
INTRODUCTION
The Mouse Double Minute 2 homolog (MDM2) is a major regulator of the tumor suppressor p53. The levels of MDM2 and p53 are tightly regulated, as they act in a negative feedback loop where p53 induces MDM2 transcription in response to genotoxic stress, whereas MDM2 binds, inhibits and directs p53 for proteasomal degradation by ubiquitinylation [1–4]. MDM2 hyperactivity through mechanisms such as gene amplification, increased transcription and enhanced translation is observed in many human cancers harboring wild-type TP53 [5–7], and MDM2 overexpression has been suggested to act as an alternative mechanism of p53 inactivation, thus promoting tumor growth.
While multiple single nucleotide polymorphisms (SNPs) have been identified in MDM2 promoter regions, so far, only a few have been assessed with respect to potential biological functions. SNP309T>G; rs2279744, located in the MDM2 promoter P2, has been shown to increase MDM2 expression by increasing the binding affinity between the promoter and the transcription factor Sp1 [8]. The same investigators found the SNP309G allele to be associated with early onset of several malignancies among individuals carrying TP53 germline mutations (Li-Fraumeni syndrome) and also lower age at diagnosis of estrogen receptor (ER) rich breast cancer among individuals with wild-type TP53 [9, 8]. Subsequent findings, with respect to the influence of SNP309 status on age at cancer onset as well as cancer risk have, however, been conflicting [10–17].
Recently, we reported a second MDM2 promoter P2 polymorphism, SNP285G>C; rs117039649 [18]. SNP285 is in complete linkage disequilibrium (LD) with SNP309; thus forming a distinct SNP285C/309G haplotype [18, 19]. The SNP285C allele diminish Sp1 binding to the MDM2 promoter, and has been found to be associated with reduced risk for breast, ovarian and endometrial cancer [18, 20–22], but was not associated with prostate or lung cancer risk [20, 21, 23].
del1518 (rs3730485), is an insertion/deletion polymorphism of 40 bps in the MDM2 promoter P1 [24, 25]. The del1518 del-allele has been shown to reduce transcription from the MDM2 P1 promoter in some cell lines [26] and LD analysis has shown that the del1518 locus has a high LD with the SNP309 locus [24]. Some small studies have assessed the potential effects of the del1518-variant on cancer risk, and reported that the del1518 del allele may be associated with an increased risk for hepatocellular carcinoma [27] but to be unrelated to risk for several other cancer forms, including cancer of the lung [24], breast [25] ovary [28] or esophagus [29] in Chinese populations.
In the present case-control study we compared del1518 distribution across ethnic cohorts (Caucasians, African Americans and Han Chinese) and assessed the association of del1518 status with the risk for breast-, lung-, colon- and prostate cancer.
RESULTS
MDM2 del1518 was determined in 7,081 Norwegian patients diagnosed with incidental cancers of the breast, colon, prostate or lung as well as 3,749 age-matched healthy individuals. In addition, to assess potential ethnic differences in del1518 distribution, we analyzed a cohort of 300 healthy African-Americans, and performed data mining from previously published del1518 genotyping of Chinese individuals (including 2,594 Chinese samples) [24, 25, 27–29].
Distribution of del1518
Among the 3,749 healthy Caucasian (Norwegian) individuals, we found the MAF of del1518 to be 0.42 and the genotype distribution to be in Hardy-Weinberg equilibrium (p = 0.5). Interestingly, this differed (p = 0.015) from the distribution observed in African Americans (MAF = 0.38). It also differed (p < 1x10-5) from the distribution previously reported in Chinese populations (MAF = 0.30) [24, 25, 27–29] (comparison of genotype distribution between all three populations, p < 1x10-5; Figure 1A).
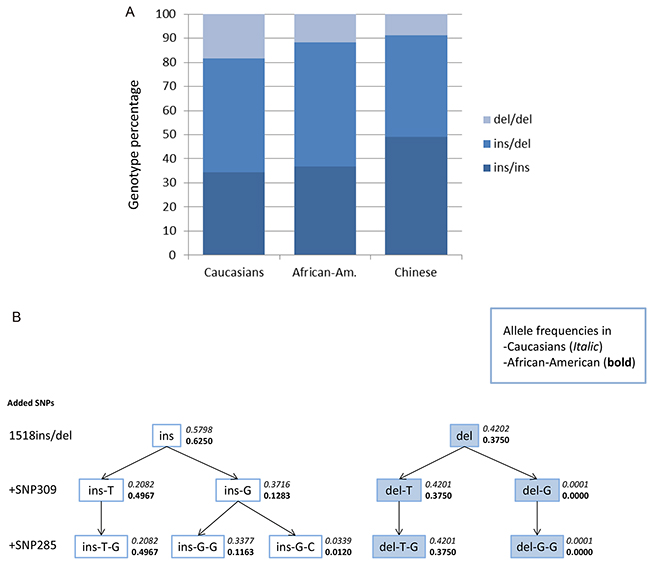
Figure 1: Genotype distribution and haplotype-tree. A. Genotype distribution of MDM2 SNP del1518 among Caucasians, African Americans and Chinese (Chinese data extracted from previous publications [24, 25, 27–29]). B. Haplotype-tree based on the three MDM2 SNPs; del1518 (rs3730485), SNP309 (rs2279744) and SNP285 (rs117039649). The tree and the frequencies given for each haplotype are based on 3,749 healthy controls in the CONOR cohort and 300 healthy African Americans.
