
INTRODUCTION
Ovarian cancer is the eighth most common cancer in women and the leading cause of death from gynecological malignancies in developed countries [1]. According to the American Cancer Society (Atlanta, GA), 21,980 new ovarian cancer cases were diagnosed in 2014, and the 5-year survival was around 44% [2]. Approximately 70% of ovarian cancer patients are diagnosed at the stage of FIGO III/IV with widespread cancer cells beyond ovaries. Despite surgical and chemotherapeutic improvements, most patients may eventually relapse and have poor prognosis after primary treatment. Numerous studies have demonstrated that EMT is associated with the motility or invasion of ovarian carcinoma. However, the detailed mechanism of EMT in ovarian cancer has yet to be defined.
Wip1, also called as PPM1D (protein phosphatase, Mg2+/Mn2+ dependent 1D), was first identified by Michele Fiscella and his colleges in 1997 [3]. Wip1 plays its role through dephosphorylating the downstream proteins including p38, p53, ATM, Chk2, and γ-H2AX that are involved in DNA damage response (DDR). Wip1 inactivates these molecules and turns back the cells to a homeostasis state when a process of DDR is completed [3–6]. Besides, Wip1 has recently been reported to be involved in regulation of cellular metastasis. Zhao et al. [7, 8] reported that Wip1 might negatively regulate neutrophil migration through regulating p38 MAPK activity. Li et al. [9] found that Wip1 knockout enhanced the migration of bone marrow mesenchymal stem cells. Paradoxically, several other studies suggest that Wip1 may enhance migration or invasion in some carcinomas [10, 11]. Although p53 is high mutated in serous ovarian cancer, the function of Wip1 in ovarian cancer metastasis is unclear.
In the present study, we investigated the biological function of Wip1 in ovarian cancer via both in vitro and in vivo assays. Our results suggest that Wip1 inhibits ovarian cancer metastasis through the ATM/Akt/Snail mediated signaling.
RESULTS
Knockdown of Wip1 promotes cell invasion and migration
To examine the expression of Wip1, serous ovarian cancer cell lines A2780, Hey, HeyA8, SKOV3, SKOV3 ip1, OVCA433 and the clear cell carcinoma cell line OVCA429 were cultured and subjected to Western blotting analysis. MCF7 and HeLa cell lines were chosen as the positive and negative controls, respectively. According to Figure 1A, A2780 and HeyA8 cells were chosen for Wip1 knockdown experiments. Reconstructed plasmid pLKO.1-shWip1 and the control pLKO.1-shGFP were used to establish A2780/shWip1and HeyA8/shWip1 cell lines and the corresponding control cell lines A2780/shGFP and Hey A8/shGFP, which were verified by Western blotting and real time-PCR (Figure 1B). We found that knockdown of Wip1 increased cell migration (Figure 1C and Figure 1D) and invasion (Figure 1E), indicating that Wip1 may suppress cell invasion and migration in serous ovarian cancer cells.
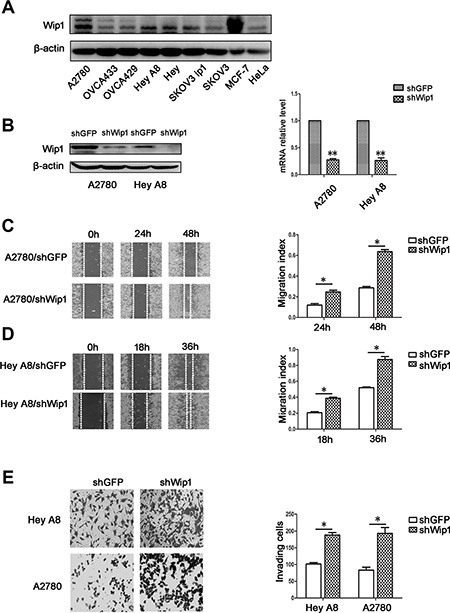
Figure 1: Knockdown of Wip1 promotes cell invasion and migration. (A) Expression of Wip1 in A2780, OVCA433, OVCA429, Hey A8, Hey, SKOV3 ip1, SKOV3, MCF7 (a positive control) and Hela (a negative control) cell lines. (B) Efficiency of Wip1 shRNA detected by Western blotting and RT-PCR. **p < 0.01. Error bars = 95% CIs. (C, D) Migration assay. Wounds were imaged at 0 h, 24 h, and 48 h (A2780) and at 0 h, 18 h, and 36 h (HeyA8). *p < 0.05. Error bars = 95% CIs. (E) Invasion was detected by Trans-well experiments. Image was acquired at 24 h. *p < 0.05. Error bars = 95% CIs.
