
INTRODUCTION
Styrene is a widely used material in production of plastic, rubber, fiberglass and etc. Occupational populations may be exposed to higher concentration of styrene, especially in the reinforced plastic industry [1, 2]. On the other hand, public exposure to this chemical may also occur due to mainstream cigarette smoke, engine exhausts, carpets and food packaging [3–6]. Inhalation is a major route for both occupational and environmental exposure to styrene [7].
Over the decades, many efforts have been made to study styrene-induced adverse health effects. In some review articles [6, 8, 9], convincing evidences have been shown that styrene induces lung cancer in mice, while evidence in humans is unclear. Many studies, but not all, have shown that styrene is genotoxic. International Agency for Research on Cancer (IARC) has classified styrene in group 2B as a possible human carcinogen, and its metabolite, styrene-7,8-oxide (SO), has been classified in group 2A, probably carcinogenic to humans [10].
Genetic variation appeared to be an important reason for the uncertainty of styrene-induced toxicity. So far the polymorphisms of some enzymes involved in styrene metabolism, such as P-450 cytochromes, microsomal epoxide hydrolase (EXPH), and glutathione S-transferases (GSTs), have been studied and reviewed for the impacts on genotoxic risk of styrene-exposed populations [9]. In general, these polymorphisms might modify individual susceptibility to styrene-induced DNA damage. But it was difficult to draw any conclusion on possible associations between genes polymorphisms and biomarkers of styrene-induced DNA damage, as most studies evaluated complex gene-gene and gene-environment interactions with small samples.
It is well-known that human aldehyde dehydrogenases (ALDHs) are involved in the metabolic pathways of endogenous and exogenous compounds. In addition, ALDH activity has been reported to represent a functional marker for cervical cancer stem cells as well as a target for novel cervical cancer therapies [11] . In the process of styrene metabolism, ALDHs participate in the formation of mandelic acid (MA) and phenylglyoxylic acid (PGA) (Figure 1), which account for more than 95% of the urinary metabolites of styrene [9]. So far, however, no data are available regarding the effects of ALDHs polymorphisms on metabolism and toxicity of styrene. In particular, deficient ALDH2 activity caused by mutant allele (ALDH2 *2) [12] may dramatically change the metabolism of styrene and its intermediates, which may in turn lead to different toxic effects among individuals exposed to styrene. The frequencies of ALDH2 genotypes vary in different ethnical groups, and the variant allele ALDH2 *2 is highly prevalent (about 30–50%) in East Asian populations, while almost all Caucasians carried the functional ALDH2 *1/*1 genotype [13–15]. Previous studies have indicated that ALDH2 *2 allele increased the risk for alcohol-related cancers [12, 16]. Moreover, the ethnical difference in ALDH2 polymorphisms could significantly influence the levels of biological exposure indices of organic solvent in urinary and blood, such as toluene, perchloroethylene and methyl ethyl ketone [17–19]. As a result, Orientals tend to have lower concentrations of urinary PGA than Caucasian under inhalational exposure to styrene [20]. Taken together, the valuable information above prompted us to examine whether ALDH2 polymorphisms exert any modifying effect on the metabolic processes and genotoxic effects of styrene.
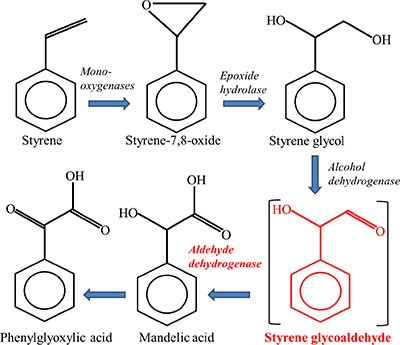
Figure 1: Major pathway of styrene metabolism.
In this study, we recruited a relative large sample of 329 workers occupationally exposed to styrene and 152 unexposed controls. A combination of three genotoxic biomarkers, DNA strand breaks, DNA-base oxidation in leukocytes, and urinary 8-OH-dG, were examined for accurate assessment of DNA damage related to styrene. Urinary MA+PGA concentrations and styrene concentration in air were used to characterize styrene exposure level.
RESULTS
Study subjects and styrene exposure
The exposed subjects were divided into three groups by workplaces. No significant differences were found between controls and exposed subjects in age, gender, smoking or drinking rate (Table 1). The mean styrene concentrations in air at workplace A, B and C were 26.21 ± 14.18, 36.75 ± 19.27 and 57.17 ± 31.01 ppm, respectively. And the total mean concentration was 40.04 ± 25.43 ppm. Meanwhile, the highest mean value of urinary MA+PGA (119.64 ± 127.18 mg/g creatinine) was found in workplace C, and the total mean value in three workplaces was 91.56 ± 146.55 mg/g creatinine.
Table 1: Characteristics of the study subjects and characterization of styrene exposure
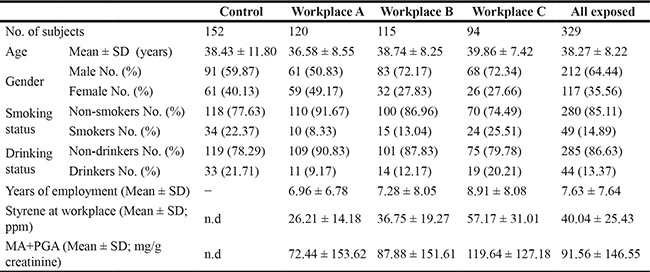
Note: MA+ PGA: mandelic acid + phenylglyoxylic acid; n.d.: not detectable.