Except for five individuals harboring the del1518 del/del – SNP309TG genotype and two individuals harboring the del1518 ins/del – SNP309GG (in the Caucasian cohort), the del1518 del variant was restricted to individuals carrying the SNP309TT or SNP309TG genotype, and del1518 del/del homozygosity was only seen among individuals carrying the SNP309TT genotype (Table 1; Figure 1B). Thus, we observed a very strong linkage disequilibrium (LD) between the del1518 variant and SNP309 (D’=0.999 for the healthy controls and D’ = 0.997 for the entire sample set, including cancer patients). This observation is in accordance with previous findings in a Chinese population [24].
Table 1: Distribution of MDM2 del1518 – SNP309 haplotypes (alleles)
Haplotype |
Healthy controls |
Breast cancer |
Colon cancer |
Lung cancer |
Prostate cancer |
|||||
|---|---|---|---|---|---|---|---|---|---|---|
del1518 - 309 |
N |
% |
n |
% |
n |
% |
n |
% |
n |
% |
ins – T |
1561 |
20,82 |
675 |
19,66 |
646 |
21,08 |
572 |
21,49 |
1058 |
21,15 |
ins – G |
2786 |
37,16 |
1296 |
37,74 |
1085 |
35,41 |
946 |
35,54 |
1854 |
3707 |
del – T |
3150 |
42,01 |
1463 |
42,60 |
1331 |
43,44 |
1143 |
42,94 |
2087 |
41,72 |
del – G |
1 |
0,01 |
0 |
0 |
2 |
0,007 |
1 |
0,04 |
3 |
0,06 |
Total |
7498 |
100 |
3434 |
100 |
3064 |
100 |
2662 |
100 |
5002 |
100 |
MDM2 del1518 and cancer risk
In order to assess the potential impact of del1518-status on cancer risk, we first performed in silico analyses of putative de novo transcription factor binding sites created by the deletion. Applying the prediction algorithm in the JASPAR database [30] with default settings (cut off threshold of 80%), we found the breakpoint of the del-variant, with flanking nucleotides, to generate new binding sites for RORA (RAR-Related Orphan Receptor A), MEF2A (Myocyte Enhancer Factor 2A) and MIZF (MBD2 (methyl-CpG-binding protein)-interacting zinc finger protein) (Figure 2).
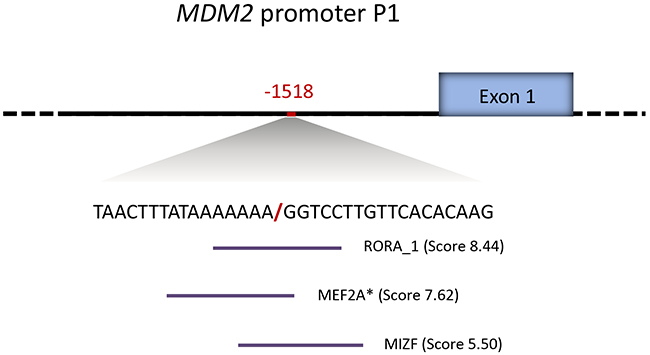
Figure 2: MDM2 promoter P1 del1518 break-point sequence context. Novel transcription factor binding sites generated by the MDM2 del1518 del allele, as predicted by the JASPAR database (indicated by horizontal lines under the sequence). The predicted binding scores are annotated in the brackets. * indicates binding sites located on the reverse strand.
To determine the potential correlation between MDM2 del1518 status and cancer risk, we analyzed the distribution of this variant in large sample sets of incident cases of four major cancer forms (1,717 breast, 1,532 colon, 1,331 lung and 2,501 prostate cancer samples) drawn from the population-based Cohort of Norway (CONOR) [31] and compared the distribution to 3,749 matched controls without any cancer from the same cohort. The genotype distributions and the results from calculations of Odds Ratios (OR) between patients and controls are summarized in Table 2.
