processing. PLoS One. 2015; 10:e0115369.
14. Lambert JC, Ibrahim-Verbaas CA, Harold D, Naj AC, Sims R, Bellenguez C, DeStafano AL, Bis JC, Beecham GW, Grenier-Boley B, Russo G, Thorton-Wells TA, Jones N, et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat Genet. 2013; 45:1452-1458.
15. Ruiz A, Heilmann S, Becker T, Hernandez I, Wagner H, Thelen M, Mauleon A, Rosende-Roca M, Bellenguez C, Bis JC, Harold D, Gerrish A, Sims R, et al. Follow-up of loci from the International Genomics of Alzheimer’s Disease Project identifies TRIP4 as a novel susceptibility gene. Transl Psychiatry. 2014; 4:e358.
16. Xiao Q, Liu ZJ, Tao S, Sun YM, Jiang D, Li HL, Chen H, Liu X, Lapin B, Wang CH, Zheng SL, Xu J and Wu ZY. Risk prediction for sporadic Alzheimer’s disease using genetic risk score in the Han Chinese population. Oncotarget. 2015; 6:36955-36964. doi: 10.18632/oncotarget.6271.
17. Jiao B, Liu X, Zhou L, Wang MH, Zhou Y, Xiao T, Zhang W, Sun R, Waye MM, Tang B and Shen L. Polygenic Analysis of Late-Onset Alzheimer’s Disease from Mainland China. PLoS One. 2015; 10:e0144898.
18. Potthoff MJ and Olson EN. MEF2: a central regulator of diverse developmental programs. Development. 2007; 134:4131-4140.
19. Lyons MR, Schwarz CM and West AE. Members of the myocyte enhancer factor 2 transcription factor family differentially regulate Bdnf transcription in response to neuronal depolarization. J Neurosci. 2012; 32:12780-12785.
20. Shepherd JD and Bear MF. New views of Arc, a master regulator of synaptic plasticity. Nat Neurosci. 2011; 14:279-284.
21. Greer PL, Zieg J and Greenberg ME. Activity-dependent transcription and disorders of human cognition. Am J Psychiatry. 2009; 166:14-15.
22. Greene CS, Penrod NM, Williams SM and Moore JH. Failure to replicate a genetic association may provide important clues about genetic architecture. PLoS One. 2009; 4:e5639.
23. Pearson TA and Manolio TA. How to interpret a genome-wide association study. JAMA. 2008; 299:1335-1344.
24. Yu JT, Song JH, Ma T, Zhang W, Yu NN, Xuan SY and Tan L. Genetic association of PICALM polymorphisms with Alzheimer’s disease in Han Chinese. J Neurol Sci. 2011; 300:78-80.
25. McKhann G, Drachman D, Folstein M, Katzman R, Price D and Stadlan EM. Clinical diagnosis of Alzheimer’s disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology. 1984; 34:939-944.
26. Yu JT, Miao D, Cui WZ, Ou JR, Tian Y, Wu ZC, Zhang W and Tan L. Common variants in toll-like receptor 4 confer susceptibility to Alzheimer’s disease in a Han Chinese population. Curr Alzheimer Res. 2012; 9:458-466.
27. Tan L, Yu JT, Zhang W, Wu ZC, Zhang Q, Liu QY, Wang W, Wang HF, Ma XY and Cui WZ. Association of GWAS-linked loci with late-onset Alzheimer’s disease in a northern Han Chinese population. Alzheimers Dement. 2013; 9:546-553.