Biomarkers for DNA damage related to styrene exposure
The levels of DNA strand breaks, DNA- base oxidation in leukocytes, and urinary 8-OH-dG concentrations were presented in Table 2. The mean level of DNA strand breaks in leukocytes, measured by a standard comet assay, was significantly higher in exposed workers (workplace A: 12.85 ± 3.75, workplace B: 13.02 ± 3.18, workplace C: 13.99 ± 2.92, and all exposed: 13.24 ± 3.36) than in controls (11.71 ± 3.88). As well, using Fpg-modified comet assay, we detected higher DNA-base oxidation in leukocytes among exposure groups as compared to control group. Furthermore, urinary 8-OH-dG, representing the whole body oxidative DNA damage, also showed significantly higher levels in various exposed groups (workplace A: 4.51 ± 1.61, workplace B: 4.72 ± 2.01, workplace C: 5.96 ± 2.08, and all exposed: 5.00 ± 1.99) than in controls (3.95 ± 1.42). These results collectively suggested the positive correlation between genetic damage and exposure level of styrene.
Table 2: DNA damage of leukocytes and biomarker of urinary nuclei acid oxidation in controls and styrene-exposed workers
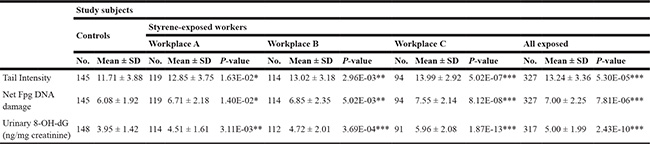
*p < 0.05, **p < 0.01, ***p < 0.001, based on independent samples t - test, and compared with the corresponding controls.
Association between ALDH2 polymorphisms and the effect of styrene exposure
The distribution of genotypes of ALDH2 and other enzyme genes was summarized in Table S1. As shown in Table 3, compared to wild-type genotype ALDH2 *1/*1, a significant lower mean value of PGA+MA excretion was found in the exposed individuals (all workplaces combined) carrying the variant allele ALDH2 *2. In addition, this significant difference could still be found in respective workplaces representing relatively small size of samples.
Table 3: Effects of ALDH2 polymorphisms on urinary excretion of styrene specific metabolites and the levels of various parameters of genetic damage in control and styrene-exposed workers
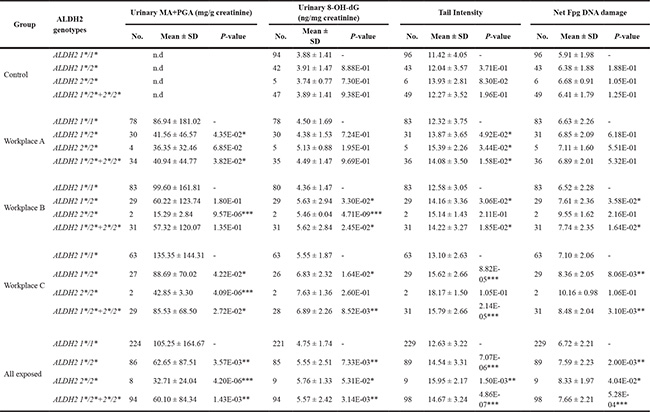
*p < 0.05, **p < 0.01, ***p < 0.001, based on independent samples t - test, and compared with the corresponding wild-type genotype. Data are expressed as mean ± S.E. n.d.: not detectable.
As for styrene-induced DNA damage, exposed workers with ALDH2 *2 allele had significantly higher levels of DNA strand breaks, DNA-base oxidation in leukocytes and urinary 8-OH-dG than individuals of wild-type genotype. When checking each workplace, significant effects of ALDH2 *2 allele were found in workplace B and C, representing middle and high level exposure. On the other hand, in workplace A with low level exposure, only DNA-base oxidation level was significantly high in ALDH2 *2 allele carriers, but not DNA strand breaks and urinary 8-OH-dG. These results were further confirmed by multivariate linear regression models including sex, age, years of employment, smoking and alcohol drinking (Table 4). In contrast, ALDH2 polymorphisms did not influence DNA damage in controls.
Table 4: Multivariate linear regression models evaluating the relationship between ALDH2 genotype and the levels of styrene specific metabolites and various parameters of genetic damage
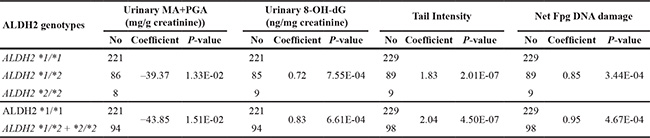
The linear model was adjusted for sex, age, years of employment, smoking and alcohol drinking, thus determining the regression coefficient and significance level (*p < 0.05, **p < 0.01, ***p < 0.001). The samples with missing values regarding any variable in the linear model were excluded.
