
INTRODUCTION
Cancer causes a wide variety of diseases and is characterized by uncontrolled growth and spread of abnormal cells and subsequent disruption of organ and tissue functions. Treatments for cancer usually include surgery followed by radiotherapies and chemotherapies [1] that inhibit cancer cell growth and proliferation. However, in contrast with localized surgery and radiotherapy, chemotherapy has systemic effects [2–4]. Among chemotherapeutic agents, phytochemicals, such as paclitaxel from the Pacific yew tree and vincristine from Vinca rosea Linn, are widely used to treat cancers and have demonstrated pro-apoptotic effects [5, 6]. Various chemopreventive and chemotherapeutic agents have been shown to induce apoptotic cell death in both in vitro and in vivo studies, suggesting that apoptosis plays a crucial role in cancer treatment [7]. Accordingly, the widely used chemotherapeutic drug 5-fluorouracil (5-FU) inhibits tumor cell growth in animal models by inducing apoptotic activation of the CD95/CD95L system [8]. The chemopreventive agent curcumin predominantly induces apoptosis via mitochondria-mediated pathways in various cancer cell types [9]. Apoptosis, or programmed cell death, is a physiological process that eliminates abnormal, misplaced, or nonfunctional cells and is critical for maintenance of tissue homeostasis [10]. Excessive apoptosis causes organ atrophy and dysfunction, whereas failure of apoptosis results in accumulation of abnormal cells, potentially leading to tumor development. Apoptosis is controlled at multiple molecular levels and involves pro- and anti-apoptotic members of the Bcl-2 protein family [11]. Many studies have shown that dietary phytochemicals induce apoptosis in cancer cells, suggesting potential for development as cancer therapeutic agents [12].
Mangrove forests are economically and ecologically important and are rich in medicinal and non-medicinal edible plants. In particular, mangroves produce a wide variety of structurally novel natural agents with biochemical profiles [13]. Avicennia marina is a mangrove species of the Acanthaceae family, and discoveries of its chemical compounds have received much attention [14]. A. marina has been used as a traditional medicine for the treatment of skin diseases, rheumatism, ulcers, and smallpox. In vitro antimalarial, antibacterial, analgesic, and cytotoxic activities of A. marina have been reported [15]. Hence, A. marina is considered a valuable source of chemical constituents with medicinal potential. Among these, luteolin 7-O-methylether, chrysoeriol 7-O-glucoside, isorhamnetin 3-O-rutinoside, tannin lapachol, and naphthoquinone analogs have been isolated, and their biological activities have been described [16]. However, few studies demonstrate in vitro and in vitro anti-cancer activities of A. marina, and its chemical constituents have not been extensively examined.
Here we evaluated the biomedical potential of A. marina plants from a mangrove forest and established the chemical composition of A. marina extracts. In particular, total phytopolyphenol contents were separated using chromatography, and A. marina fractions were evaluated for anti-cancer effects in in vitro and in vivo models.
RESULTS
Polyphenol contents and anticancer activities against breast and liver cancer cell lines
Plant extracts that are rich in polyphenols have been safely used as traditional Chinese medicines for many centuries. Thus, water (H2O), ethanol (EtOH), methanol (MeOH), and ethyl acetate (EtOAc) extracts of A. marina were evaluated for phenol and flavonoid contents. As shown in Table 1A, EtOAc extracts of A. marina leaves had the highest phenol (80.96 ± 0.78 mg/g) and flavonoid (18.6 ± 2.01 mg/g) contents, followed by H2O and MeOH extracts of A. marina leaves. Similarly, EtOAc extracts of A. marina seeds were richer in phenols and flavonoids than H2O and MeOH extracts. Although mineral contents in A. marina leaves have not been determined previously, these were similar to those reported in previous studies of medicinal plants. Specifically, inductively coupled plasma atomic emission spectroscopy (ICP-AES) revealed the presence of the trace metal elements lead (Pb), zinc (Zn), nickel (Ni), indium (In), iron (Fe), aluminum (Al), arsenic (As), copper (Cu), cadmium (Cd), chromium (Cr), and silver (Ag) in A. marina leaves that were collected from the Xinfeng mangrove conservation area in Taiwan. Average concentrations of Rb, Zn, Ni, In, Fe, and Al in A. marina leaves were 10.3, 15.5, 2.7, 2.6, 128.7, and 93.3 mg/kg, respectively (Table 1B). The absence of detectable As, Cu, Cd, Cr, and Ag in the present A. marina leaves was considered favorable for clinical application without toxicity.
Table 1A: Total phenol and flavonoid in Avicennia marina leaves extraction
|
A. marina Extracts |
||||||
|---|---|---|---|---|---|---|---|
Content of phytochemicals (mg/g) |
Leaf |
Seed |
|||||
MeOH |
EtOH |
H2O |
EtOAc |
EtOH |
H2O |
EtOAc |
|
Total phenols |
46.96 |
22.82 |
47.06 |
80.96 |
49.96 |
36.08 |
82.23 |
± 0.24 |
± 1.80 |
± 2.15 |
± 0.78 |
± 3.85 |
± 6.85 |
± 1.12 |
|
Total flavonoids |
9.26 |
11.96 |
6.83 |
18.69 |
3.15 |
1.47 |
4.72 |
± 1.05 |
± 3.16 |
± 1.57 |
± 2.01 |
± 1.02 |
± 0.08 |
± 0.58 |
|
Table 1B: Concentrations of trace metal elements in Avicennia marina leaves
|
Ag |
Al |
As |
Cu |
Cd |
Cr |
Fe |
Ni |
In |
Rb |
Zn |
|---|---|---|---|---|---|---|---|---|---|---|---|
(mg/kg)a |
|
||||||||||
A. marina |
NDb |
93.3 |
ND |
ND |
ND |
ND |
128.7 |
2.7 |
2.6 |
10.3 |
12.7 |
aConcentration of trace metal elements (mg/kg dry matter) in Avicennia marina leaves.
bND, not detected.
