
INTRODUCTION
Colorectal cancer (CRC) is the second leading cause of cancer death in developed countries [1]. It is a common complex human disease caused by the combination of genetic variants and environmental factors [2–3]. Large-scale genome-wide association studies (GWAS) and pathway analysis of GWAS datasets have been widely performed to identify common genetic variants or pathways contributing to human disease susceptibility [1, 4–15].
In 2008, Houlston et al. conducted a meta-analysis of two CRC GWAS including 13,315 individuals in discovery dataset and 27,418 subjects from eight independent case-control series in replication dataset [16]. They identified four new CRC risk variants at 14q22.2 (rs4444235, bone morphogenetic protein 4 (BMP4); P = 8.10E-10), 16q22.1 (rs9929218, cadherin 1 (CDH1); P = 1.20E-8), 19q13.1 (rrs9929218, rhophilin, Rho GTPase binding protein 2 (RHPN2); P = 4.60E-09) and 20p12.3 (rs961253; P = 2.00E-10) [16]. These findings underscore the value of large sample series for discovery and follow-up of genetic variants contributing to the etiology of CRC.
Followed the original CRC GWAS, several GWAS and candidate variant studies also investigate the association between rs9929218 and CRC in European, Asian and American populations [16–25]. However these studies reported consistent and inconsistent results. Evidence shows that rs9929218 variant may regulate different gene expressions in different human tissues, tumor tissues and normal tissues, and may have different regulatory mechanisms [26]. Here, we evaluated this association using large-scale samples from 16 studies (n=131768, 53656 CRC cases and 78112 controls) using meta-analysis method. To validate the effect of rs9929218 variant on CDH1 mRNA expression level, we further performed a functional analysis using two large-scale expression datasets.
RESULTS
Study selection
We selected 57 and 140 articles from PubMed and Google Scholar databases. Using the inclusion criteria and exclusion criteria, 11 articles including 16 independent studies were finally selected for our following analysis. More detailed information about the inclusion or exclusion of selected studies was described in Figure 1. The main characteristics of the included studies are described in Table 1.
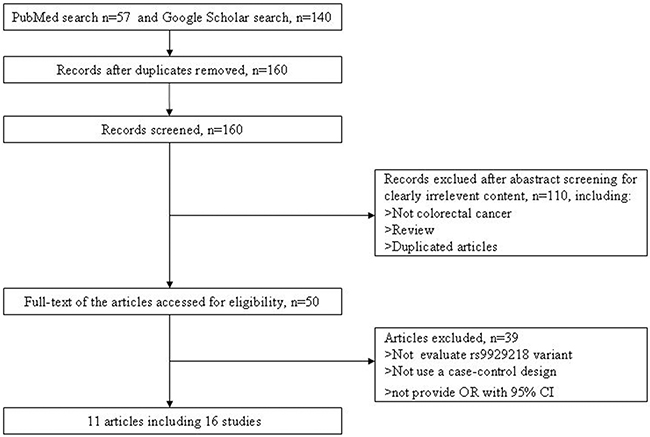
Figure 1: Flow diagram of selection of studies included in the current meta-analysis for the association between rs9929218 polymorphism and CRC.