Association between other genetic polymorphisms and biomarkers of exposure and genotoxic effect
Table 5 showed that none of three biomarkers representing genotoxic endpoints was significantly affected by CYP2E1, GSTM1, GSTT1 or EPHX1 polymorphisms in all styrene-exposed workers. Only the variant homozygous CYP2E1 Rsal c2/c2 and Dral C/C were found to be associated with lower concentration of urinary MA+PGA. We also examined these genetic polymorphisms for each workplace as well as controls (Tables S2–S5). Significant signals were observed only in workplace B, but not in workplace A, C and controls.
Table 5: Effects of GSTM1, GSTT1, EPHX1 and CYP2E1 polymorphisms on urinary excretion of styrene specific metabolites and the levels of various parameters of genetic damage in all styrene-exposed workers
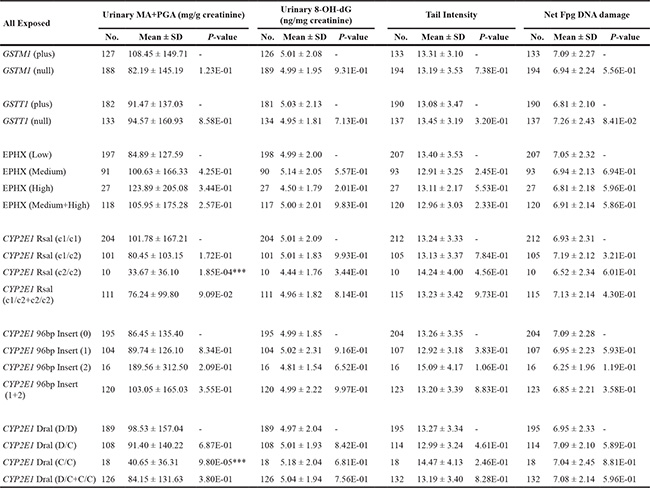
***p < 0.001, based on independent samples t - test, and compared with the corresponding wild-type genotype.
The above results suggested a prominent role of ALDH2 polymorphisms, which might overshadow the effects of other genetic polymorphisms on styrene metabolism and styrene-related DNA damage. So we further stratified the exposed samples by focusing on the wild-type genotype ALDH2 *1/*1 and re-analyzed the effects of other genetic polymorphisms (Table 6). For example, CYP2E1 Rsal variant homozygous c2/c2 and heterozygous c1/c2 genotypes exhibited significantly lower mean level of urinary MA+PGA than wild-type c1/ c2 genotypes. Also, GSTM1 null individuals had lower MA+PGA concentration than GSTM1 plus ones. When compared with respective wild-type genotypes, CYP2E1 Rsal and 96bp insert variant homozygous genotypes were found to be related with decreased DNA-base oxidation in leukocytes.
Table 6: Analysis of the influence of GSTM1, GSTT1, EPHX1 and CYP2E1 polymorphisms on urinary excretion of styrene specific metabolites and the levels of various parameters of genetic damage in exposed workers carrying ALDH2 *1/*1
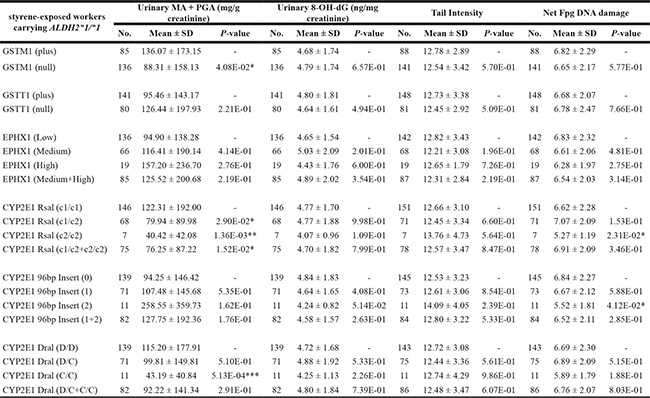
*p < 0.05, **p < 0.01, ***p < 0.001, based on independent samples t - test, and compared with the corresponding wild-type genotype.
DISCUSSION
In this study, we investigated the urinary excretion of styrene metabolites and various parameters of DNA damage, and analyzed if the genetic polymorphisms of styrene-metabolizing enzymes exert any modification. Considering the confusing results about the role of genetic polymorphisms as reported, we felt that a large number of styrene-exposed workers and comparable controls should be necessary to improve the statistical power. To our best knowledge, this is the largest sample size so far used for studying the functional relevance of these polymorphisms in the occupational exposure of styrene.
One study [21] proposed that styrene exposure may increase oxidative stress and in turn result in the oxidative DNA damage. Using Fpg-modified comet assay, we found that DNA-base oxidation in leukocytes was induced by styrene exposure. The result was consistent with prior studies [22, 23], which demonstrated that occupational exposure to styrene in workers significantly increased the levels of leukocyte 8-OH-dG.
Instead of 8-OH-dG in leukocytes, we analyzed urinary 8-OH-dG to evaluate the oxidative stress of whole body. The results were in line with other two biomarkers (DNA strand breaks and DNA-base oxidation). Similar results were also observed in an earlier study [24], though not reaching statistical significance with controls. This discrepancy could be explained by variations in sample size, exposure levels of styrene, ethnicity, and measurement methods of urinary 8-OH-dG. Taken together, our data and previous observations suggested that oxidative stress played a key role in styrene-induced genetic damage.