Wip1 suppresses EMT
As shown in Figure 2A, cells expressing Wip1 shRNA displayed a thinner and longer morphology than did control cells expressing shGFP. To illustrate the potential mechanism, MMPs and EMT-related molecules were examined by using Western blotting. As shown in Figure 2B, knockdown of Wip1 decreased the expression of E-cadherin, while remarkably increased the expression of N-cadherin, snail and slug. The expression of vimentin, MMP9, MMP2 was slightly increased, but no changes of β-catenin were conceived. Since Wip1 is a serine-threonine protein phosphatase, we speculated that a serine/threonine protein kinase might be closely regulated by Wip1 through protein dephosphorylation.
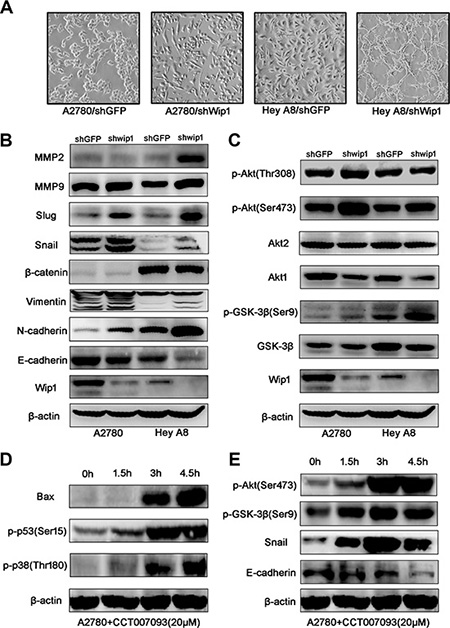
Figure 2: Wip1 knockdown activates EMT and the Akt/GSK-3β signaling. (A) Morpgological change in cells expressing Wip1 shRNA. (B) Knockdown of Wip1 reduces E-cadherin, but increases N-cadherin, snail, slug, vimentin, MMP9, MMP2, but did not influence expression of β-catenin. (C) Wip1 knockdown promotes the phospharylation of GSK-3β (Ser9) and Akt (Ser473). (D, E) Treatment with CCT007093 activated the Akt/GSK-3β signaling pathway and EMT.
Akt, a serine-threonine protein kinase, plays a central role in a variety of oncogenic processes. It is well known that the Akt activation associated with tumor metastasis is mediated through the phosphorylation at Ser473 [12]. Previous reports also suggest that the activation of Akt may induce the phosphorylation of GSK-3β to suppress the GSK-3β/snail/E-cadherin signaling [12–14]. Therefore, we tested the alterations of these molecules. As shown in Figure 2C, knockdown of Wip1 decreased the expression of Akt1 and GSK-3β, but increased the phosphorylation of GSK-3β at Ser9 and Akt at Ser473. However, no alterations of Akt2 and p-Akt (Thr308) were observed. To validate the above results, CCT007093 (10 μM), a specific inhibitor of Wip1 [15], was used to perform a time course inhibition assay. As shown in Figure 2D, treatment of cells with CCT007093 activated p38 and p53, and enhanced the expression of pro-apoptotic protein Bax, which is consistent with other reports [16, 17]. Meanwhile, CCT007093 treatment decreased E-cadherin and increased pGSK-3βSer9 and pAktSer473 in a time-dependent manner (Figure 2E). These results revealed that the inhibition of Wip1 activity by using the specific inhibitor yielded a similar effect to shRNA transfection. Collectively, these data suggest that inhibition of Wip1 may activate the Akt/GSK-3β/snail signaling.
Wip1 overexpression decreases cellular migration and invasion ability
As silencing of Wip1 promoted metastasis in serous ovarian cancer cells, we examined whether overexpression of Wip1 could suppress cellular motility. Thus, the pCDH-PPM1D plasmid was generated and transfected into SKOV3 and OVCA433 cells. Compared with in control cells, the expression of Wip1 was dramatically increased in SKOV3/Wip1 cDNA and OVCA433/Wip1 cDNA cells, which was assessed by Western blotting and real time-PCR (Figure 3A). Remarkably, overexpression of Wip1 decreased invasion and migration in vitro (Figure 3B–3D) by upregulation of E-cadherin and down regulation of snail, pAktSer473, and pGSK-3βSer9 (Figure 3E).
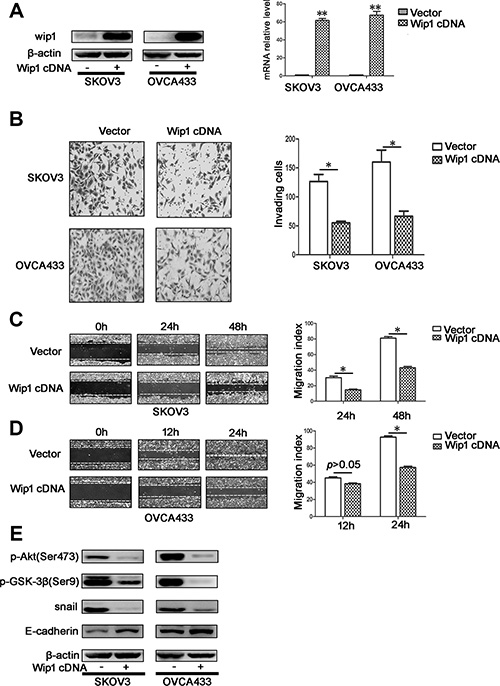
Figure 3: Overexpression of Wip1 decreases cellular migration and invasion in vitro. (A) Overexpession of Wip1 detected by Western blotting and real time-PCT in SKOV3 and OVCA433 cell lines. **p < 0.01. Error bars = 95% CIs. (B) Cellular invasion was detected by trans-well experiment. The invaded cells were imaged at 24 h. *p < 0.05. Error bars = 95% CIs. (C, D) Cellular migration was detected by Scratch assay. The wounds were imaged at 0 h, 24 h, 48 h (SKOV3) and at 0 h, 12 h, 24 h (OVCA433). *p < 0.05. Error bars = 95% CIs. (E) Overexpression of Wip1 suppressed Akt/GSK-3β signaling and EMT.