Table 2: MDM2 del1518 distribution and cancer risk (OR)
Cases/controls |
Genotype del1518 n (%) |
OR (95% CI) del1518 |
Fisher exact |
OR (95% CI) del1518 |
Fisher exact |
||
|---|---|---|---|---|---|---|---|
ins/ins |
ins/del |
del/del |
Dominant modela |
Recessive modelb |
|||
Healthy Controls |
1285 (34.3) |
1777 (47.4) |
687 (18.3) |
1.00 |
- |
1.00 |
- |
Women |
636 (34.0) |
877 (46.9) |
359 (19.2) |
1.00 |
- |
1.00 |
- |
Men |
649 (34.6) |
900 (47.9) |
328 (17.5) |
1.00 |
- |
1.00 |
- |
Colon cancer |
478 (31.2) |
775 (50.6) |
279 (18.2) |
1.15 (1.01-1.31) |
0.034 |
0.99 (0.85-1.16) |
0.938 |
Women |
237 (30.4) |
393 (50.5) |
149 (19.1) |
1.18 (0.98-1.41) |
0.077 |
1.00 (0.81-1.23) |
1.000 |
Men |
241 (32.0) |
382 (50.7) |
130 (17.3) |
1.12 (0.94-1.35) |
0.218 |
0.99 (0.79-1.23) |
0.955 |
Left side |
190 (30.2) |
331 (52.5) |
109 (17.3) |
1.21 (1.01-1.45) |
0.045 |
0.93 (0.75-1.17) |
0.577 |
Right side |
261 (31.5) |
410 (49.5) |
158 (19.1) |
1.14 (0.97-1.33) |
0.133 |
1.05 (0.87-1.27) |
0.621 |
Lung cancer |
447 (33.6) |
624 (46.9) |
260 (19.5) |
1.03 (0.90-1.18) |
0.662 |
1.08 (0.92-1.27) |
0.346 |
Women |
155 (31.2) |
247 (49.7) |
95 (19.1) |
1.14 (0.92-1.40) |
0.261 |
1.00 (0.78-1.28) |
1.000 |
Men |
292 (35.0) |
377 (45.2) |
165 (19.8) |
0.98 (0.83-1.16) |
0.827 |
1.17 (0.95-1.43) |
0.161 |
Breast cancerc |
581 (33.8) |
809 (47.1) |
327 (19.0) |
1.01 (0.88-1.16) |
0.944 |
0.99 (0.84-1.17) |
0.932 |
Prostate cancerd |
836 (33.4) |
1240 (49.6) |
425 (17.0) |
1.05 (0.93-1.19) |
0.439 |
1.02 (0.85-1.22) |
0.853 |
adel/del + ins/del versus ins/ins
bdel/del versus ins/del + ins/ins
ccompared to female controls only
dcompared to male controls only
In overall assessments applying a dominant model, the del1518 del allele was associated with an increased risk of colon cancer (OR = 1.15; 95 % CI = 1.01 – 1.31; Table 2, Figure 3, Supplementary Table S1). Notably, this association was more pronounced among patients with left sided colon cancer (dominant model: OR = 1.21; 95 % CI = 1.01 - 1.45), compared to patients with right sided tumors (dominant model: OR = 1.14; 95 % CI = 0.97 – 1.33; Table 2, Figure 3, Supplementary Table S1). For the three other cancer forms, no statistically significant association between del1518 status and cancer risk was recorded (Table 2, Figure 3, Supplementary Table S1).
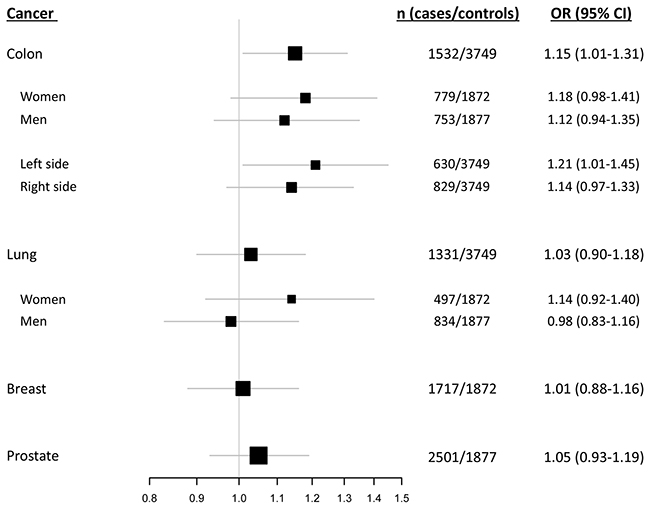
Figure 3: del1518 polymorphism and cancer risk. Forest plot illustrating the effect of the MDM2 del1518 polymorphism on the risk of breast-, colon-, lung- and prostate cancer, applying the dominant model (comparison of del1518 del/del + del/ins genotypes versus the ins/ins genotype).