Consistent with many previous studies [9], our current results indicated a positive correlation between styrene exposure and DNA strand breaks in leukocytes. However, ethnic differences in styrene-related health outcomes may exist. For instance, one study showed that high exposure styrene (46.74 ppm) in Caucasians induced significant DNA damage and decreased the DNA repair capacities [25], but not in Caucasians exposed to lower levels of styrene (19.13 ppm) [26]. A recent study, however, showed that exposure to styrene of less than 20 ppm was associated with significantly increased genotoxic risk and decreased DNA repair capacity in Asian workers [22]. These inconsistent results suggest that East Asians are more sensitive to styrene-induced toxicity.
Ethnic differences were also found in metabolism of styrene, as a previous study described lower concentrations of urinary PGA in Orientals than in Caucasians [20]. Such difference may be explained by physiological factors such as body size, body composition, and rental function, as well as genetic background.
In this study, we focused on the effects of ALDH2 polymorphisms on styrene metabolism and styrene-related DNA damage. Our results indicated that variant allele ALDH2 *2 could significantly decrease the excretion of urinary MA+PGA in workers exposed to styrene. It is believed that the ALDH2 polymorphisms alter the enzyme activity and influence the oxidation of styrene glycol to MA, which may subsequently result in the accumulation of upstream product of MA, i.e., styrene glycoaldehydes. In animal experiments of styrene exposure by either oral administration or inhalation, our results also showed that Aldh2 knockout mice had lower concentrations of MA and PGA and higher DNA damage than those of wild-type mice (unpublished data). Also, previous studies [18, 27–29] have demonstrated that clearance of other aldehyde intermediates (e.g. acetaldehyde, ethylene glycoaldehyde, methoxyacetaldehyde, benzaldehyde, and propionaldehyde) were modulated by ALDH2 polymorphisms. These strong evidences together showed that individuals with ALDH2 variant allele have more difficulty of clearing aldehydes. Furthermore, accumulation of the aldehyde intermediate appeared to be related with increased DNA damage induced by styrene in occupational workers, as allelic variation of ALDH2 increased the levels of DNA strand breaks, DNA-base oxidation of leukocytes and urinary 8-OH-dG as comparison to wild-type genotype. It is well known that aldehydes are highly reactive molecules that react with biomacromolecules (e.g. proteins, DNA) to yield covalent adducts, which are likely to cause a series of functional alteration of pathology [30, 31]. Existing evidences showed that aldehydes (such as formaldehyde, acetaldehyde, crotonaldehyde, furfural, acrolein, methoxyacetaldehyde, benzaldehyde and aldehydes intermediates of some drugs) could cause toxic side-effect and even increased cancer risk. The metabolism and toxic effects of some aldehydes have been proven to be modified by ALDH2 polymorphisms [12, 32–37]. So the regulatory role of ALDH2 polymorphisms in detoxification of aldehyde intermediate would be crucial in understanding the mechanism underlying the inter-individual difference on styrene-induced toxicity. In addition, increased genetic toxicity of styrene in relation to ALDH2 polymorphisms is possibly related to other toxic products of styrene, such as styrene-7,8-oxide (SO), which is well documented for playing an important role in risk assessment of styrene. Since we cannot exclude the possibility that ALDH2 activity might also influence the metabolic fate of upstream products of aldehyde intermediate, more experiments with animal or in vitro system are needed to address this concern.
Regarding other polymorphisms of enzymes, our results suggested that ALDH2 activity appeared to mask some modifications of CYP2E1 polymorphisms on styrene metabolism and genotoxic effects, since we found that heterozygous c1/c2 of CYP2E1 significantly decreased the excretion of urinary MA+PGA among the styrene-exposed workers bearing ALDH2 *1/*1 genotype. Moreover, the similar results were also found in styrene-related DNA damage. Our positive results regarding association between CYP2E1 polymorphisms and styrene metabolism can be explained by an earlier study, showing that CYP2E1 c1/c2 decreased the expression of CYP2E1 mRNA [38].
GSTs could protect from styrene-induced toxicity through its catalyzing GSH conjugation of SO. We found that GSTM1 null genotype significantly decreased the MA+PGA concentrations in individuals carrying ALDH2*1/*1, but not in all exposed workers. Also, homozygous deletions of GSTM1 and GSTT1 did not significantly affect the genetic damage induced by styrene exposure. Previous studies indicated the contradictory results about associations between GSTs polymorphisms and biomarkers for styrene metabolism and genotoxic effects [9]. In fact, these limited data are difficult to be explained using effects of GSTs polymorphisms on styrene metabolism alone, because GSH conjugation of SO via GSTs accounts for only about 1% of all absorbed styrene [1].
In theory, the EPHX1 gene encoding microsomal epoxide hydrolase is considered as a more important enzyme for detoxifying styrene-induced toxicity than GSTs, since EPHX is a key enzyme involved in the metabolism of SO. A review [9], however, suggested that no solid evidences showed that the activity of EPHX, based on the EPHX1 polymorphisms in exon 3 and exon 4, influenced the levels of urinary MA+PGA and was related to risk of SO toxicity in styrene-exposed workers. Our data indicated the correlation between higher activity of EPHX and increased urinary MA+PGA concentrations and decreased genetic damage, even though statistical significance was not achieved.