Wip1 negatively regulates tumor metastasis
To test how Wip1 regulates tumor metastasis in vivo, animals were introperitoneally injected with A2780 shWip1, Hey A8 shWip1, SKOV3/Wip1 cDNA and OVCA433/Wip1 cDNA cells and the corresponding control cells, respectively. The mice were sacrificed before natural death, and the number of nodules and their weights were measured. As shown in Figure 4A, we found that the intraperitneal injection of A2780 cells only caused mesenteric metastases, but no liver metastases. Knockdown of Wip1 increased mesenteric metastases in A2780 cells, including both the weight of tumor nodules (Figure 4B) and average numbers (Figure 4C). Introperitoneal injection of Hey A8 or SKOV3 cells presented both mesenteric and liver metastases (Figure 4A). Wip1 knockdown in Hey A8 cells increased both liver tumor burden and mesenteric metastases although there was not statistical significance (Figure 4B and 4C). On the contrary, overexpression of Wip1 in SKOV3 cells decreased mesenteric metastases (p < 0.05), but did not show significant differences compared with liver metastases (Figure 4B and 4C). Both xenograft mice injected with OVCA433/Wip1 cDNA and control cells did not form any tumors in vivo (Figure 4A). Meanwhile, as shown in Figure 4D, knockdown of Wip1 in Hey A8 cells increased the volume of ascites, while overexpression of Wip1 in SKOV3 cells decreased the volume of ascites. Two different patterns of mesenteric metastases and liver metastases were shown in Figure 4E.
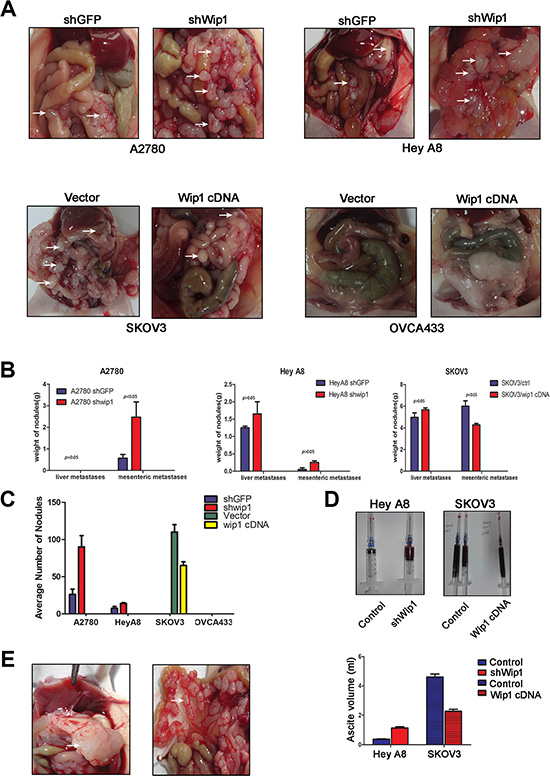
Figure 4: Wip1 suppresses tumor metastasis in vivo. (A) In intraperitoneal models, knockdown of Wip1 promotes formation of metastatic nodules in small bowels and mesenteric surface and liver (A2780 and Hey A8). Overexpression of Wip1 in SKOV3 decreased mesenteric metastases. Injection of OVCA433 did not form tumors in vivo. White arrows referred to metastatic loci. (B) Weight of nodules. Error bars = 95% CIs. (C) Quantification of nodules. Error bars = 95% CIs. (D) Measurement of ascite volume. (E) Different metastatic patterns in our experiment: liver metastasis and mesenteric metastasis.
Wip1 supresses cellular motility through the Akt/GSK-3β/snail signaling
The above data indicate that snail may be the major transcriptional factor during EMT process of ovarian carcinoma cells. To test whether snail truly contributes to the metastasis induced by Wip1 knockdown, we transiently transfected snail shRNA into the A2780/shWip1 cells. As shown in Figure 5A, knockdown of snail attenuated the cellular invasion of A2780/shWip1 cells. The expression level of E-cadherin was also increased in snail silencing cells compared with in control cells (Figure 5B). The above results suggest that the EMT transcriptional factor snail may be a direct downstream target of Wip1.