Potential interactions between MDM2 del1518 status and MDM2 SNP309
In the samples investigated in this study we have previously reported the MDM2 SNP309GG/TG genotypes to be associated with reduced risk of lung cancer among women, but not associated with risk of breast-, colon- or prostate cancer [20]. Given that the del1518 del was strongly linked to the SNP309T allele, we further refined our analyses by removing individuals harboring the SNP309GG genotype and assessed the effect of del1518 separately in individuals harboring the SNP309TG or SNP309TT genotype. Among individuals carrying the SNP309TT genotype, no association was recorded in either analysis (Table 3A, Figure 4A). However, among individuals with the SNP309TG genotype, the del1518 del allele was associated with increased risk for colon cancer (OR = 1.21; 95 % CI = 1.01 – 1.46; Table 3B, Figure 4B), indicating a possible synergism between the del1518 del-allele and the SNP309G-allele.
Table 3A: MDM2 del1518 among SNP309TT
Cases/controls |
Genotype del1518 n (%) |
OR (95% CI) del1518 |
Fisher exact |
OR (95% CI) del1518 |
Fisher exact |
||
|---|---|---|---|---|---|---|---|
ins/ins |
ins/del |
del/del |
Dominant modela |
Recessive modelb |
|||
Healthy Controls |
155 (10.6) |
623 (42.6) |
686 (46.9) |
1.00 |
- |
1.00 |
|
Women |
70 (12.3) |
311 (41.1) |
359 (46.6) |
1.00 |
- |
1.00 |
|
Men |
85 (10.2) |
312 (42.9) |
327 (46.9) |
1.00 |
- |
1.00 |
|
Colon cancer |
71 (11.3) |
283 (44.9) |
277 (43.9) |
0.93 (0.69-1.26) |
0.646 |
0.89 (0.74-1.07) |
0.958 |
Women |
34 (10.6) |
140 (43.5) |
148 (46.0) |
0.89 (0.57-1.36) |
0.576 |
0.90 (0.69-1.17) |
0.463 |
Men |
37 (12.0) |
142 (46.3) |
129 (4.8) |
0.98 (0.65-1.48) |
0.916 |
0.87 (0.67-1.14) |
0.338 |
Lung cancer |
63 (11.1) |
245 (43.2) |
259 (45.7) |
0.95 (0.70-1.29) |
0.749 |
0.95 (0.79-1.16) |
0.655 |
Women |
23 (10.1) |
110 (48.3) |
95 (41.7) |
0.93 (0.57-1.53) |
0.797 |
0.76 (0.56-1.02) |
0.081 |
Men |
40 (11.8) |
135 (39.8) |
164 (48.4) |
0.99 (0.67-1.48) |
1.000 |
1.14 (0.88-1.47) |
0.355 |
Breast cancerc |
66 (9.8) |
279 (41.5) |
327 (48.7) |
1.00 (0.67-1.37) |
0.857 |
1.01 (0.82-1.24) |
0.958 |
Prostate cancerd |
108 (10.9) |
456 (46.2) |
424 (42.9) |
1.08 (0.80-1.47) |
0.643 |
0.91 (0.75-1.11) |
0.375 |
adel/del + ins/del versus ins/ins
bdel/del versus ins/del + ins/ins
ccompared to female controls only
dcompared to male controls only
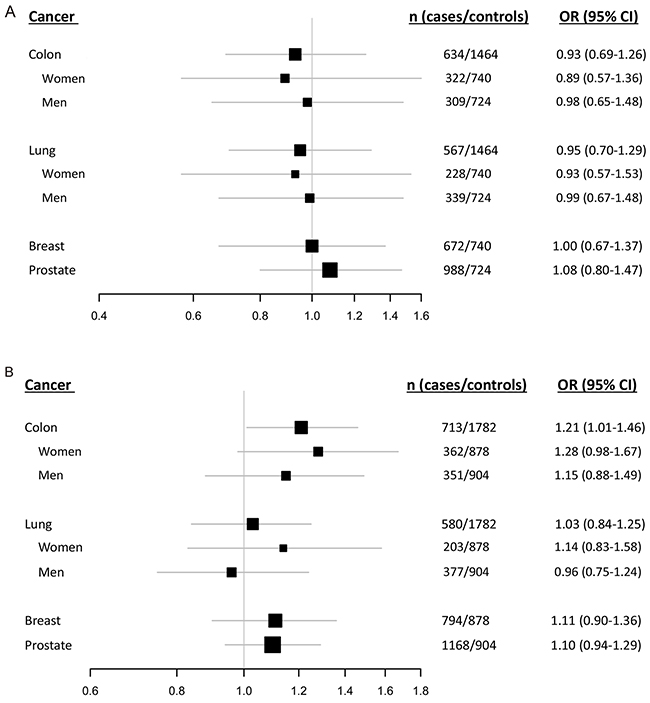
Figure 4: Impact of the del1518 polymorphism on cancer risk among SNP309. Forest plots illustrating the effect of the MDM2 del1518 polymorphism on the risk of breast-, colon-, lung- and prostate cancer, applying the dominant model (comparison of del1518 del/del + del/ins genotypes versus the ins/ins genotype) among individuals carrying the SNP309TT genotype A. and (del1518 del/ins versus ins/ins) among individuals harboring the SNP309TG genotype B. (Due to the strong linkage between the del1518 del-allele and the SNP309T-allele, the group of homozygous del/del was non-evaluable within individuals with the SNP309TG-genotype).