In general, our study initially identified the in vivo associations between ALDH2 polymorphisms and biomarkers of styrene metabolisms and related genotoxic effects in exposed workers. In the meantime, some results need to be further confirmed. First of all, because of the instability of aldehyde and some technical difficulties, we used urinary MA+PGA as the indirect biomarker reflecting the accumulation of styrene glycoaldehyde, rather than directly measuring the aldehyde in blood. Secondly, additional in vitro and in vivo studies are still needed to investigate the toxicological mechanisms related to styrene glycoaldehyde. Furthermore, a well-designed animal model such as Aldh2 knockout mouse is required to confirm ALDH2 polymorphisms involved in inter-individual variation in the study of styrene exposure, in order to minimize potential confounding factors that can be hardly avoided in human epidemiological studies. Despite these, the results obtained in this study clearly suggested that ALDH2 polymorphisms play a prominent role in styrene metabolism and related genotoxicity. Combined with accumulated evidences regarding the function of ALDH2 in the detoxification of toxic and carcinogenic aldehyde intermediates [12, 39, 40], it is reasonable to consider individual ALDH2 polymorphisms in quantitatively assessing the risk of styrene exposure.
MATERIALS AND METHODS
Study subjects
This cross-sectional study was approved by the Research Ethics Committees of the National Institute of Occupational Safety and Health, Japan, and National Institute for Occupational Health and Poison Control, Chinese Center for Disease Control and Prevention. In this study, all the participants were interviewed by an occupational physician using a detailed questionnaire, which included demographic information, occupational history, individual lifestyle, and personal medical history, and styrene-exposed participants worked for at least six months. The study was conducted on healthy individuals and exclusion criteria included recent exposure to mutagenic agents (such as X-ray), current drug use, and recent viral infections. Finally, 329 styrene-exposed workers were recruited from three different workplaces, defined as workplace A, B and C, of the fiberglass-reinforced plastic factory in China, and 152 unexposed clerks were also recruited from the same factory.
Blood and urine sampling
In an annual physical examination, blood and urine samples were collected from 8 a.m to 11 a.m on Wednesday, Thursday and Friday, before subjects started their work. All samples were placed in an icebox and were immediately transported to the National Institute of Occupational Health and Poison Control, China CDC. Part of the blood samples was immediately used for the analysis of comet assay on the same day of sampling. The remaining blood, serum and urine samples were stored at – 80 ºC. All samples were coded and analyzed under blind conditions.
Styrene exposure at the workplace
In each workplace, 10 workers located at different sites were randomly selected to wear personal dosimeters (3M, St. Paul, MN) close to the breathing zone for 8 h (an entire work shift). The concentration of airborne styrene at the workplace was analyzed by an Agilent gas chromatography (Santa Clara, CA) with the column HP-INNOWax according to the National Institute for Occupational Safety and Health method 1501 [41].
Analysis of urinary styrene metabolites
Urinary MA and PGA concentrations were determined as described in a prior study [42]. An aliquot of 100 μl urine sample was diluted with 1ml pure water. Mixed solution was filtered using syringe-driven filter unit (Millipore, Billerica, MA). 20 μl of each sample was injected into the HPLC system (Agilent model 1100 series) equipped with a UV detector. The column (GL Sciences, Tokyo, Japan) used was 150 mm in length and 4.6 mm in inner diameter, and was packed with inertsil ODS (diameter of the granules, 5 μm). The final concentrations of MA and PGA were expressed as mg/g Creatinine (Cr.), and Cr. concentrations in the urine samples were determined according to Jaffe’s method. Sample with Cr. Concentrations lower than 0.3 g/L or higher than 3.0 g/L were excluded from statistical analysis according to the American Conference of Governmental Industrial Hygienists recommendation [43].
Measurement of urinary 8-OH-dG
Urinary 8-OH-dG concentration was determined by HPLC-ECD using a two-separation method as described previously [44] and expressed as mg/g Creatinine. Urine samples were thawed and mix with a solution containing the ribonucleoside marker 8-hydroxygunosine (8- OH- G). After mixed solutions were put at 4ºC for 2 h, then centrifuged at 13,000 rpm for 5 min, and the 20 μl aliquots of the supernatants were used for the analysis. Firstly, 8-OH-dG fraction in supernatant was separated by an anion exchange column (MCI GEL CA08F, 7 μm, 1.5 × 150 mm), and collected based on the 8-OH-G peak. Secondly, collected 8-OH-dG fraction was further separated by a reverse phase column (Shiseido, Capcell Pak C18, 5 μm, 4.6 × 250 mm), then and was detected by a Coulochem II electrochemical detector (ESA) with a guard cell (5020) and an analytical cell (5011). Applied voltage was set at 400 mV in guard cell and was set at 190 mV and 350 mV in analytical cell. The value of urinary 8-OH-dG was expressed as mg/g Cr.
Standard comet assay
The comet assay was carried out under alkaline conditions, basically as a prior description [45] with some modification [46]. Finally, 100 cells were scored per sample using the Comet Assay IV capture system (Perceptive Instruments, Suffolk, UK). The tail intensity (TI), defined as the percentage of DNA migrated from the head of the comet into the tail, was measured for each nucleus.