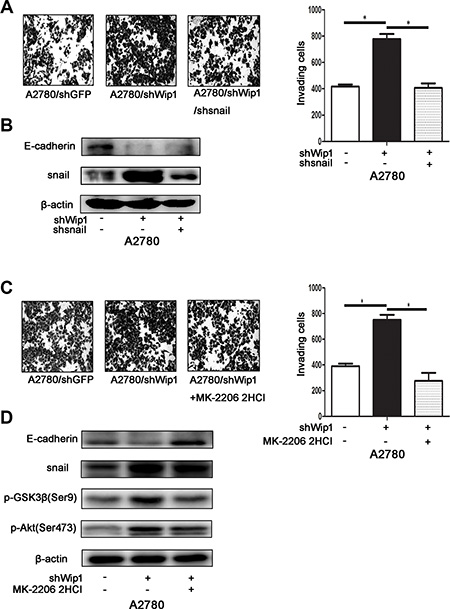
Figure 5: Wip1 suppresses cellular motility through Akt/GSK-3β/snail pathway signaling. (A) Knockdown of snail in A2780/shWip1 neutralized the increased invading cells. Image was acquired at 36 h. *p < 0.05. Error bars = 95% CIs. (B) The decreased expression of E-cadherin Snail knockdown neutralized. (C) Cell invasion of A2780/shWip1 after MK-2206 2HCl treatment. Image was acquired at 36 h. *p < 0.05. Error bars = 95% CIs. (D) Detection of pGSK-3βSer9, snail and E-cadherin in cells treated with MK-2206 2HCl.
Similarly, to verify if the Akt/GSK-3β signaling essentailly promotes the snail mediated cell metastasis, we used the Akt inhibitor MK-2206 2HCl to inhibit the phosphorylation of Akt at Ser473 in A2780/shWip1 cells. As in Figure 5C, MK-2206 2HCl treatment also reduced the invasive ability of A2780/shWip1 cells. Both pGSK-3βSer9 and snail were downregulated after MK-2206 2HCl treatment, while the expression level of E-cadherin was increased (Figure 5D).
ATM links Wip1 and the Akt/GSK-3β/snail signaling
To analyze the physical interaction between Wip1 and Akt/GSK-3β, we performed co-immunoprecipitation experiment. However, the binding between Akt or GSK-3β with Wip1 was not found (Data not shown). We therefore speculate other proteins may mediate the activation of Akt/ GSK-3β.
ATM, an important kinase that is activated through serine phosphorylation during DNA damage response, has been reported to be dephosphorylated by Wip1 at Ser1981 [5]. Furthermore, the PI3K domain of ATM could specifically regulate Akt (Ser473) phosphorylation [18]. To verify the potential role of ATM in the Akt/GSK-3β signaling, p-ATM was detected after cells were treated with CCT007093. As shown in Figure 6A, inhibition of Wip1 increased ATM activation in a time-dependent manner, which was consistent with a previous study [5]. We further used KU55933 [19], a specific ATM inhibitor, to evaluate the relationship between ATM and Akt. As shown in Figure 6B, the expression levels of p-AktSer473 and pGSK-3βSer9 were markedly decreased in cells treated with KU55933, indicating that ATM may activate the Akt/GSK-3β signaling.
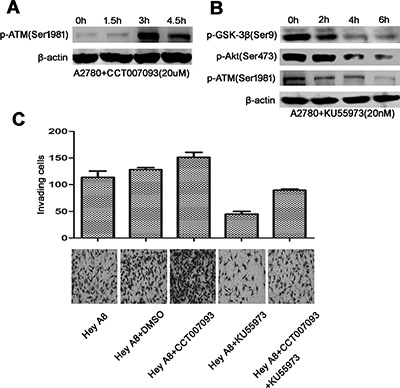
Figure 6: ATM links Wip1 and the Akt/GSK-3β/snail signaling. (A) Inhibition of Wip1 by CCT007093 increased ATM activation in a time-dependent manner. (B) KU55973 suppressed the Akt/GSK-3β signaling. Cell pellets for Western blotting were collected at 0 h, 2 h, 4 h, and 6 h. (C) Invasion array for Hey A8: Inhibition of Wip1 by CCT007093 treatment increased the number of trans-membrane cells (p < 0.05). Error bars = 95% CIs. Inhibition of ATM by KU55973 suppressed cellular invasion (p < 0.05). Error bars = 95% CIs. Co-treatment of Hey A8 cells with CCT007093 and KU55973 neutralized the increased invading cells compared with Hey A8+CCT007093 (p < 0.05). Error bars = 95% CIs.
To further illustrate the impact of the Wip1/ATM/Akt signaling on cellular metastasis, we performed an invasion assay by using the high-invasive cell line Hey A8. As in Figure 6C, inhibition of Wip1 increased the number of trans-membrane cells, while inhibition of ATM suppressed the cellular invasion through rescuing the effect of Wip1 inhibition. These data suggest that the ATM/Akt mediated signaling may exert important functions on ovarian cancer metastasis in association with Wip1.