To determine whether high phenol and flavonoid contents were associated with anticancer activities, cytotoxic effects of H2O, EtOH, MeOH, and EtOAc extracts of A. marina leaves were compared using MTT assays in normal NIH3T3 cells, and in breast (AU565, MDA-MB-231 and BT483) and liver (HepG2 and Huh7) cancer cell lines (Figure 1B). In these experiments, EtOH and EtOAc extracts of A. marina leaves inhibited cell growth in cancer cell lines more than in normal NIH 3T3 cells. Inhibition of cancer cell growth was greater with EtOAc extracts than with EtOH and MeOH extracts, but was dose-dependent in all cases, and similar observations were made after treatments with EtOAc extracts of A. marina seeds (data not shown). However, whereas growth inhibition by H2O extracts of A. marina leaves was significant at 1000 μg/mL, H2O extracts had no effect at 200 μg/mL (data not shown). These data suggest that EtOAc extracts of A. marina leaves have the highest polyphenol contents and anticancer activities. Accordingly, EtOAc extracts did not inhibit proliferation of NIH 3T3 cells at 40–80-μg/mL, but significantly inhibited cell proliferation in AU565, BT483, HepG2, and Huh7 cancer cells. Thus, subsequent analyses were performed after treatments with EtOAc extracts at 40–80 μg/mL.
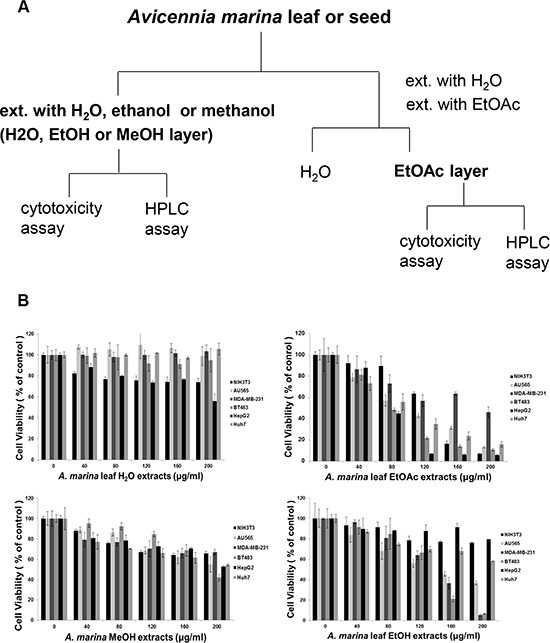
Figure 1: (A) Procedure for obtaining extracts from Avicennia marina; (B) Comparisons of cell viability after treatments with water (H2O), ethanol (EtOH), methanol (MeOH), and ethyl acetate (EtOAc) extracts of A. marina leaves; MTT assays showed differing responses of cell lines after 48-h treatments with differing extracts of A. marina leaves.
Soft agar colony formation assays can be used to separate cancer cells and to confirm phenotypes, and were used to determine the effects of EtOAc extracts of A. marina leaves on anchorage-independent growth in breast (AU565 and BT483) and liver (HepG2 and Huh7) cancer cells. After 14–21 days of treatment with the EtOAc extract, cultured cells showed reduced soft agar colony formation, with decreased colony sizes and a smaller numbers of AU565, BT483, HepG2, and Huh7 cancer cell colonies (Figure 2).
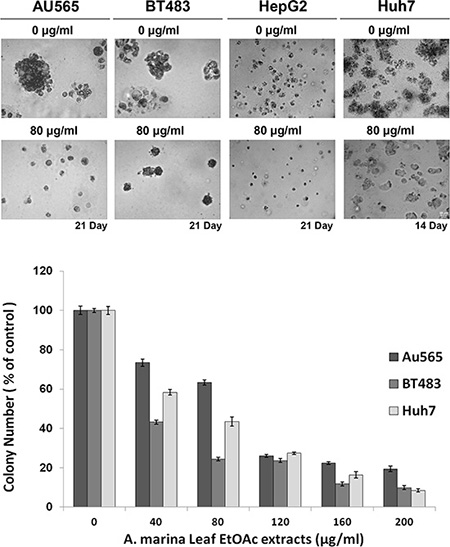
Figure 2: Effects of EtOAc extracts from A. marina leaves on anchorage-independent growth of AU565, BT483, HepG2, and Huh7 cancer cells. Cells were treated with EtOAc extracts at 40–200 μg/mL in 0.35% agarose containing 10% FCS over 0.7% agarose containing 10% FCS. Cell colonies were observed after 14–21 days incubation at 37°C in 5% CO2 under light microscopy. Bars on images represent 20 m. Colonies of > 50 μm were counted after the incubation period and numbers are presented as percentages of the control.
Chemical composition of EtOAc extracts of A. marina leaves
Polyphenol concentrations of H2O, EtOH, and EtOAc extracts of A. marina leaves were determined using HPLC, and included the 14 flavonoids apigenin, chrysin, catechin, epigallocatechin gallate (EGCG), picatechin gallate (ECG), kaempferol, luteolin, narigenin, myricetin, quercetin, rutin, resveratrol, theaflavin (TF1), and theaflavin-3-gallate (TF2), and the 7 phenolic acids caffeic acid, chlorogenic acid, p-coumaric acid, corilagin, ellagic acid, gallic acid, and syringic acid (Figure 3A and Table 2A).
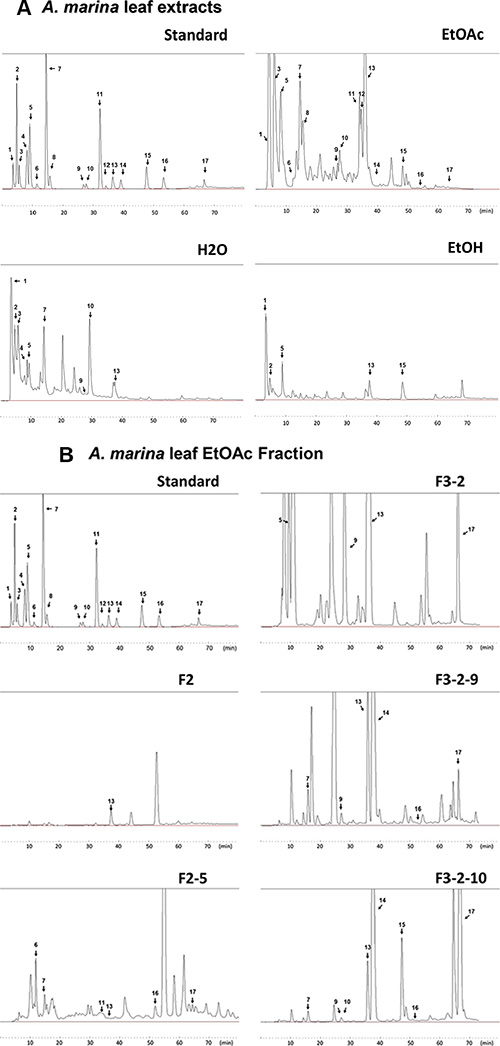
Figure 3: Analyses of polyphenol contents in A. marina leaves. (A) Polyphenol contents of H2O, EtOAc, and EtOH extracts of A. marina leaves were analyzed using HPLC; HPLC chromatograms of the polyphenol standard mixture were recorded at 280 nm; Peaks: 1, ellagic acid; 2, ellagic acid + gallic acid; 3, chlorogenic acid; 4, catechin; 5, syringic acid + epigallocatechin gallate (EGCG) + rutin + caffeic acid; 6, corilagin; 7, narigenin + p-coumaric acid; 8, picatechin gallate (ECG); 9, myricetin; 10, theaflavin (TF1); 11, resveratrol; 12, theaflavin-3-gallate (TF2); 13, luteolin; 14, quercetin; 15, apigenin; 16, kaempferol; 17, chrysin. (B) Chromatograms of fractions separated from EtOAc extracts of A. marina leaves.