Table 1: Main characteristics of selected studies in following meta-analysis
Study |
Year |
Ethnicity or country |
Population |
Case # |
Control # |
P value |
|---|---|---|---|---|---|---|
Ho [17] |
2011 |
China |
Asian |
892 |
890 |
0.965 |
Xiong [18] |
2010 |
China |
Asian |
2124 |
2124 |
0.14 |
Li [50] |
2012 |
China |
Asian |
229 |
267 |
0.484 |
Zhang [19] |
2014 |
China, South Korea and Japan |
Asian |
14963 |
31945 |
0.03 |
He [20] |
2011 |
European American |
American |
1171 |
1534 |
NA |
He [20] |
2011 |
African American |
American |
382 |
510 |
NA |
He [20] |
2011 |
Native Hawaiian |
American |
323 |
472 |
NA |
He [20] |
2011 |
Japanese Americans |
American |
1042 |
1426 |
NA |
He [20] |
2011 |
Latino |
American |
393 |
524 |
NA |
Hutter [21] |
2012 |
United States, Canada and Europe |
American |
7016 |
9723 |
0.01 |
Wang [22] |
2013 |
African American |
American |
1894 |
4703 |
0.12 |
Kupfer [23] |
2010 |
African American |
American |
795 |
985 |
0.44 |
Kupfer [23] |
2010 |
European American |
American |
399 |
367 |
0.10 |
Mates [25] |
2010 |
Romanian |
European |
92 |
96 |
0.303 |
Holst [24] |
2010 |
Swedish |
European |
1755 |
1691 |
0.051 |
Houlston [16] |
2008 |
United Kingdom |
European |
20186 |
20855 |
1.20E-08 |
n =131768 |
n=53656 |
n=78112 |
NA, not reported;
Heterogeneity test
We evaluated the genetic heterogeneity using Cochran’s Q statistic and I2 statistic, respectively. Using Cochran’s Q statistic, we identified no significant heterogeneity among these 16 studies with Cochran’s Q statistic = 17.62, and degrees of freedom = 15, and P=0.2829. We identified I2 = 14.9%, which indicates no statistically significant heterogeneity. We further performed a subgroup heterogeneity test in Asian, American and European populations, respectively. In the end, we found no significant genetic heterogeneity in Asian population with I2 = 0 and P=0.70, American population with I2 = 29% and P=0.19, and European population I2 = 11.2 and P=0.32.
Meta-analysis
Considering no significant heterogeneity in all the selected studies, we calculated the overall OR by the fixed effect model. The meta-analysis results showed significant association between rs9929218 and CRC (P=6.16E-21, OR=0.92, 95% CI 0.91-0.94). Detailed results are described in Figure 2.
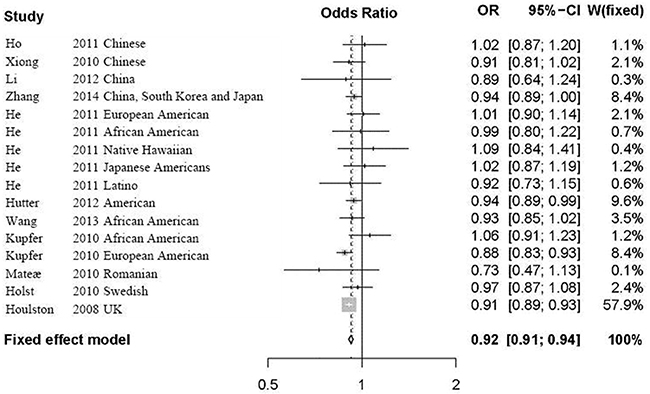
Figure 2: Forest plot for the meta-analysis of rs9929218 polymorphism using an additive model. According to Table 1, 11 articles including 16 independent studies were finally selected for our following analysis. In Figure 2, we list the name of the first author, the year of publication, the population or ethnicity, the OR with 95% CI and the weight in meta-analysis. The overall OR was calculated by the fixed effect model. OR, odds ratio; CI, confidence interval; fixed, fixed effect model.
We further performed a subgroup meta-analysis in Asian, American and European populations, respectively. The results further supported the significant association between rs9929218 and CRC in Asian population (P=0.019, OR=0.94, 95% CI 0.90-0.99), American population (P=5.9E-05, OR=0.94, 95% CI 0.91-0.97), and European population (P=8.91E-17, OR=0.91, 95% CI 0.90-0.93).
The sensitivity analysis showed that the association between rs9929218 and CRC did not vary substantially. We also excluded the most two large-scale studies from Houlston [16] and Zhang [19] at a time. We observed no heterogeneity in other studies with P=0.3517 and I2 = 9.2%, and identified significant association between rs9929218 and CRC with P=1.79E-05. Evidence from the linear regression test suggests no significant publication bias with P= 0.03689. The funnel plot is a symmetrical inverted funnel as described in Figure 3.