FPG-modified comet assay
Details of analytical conditions about the formamido-pyrimidine-DNA-glycosylase (FPG)-modified comet assay were described in a previous study [47]. Briefly, samples were treated with 1 U FPG enzyme (Trevigen, Gaithersburg, MD, USA) per gel in 50 μl FPG buffer (40 mmol/L HEPES, 0.1 mol/L KCl, 0.5 mmol/L EDTA, 0.2 mg/mL bovine serum albumin, pH 8) for 60 min at 37°C, the following procedure was in line with the standard come assay. The net FPG DNA damage was considered as the differences in measured TI values between samples obtained with standard alkaline comet assay (basic DNA damage) and FPG-modified comet assay (total DNA damage).
DNA isolation and genotyping
Genomic DNA was isolated from whole blood samples using EZ1 Blood DNA kit (Qiagen, Hilden, Germany). Genotyping for ALDH2, CYP2E1 (5´-flanking region, RsaI/PstI, 96-bp insertion, and intron 6, DraI), EPHX (exon 3, Tyr113His, EcoRV and exon 4, His139Arg, RsaI), GSTM1, and GSTT1 was carried out as described previously [48–52]. Approximately 30% of all samples with clear genotypes were regenotyped once in independent experiments, and repeat results were 100% consistent with previous results. To Inconclusive samples were reanalyzed twice or more times, and only concordant results from the analyses were accepted.
The EPHX enzymatic activity in individuals was classified based on the results of genotyping of polymorphisms in exons 3 and 4 [9]. Low activity: His/His–His/His; His/His–His/Arg; Tyr/His–His/His; His/His–Arg/Arg. Medium activity: Tyr/Tyr–His/His; Tyr/His–His/Arg; Tyr/His–Arg/Arg. High activity: Tyr/Tyr–Arg/Arg; Tyr/Tyr–His/Arg.
Statistical analysis
The deviation of genotype distribution from Hardy-Weinberg equilibrium was examined with Chi-square test in both control and exposed groups. Then, the independent t - test was performed to compare the levels of biomarkers (i.e. DNA strand breaks, DNA-base oxidation in leukocytes, urinary 8-OH-dG concentrations, and urinary MA+PGA) (1) between exposure and control groups, and (2) between the subjects carrying mutant allele and those of wild-type genotype. The Levene’s test was used to assess the equality of variances of data. If necessary, the data were transformed in order to restore equal variances. A p-value < 0.05 was used as the threshold of statistical significance. In addition, the styrene-ALDH2 genotypes interaction on the degree of genetic damage was further confirmed by multivariate linear regression models adjusting the effects of sex, age, years of employment, smoking and alcohol drinking. All analyses were performed using the epicalc package of R software (cran.r-project.org/package=epical).
ACKNOWLEDGMENTS AND FUNDING
We appreciate the assistance of Dr. Kejian Wang with statistical analysis and data visualization. We are grateful to Ms. Yukie Uchiyama for her assistance in the measurement of urinary 8-OH-dG. This work was supported by grant-in-aid for project research from the Japan National Institute of Occupational Safety and Health, Japan (F22–13), and Start-up Funds of Minjiang Scholar (510246) and Open-test Funds of valuable Instruments and Equipment (2016T005), Fujian, China.
CONFLICTS OF INTEREST
The authors declare that there are no conflicts of interest.
REFERENCES
1. Sumner SJ, Fennell TR. Review of the metabolic fate of styrene. Crit Rev Toxicol. 1994; 24:S11–33.
2. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Some traditional herbal medicines, some mycotoxins, naphthalene and styrene. IARC Monogr Eval Carcinog Risks Hum. 2002; 82:1–556.
3. Miller RR, Newhook R, Poole A. Styrene production, use, and human exposure. Crit Rev Toxicol. 1994; 24:S1–10.
4. Newhook R, Caldwell I. Exposure to styrene in the general Canadian population. IARC Sci Publ. 1993; 127:27–33.
5. Hodgson AT, Wooley JD, Daisey JM. Emissions of volatile organic compounds from new carpets measured in a large-scale environmental chamber. Air Waste. 1993; 43:316–324.
6. Gelbke HP, Banton M, Faes E, Leibold E, Pemberton M, Duhayon S. Derivation of safe health-based exposure limits for potential consumer exposure to styrene migrating into food from food containers. Food Chem Toxicol. 2014; 64:258–269.
7. Limasset JC, Simon P, Poirot P, Subra I, Grzebyk M. Estimation of the percutaneous absorption of styrene in an industrial situation. Int Arch Occup Environ Health. 1999; 72:46–51.
8. Henderson LM, Speit G. Review of the genotoxicity of styrene in humans. Mutat Res. 2005; 589:158–191.
9. Rueff J, Teixeira JP, Santos LS, Gaspar JF. Genetic effects and biotoxicity monitoring of occupational styrene exposure. Clin Chim Acta. 2009; 399:8–23.
10. IARC. Some industrial chemicals. IARC Monographs on the evaluation of the carcinogenic risk of chemicals to humans. International Agency for Research on Cancer. 1994; vol. 60.
11. Liu SY, Zheng PS. High aldehyde dehydrogenase activity identifies cancer stem cells in human cervical cancer. Oncotarget. 2013; 4:2462–2475. doi: 10.18632/oncotarget.1578.