Table 2A: Contents (micrograms per gram) of phenolic acids in different extracts of A. marina leaf
Peak |
Retention time (min) |
Leaf (EtOAc) |
Leaf (H2O) |
Leaf (EtOH) |
Comparison with standard |
|---|---|---|---|---|---|
1. |
3.52 |
40.367 ± 1.232 |
28.102 ± 1.369 |
19.331 ± 2.133 |
Elliagic Acid (1) |
2. |
4.72 |
ND |
1.984 ± 0.228 |
0.276 ± 0.013 |
Elliagic Acid (2), Gallic Acid |
3. |
5.55 |
38.60 ± 2.117 |
8.918 ± 0.139 |
ND |
Chlorogenic Aicd |
4. |
8.17 |
ND |
2.149 ± 1.131 |
ND |
Catechin |
5. |
9.03 |
31.120 ± 1.01 |
7.011 ± 1.237 |
6.051 ± 2.526 |
Syring Acid, EGCG, Rutin, Caffeic Acid |
6. |
11.28 |
2.123 ± 1.231 |
ND |
ND |
Corilagin |
7. |
14.35 |
10.214 ± 0.912 |
5.854 ± 0.243 |
ND |
Naringic, p-Coumaric Acid |
8. |
15.60 |
40.062 ± 2.117 |
ND |
ND |
ECG |
9. |
26.78 |
11.183 ± 0.182 |
9.985 + 2.496 |
ND |
Myricetin |
10. |
27.67 |
19.303 ± 2.362 |
39.837 ± 5.199 |
ND |
TF1 |
11. |
32.25 |
6.487 ± 0.325 |
ND |
ND |
Resveratrol |
12. |
34.22 |
156.381 ± 5.475 |
ND |
ND |
TF2 |
13. |
36.32 |
29.335 ± 2.311 |
2.298 ± 0.011 |
4.504 + 1.715 |
Luteolin |
14. |
39.03 |
0.941 ± 0.140 |
ND |
ND |
Quercetin |
15. |
47.55 |
4.924 ± 0.987 |
ND |
3.82 ± 0.615 |
Apigenin |
16. |
53.22 |
0.231 ± 0.009 |
ND |
ND |
Kaempferol |
17. |
66.55 |
0.810 ± 0.012 |
ND |
ND |
Chrysin |
Table 2B: Contents (micrograms per gram) of phenolic acids in EtOAc extracts fraction of A. marina leaf
Peak |
Retention time (min) |
F2 |
F2-5 |
F3 |
F3-2-9 |
F3-2-10 |
Comparison with standard |
|---|---|---|---|---|---|---|---|
1. |
3.52 |
ND |
ND |
ND |
ND |
ND |
Elliagic Acid (1) |
2. |
4.72 |
ND |
ND |
ND |
ND |
ND |
Eliiagic Acid (2), Gallic Acid |
3. |
5.55 |
ND |
ND |
ND |
ND |
ND |
Chlorogenic Aicd |
4. |
8.17 |
ND |
ND |
ND |
ND |
ND |
Catechin |
5. |
9.03 |
ND |
ND |
38.720 |
ND |
ND |
byring Acia, LUUU, Kutin, Caffeic Acid |
6. |
11.28 |
ND |
237.840 |
ND |
ND |
ND |
Corilagin |
7. |
14.35 |
ND |
6.845 |
ND |
0.234 |
0.367 |
Naringic, p-Coumaric Acid |
8. |
15.60 |
ND |
ND |
ND |
ND |
ND |
ECG |
9. |
26.78 |
ND |
ND |
686.320 |
3.890 |
2.361 |
Myricetin |
10. |
27.67 |
ND |
ND |
ND |
ND |
0.697 |
TF1 |
11. |
32.25 |
ND |
0.123 |
ND |
ND |
ND |
Resveratrol |
12. |
34.22 |
ND |
ND |
ND |
ND |
ND |
TF2 |
13. |
36.32 |
2.234 |
0.332 |
92.130 |
38.110 |
19.653 |
Luteolin |
14. |
39.03 |
ND |
ND |
ND |
98.321 |
88.652 |
Quercetin |
15. |
47.55 |
ND |
ND |
ND |
ND |
38.146 |
Apigenin |
16. |
53.22 |
ND |
1.328 |
ND |
0.653 |
0.563 |
Kaempferol |
17. |
66.55 |
ND |
1.398 |
110.690 |
22.095 |
132.780 |
Chrysin |
Identification and separation of EtOAc extracts of A. marina leaves
To identify bioactive components of A. marina leaves, EtOAc extracts were purified using column chromatography (Figure 4A). The plant material (20 kg) of A. marina was immersed in 200 L distilled water for 30 min and then boiled at 95°C for 30 min. The dried extract was partitioned between H2O and EtOAc to yield 35.69 g of a dried EtOAc extract and an aqueous residue. The EtOAc extract was subjected to silica gel medium pressure column chromatography (gradient mixtures, from 100% CH2Cl2 to 100% MeOH). The eluents were pooled to form 6 fractions on the basis of TLC analysis. Fraction 3 (8.29 g) was further purified on a silica gel column chromatography (hexane-EtOAc-Acetone, 10:3:3) to give fractions F3-1 to F3-6. Fraction F3-2 (224.7 mg) was subjected to Sephadex LH-20 column chromatography (MeOH) to give fractions F3-2-1 to F3-2-12.