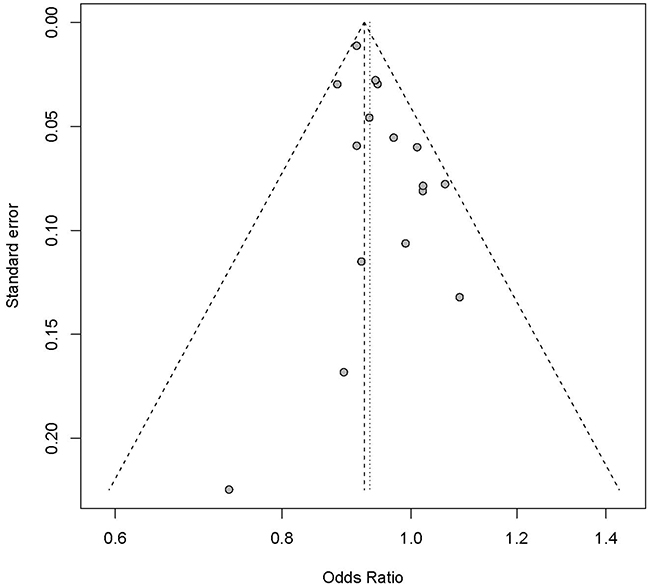
Figure 3: Funnel plot for publication bias analysis of the selected studies investigating the association between rs9929218 polymorphism and CRC. The X-axis stands for the ORs and the Y-axis is the standard error for each of the 16 studies. A linear regression based approach proposed by Egger et al. is used to evaluate the asymmetry of the funnel plot.
eQTL analysis
Using the eQTL dataset from the human peripheral blood, we identified that the rs9929218 minor allele (A) can significantly regulate ZFP90 expression with P=4.95E-152 by a cis-effect, and MCOLN2 expression with P=4.22E-06 by a trans-effect. Using GTEx, we further found that rs9929218 minor allele (A) can significantly regulate ZFP90 expression in the whole blood, Testis and Muscle-Skeletal tissues with P=6.60E-14, P=5.90E-06, and P=1.60E-05 by a cis-regulation. Meanwhile, rs9929218 can also regulate CDH1 expression in Nerve-Tibial and Spleen tissues with P=2.80E-11 and P=1.20E-06, respectively (Table 2)
Table 2: rs9929218 and gene expression in GTEx dataset
SNP |
Gene Symbol |
P value |
Effect size |
Tissue |
Cis or trans |
|---|---|---|---|---|---|
rs9929218 |
ZFP90 |
6.60E-14 |
-0.36 |
Whole Blood |
cis |
rs9929218 |
CDH1 |
2.80E-11 |
0.22 |
Nerve - Tibial |
cis |
rs9929218 |
CDH1 |
1.20E-06 |
-0.66 |
Spleen |
cis |
rs9929218 |
ZFP90 |
5.90E-06 |
0.46 |
Testis |
cis |
rs9929218 |
RP11-354M1.2 |
8.90E-06 |
-0.62 |
Spleen |
cis |
rs9929218 |
ZFP90 |
1.60E-05 |
-0.26 |
Muscle - Skeletal |
cis |
Effect size >0 and <0 mean that this SNP allele regulated increased and reduced gene expression, respectively.
DISCUSSION
GWAS reported rs9929218 to be significantly associated with CRC in European ancestry [16]. Following studies investigated this association and reported inconsistent results. Here, using a comprehensive search in the PubMed and Google Scholar, we reevaluated this association using large-scale samples from 16 studies. Using two genetic heterogeneity test methods Cochran’s Q-statistic and I2statistic, we did not identify significant heterogeneity among these 16 studies. Using a fixed effect model for meta-analysis to calculate the overall OR, we identified significant association between rs9929218 and CRC (P=6.16E-21, OR=0.92, 95% CI 0.91-0.94). The sensitivity analysis and publication bias further supported our results to be robust. After excluding the two large-scale studies from Houlston [16] and Zhang [19], we still identified significant association between rs9929218 and CRC.
Evidence shows that eQTL may modify gene expression and influence risk for human diseases [27–29]. In order to validate the effect of rs9929218 variant on CDH1 mRNA expression level, we performed eQTLs analysis using a large-scale dataset from the peripheral blood samples and multiple human tissues from GTEx. We identified significant regulation relations between rs9929218 variant and gene expression including CDH1, ZFP90, RP11-354M1.2 and MCOLN2 by both cis-effect and trans-effect. Recently, Closa et al. analyzed the association between genotypes for 26 GWAS SNPs and the expression of genes within a 2 Mb region (cis-eQTLs) using Affymetrix Human Genome U219 expression arrays with 47 samples from healthy colonic mucosa and 97 samples from normal mucosa adjacent to colon cancer [26]. In all the samples, the cis-eQTL analysis showed significant association between rs9929218 variant and GFOD2 expression with P=3.90E-03 [26]. Closa et al. further reported significant association of rs9929218 variant with expression of TSNAXIP1, HSF4 and NUTF2 in tumor tissue eQTL analysis with P=5.50E-03, P=6.70E-03 and P=1.70E-03, respectively, but not in normal tissue eQTL analysis with P=0.59, P=0.54 and P=0.27, respectively [26].