Correlation of Wip1with p-ATM, p-AKT, snail and E-cadherin in ovarian cancer tissues
We next tested the endogenous basal level of Wip1 and pAKT, SNAIL, p-ATM, E-cadherin in the ovarian cancer cell lines. As shown in Figure 7A, Wip1 was correlated with the endogenous expression of p-ATM, pAKT, snail, but not E-cadherin To investigate the correlation of Wip1 with p-ATM, p-AKT, snail and E-cadherin in ovarian cancer tissues, we performed immunostaining using antibodies against Wip1, p-ATM, p-AKT, snail and E-cadherin in 53 high-grade serous ovarian cancer specimens. As shown In Table 1, the expression of Wip1 was negatively correlated with p-ATM, p-AKT, snail (p < 0.05), while positively correlated with E-cadherin expression, although the p value > 0.05. Representative images in the same tissue were shown in Figure 7B.
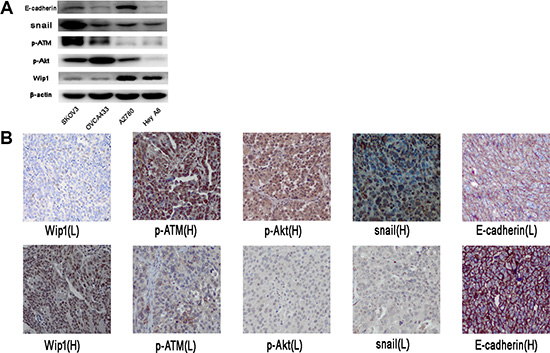
Figure 7: Correlation of Wip1with p-ATM, p-AKT, snail and E-cadherin in ovarian cancer tissues. (A) Endogenous basal level of Wip1 and pAKT, SNAIL, p-ATM, E-cadherin in SKOV3, OVCA433, A2780 and Hey A8. (B) Representative images showing the level of Wip1, p-ATM, pAKT, snail and E-cadherin in the same tissue.
Table 1: Correlation of Wip1with p-ATM, p-AKT, snail and E-cadherin in ovarian cancer tissues
N = 53 |
p-ATM |
p-AKT |
snail |
E-cadherin |
|---|---|---|---|---|
Wip1 |
Low High |
Low High |
Low High |
Low High |
Low |
9 23 |
4 28 |
8 24 |
26 6 |
High |
14 7 |
9 12 |
13 8 |
16 5 |
P value |
0.004 |
0.021 |
0.038 |
0.454 |
DISCUSSION
Our study show that knockdown of Wip1 facilitates an epithelial-mesenchymal transition through the activation of the Akt/GSK-3β signaling, leading to the enhanced invasion, migration and metastasis of serous ovarian cancer cells, while overexpression of Wip1 entirely reverses these effects. Since knockdown of snail and inhibition of the Akt/GSK-3β signaling inhibited the motility of cells with Wip1 knockdown, we propose that Akt, GSK-3β, and snail are downstream targets of Wip1, while ATM may link Wip1 with the Akt/GSK-3β/snail signaling (Figure 8).
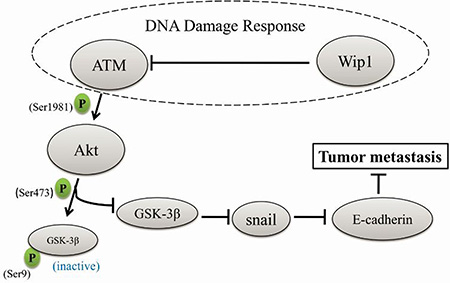
Figure 8: A schematic diagram of how Wip1 suppresses cellular metastasis through the ATM/Akt/Snail mediated signaling. Both Wip1 and ATM are important regulators in DDR. Wip1 dephosphorylates ATM at Ser1981, and ATM activation phosphorylates Akt at Ser473. Akt activation can phosphorylate GSK-3β at Ser9, which lead to the degradation of GSK-3β, while GSK-3β attenuates snail expression and upregulates E-cadherin expression.
Previous studies have shown that Wip1 regulates cellular metastasis in different cell types including neutraphil [7, 8], bone marrow mesenchymal stem cell [9], salivary adenoid cystic carcinoma [10], while we show here that Wip1 suppresses cellular metastasis in ovarian carcinoma. Despite the oncogenic role of Wip1 was reported in others carcinomas, we revealed an antitumor function of Wip1 in serous ovarian cancer cells. Since Wip1 overexpression in both p53 null (SKOV3) and wild type (OVCA433) cell lines suppressed the cellul motility in our results, we presume that Wip1 may also inhibit ovarian cancer metastasis in a p53-independent manner.