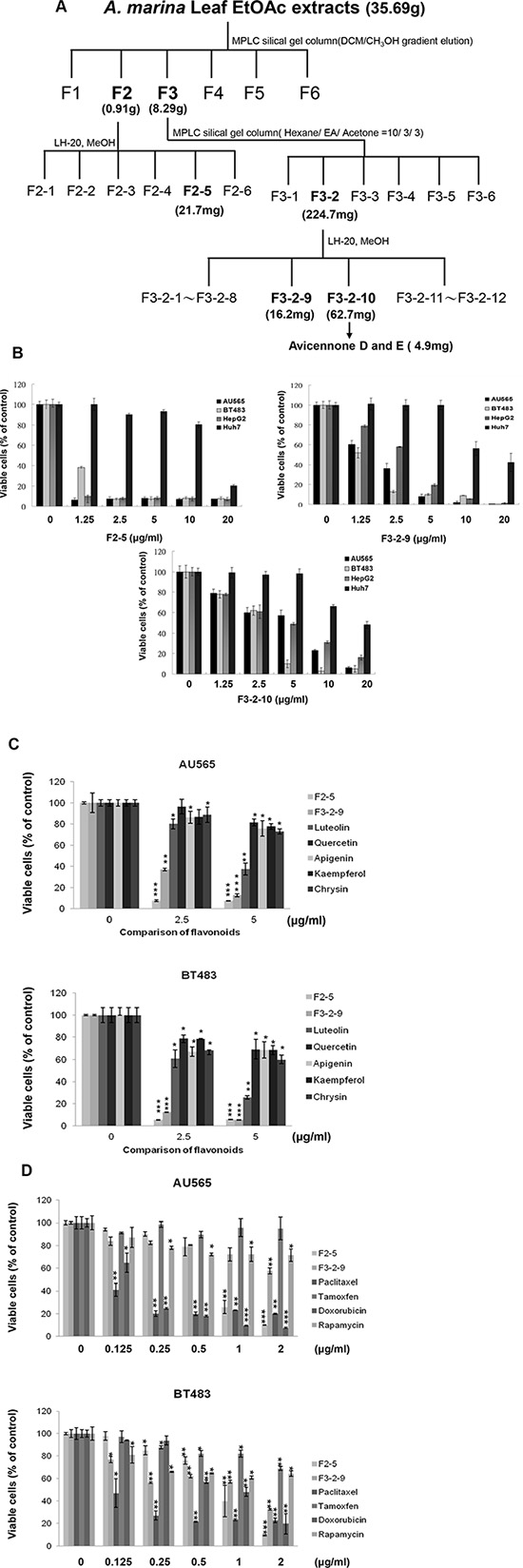
Figure 4: (A) Procedure for fractionation of EtOAc extracts from A. marina leaves; (B) Antiproliferative effects of F2-5, F3-2-9, and F3-2-10 in AU565, BT483, HepG2, and Huh7 cancer cells were determined using MTT assays. (C) Comparisons of antiproliferative effects of F2-5, F3-2-9, luteolin, quercetin, apigenin, kaempferol, and chrysin in AU565 and BT483 breast cancer cells; (D) Comparisons of antiproliferative effects of F2-5, F3-2-9, paclitaxel, tamoxifen, tdoxorubicin, and rapamycin on AU565 and BT483 breast cancer cells. All values are expressed as means ± standard errors of the mean (SE) and differences were identified using t-tests; *p < 0.05, **p < 0.01, ***p < 0.001.
Results showed that the fractions 2-5 (F2-5), 3-2-9 (F3-2-9), and 3-2-10 (F3-2-10) from EtOAc extracts of A. marina leaves had the highest anticancer effects (Figure 4B). Specifically, percentage inhibition was observed in the order F2-5 > F3-2-9 > F3-2-10, and F2-5 significantly inhibited cell proliferation in AU565, BT483, HepG2, and Huh7 cells in a dose-dependent manner, with half inhibitory concentrations (IC50) of 0.75, 0.85, 0.79, and 15.6 μg/mL, respectively. Similar results were observed following treatments with F3-2-9 and F3-2-10, with IC50 values of 2.1, 1.2, 3.2, and 9.8 μg/mL, and 5.6, 3.8, 5.1, and 19.2 μg/mL, respectively. In further HPLC analyses of F2-5, F3-2-9, and F3-2-10 (Figure 3B and Table 2B), F3-2-9 contained two major HPLC peaks with retention times of 36–40 min that corresponded with luteolin and quercetin standards. The major HPLC peak from F2-5 had a retention time of 55 minutes, and did not match any of the 21 standards. Subsequent NMR analyses showed that the most abundant components were the same in F3-2-9 and F3-2-10, but differed from those in F2-5 (data not shown).
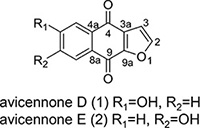
The bioactive fraction F3-2-10 further purified by HPLC to yield 1 and 2 (4.9 mg). The chemical structures of compounds 1 and 2 were identified by NMR spectroscopy. Compounds 1 and 2, which were chromatographically inseparable could be obtained only as a mixture (1:1, according to 1H & 13C NMR data). The identical molecular formula of 1 and 2 was established as C12H6O4 by ESIMS at m/z 213.0 [M-H]- and NMR data. A naphthoquinone skeleton was suggested by the absorption bands of UV spectrum at λmax 267, 307, and 350 nm. One set of 1H NMR signals of compound 1 indicated three aromatic protons (δH 8.01, 7.47, 7.10) in an AMX spin system and two AB-type olefinic protons (δH 7.96, d, J = 1.9 Hz, δH 6.99, d, J = 1.9 Hz) of a furan ring (Table 3). In the 13C NMR spectrum, two sets of signals, each representing 12 carbons, could be distinguished. The NMR data of the mixture of compounds 1 and 2 indicated them to carry a hydroxyl substituent on the aromatic ring. Based on above data and with those reported in the literature, 1 was determined to be 6-hydroxynaphtho [2, 3-b] furan-4, 9-dione and 2 as 7-hydroxynaphtho [2, 3-b] furan-4, 9-dione, and these compounds were named avicennone D and avicennone E, respectively. In contrast with previously reported 1H-NMR and 13C-NMR data, the F3-2-10 fraction was identified as a mixture of avicennone D and E (Table 3). To our knowledge, this is the first study to demonstrate apoptotic mechanisms of avicennone D and E from a HPLC fraction from an EtAOc extract of A. marina leaves in breast and liver cancer cells.