Table 3B: MDM2 del1518 among SNP309TG
Cases/controls |
Genotype del1518 n (%) |
OR (95% CI) del1518 |
Fisher exact |
||
|---|---|---|---|---|---|
ins/ins |
ins/del |
del/dela |
Ins/del vs. ins/ins |
||
Healthy Controls |
628 (35.2) |
1154 (64.8) |
- |
1.00 |
- |
Women |
312 (35.5) |
566 (64.5) |
|||
Men |
316 (35.0) |
588 (65.0) |
|||
Colon cancer |
221 (31.0) |
492 (69.0) |
- |
1.21 (1.01-1.46) |
0.044 |
Women |
109 (30.1) |
253 (69.9) |
1.28 (0.98-1.67) |
0.075 |
|
Men |
112 (31.9) |
239 (68.1) |
1.15 (0.88-1.49) |
0.320 |
|
Lung cancer |
201 (34.7) |
379 (65.3) |
- |
1.03 (0.84-1.25) |
0.841 |
Women |
66 (32.5) |
137 (67.5) |
1.14 (0.83-1.58) |
0.462 |
|
Men |
135 (35.8) |
242 (64.2) |
0.96 (0.75-1.24) |
0.797 |
|
Breast cancerb |
264 (33.3) |
530 (66.8) |
- |
1.11 (0.90-1.36) |
0.328 |
Prostate cancerc |
386 (33.1) |
782 (67.0) |
- |
1.10 (0.94-1.29) |
0.234 |
ano observations since del1518 del is linked to SNP309T
bcompared to female controls only
ccompared to male controls only
Similar subgroup analyses of the three remaining cancer forms showed no association between del1518 status and risk for malignancy at any site (Table 3A–3B, Figure 4A–4B).
DISCUSSION
The findings by Levine’s group [8] and our group [18], suggesting that both the MDM2 promoter SNP’s 309 and 285 may have an effect on cancer risk, implies the importance of fine tuning of MDM2 levels with respect to carcinogenesis, and also indicate that other MDM2 promoter variants could potentially be of importance.
Assessments of the promoter P1 del1518 variant have previously been performed in a few relatively small Chinese cohorts [24, 25, 27–29]. In the present study, we determined the distribution of del1518 status in a large Caucasian cohort. A major strength of our study is the large number of cancer patients and controls compared to other studies assessing del1518-status with respect to cancer risk. Also, the population based design, using incident cancers (breast, lung, colon and prostate) for whom DNA samples were collected years prior to diagnosis and controls matched by gender, age, and residential area, is a strong feature.
In line with previous risk assessments in Chinese populations [24, 25, 28], we found no association between del1518 status and risk for cancer of the breast, lung or prostate. We did, however, observe a positive association with colon cancer risk. Somewhat surprisingly, we found an effect on colon cancer among individuals heterozygous, but not homozygous, for del1518. However, as the del1518 variant is linked to the SNP309T allele, homozygosity for del1518 is restricted to individuals harboring the SNP309TT genotype. Additionally, after stratifying our data according to SNP309 status, we found heterozygousity for the del1518 del allele to be associated with an increased risk of colon cancer among individuals carrying the SNP309TG genotype, but not among SNP309TT homozygous individuals. Although the statistical significance was borderline, this may indicate a synergistic effect of these two polymorphisms. Notably, studies on other genes have revealed that individuals harboring heterozygous mutations may be at risk of different diseases compared to homozygous carriers [32, 33]. Such effects may potentially differ between cancer forms as well; previously, we reported that the SNP285C allele significantly reduces the risk for breast cancer among SNP309GG homozygous individuals, whereas in ovarian cancer, the effect of SNP285C was significant among SNP309TG individuals only [20, 18]. Here, the possibility exists that del1518 may be detrimental in concert with SNP309G heterozygosity (individuals harboring the SNP309TG genotype), but not exert a similar effect among SNP309TT carriers.