It is reported that the rs9929218 variant and rs1862748 are in linkage disequilibrium (LD) with rs28626308, which results in a non-synonymous change in RHPN2 protein [30]. In addition to rs9929218 variant, another two CDH1 variants rs4939827 and rs12953717 are in LD with rs7199991, which is genetically associated with four expression quantitative trait loci that correlate with expression of the upstream gene ZFP90 [30]. The rs9929218 is located at 16q22.1 (CDH1/ CDH3). Carvajal-Carmona et al. conducted a fine-mapping of CRC susceptibility loci at 8q23.3, 16q22.1 and 19q13.11 to refine the association signals [31]. Their findings suggest that ZFP90, rather than CDH1 or CDH3, is the most likely target of the 16q22.1 genetic variation associated with increased CRC risk [31].
Here, we reinvestigated the association between rs9929218 and CRC using an additive model as descried in Figure 2. It is reported that this model performs well when the true underlying genetic model is uncertain [32]. Analyzing the association between rs9929218 and CRC using dominant and recessive models is also required. However, the original genotype data are not publicly available. Future studies with genotype data are required to replicate our findings. In summary, our findings show that rs9929218 polymorphism contributes to CRC susceptibility.
METHODS AND MATERIALS
Search strategy
We searched the PubMed database to select all possible studies with the combined key words including ‘CDH1’, ‘rs9929218’, ‘polymorphism’, or ‘variant’ and ‘colorectal cancer’, ‘colon cancer’, ‘rectal cancer’, or ‘bowel cancer’. We also used the Google Scholar to query the articles citing CRC GWAS and select all associated publications. There was no language and sample limitation, and the literature search was updated on October 28, 2015. This search was performed by two independent individuals to identify potential publications meeting our inclusion criteria (see below), with differences resolved by discussion with a third reviewer. More detailed information about the search strategy is described in previous studies [33–35].
Inclusion and exclusion criteria
The following inclusion criteria were used to select all potential publications for the meta-analysis: (1) use a case-control design; (2) evaluate the association between rs9929218 and CRC; (3) provide odds ratio (OR) with 95% confidence interval (CI) for allele model (see below); or (4) provide sufficient data to calculate the OR and 95% CI for allele model; For any publication, it is excluded from the meta-analysis, if (1) not for cancer research; (2) not use a case-control design including case population; case reports, conference abstract, reviews; (3) not provide OR with 95% CI for allele model (see below); or (4) not provide sufficient data to calculate the OR and 95% CI for allele model; More detailed information is described in previous studies [33–35].
Data extraction
Two authors reviewed and extracted information from all eligible publications independently, according to the inclusion and exclusion criteria listed above. The following items were extracted including (1) the name of the first author; (2) the year of publication; (3) the population and ethnicity; (4) the numbers of CRC cases and controls; (5) the OR with 95% CI or (6) to calculate the OR and 95% CI; If there is a conflict, both reviewers discuss with third reviewer and reach an agreement. For rs9929218 polymorphism A/G alleles, we selected the additive genetic model: A allele versus G allele for further meta-analysis [36–40].
Heterogeneity test
Genetic heterogeneity is evaluated using two methods including Cochran’s Q-statistic and statistic [41–46]. Cochran’s Q statistic approximately follows a χ2 distribution with k-1 degrees of freedom (k stands for the number of studies for analysis) [41–46]. A significant Q-statistic (P < 0.01) indicates heterogeneity among selected studies. I2 is a measure of heterogeneity and a statistic that indicates the percentage of variance in a meta-analysis that is attributable to study heterogeneity [41–46]. The intervals including 0-25%, 25-50%, 50-75% and 75-100%, represent the low, moderate, large and extreme heterogeneity [41–46]. The interval I2 > 50% indicates statistically significant heterogeneity.