Table 3: 1H NMR and 13C NMR data of avicennone D (1) and avicennone E (2) (CD3OD)a
atom number |
1 |
2 |
||
|---|---|---|---|---|
δC (mult.) |
δH (mult. J in Hz) |
δC (mult.) |
δH (mult. J in Hz) |
|
2 |
150.29 (d) |
7.96 (d, 1.9) |
150.82 (d) |
7.98 (d, 1.9) |
3 |
109.17 (d) |
6.99 (d, 1.9) |
109.40 (d) |
6.99 (d, 1.9) |
3a |
131.32 (s) |
|
132.07 (d) |
|
4 |
181.93 (s) |
|
181.24 (t) |
|
4a |
137.12 (s) |
|
126.56 (s) |
|
5 |
114.44 (d) |
7.47 (d, 2.6) |
130.68 (d) |
8.03 (d, 8.5) |
6 |
164.73 (s) |
|
121.10 (d) |
7.12 (dd, 8.5, 2.6) |
7 |
121.2 (d) |
7.10 (dd, 8.5, 2.6) |
164.78 (s) |
|
8 |
130.55 (d) |
8.01 (d, 8.5) |
114.14 (d) |
7.49 (d, 2.6) |
8a |
125.73 (s) |
|
136.29 (s) |
|
9 |
174.42 (s) |
|
174.81 (s) |
|
9a |
154.56 (s) |
|
154.06 (s) |
|
aReference to residual solvent CD3OD signals at δH 3.3 and δC 49.0 and measured at 25oC, 600 MHz for 1H and 150 MHz for 13C. 13C multiplicities were assigned from DEPT experiments.
Major HPLC peaks from F2-5, F3-2-9, and F3-2-10 showed retention times of 36–66 minutes, and corresponded with luteolin, quercetin apigenin, kaempferol, and chrysin standards (Figure 3B and Table 2B). Thus, the anticancer properties of these polyphenols were compared with those of F2-5 and F3-2-9 in AU565 and BT483 cells. In these experiments, anticancer activities were ranked in the same decreasing order of F2-5 > F3-2-9 > luteolin > quercetin, apigenin, kaempferol, and chrysin in both cell lines (Figure 4C). In comparisons with paclitaxel, tamoxifen, doxorubicin, and rapamycin, antiproliferative activities decreased in the order paclitaxel > doxorubicin > F2-5 > F3-2-9 or rapamycin > tamoxifen in both cell lines. Similar results were also observed in BT483 cells (Figure 4D). Taken together, these data indicate the efficacy of F2-5 and F3-2-9 as treatments for breast cancer.
Fractions of EtOAc extracts from A. marina leaves induced apoptosis in breast and liver cancer cell lines
To identify antiproliferative mechanisms of EtOAc extracts and their fractions, cell cycle analyses were performed in breast and liver cells after 48-h treatments using flow cytometry. Treatment of AU565, BT483, and HepG2 cells with 80–200-μg/mL EtOAc extracts for 48 h significantly increased percentages of cells in the sub-G1 phase (44.56%–82.22%; Figure 5A). However, Huh7 cells treated with EtOAc extracts for 48 h were arrested in the G1 phase at lower doses (40–80 μg/mL), and apoptosis was observed after treatments with 120–200-μg/mL EtOAc extract, and after treatments with 1-μg/mL F2-5, 5-μg/mL F3-2-9, or 5-μg/mL F3-2-10 (Figure 5B). In further experiments, apoptosis was assessed in AU565, BT483, and HepG2 cells after treatments with EtOAc extracts (40–200 μg/mL) using trypan blue exclusion, Hoechst, and DNA fragmentation assays. In these experiments, significant time- and dose-dependent cell death was observed in trypan blue exclusion assays (Figure 6A) and corresponded with dose-dependent DNA fragmentation (Figure 6B). Fluorescence photomicrographs of cells stained with Hoechst 33258 confirmed the induction of apoptosis following treatment with EtOAc extracts at 40–80 μg/mL for 48 h. Specifically, control cells showed round and homogeneous nuclei, whereas cells treated with 80-μg/mL EtOAc extracts showed condensed and fragmented nuclei (Figure 6C). Similar observations were made in cells treated with 1-μg/mL F2-5, 5-μg/mL F3-2-9, or 5-μg/mL F3-2-10.
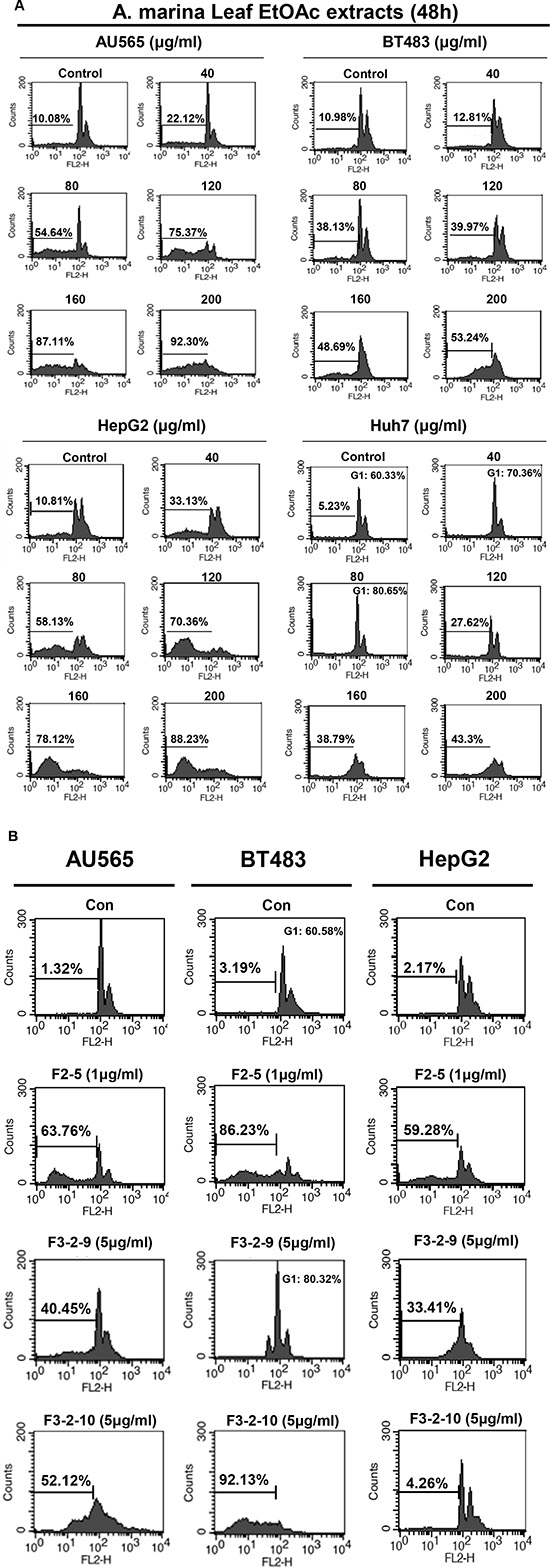
Figure 5: Effects of (A) EtOAc extracts of A. marina leaves and (B) fractions F2-5, F3-2-9, and F3-2-10 on cell cycle distributions of AU565, BT483, HepG2, and Huh7 cancer cells. Cell cycle distributions were assessed using flow cytometry.