Left sided and right sided colon cancers are known to be different with respect to histological and molecular characteristics [34], and also the genetic mechanisms initiating the tumorigenic process have been found to be dissimilar [35]. There are evidence that hereditary nonpolyposis colon cancer (HNPCC) in general are found in right sided colon cancer [36], while familial adenomatous polyposis (FAP) dominates in left sided colon cancer [37]. However, although we only observed an increased OR among individuals suffering from left sided colon cancer, a clear trend for increased risk was observed also among patients harboring right sided colon cancer. The lack of statistical significance in this group may be a result of limited statistical power. Thus, whether the del1518 polymorphism is associated with increased risk for both left sided – and right sided colon cancer may need to be explored in independent populations.
Previously, we showed that MDM2 SNP285 is part of a predicted estrogen response element (ERE), where both Sp1 and ERα may bind and affect tumorigenesis [21]. However, when assessing SNP285 and cancer risk among “gender neutral” cancer forms (i.e., lung and colon cancer), we observed only moderate gender differences [20]. In the present study, corresponding to our previous findings regarding SNP285, we found no gender specific effect of del1518 on cancer risk in either cancer form. Taken together, these data indicate that the effect on cancer risk of the germline variations in the MDM2 promoter regions, in general, are more dependent on tissue type than on gender.
In conclusion, we find an association of del1518 with the risk of colorectal cancer and no association for cancer of the prostate, breast or lung. The finding for colon cancer, however, warrants confirmation by others.
MATERIALS AND METHODS
Study populations
To assess potential impact of del1518 status on cancer risk, we genotyped this variant in blood DNA from 10,830 individuals drawn from the Norwegian population-based Cohort of Norway (CONOR) study [31], previously described in [20]. This sample set included 3,749 cancer-free controls as well as incident cases of four major cancer forms: cancer of the breast (n=1,717), prostate (n=2,501), lung (n=1,331) and colon (n=1,532).
The African-American cohort (n=300) was from the Laboratory of Human Carcinogenesis, Center for Cancer Research, National Cancer Institute Bethesda, USA. These samples were collected as part of an ongoing case control study with previously described inclusion criteria [38]. Briefly, population controls were identified from the Department of Motor Vehicles, MD, USA and frequency matched to cases by age and gender. Written informed consent was obtained from all participants, and the study was approved by the Institutional Review Boards of the participating institutions.
In silico predictions
Potential novel binding sites for transcription factors, generated by the deletion variant of the MDM2 del1518 polymorphism, were predicted using the JASPAR database [30] (http://jaspardev.genereg.net). Predictions were made using the default settings of the database (cut off threshold of 80%).
MDM2 del1518 genotyping
All samples were genotyped for MDM2 del1518 by using DNA extracted from peripheral white blood cells. The region of the MDM2 promoter P1 containing the del1518 was amplified by PCR using the VWR Taq DNA Polymerase system (VWR), with a primer pair previously described [27]. The amplification was performed in a total reaction volume of 25 µl, containing 2.5 µl 10x Key Buffer, 0.2 mM dNTPs, 0.2 µM each primer, 1.25 U Taq polymerase and 1 µl template DNA (10-100 µg). The thermo-cycling conditions were an initial step of 94 °C for 5 min, 35 cycles of 94 °C for 30 sec, 58 °C for 30 sec and 72 °C for 30 sec, followed by a final elongation step of 72 °C for 10 min. The PCR products were separated by electrophoresis in a 3% agarose gel and visualized by GelRedTM Nucleic Acid Gel Stain (BIOTIUM). The del1518 insertion and deletion alleles were observed as 481 bps – and 441 bps bands respectively.
All samples were previously genotyped for MDM2 SNP285 and SNP309 status [20] using custom LightSNiP assays (TIB MOLBIOL Syntheselabor GmbH, Berlin, Germany) as described in detail elsewhere [39].
Statistical analysis
Potential deviations from Hardy-Weinberg equilibrium in cancer patients as well as healthy controls were evaluated using the Chi-square test. Possible associations between MDM2 del1518 and cancer risk, both in total and in stratified groups, were estimated by Odds Ratios (OR) and Fisher exact tests. ORs are given with 95% confidence intervals (CI) and p-values from Fisher exact tests are given as two-sided and cumulative.
All statistical analyses were performed using the IBM SPSS statistics (version 19) software package.
ACKNOWLEDGMENTS
We thank Beryl Leirvaag for technical assistance. Most of this work was performed in the Mohn Cancer Research Laboratory.
CONFLICTS OF INTEREST
The authors declared no conflict of interest.
GRANT SUPPORT
This study was supported by grants from the Norwegian Cancer Society, the Norwegian Health Region West funding body, the Norwegian Research Council and the Bergen Research Foundation.
REFERENCES
1. Haupt Y, Maya R, Kazaz A, Oren M. Mdm2 promotes the rapid degradation of p53. Nature. 1997;387:296-9.
2. Honda R, Tanaka H, Yasuda H. Oncoprotein MDM2 is a ubiquitin ligase E3 for tumor suppressor p53. FEBS Letters. 1997;420:25-7.