Statistical analysis
Here, we used two kinds of meta-analysis models including the fixed effect model (Mantel-Haenszel) and random-effect model (DerSimonian-Laird) to calculate the pooled OR. If there is no significant heterogeneity among the included studies, the pooled OR is calculated by the fixed effect model; otherwise the OR is calculated by random-effect model [41–46]. Z test is used to determine the significance of OR. R Package is used to compute all statistical tests [41–46]. A significant levels (P < 0.01) indicates significant association between rs9929218 and CRC. Using sensitivity analysis, each study is omitted at a time to assess the influence of single study on the association between rrs9929218 and CRC. A funnel plot proposed by Egger et al. is used to investigate potential publication bias [41–46]. A linear regression based approach proposed by Egger et al. is used to test for publication bias, and provide statistical evidence to evaluate the asymmetry of the funnel plot to. with the P < 0.01 indicating that there was a significant publication bias [47]. All statistical analyses were performed using R Package [41–46].
eQTL analysis in human peripheral blood
To validate the effect of rs9929218 variant on CDH1 mRNA expression level, we performed an expression quantitative trait loci (eQTL) analysis using a large-scale eQTL dataset, which is from a meta-analysis in non-transformed peripheral blood samples from 5,311 individuals with replication in 2,775 individuals [48].
eQTLs analysis using Genotype-Tissue Expression
We also investigate the effect of rs9929218 variant on CDH1 mRNA expression using the Genotype-Tissue Expression (GTEx) project (http://www.gtexportal.org/home/), which provides a scientific resource to study human gene expression and regulation and its relationship to genetic variation [49]. Here, we selected all the human tissues with the number of genotyped samples > 60 in the following eQTL analysis [49].
ACKNOWLEDGMENTS
This work was supported by funding from the National Nature Science Foundation of China (Grant No. 81300945), Heilongjiang Postdoctoral Financial Assistance (Grant No. LBH-Z15141), and Health and Family Planning Commission of Heilongjiang Province Research Projects (Grant No. 2014-349).
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
REFERENCES
1. Quan B, Qi X, Yu Z, Jiang Y, Liao M, Wang G, Feng R, Zhang L, Chen Z, Jiang Q and Liu G. Pathway analysis of genome-wide association study and transcriptome data highlights new biological pathways in colorectal cancer. Mol Genet Genomics. 2015; 290:603-610.
2. Liao M, Wang G, Quan B, Qi X, Yu Z, Feng R, Zhang L, Jiang Y, Zhang Y and Liu G. Analyzing large-scale samples confirms the association between rs16892766 polymorphism and colorectal cancer susceptibility. Sci Rep. 2015; 5:7957.
3. He D, Ma L, Feng R, Zhang L, Jiang Y, Zhang Y and Liu G. Analyzing large-scale samples highlights significant association between rs10411210 polymorphism and colorectal cancer. Biomed Pharmacother. 2015; 74:164-168.
4. Liu G, Jiang Y, Wang P, Feng R, Jiang N, Chen X, Song H and Chen Z. Cell adhesion molecules contribute to Alzheimer’s disease: multiple pathway analyses of two genome-wide association studies. J Neurochem. 2012; 120:190-198.
5. Liu G, Jiang Y, Chen X, Zhang R, Ma G, Feng R, Zhang L, Liao M, Miao Y, Chen Z, Zeng R and Li K. Measles contributes to rheumatoid arthritis: evidence from pathway and network analyses of genome-wide association studies. PLoS One. 2013; 8:e75951.
6. Liu G, Yao L, Liu J, Jiang Y, Ma G, Chen Z, Zhao B and Li K. Cardiovascular disease contributes to Alzheimer’s disease: evidence from large-scale genome-wide association studies. Neurobiol Aging. 2014; 35:786-792.
7. Shang H, Liu G, Jiang Y, Fu J, Zhang B, Song R and Wang W. Pathway analysis of two amyotrophic lateral sclerosis GWAS highlights shared genetic signals with Alzheimer’s disease and Parkinson’s disease. Mol Neurobiol. 2015; 51:361-369.