In advanced ovarian cancer patients, cancer cells always seed into the peritoneal cavity, we demonstrated that knockdown of Wip1 in both A2780 and Hey A8 cells increased the tumor nodules located at mesenteric or liver, while Wip1 overexpression in SKOV3 cells alleviated the tumor burden. The fact that the volume of ascites was affected by Wip1 knockdown or overexpression suggests a potential role of Wip1 in tumor microenvironment. These results may be explained by Zhao et al. [8] that neutrophils in Wip1-deficient mice produce more cytokines (IL-6, TNF-α, IL-1β, and so on) and granule proteins (elastase, lactoferrin, MPO, and MMP9) after LPS stimulation. Therefore, Wip1 overexpression may be a potential therapeutic choice to block the metastasis and recurrence of serous ovarian cancer, however, further study is needed to better understand the function of Wip1 in tumor microenvironment.
Taken together, our study suggest that Wip1 blocks cellular metastasis of serous ovarian cancer through the ATM/Akt/snail mediated signaling, which may be used to improve the efficiency of ovarian cancer treatment.
MATERIALS AND METHODS
Cell Lines and cell culture
Ovarian cancer lines A2780, OVCA429, Hey, HeyA8, SKOV3, SKOV3 ip1, OVCA433, breast cell line MCF7, cervical cancer cell line HeLa, and lentiviral packaging cell line 293T were obtained from the American Type Culture Collection (ATCC). 293T cell was cultured in DMEM media, and the other cells were cultured in RPMI 1640 medium supplemented with 10% (v/v) FBS and 1% penicillin and streptomycin at 37°C under a humidified condition with 95% air and 5% CO2.
Plasmid constructs
DNA oligonucleotides used to generate shRNAs against the open reading frame of mRNA were 5′-CCCTTCTCGTGTTTGCTTAAA-3′ (for Wip1) and 5′-GACTACCGCTGCTCCATTCCA-3′ (for snail) to silence the expression of Wip1 and snail respectively. The shRNAs were cloned into pLKO.1 TRC Cloning Vector (Addgene) according to the manufacturer’s protocol. The pLKO.1-shGFP was used as a control vector.
The pCMV6-AN-HA plasmid (Origene) containing full-length wild-type PPM1D cDNA was kindly gifted from Dr Nazneen Rahman [20]. The full-length wild-type PPM1D was amplified using the following: Forward: 5′-CTAGCTAGCATGGCGGGGCT GTACTCGCTGGGA-3′ and Reverse: 5′-ATAAGAATG CGGCCGC TCAGCAAACACAAACAGTTTTCCT-3′, the PCR product was digested with Nhe1 and Not1 and then cloned into the pCDH–CMV-MCS-EF1-PURO (Addgene), yielding pCDH-PPM1D construct. The pCDH empty vector was used as a control vector.
Lentivirus infection and transient transfection with Plasmid DNA
Lentiviruses harboring shWip1 or Wip1 cDNA were produced by transfecting pLKO.1/shWip1 or pCDH-PPM1D into 293T cells. Serous ovarian cancer cells were infected twice for a total of 6 days (3 days for each infection) and the positive clones were selected with puromycin for 10–14 days to establish stable cell lines expressing shWip1 or Wip1 cDNA and the corresponding control vectors. The shRNA for snail was transiently transfected into cells by FuGENE HD (Roche) according to the manufacturer’s protocol.
Cell invasion and migration
Cell invasion was assessed using a 24-well inserts (BD Biosciences) with an 8-μm pores according to the manufacturer’s instructions. Briefly, 5 × 104 cells were seeded onto upper chamber which contained the Matrigel layer and allowed to invade at 37°C for 24 h toward a lower reservoir with 10% FBS. Cells were then fixed in 100% methanol for 30 minutes and stained with Crystal violet for 10 minutes. The invasive cells were counted as those passed through the membrane. All cells were counted at ×200 magnification under a microscope.
To examine cell migration, cells were incubated in 6-well plate over night to yield monolayer of 100% confluence for scratch assay. Scratched were made using a pipette tip and photographed immediately (time 0), and at 18/24 h and 36/48 h. The distance migrated by cells monolayer to close the scratch area during the time period was measured. Both the assays were repeated three times in duplicate. Migration index is defined as the distance migrated by shRNA or cDNA treated relative to the distance migrated by the control.
The snail shRNA (4 μg) was transiently transfected into A2780/shWip1 with FuGENE HD (12 μl) for 48 hours, then the cell culture medium was removed and cells were collected for the trans-well experiments.
Real time fluorescence quantitative polymerase chain reaction
Total RNA was isolated with Trizol reagent (Invitrogen), and 1 μg RNA was reversely transcribed into cDNA using the Exscript RT-PCR kit (TaKaRa) according to the manufacturer’s instructions. Primer pairs for cDNA amplification were as follows: 5′ -GC CAGAACTTCCCAAGGAAAG-3′ (forward) and 5′-GG TTCAGGTGACACCACAAATTC-3′ (reverse) for Wip1; 5′-GGCCTCCAAGGAGTAAGACC-3′ (forward) and 5′-CAAGGGGTCTACATGGCAAC-3′ (reverse) for GA PDH. All amplifications and detections were carried out in the Applied Biosystems Prism 7900 system (Applied Biosystems, Foster City, CA) using the ExScript Sybr green QPCR kit (TaKaRa, Japan) and the following program: 1 cycle of 15min at 95°C followed by 35 cycles of (10 sec at 95°C, 20 sec at 60°C). Statisticalanalyses were performed using the 2−ΔΔCT relative quantification method. The assay was performed three times in triplicate.