3. Kubbutat MHG, Jones SN, Vousden KH. Regulation of p53 stability by Mdm2. Nature. 1997;387:299-303.
4. Momand J, Zambetti GP, Olson DC, George D, Levine AJ. The mdm-2 oncogene product forms a complex with the p53 protein and inhibits p53-mediated transactivation. Cell. 1992;69:1237-45.
5. Landers JE, Cassel SL, George DL. Translational enhancement of mdm2 oncogene expression in human tumor cells containing a stabilized wild-type p53 protein. Cancer Res. 1997;57:3562-8.
6. Momand J, Jung D, Wilczynski S, Niland J. The MDM2 gene amplification database. Nucleic Acids Res. 1998;26:3453-9.
7. Oliner JD, Kinzler KW, Meltzer PS, George DL, Vogelstein B. Amplification of a gene encoding a p53-associated protein in human sarcomas. Nature. 1992;358:80-3.
8. Bond GL, Hu W, Bond EE, Robins H, Lutzker SG, Arva NC, Bargonetti J, Bartel F, Taubert H, Wuerl P, Onel K, Yip L, Hwang S-J et al. A Single Nucleotide Polymorphism in the MDM2 Promoter Attenuates the p53 Tumor Suppressor Pathway and Accelerates Tumor Formation in Humans. Cell. 2004;119:591-602.
9. Bond GL, Hirshfield KM, Kirchhoff T, Alexe G, Bond EE, Robins H, Bartel F, Taubert H, Wuerl P, Hait W, Toppmeyer D, Offit K, Levine AJ. MDM2 SNP309 Accelerates Tumor Formation in a Gender-Specific and Hormone-Dependent Manner. Cancer Res. 2006;66:5104-10.
10. Bai J, Dai J, Yu H, Shen H, Chen F. Cigarette smoking, MDM2 SNP309, gene-environment interactions, and lung cancer risk: a meta-analysis. Journal of toxicology and environmental health Part A. 2009;72:677-82.
11. Economopoulos KP, Sergentanis TN. Differential effects of MDM2 SNP309 polymorphism on breast cancer risk along with race: a meta-analysis. Breast cancer research and treatment. 2010;120:211-6.
12. Hu Z, Jin G, Wang L, Chen F, Wang X, Shen H. MDM2 promoter polymorphism SNP309 contributes to tumor susceptibility: evidence from 21 case-control studies. Cancer epidemiology, biomarkers & prevention. 2007;16:2717-23.
13. Liu G, Jiang D, Shen S, Yu L. Murine double minute 2 promoter SNP309 polymorphism and prostate cancer risk: a meta-analysis. International journal of urology. 2012;19:914-20.
14. Wan Y, Wu W, Yin Z, Guan P, Zhou B. MDM2 SNP309, gene-gene interaction, and tumor susceptibility: an updated meta-analysis. BMC Cancer. 2011;11:208.
15. Wilkening S, Bermejo JL, Hemminki K. MDM2 SNP309 and cancer risk: a combined analysis. Carcinogenesis. 2007;28:2262-7.
16. Zhao E, Cui D, Yuan L, Lu W. MDM2 SNP309 polymorphism and breast cancer risk: a meta-analysis. Molecular biology reports. 2012;39:3471-7.
17. Gui XH, Qiu LX, Zhang HF, Zhang DP, Zhong WZ, Li J, Xiao YL. MDM2 309 T/G polymorphism is associated with lung cancer risk among Asians. Eur J Cancer. 2009;45:2023-6.
18. Knappskog S, Bjornslett M, Myklebust LM, Huijts PE, Vreeswijk MP, Edvardsen H, Guo Y, Zhang X, Yang M, Ylisaukko-Oja SK, Alhopuro P, Arola J, Tollenaar RA et al. The MDM2 promoter SNP285C/309G haplotype diminishes Sp1 transcription factor binding and reduces risk for breast and ovarian cancer in Caucasians. Cancer Cell. 2011;19:273-82.
19. Paulin F, O’Neill M, McGregor G, Cassidy A, Ashfield A, Ali C, Munro A, Baker L, Purdie C, Lane D, Thompson A. MDM2 SNP309 is associated with high grade node positive breast tumours and is in linkage disequilibrium with a novel MDM2 intron 1 polymorphism. BMC Cancer. 2008;8:281.
20. Gansmo LB, Knappskog S, Romundstad P, Hveem K, Vatten L, Lonning PE. Influence of MDM2 SNP309 and SNP285 status on the risk of cancer in the breast, prostate, lung and colon. Int J Cancer. 2015;137:96-103.