8. Xiang Z, Xu M, Liao M, Jiang Y, Jiang Q, Feng R, Zhang L, Ma G, Wang G, Chen Z, Zhao B, Sun T, Li K and Liu G. Integrating Genome-Wide Association Study and Brain Expression Data Highlights Cell Adhesion Molecules and Purine Metabolism in Alzheimer’s Disease. Mol Neurobiol. 2015; 52:514-521.
9. Zhao X, Gu J, Li M, Xi J, Sun W, Song G and Liu G. Pathway analysis of body mass index genome-wide association study highlights risk pathways in cardiovascular disease. Sci Rep. 2015; 5:13025.
10. Jiang Q, Jin S, Jiang Y, Liao M, Feng R, Zhang L, Liu G and Hao J. Alzheimer’s Disease Variants with the Genome-Wide Significance are Significantly Enriched in Immune Pathways and Active in Immune Cells. Mol Neurobiol. 2016.
11. Wei J, Li M, Gao F, Zeng R, Liu G and Li K. Multiple analyses of large-scale genome-wide association study highlight new risk pathways in lumbar spine bone mineral density. Oncotarget. 2016; 7:31429-39. doi: 10.18632/oncotarget.8948.
12. Liu G and Li K. CHCHD2 and Parkinson’s disease. Lancet Neurol. 2015; 14:679-680.
13. Li K, Jiang Q, Xu A and Liu G. REST rs3796529 variant does not confer susceptibility to Alzheimer’s disease. Ann Neurol. 2015; 78:835-836.
14. Jiang Q and Liu G. REST rs3796529 variant does not influence human subcortical brain structures. Ann Neurol. 2016; 79:334-335.
15. Liu G. No association of TREM1 rs6910730 and TREM2 rs7759295 with Alzheimer disease. Ann Neurol. 2015; 78:659.
16. Houlston RS, Webb E, Broderick P, Pittman AM, Di Bernardo MC, Lubbe S, Chandler I, Vijayakrishnan J, Sullivan K, Penegar S, Carvajal-Carmona L, Howarth K, Jaeger E, et al. Meta-analysis of genome-wide association data identifies four new susceptibility loci for colorectal cancer. Nat Genet. 2008; 40:1426-1435.
17. Ho JW, Choi SC, Lee YF, Hui TC, Cherny SS, Garcia-Barcelo MM, Carvajal-Carmona L, Liu R, To SH, Yau TK, Chung CC, Yau CC, Hui SM, et al. Replication study of SNP associations for colorectal cancer in Hong Kong Chinese. Br J Cancer. 2011; 104:369-375.
18. Xiong F, Wu C, Bi X, Yu D, Huang L, Xu J, Zhang T, Zhai K, Chang J, Tan W, Cai J and Lin D. Risk of genome-wide association study-identified genetic variants for colorectal cancer in a Chinese population. Cancer Epidemiol Biomarkers Prev. 2010; 19:1855-1861.
19. Zhang B, Jia WH, Matsuda K, Kweon SS, Matsuo K, Xiang YB, Shin A, Jee SH, Kim DH, Cai Q, Long J, Shi J, Wen W, et al. Large-scale genetic study in East Asians identifies six new loci associated with colorectal cancer risk. Nat Genet. 2014; 46:533-542.
20. He J, Wilkens LR, Stram DO, Kolonel LN, Henderson BE, Wu AH, Le Marchand L and Haiman CA. Generalizability and epidemiologic characterization of eleven colorectal cancer GWAS hits in multiple populations. Cancer Epidemiol Biomarkers Prev. 2011; 20:70-81.
21. Hutter CM, Chang-Claude J, Slattery ML, Pflugeisen BM, Lin Y, Duggan D, Nan H, Lemire M, Rangrej J, Figueiredo JC, Jiao S, Harrison TA, Liu Y, et al. Characterization of gene-environment interactions for colorectal cancer susceptibility loci. Cancer Res. 2012; 72:2036-2044.
22. Wang H, Haiman CA, Burnett T, Fortini BK, Kolonel LN, Henderson BE, Signorello LB, Blot WJ, Keku TO, Berndt SI, Newcomb PA, Pande M, Amos CI, et al. Fine-mapping of genome-wide association study-identified risk loci for colorectal cancer in African Americans. Hum Mol Genet. 2013; 22:5048-5055.