12. Seitz HK, Stickel F. Molecular mechanisms of alcohol-mediated carcinogenesis. Nat Rev Cancer. 2007; 7:599–612.
13. Brennan P, Lewis S, Hashibe M, Bell DA, Boffetta P, Bouchardy C, Caporaso N, Chen C, Coutelle C, Diehl SR, Hayes RB, Olshan AF, Schwartz SM, et al. Pooled analysis of alcohol dehydrogenase genotypes and head and neck cancer: a HuGE review. Am J Epidemiol. 2004; 159:1–16.
14. Secretan B, Straif K, Baan R, Grosse Y, El Ghissassi F, Bouvard V, Benbrahim-Tallaa L, Guha N, Freeman C, Galichet L, Cogliano V, Group WHOIAfRoCMW. A review of human carcinogens—Part E: tobacco, areca nut, alcohol, coal smoke, and salted fish. Lancet Oncol. 2009; 10:1033–1034.
15. Au Yeung SL, Jiang C, Cheng KK, Liu B, Zhang W, Lam TH, Leung GM, Schooling CM. Is aldehyde dehydrogenase 2 a credible genetic instrument for alcohol use in Mendelian randomization analysis in Southern Chinese men? Int J Epidemiol. 2013; 42:318–328.
16. Shin CM, Kim N, Cho SI, Kim JS, Jung HC, Song IS. Association between alcohol intake and risk for gastric cancer with regard to ALDH2 genotype in the Korean population. Int J Epidemiol. 2011; 40:1047–1055.
17. Jang JY, Kang SK, Chung HK. Biological exposure indices of organic solvents for Korean workers. Int Arch Occup Environ Health. 1993; 65:S219–222.
18. Kawamoto T, Koga M, Murata K, Matsuda S, Kodama Y. Effects of ALDH2, CYP1A1, and CYP2E1 genetic polymorphisms and smoking and drinking habits on toluene metabolism in humans. Toxicol Appl Pharmacol. 1995; 133:295–304.
19. Kawamoto T, Matsuno K, Kodama Y, Murata K, Matsuda S. ALDH2 polymorphism and biological monitoring of toluene. Arch Environ Health. 1994; 49:332–336.
20. Jang JY, Droz PO, Berode M. Ethnic differences in biological monitoring of several organic solvents. I. Human exposure experiment. Int Arch Occup Environ Health. 1997; 69:343–349.
21. Marczynski B, Peel M, Baur X. New aspects in genotoxic risk assessment of styrene exposure--a working hypothesis. Med Hypotheses. 2000; 54:619–623.
22. Wongvijitsuk S, Navasumrit P, Vattanasit U, Parnlob V, Ruchirawat M. Low level occupational exposure to styrene: its effects on DNA damage and DNA repair. Int J Hyg Environ Health. 2011; 214:127–137.
23. Marczynski B, Rozynek P, Elliehausen HJ, Korn M, Baur X. Detection of 8-hydroxydeoxyguanosine, a marker of oxidative DNA damage, in white blood cells of workers occupationally exposed to styrene. Arch Toxicol. 1997; 71:496–500.
24. Manini P, De Palma G, Andreoli R, Marczynski B, Hanova M, Mozzoni P, Naccarati A, Vodickova L, Hlavac P, Mutti A, Vodicka P. Biomarkers of nucleic acid oxidation, polymorphism in, and expression of, hOGG1 gene in styrene-exposed workers. Toxicol Lett. 2009; 190:41–47.
25. Fracasso ME, Doria D, Carrieri M, Bartolucci GB, Quintavalle S, De Rosa E. DNA single- and double-strand breaks by alkaline- and immuno-comet assay in lymphocytes of workers exposed to styrene. Toxicol Lett. 2009; 185:9–15.
26. Vodicka P, Tuimala J, Stetina R, Kumar R, Manini P, Naccarati A, Maestri L, Vodickova L, Kuricova M, Jarventaus H, Majvaldova Z, Hirvonen A, Imbriani M, et al. Cytogenetic markers, DNA single-strand breaks, urinary metabolites, and DNA repair rates in styrene-exposed lamination workers. Environ Health Perspect. 2004; 112:867–871.
27. Chen YC, Peng GS, Wang MF, Tsao TP, Yin SJ. Polymorphism of ethanol-metabolism genes and alcoholism: correlation of allelic variations with the pharmacokinetic and pharmacodynamic consequences. Chem Biol Interact. 2009; 178:2–7.
28. Kitagawa K, Kawamoto T, Kunugita N, Tsukiyama T, Okamoto K, Yoshida A, Nakayama K, Nakayama K. Aldehyde dehydrogenase (ALDH) 2 associates with oxidation of methoxyacetaldehyde; in vitro analysis with liver subcellular fraction derived from human and Aldh2 gene targeting mouse. FEBS Lett. 2000; 476:306–311.
29. Wang RS, Ohtani K, Suda M, Kitagawa K, Nakayama K, Kawamoto T, Nakajima T. Reproductive toxicity of ethylene glycol monoethyl ether in Aldh2 knockout mice. Ind Health. 2007; 45:574–578.
30. Langevin F, Crossan GP, Rosado IV, Arends MJ, Patel KJ. Fancd2 counteracts the toxic effects of naturally produced aldehydes in mice. Nature. 2011; 475:53–58.
31. Lee HW, Wang HT, Weng MW, Hu Y, Chen WS, Chou D, Liu Y, Donin N, Huang WC, Lepor H, Wu XR, Wang H, Beland FA, et al. Acrolein- and 4-Aminobiphenyl-DNA adducts in human bladder mucosa and tumor tissue and their mutagenicity in human urothelial cells. Oncotarget. 2014; 5:3526–3540. doi: 10.18632/oncotarget.1954.