21. Knappskog S, Trovik J, Marcickiewicz J, Tingulstad S, Staff AC, MoMa TECsg, Romundstad P, Hveem K, Vatten L, Salvesen HB, Lonning PE. SNP285C modulates oestrogen receptor/Sp1 binding to the MDM2 promoter and reduces the risk of endometrial but not prostatic cancer. Eur J Cancer. 2012;48:1988-96.
22. Piotrowski P, Lianeri M, Rubis B, Knula H, Rybczynska M, Grodecka-Gazdecka S, Jagodzinski PP. Murine double minute clone 2,309T/G and 285G/C promoter single nucleotide polymorphism as a risk factor for breast cancer: a Polish experience. The International journal of biological markers. 2012;27:e105-10.
23. Ryan BM, Calhoun KM, Pine SR, Bowman ED, Robles AI, Ambs S, Harris CC. MDM2 SNP285 does not antagonize the effect of SNP309 in lung cancer. Int J Cancer. 2012;131:2710-6.
24. Hu Z, Ma H, Lu D, Qian J, Zhou J, Chen Y, Xu L, Wang X, Wei Q, Shen H. Genetic variants in the MDM2 promoter and lung cancer risk in a Chinese population. Int J Cancer. 2006;118:1275-8.
25. Ma H, Hu Z, Zhai X, Wang S, Wang X, Qin J, Jin G, Liu J, Wei Q, Shen H. Polymorphisms in the MDM2 promoter and risk of breast cancer: a case-control analysis in a Chinese population. Cancer letters. 2006;240:261-7.
26. Lalonde ME, Ouimet M, Lariviere M, Kritikou EA, Sinnett D. Identification of functional DNA variants in the constitutive promoter region of MDM2. Human genomics. 2012;6:15.
27. Dong D, Gao X, Zhu Z, Yu Q, Bian S, Gao Y. A 40-bp insertion/deletion polymorphism in the constitutive promoter of MDM2 confers risk for hepatocellular carcinoma in a Chinese population. Gene. 2012;497:66-70.
28. Kang S, Wang DJ, Li WS, Wang N, Zhou RM, Sun DL, Duan YN, Li SZ, Li XF, Li Y. Association of p73 and MDM2 polymorphisms with the risk of epithelial ovarian cancer in Chinese women. International journal of gynecological cancer. 2009;19:572-7.
29. Ma J, Zhang J, Ning T, Chen Z, Xu C. Association of genetic polymorphisms in MDM2, PTEN and P53 with risk of esophageal squamous cell carcinoma. Journal of human genetics. 2012;57:261-4.
30. Sandelin A, Alkema W, Engstrom P, Wasserman WW, Lenhard B. JASPAR: an open-access database for eukaryotic transcription factor binding profiles. Nucleic Acids Res. 2004;32:D91-4.
31. Naess O, Sogaard AJ, Arnesen E, Beckstrom AC, Bjertness E, Engeland A, Hjort PF, Holmen J, Magnus P, Njolstad I, Tell GS, Vatten L, Vollset SE et al. Cohort profile: cohort of Norway (CONOR). Int J Epidemiol. 2008;37:481-5.
32. di Masi A, Antoccia A. NBS1 Heterozygosity and Cancer Risk. Current Genomics. 2008;9:275-81.
33. Smirnov DA, Cheung VG. ATM gene mutations result in both recessive and dominant expression phenotypes of genes and microRNAs. Am J Hum Genet. 2008;83:243-53.
34. Iacopetta B. Are there two sides to colorectal cancer? International Journal of Cancer. 2002;101:403-8.
35. Albuquerque C, Bakker ER, van Veelen W, Smits R. Colorectal cancers choosing sides. Biochim Biophys Acta. 2011;1816:219-31.
36. Rijcken FEM, Hollema H, Kleibeuker JH. Proximal adenomas in hereditary non-polyposis colorectal cancer are prone to rapid malignant transformation. Gut. 2002;50:382-6.
37. Lynch HT, Smyrk T. Hereditary nonpolyposis colorectal cancer (Lynch syndrome): An updated review. Cancer. 1996;78:1149-67.
38. Robles AI, Yang P, Jen J, McClary AC, Calhoun K, Bowman ED, Vähäkangas K, Greathouse KL, Wang Y, Olivo-Marston S, Wenzlaff AS, Deng B, Schwartz AG et al. A DRD1 Polymorphism Predisposes to Lung Cancer among Those Exposed to Secondhand Smoke during Childhood. Cancer Prevention Research. 2014;7:1210-8.
39. Knappskog S, Gansmo LB, Dibirova K, Metspalu A, Cybulski C, Peterlongo P, Aaltonen LA, Vatten L, Romundstad P, Hveem K, Devilee P, Evans DG, Lin D et al. Population distribution and ancestry of the cancer protective MDM2 SNP285 (rs117039649). Oncotarget. 2014;5:8223-34. doi: 10.18632/oncotarget.1910.