23. Kupfer SS, Anderson JR, Hooker S, Skol A, Kittles RA, Keku TO, Sandler RS and Ellis NA. Genetic heterogeneity in colorectal cancer associations between African and European americans. Gastroenterology. 2010; 139:1677-1685, 1685 e1671-1678.
24. von Holst S, Picelli S, Edler D, Lenander C, Dalen J, Hjern F, Lundqvist N, Lindforss U, Pahlman L, Smedh K, Tornqvist A, Holm J, Janson M, et al. Association studies on 11 published colorectal cancer risk loci. Br J Cancer. 2010; 103:575-580.
25. Mates IN, Csiki I, Mates D, Constantinescu V, Badea P, Dinu D, Constantin A and Constantinoiu S. Association of common genetic variants with colorectal cancer risk in a Romanian sample. Chirurgia (Bucur). 2010; 105:749-757.
26. Closa A, Cordero D, Sanz-Pamplona R, Sole X, Crous-Bou M, Pare-Brunet L, Berenguer A, Guino E, Lopez-Doriga A, Guardiola J, Biondo S, Salazar R and Moreno V. Identification of candidate susceptibility genes for colorectal cancer through eQTL analysis. Carcinogenesis. 2014; 35:2039-2046.
27. Liu G, Bao X and Wang R. Expression quantitative trait loci regulate HNF4A and PTBP1 expression in human brains. Proc Natl Acad Sci U S A. 2015; 112:E3975.
28. Bao X, Liu G, Jiang Y, Jiang Q, Liao M, Feng R, Zhang L, Ma G, Zhang S, Chen Z, Zhao B, Wang R and Li K. Cell adhesion molecule pathway genes are regulated by cis-regulatory SNPs and show significantly altered expression in Alzheimer’s disease brains. Neurobiol Aging. 2015; 36:2904 e2901-2907.
29. Liu G, Liu Y, Jiang Q, Jiang Y, Feng R, Zhang L, Chen Z, Li K and Liu J. Convergent Genetic and Expression Datasets Highlight TREM2 in Parkinson’s Disease Susceptibility. Mol Neurobiol. 2015.
30. Kupfer SS, Skol AD, Hong E, Ludvik A, Kittles RA, Keku TO, Sandler RS and Ellis NA. Shared and independent colorectal cancer risk alleles in TGFbeta-related genes in African and European Americans. Carcinogenesis. 2014; 35:2025-2030.
31. Carvajal-Carmona LG, Cazier JB, Jones AM, Howarth K, Broderick P, Pittman A, Dobbins S, Tenesa A, Farrington S, Prendergast J, Theodoratou E, Barnetson R, Conti D, et al. Fine-mapping of colorectal cancer susceptibility loci at 8q23.3, 16q22.1 and 19q13.11: refinement of association signals and use of in silico analysis to suggest functional variation and unexpected candidate target genes. Hum Mol Genet. 2011; 20:2879-2888.
32. Gogele M, Minelli C, Thakkinstian A, Yurkiewich A, Pattaro C, Pramstaller PP, Little J, Attia J and Thompson JR. Methods for meta-analyses of genome-wide association studies: critical assessment of empirical evidence. Am J Epidemiol. 2012; 175:739-749.
33. Liu G, Zhang L, Feng R, Liao M, Jiang Y, Chen Z, Zhao B and Li K. Lack of association between PICALM rs3851179 polymorphism and Alzheimer’s disease in Chinese population and APOEepsilon4-negative subgroup. Neurobiol Aging. 2013; 34:1310 e1319-1310.
34. Liu G, Zhang S, Cai Z, Ma G, Zhang L, Jiang Y, Feng R, Liao M, Chen Z, Zhao B and Li K. PICALM gene rs3851179 polymorphism contributes to Alzheimer’s disease in an Asian population. Neuromolecular Med. 2013; 15:384-388.
35. Liu G, Xu Y, Jiang Y, Zhang L, Feng R and Jiang Q. PICALM rs3851179 Variant Confers Susceptibility to Alzheimer’s Disease in Chinese Population. Mol Neurobiol. 2016.
36. Liu G, Zhang S, Cai Z, Li Y, Cui L, Ma G, Jiang Y, Zhang L, Feng R, Liao M, Chen Z, Zhao B and Li K. BIN1 gene rs744373 polymorphism contributes to Alzheimer’s disease in East Asian population. Neurosci Lett. 2013; 544:47-51.