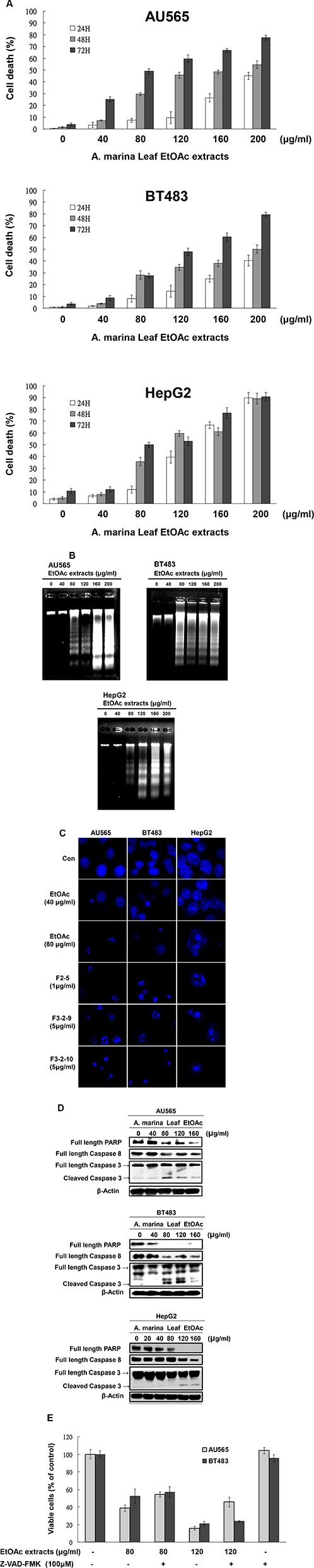
Figure 6: EtOAc extracts of A. marina leaves induced apoptosis in AU565, BT483, and HepG2 cells. Cytotoxic effects of EtOAc extracts of A. marina leaves in AU565, BT483, and HepG2 cancer cells were determined using trypan blue dye exclusion assays (A), DNA fragmentation assays (B), and Hoechst 33258 Dye assays (C) as described in the Materials and Methods. (D) Effects of EtOAc extracts of A. marina leaves on full-length PARP, full-length caspase 8, and cleaved caspase 3 production in AU565, BT483, and HepG2 cancer cells; Cells were treated with various concentrations of EtOAc extracts for 48 h in 2% FBS media. Total protein was collected and western blotting analyses were performed as described in the Materials and Methods. (E) Effects of EtOAc extracts of A. marina leaves on AU565 and BT483 cancer cell viability with or without pretreatment with Z-VAD-FMK caspase inhibitor. Cell viability was determined using MTT assays.
Flow cytometry and DNA fragmentation data indicated that treatments with EtOAc extracts trigger apoptosis in AU565, BT483, and HepG2 cells. Accordingly, western blot analyses showed decreased protein expression of full-length PARP, full-length caspase 8, and increased expression of cleaved caspase 3 after treatment with 80-μg/mL EtOAc extracts for 48 h (Figure 6D). In further investigations of caspase-dependent mechanisms (Figure 6E), pretreatment of AU565 cells with the caspase inhibitor Z-VAD-FMK blocked EtOAc extract-mediated apoptosis, but did not block apoptosis in BT483 cells. These results suggest roles of both caspase-dependent and -independent mechanisms in BT483 cells.
Anti-motility activities of EtOAc extracts of A. marina leaves in MDA-MB-231 cells
The effects of EtOAc extracts of A. marina leaves on cell motility were examined using wound-healing assays. In these experiments, significant cytotoxic activity was observed in MDA-MB-231 cells after treatment with 80-μg/mL EtOAc extracts for 48 h. To decrease interference in assessments of motility, wound-healing assays were performed after 24–48 h exposure to 10–40 μg/mL EtOAc extracts (Figure 7A). These assays showed inhibition of motility but not growth in MDA-MB-231 cells. Because cell motility has been associated with expression of specific cell adhesion proteins such as MMP2, MMP9, and EMT-related proteins, expression levels of these and fibronectin, snail protein, slug protein, vimentin, LRP6, and cyclin B were determined in MDA-MB-231 cells. In these experiments, treatments with 10–20-μg/mL EtOAc extracts decreased MMP2, MMP9, cyclin B, vimentin, and snail protein expression in MDA-MB-231 cells after 24 h (Figure 7B).
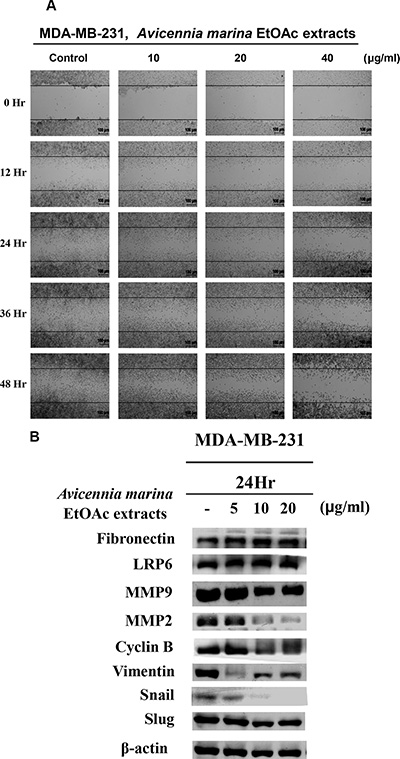
Figure 7: (A) Effects of 0–48-h treatments with EtOAc extracts of A. marina leaves on migration of MDA-MB-231 cells were examined using wound-healing assays. (B) LRP6, MMP9, MMP2, cyclin B, vimentin, snail protein, slug protein expression was determined using western blotting after 24-h exposures to various concentrations of EtOAc extracts of A. marina leaves as indicated.