Western analysis
Cell lysates containing 30ug protein were analyzed. The following primary antibodies were used: antibodies to Wip1 (H300), p-p53 (Ser15), Bax, E-cadherin, N-cadherin, β-catenin, slug, snail, vimentin, Akt1, Akt2, p-Akt (Ser473), MMP9 and MMP2 were from Santa Cruz. Antibodies for β-actin, p-p38 (Thr180), p-Akt (Thr308) were from CST (Cell Signaling Technologies), GSK-3β, p-GSK-3β (Ser9) were from BD biosciences, p-ATM (Ser1981) was from Abcam. Immunoreactivity was conducted according to standard procedures.
Inhibitor assay
CCT007093 (sigma-Aldrich, C9369), a Wip1 inhibitor, was prepared in DMSO at a stock concentration of 10 mM. A2780 was detached by trypsinization and washed with PBS twice. Same number of cells were plated in 6 wells dish for 24 h, and incubated with 20 μM CCT007093 for 0 h, 1.5 h, 3 h, and 4.5 h, then the samples were collected for Western blotting.
KU55933 (Selleck chem, S1092), an ATM inhibitor, was prepared in DMSO at a stock concentration of 50 μM. Same number of A2780 cells was incubated with 20 nM KU55933 for 0 h, 2 h, 4 h, and 6 h respectively, cell pellets were collected for Western blotting. For the invasion assay, same number of HeyA8 was plated into 24-well plate overnight and incubated with RPMI 1640 medium, RPMI 1640 + DMSO, RPMI 1640 + CCT007093 (20 μM), RPMI 1640 + KU55933 (20 nM), RPMI 1640 + CCT007093 (20 μM) + KU55933 (20 nM) respectively for 2 h, then those cells were collected for trans-well experiment.
MK-2206 2HCl (Selleck chem, S1078), an Akt inhibitor, was prepared in DMSO at a stock concentration of 10 mM. A2780/shWip1 was incubated with MK-2206 2HCl (10 nM) for 24 h, then cells were collected for invasion assay.
Animal experiments
In intraperitoneal tumor model, four mice (Balb/c nude mice) were used for each cell line and each mouse received one injection of 1 × 107cells. Mice were observed for the lethargy and abdominal enlargement every three days, and sacrificed before natural death occured. Tumor nodules and ascites were counted, weighed and measured.
Imunohistochemical staining
Ovarian tissue samples from 53 high grade serous ovarian cancer patients who underwent primary debulking surgery at Fudan University Shanghai Cancer Center were included in our study. The expression of Wip1, p-ATM (Ser1981), p-AKT (Ser473), snail and E-cadherin was detected by immunohistochemical staining. The expression levels were judged based on the intensity of staning (IS) and the percentage of positive cells (PPC) as previously reported [21]. The final score (PPC×IS) was graded as “Low” for < 4 and “High” for ≥ 4.
ACKNOWLEDGMENTS AND FUNDING
This study was supported by grants from the National Natural Science Foundation of China (No. 81572553, 81372797 for G. Yang); Shanghai Municipal Commission of Health and Family Planning Science Foundation (20124009) for RY. Zang.
CONFLICTS OF INTEREST
The authors indicate no conflicts of interest.
REFERENCES
1. Jemal A, Bray F, Center MM, Ferlay J, Ward E, Forman D. Global cancer statistics. CA Cancer J Clin. 2011; 61:69–90.
2. DeSantis CE, Lin CC, Mariotto AB, Siegel RL, Stein KD, Kramer JL, Alteri R, Robbins AS, Jemal A. Cancer treatment and survivorship statistics, 2014. CA Cancer J Clin. 2014; 64:252–71.
3. Fiscella M, Zhang H, Fan S, Sakaguchi K, Shen S, Mercer WE, Vande Woude GF, O’Connor PM, Appella E. Wip1, a novel human protein phosphatase that is induced in response to ionizing radiation in a p53-dependent manner. Proc Natl Acad Sci U S A. 1997; 94:6048–53.
4. Moon SH, Nguyen TA, Darlington Y, Lu X, Donehower LA. Dephosphorylation of gamma-H2AX by WIP1: an important homeostatic regulatory event in DNA repair and cell cycle control. Cell cycle. 2010; 9:2092–96.
5. Shreeram S, Demidov ON, Hee WK, Yamaguchi H, Onishi N, Kek C, Timofeev ON, Dudgeon C, Fornace AJ, Anderson CW, Minami Y, Appella E, Bulavin DV. Wip1 phosphatase modulates ATM-dependent signaling pathways. Mol Cell. 2006; 23:757–64.