32. Grilo NM, Charneira C, Pereira SA, Monteiro EC, Marques MM, Antunes AM. Bioactivation to an aldehyde metabolite—possible role in the onset of toxicity induced by the anti- HIV drug abacavir. Toxicol Lett. 2014; 224:416–423.
33. O’Brien PJ, Siraki AG, Shangari N. Aldehyde sources, metabolism, molecular toxicity mechanisms, and possible effects on human health. Crit Rev Toxicol. 2005; 35:609–662.
34. Weng Z, Ohtani K, Suda M, Yanagiba Y, Kawamoto T, Nakajima T, Wang RS. Assessment of the reproductive toxicity of inhalation exposure to ethyl tertiary butyl ether in male mice with normal, low active and inactive ALDH2. Arch Toxicol. 2014; 88:1007–1021.
35. Weng Z, Suda M, Ohtani K, Mei N, Kawamoto T, Nakajima T, Wang RS. Aldh2 knockout mice were more sensitive to DNA damage in leukocytes due to ethyl tertiary butyl ether exposure. Ind Health. 2011; 49:396–399.
36. Weng Z, Suda M, Ohtani K, Mei N, Kawamoto T, Nakajima T, Wang RS. Differential genotoxic effects of subchronic exposure to ethyl tertiary butyl ether in the livers of Aldh2 knockout and wild-type mice. Arch Toxicol. 2012; 86:675–682.
37. Weng Z, Suda M, Ohtani K, Mei N, Kawamoto T, Nakajima T, Wang RS. Subchronic exposure to ethyl tertiary butyl ether resulting in genetic damage in Aldh2 knockout mice. Toxicology. 2013; 311:107–114.
38. Prieto-Castello MJ, Cardona A, Marhuenda D, Roel JM, Corno A. Use of the CYP2E1 genotype and phenotype for the biological monitoring of occupational exposure to styrene. Toxicol Lett. 2010; 192:34–39.
39. Speit G, Henderson L. Review of the in vivo genotoxicity tests performed with styrene. Mutat Res. 2005; 589:67–79.
40. Vasiliou V, Pappa A. Polymorphisms of human aldehyde dehydrogenases. Consequences for drug metabolism and disease. Pharmacology. 2000; 61:192–198.
41. NIOSH. U.S. Department of Health and Human Services-Centers for Diseases Control and Prevention, National Institute for Occupational Safety and Health. Manual of Analytical Methods. 1994; Cincinnati, OH, USA.
42. Eitaki Y, Kawai T, Kishi R, Sakurai H, Ikeda M. Stability in urine of authentic phenylglyoxylic and mandelic acids as urinary markers of occupational exposure to styrene. J Occup Health. 2008; 50:221–228.
43. ACGIH. Threshold limit values (TLVs) for chemical substances and physical agents and biological exposure indices (BEIs). In: American Conference of Governmental Industrial Hygienists. 2004; Cincinnati (OH).
44. Kasai H. A new automated method to analyze urinary 8-hydroxydeoxyguanosine by a high-performance liquid chromatography-electrochemical detector system. J Radiat Res. 2003; 44:185–189.
45. Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp Cell Res. 1988; 175:184–191.
46. Weng Z, Lu Y, Weng H, Morimoto K. Effects of the XRCC1 gene-environment interactions on DNA damage in healthy Japanese workers. Environ Mol Mutagen. 2008; 49:708–719.
47. Collins AR, Duthie SJ, Dobson VL. Direct enzymic detection of endogenous oxidative base damage in human lymphocyte DNA. Carcinogenesis. 1993; 14:1733–1735.
48. Le Marchand L, Donlon T, Seifried A, Wilkens LR. Red meat intake, CYP2E1 genetic polymorphisms, and colorectal cancer risk. Cancer Epidemiol Biomarkers Prev. 2002; 11:1019–1024.
49. Sarmanova J, Tynkova L, Susova S, Gut I, Soucek P. Genetic polymorphisms of biotransformation enzymes: allele frequencies in the population of the Czech Republic. Pharmacogenetics. 2000; 10:781–788.
50. Teixeira JP, Gaspar J, Martinho G, Silva S, Rodrigues S, Mayan O, Martin E, Farmer PB, Rueff J. Aromatic DNA adduct levels in coke oven workers: correlation with polymorphisms in genes GSTP1, GSTM1, GSTT1 and CYP1A1. Mutat Res. 2002; 517:147–155.
51. Teixeira JP, Gaspar J, Silva S, Torres J, Silva SN, Azevedo MC, Neves P, Laffon B, Mendez J, Goncalves C, Mayan O, Farmer PB, Rueff J. Occupational exposure to styrene: modulation of cytogenetic damage and levels of urinary metabolites of styrene by polymorphisms in genes CYP2E1, EPHX1, GSTM1, GSTT1 and GSTP1. Toxicology. 2004; 195:231–242.
52. Wang RS, Nakajima T, Kawamoto T, Honma T. Effects of aldehyde dehydrogenase-2 genetic polymorphisms on metabolism of structurally different aldehydes in human liver. Drug Metab Dispos. 2002; 30:69–73.