EtOAc and EtOH extracts, and F2-5 from A. marina leaves inhibited growth of MDAMB-231 xenograft tumors in nude mice
To determine whether A. marina extracts inhibit breast cancer cell growth in vivo, human MDAMB-231 breast cancer cells were xeno-transplanted into the back skin of BALB/c nude mice and the effects of EtOAc extracts and EtOH extracts of A. marina leaves were investigated (Figure 8). In these animals, mean tumor volumes and weights were decreased by treatments with EtOAc and EtOH extracts at 200 mg/kg/day compared with DMSO-treated controls on day 23. However, no significant changes in relative weights of hearts, livers, spleens, or kidneys were observed in comparisons of EtOAc and EtOH extract-treated animals and the DMSO-treated control group. Finally, no differences in serum aspartate aminotransferase (AST) and creatinine values were found between treatment and control groups (data not shown), indicating limited side effects of A. marina extracts.
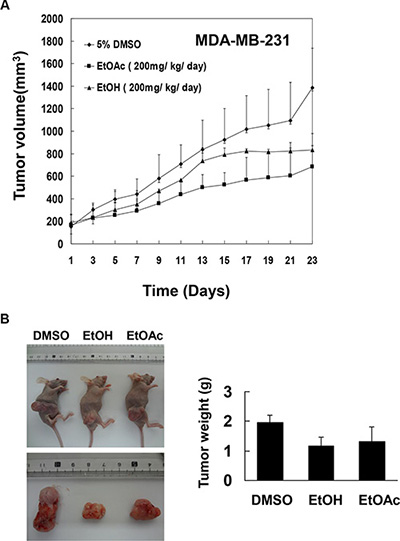
Figure 8: Effects of EtOAc and EtOH extracts of A. marina leaves on tumor growth in nude mice carrying MDAMB-231 xenografts. Cells were subcutaneously implanted into the legs of female nude mice to induce tumor xenografts. Treatment groups were intraperitoneally injected with dimethyl sulfoxide (DMSO), EtOAc extracts (200 mg/kg/day), or EtOH extracts (200 mg/kg/day) for 23 days, and tumor volumes (A) and tumor weights (B) were determined.
DISCUSSION
Mangroves grow under environmental conditions of high visible and UV radiation [17]. Both visible and UV radiation have been shown to be harmful to the photosynthetic apparatus in plants. Accordingly, higher plants adapt to high irradiance by accumulating UV-B-absorbing phenolic compounds such as flavonoids. Such flavonoids and phenolic acids also have antioxidant properties and health benefits in mammals, and flavonoids are the most abundant polyphenols in plant foods. Previous studies show that higher intakes of flavonoid-rich foods are associated with lower risks of various diseases, including cardiovascular disease and cancer [18]. A. marina is the most abundant and dominant mangrove species and has a long history of use as a traditional medicine for the treatment for skin diseases, rheumatism, ulcers, and smallpox. However, few previous studies have characterized the relationship between anticancer activities and polyphenol contents. The present study shows anticancer activities of four different extracts of A. marina in three human breast cancer cell lines, two liver human cancer cell lines, and in one normal cell line, and in a mouse xenograft model. Taken together, these data indicate chemotherapeutic potential of A. marina extracts and fractions, and the ensuing anticancer efficacies were correlated with total phenol contents. The present findings also show that treatment with A. marina leaf extracts and fractions leads to activation of PARP, caspase 3, and caspase 8 in AU565, BT483, and HepG2 cancer cell lines. EtOAc and EtOH extracts inhibited MDA-MB-231 xenograft tumor growth in nude mice. Accordingly, treatment with low-dose (10–20 μg/mL) EtOAc extracts for 24 h decreased cell migration by downregulating protein expression of MMP2, MMP9, cyclin B, vimentin, and snail protein in MDA-MB-231 cells, indicating decreased breast cancer cell growth both in vivo and in vitro.
Mangroves produce a wide variety of secondary metabolites with demonstrated anticancer properties, including alkaloids, triterpenoids, flavonoids, and tannins. Here EtOAc extracts of A. marina leaves had the highest phenol and flavonoid contents, and also had the best anticancer activities. The present F2-5, F3-2-9, and F3-2-10 fractions showed higher cytotoxic effects than the raw EtOAc extracts. In particular, F2-5 and F3-2-9 fractions exhibited strong inhibitory effects on AU565 cells, with IC50 values of 0.75 μg/mL and 2.1 μg/mL, respectively.
Because the criteria of the American National Cancer Institute consider IC50 values of < 30 μg/mL indicative of a promising extract [19], F2-5 and F3-2-9 can be considered as potential sources of anticancer drugs. However, whether EtOAc extracts of A. marina leaves contain other secondary metabolites, such as alkaloids, triterpenoids, and tannins, requires confirmation in further studies.
The present HPLC data showed that F3-2-9 contains two major peaks with retention times close to those of luteolin and quercetin standards. Luteolin is present in a fraction from a MeOH extract of A. marina that had high anticancer activity [20]. However, the present EtOAc extracts had higher cytotoxic effects than MeOH extracts. Hence, further structural comparisons of these fractions are warranted to elucidate anticancer mechanisms. Here reversed-phased HPLC of the F3-2-10 fraction yielded avicennones D and E, which were previously isolated from in 2007 by Li et al. [21]. However, corresponding anticancer activities of these compounds remain uncharacterized. Hence, the present data are the first to indicate apoptotic mechanisms of avicennone D and E in HPLC fractions from EtOAc extracts of A. marina leaves in breast and liver cancer cells.
In summary, EtOAc extracts of A. marina leaves induce apoptosis and inhibit migration of breast and liver cancer cells. These results warrant further investigation of A. marina leaf extracts to identify candidate drugs for the treatment of breast and liver cancers, either alone or in combination with other drugs.