6. Bulavin DV, Demidov ON, Saito S, Kauraniemi P, Phillips C, Amundson SA, Ambrosino C, Sauter G, Nebreda AR, Anderson CW, Kallioniemi A, Fornace AJ Jr, Appella E. Amplification of PPM1D in human tumors abrogates p53 tumor-suppressor activity. Nat Genet. 2002; 31:210–15.
7. Liu G, Hu X, Sun B, Yang T, Shi J, Zhang L, Zhao Y. Phosphatase Wip1 negatively regulates neutrophil development through p38 MAPK-STAT1. Blood. 2013; 121:519–29.
8. Sun B, Hu X, Liu G, Ma B, Xu Y, Yang T, Shi J, Yang F, Li H, Zhang L, Zhao Y. Phosphatase Wip1 negatively regulates neutrophil migration and inflammation. J Immunol. 2014; 192:1184–95.
9. Tang Y, Liu L, Sheng M, Xiong K, Huang L, Gao Q, Wei J, Wu T, Yang S, Liu H, Mu Y, Li K. Wip1 knockout inhibits the proliferation and enhances the migration of bone marrow mesenchymal stem cells. Exp Cell Res. 2015; 334:310–22.
10. Tang YL, Liu X, Gao SY, Feng H, Jiang YP, Wang SS, Yang J, Jiang J, Ma XR, Tang YJ, Chen Y, Liang XH. WIP1 stimulates migration and invasion of salivary adenoid cystic carcinoma by inducing MMP-9 and VEGF-C. Oncotarget. 2015; 6:9031–44. doi: 10.18632/oncotarget.3320.
11. Buss MC, Remke M, Lee J, Gandhi K, Schniederjan MJ, Kool M, Northcott PA, Pfister SM, Taylor MD, Castellino RC. The WIP1 oncogene promotes progression and invasion of aggressive medulloblastoma variants. Oncogene. 2015; 34:1126–40.
12. Qiao M, Sheng S, Pardee AB. Metastasis and AKT activation. Cell cycle. 2008; 7:2991–96.
13. Batlle E, Sancho E, Franci C, Dominguez D, Monfar M, Baulida J, Garcia De Herreros A. The transcription factor snail is a repressor of E-cadherin gene expression in epithelial tumour cells. Nat Cell Biol. 2000; 2:84–89.
14. Zhou BP, Deng J, Xia W, Xu J, Li YM, Gunduz M, Hung MC. Dual regulation of Snail by GSK-3beta-mediated phosphorylation in control of epithelial-mesenchymal transition. Nat Cell Biol. 2004; 6:931–40.
15. Rayter S, Elliott R, Travers J, Rowlands MG, Richardson TB, Boxall K, Jones K, Linardopoulos S, Workman P, Aherne W, Lord CJ, Ashworth A. A chemical inhibitor of PPM1D that selectively kills cells overexpressing PPM1D. Oncogene. 2008; 27:1036–44.
16. Takekawa M, Adachi M, Nakahata A, Nakayama I, Itoh F, Tsukuda H, Taya Y, Imai K. p53-inducible wip1 phosphatase mediates a negative feedback regulation of p38 MAPK-p53 signaling in response to UV radiation. EMBO J. 2000; 19:6517–26.
17. Goloudina AR, Mazur SJ, Appella E, Garrido C, Demidov ON. Wip1 sensitizes p53-negative tumors to apoptosis by regulating the Bax/Bcl-xL ratio. Cell cycle. 2012; 11:1883–87.
18. Viniegra JG, Martinez N, Modirassari P, Hernandez Losa J, Parada Cobo C, Sanchez-Arevalo Lobo VJ, Aceves Luquero CI, Alvarez-Vallina L, Ramon y Cajal S, Rojas JM, Sanchez-Prieto R. Full activation of PKB/Akt in response to insulin or ionizing radiation is mediated through ATM. J Biol Chem. 2005; 280:4029–36.
19. Li Y, Yang DQ. The ATM inhibitor KU-55933 suppresses cell proliferation and induces apoptosis by blocking Akt in cancer cells with overactivated Akt. Mol Cancer Ther. 2010; 9:113–25.
20. Ruark E, Snape K, Humburg P, Loveday C, Bajrami I, Brough R, Rodrigues DN, Renwick A, Seal S, Ramsay E, Duarte Sdel V, Rivas MA, Warren-Perry M, et al. Mosaic PPM1D mutations are associated with predisposition to breast and ovarian cancer. Nature. 2013; 493:406–10.
21. Wang ZL, Liu Y, Lu LL, Yang LN, Yin S, Wang Y, Qi ZH, Meng J, Zang RY, Yang G. Fibrillin-1, induced by Aurora-A but inhibited by BRCA2, promotes ovarian cancer metastasis. Oncotarget. 2015; 6:6670–83. doi: 10.18632/oncotarget.3118.