37. Liu G, Wang H, Liu J, Li J, Li H, Ma G, Jiang Y, Chen Z, Zhao B and Li K. The CLU gene rs11136000 variant is significantly associated with Alzheimer’s disease in Caucasian and Asian populations. Neuromolecular Med. 2014; 16:52-60.
38. Liu G, Li F, Zhang S, Jiang Y, Ma G, Shang H, Liu J, Feng R, Zhang L, Liao M, Zhao B and Li K. Analyzing large-scale samples confirms the association between the ABCA7 rs3764650 polymorphism and Alzheimer’s disease susceptibility. Mol Neurobiol. 2014; 50:757-764.
39. Shen N, Chen B, Jiang Y, Feng R, Liao M, Zhang L, Li F, Ma G, Chen Z, Zhao B, Li K and Liu G. An Updated Analysis with 85,939 Samples Confirms the Association Between CR1 rs6656401 Polymorphism and Alzheimer’s Disease. Mol Neurobiol. 2015; 51:1017-1023.
40. Zhang S, Zhang D, Jiang Y, Wu L, Shang H, Liu J, Feng R, Liao M, Zhang L, Liu Y, Liu G and Li K. CLU rs2279590 polymorphism contributes to Alzheimer’s disease susceptibility in Caucasian and Asian populations. J Neural Transm (Vienna). 2015; 122:433-439.
41. Chen H, Wu G, Jiang Y, Feng R, Liao M, Zhang L, Ma G, Chen Z, Zhao B, Li K, Yu C and Liu G. Analyzing 54,936 Samples Supports the Association Between CD2AP rs9349407 Polymorphism and Alzheimer’s Disease Susceptibility. Mol Neurobiol. 2015; 52:1-7.
42. Li X, Shen N, Zhang S, Liu J, Jiang Q, Liao M, Feng R, Zhang L, Wang G, Ma G, Zhou H, Chen Z, Jiang Y, Zhao B, Li K and Liu G. CD33 rs3865444 Polymorphism Contributes to Alzheimer’s Disease Susceptibility in Chinese, European, and North American Populations. Mol Neurobiol. 2015; 52:414-421.
43. Zhang S, Li X, Ma G, Jiang Y, Liao M, Feng R, Zhang L, Liu J, Wang G, Zhao B, Jiang Q, Li K and Liu G. CLU rs9331888 Polymorphism Contributes to Alzheimer’s Disease Susceptibility in Caucasian But Not East Asian Populations. Mol Neurobiol. 2016; 53:1446-1451.
44. Chen G, Fu X, Wang G, Liu G and Bai X. Genetic Variant rs10757278 on Chromosome 9p21 Contributes to Myocardial Infarction Susceptibility. Int J Mol Sci. 2015; 16:11678-11688.
45. Li Y, Song D, Jiang Y, Wang J, Feng R, Zhang L, Wang G, Chen Z, Wang R, Jiang Q and Liu G. CR1 rs3818361 Polymorphism Contributes to Alzheimer’s Disease Susceptibility in Chinese Population. Mol Neurobiol. 2015.
46. Liu G and Jiang Q. Alzheimer’s disease CD33 rs3865444 variant does not contribute to cognitive performance. Proc Natl Acad Sci U S A. 2016; 113:E1589-1590.
47. Song F, Khan KS, Dinnes J and Sutton AJ. Asymmetric funnel plots and publication bias in meta-analyses of diagnostic accuracy. Int J Epidemiol. 2002; 31:88-95.
48. Westra HJ, Peters MJ, Esko T, Yaghootkar H, Schurmann C, Kettunen J, Christiansen MW, Fairfax BP, Schramm K, Powell JE, Zhernakova A, Zhernakova DV, Veldink JH, et al. Systematic identification of trans eQTLs as putative drivers of known disease associations. Nat Genet. 2013; 45:1238-1243.
49. Human genomics. The Genotype-Tissue Expression (GTEx) pilot analysis: multitissue gene regulation in humans. Science. 2015; 348:648-660.
50. Li FX, Yang XX, Hu NY, Du HY, Ma Q and Li M. Single-nucleotide polymorphism associations for colorectal cancer in southern chinese population. Chin J Cancer Res. 2012; 24:29-35.