MATERIALS AND METHODS
Preparation of A. marina extracts and ethyl acetate (EtOAc) fractions
Raw A. marina materials were collected from the Xinfeng mangrove conservation area in Taiwan. To prepare water (H2O) extracts, 100 g of dried A. marina leaves or seeds were immersed in 1000 mL of distilled water for 30 min and were then boiled at 95°C for 30 min. After cooling, the mixture was filtered through a filter paper and water extracts were obtained after evaporation to dryness in a vacuum rotary evaporator at 80°C. To prepare methanol (MeOH) and ethanol (EtOH) extracts, 100 g of A. marina dried leaves or seeds were immersed in 1000 mL of MeOH or EtOH for 24 h at 4°C and were then filtered using filter papers. Filtrates were then evaporated under vacuum at 55°C. To prepare ethyl acetate (EtOAc) extracts, water extracts were reconstituted in distilled water and were then partitioned sequentially with equal volumes of EtOAc as indicated in Figure 1A. EtOAc soluble fractions were then fractionated using various column chromatography protocols as indicated in Figure 4A. Tested concentrations were expressed according to dry weights of extracts as indicated in Table 1. Furthermore, the chemical structures were identified by NMR spectroscopy.
Cell viability assays
Cells were treated with A. marina extracts for 48 h and cell viability was determined using MTT [3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide] assays. Briefly, MTT-formazan crystals were formed in metabolically viable cells and were dissolved in 500 μl of DMSO for spectrophotometric determinations at a wavelength of 550 nm.
Determinations of total phenol and flavonoid contents
Total phenol contents were determined spectrophotometrically using gallic acid as a standard according to a previously described Folin-Ciocalteu method [22]. Total flavonoid contents were also determined spectrophotometrically using quercetin as a standard, according to a previously reported method [23].
Determination of trace metal elements in A. marina leaves using inductively coupled plasma atomic emission spectroscopy (ICP-AES)
ICP-AES was used for rapid, precise, and accurate determinations of trace metal elements in digested A. marina leaves. Briefly, 0.5-g leaf extracts were digested by heating in a sealed teflon microwave digestion vessel (MARS Xpress TFM high pressure reaction vessel) containing 10 mL of concentrated nitric acid. Teflon vessels were then incubated at 180°C for 10 min and after cooling to the room temperature, contents of vessels were thoroughly transferred into 50 mL polypropylene test tubes and were diluted to 10 mL in deionized water. Sample solutions were then filtered and concentrations of trace metal elements were measured directly using ICP-AES (PerkinElmer ICP Optima 21000bv).
High performance liquid chromatography (HPLC) analyses of A. marina extracts
Concentrations of polyphenol compounds in A. marina extracts were determined using HPLC with a 250 × 4.6 mm i.d., 5 μm Thermo 5 C18-MS packed column (Nacalei Tesque, Inc., Kyoto, Japan). Gradient elution was performed using mobile phases comprising (A) 15% acetonitrile, 4% ethyl acetate, 0.1% formic acid and 80.9% ddH2O, and (B) 45% acetonitrile, 4% ethyl acetate, 0.1% formic acid and 50.9% ddH2O. The flow rate was 1 mL/min and polyphenols were detected spectrophotometrically at 280 nm.
Soft agar colony formation assays
Equal volumes of single-cell suspensions were treated with EtOAc extracts of A. marina leaves in DMEM containing 20% fetal bovine serum (FBS), and were then mixed with 0.7% agarose. Mixtures were plated on 6-cm culture dishes on top of a base layer of 0.7% agarose containing 10% FBS, and were allowed to gel. After 14–21-days incubation, colonies of > 60 μm were counted using a light microscope.
Flow cytometry
Flow cytometric analyses were performed as previously described [24], and fluorescent propidium idodide (PI)-DNA complexes were quantified using FAC-Scan cytometry.
Trypan blue exclusion assays
Trypan blue dye exclusion assays were used to determine cytotoxic effects of A. marina leaf extracts in AU565, BT483, and HepG2 cells. Briefly, cells were seeded in six-well plates at a density of 5 × 104 cells in DMEM containing 10% FBS, and were then incubated for 24 h at 37°C. Culture medium was replaced and cells were incubated with or without the indicated concentrations of A. marina leaf extracts for 24–72 h in DMEM containing 2% FBS. Cells were then harvested by trypsinization and were mixed with trypan blue solution, and dead stained cells were counted using a hemocytometer under a microscope.
DNA fragmentation assays
DNA fragmentation assays were performed as previously described [19]. Briefly, cells were incubated with reagents at various concentrations for 48 h and were then lysed using lysis buffer. DNA was then extracted using phenol/chloroform/isoamyl alcohol and fragments were visualized using 2% agarose gel electrophoresis.
Analyses of nuclear morphology
Cells were plated on coverslips in six-well plates. After treatment with A. marina leaf extracts for 48 h, cells were fixed with 4% formaldehyde for 20 min and were then incubated in 4-mg/mL Hoechst 33258 for 30 min. Coverslips were washed and mounted in Vectashield (Vector Laboratories, Burlingame, CA) and nuclear chromatin morphology was viewed under a microscope.
Western blot analyses
Western blot analyses were performed as previously described [19], and immunoblots were visualized using enhanced chemiluminescent reagent.
Animals and treatments
Female BALB/c nude mice (6–8 weeks old) were purchased from the National Laboratory Animal Breeding and Research Center (Taipei, Taiwan). A total of 2 × 106 MDA-MB-231 cells were subcutaneously implanted into the legs of female nude mice. Mice were then randomly divided into three groups (four mice per group) for intraperitoneal treatments with DMSO, EtOH (200 mg/kg/day), and EtOAc extracts (200 mg/kg/day) for 23 days. Tumor volumes were calculated using the following formula: Volume (mm3) = A × B2/2, where A is the longest diameter (mm) and B is the shortest diameter (mm).
Statistical analyses
All data are expressed as means ± standard errors of the mean (SE) of at least three separate experiments for each group. Differences were identified using Student’s t-tests and were considered significant when *p < 0.05, **p < 0.01, or ***p < 0.001.
ACKNOWLEDGMENTS AND FUNDING
This work was supported by grants from the National Science Council, Taiwan. (NSC 100-2313-B-134-001-MY3, MOST 103-2313-B-134-001-MY3, MOST 104-2320-B-077-003 - and MOHW104-NRICM-D-315-133106).
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Willems L, Suarez F, Messas E, Baubion N, Decaudin D, Fourquet A, Ghez D, Delarue R, Hermine O, Buzyn A, Varet B, Rubio MT. [High risk of cardiac dysfunction after treatment of secondary acute myeloid leukemia following chemotherapy and radiotherapy for breast cancer]. Bull Cancer. 2010; 97:245–254.
2. Perez-Herrero E, Fernandez-Medarde A. Advanced targeted therapies in cancer: Drug nanocarriers, the future of chemotherapy. Eur J Pharm Biopharm. 2015; 93:52–79.
3. Van Uden DJ, van Laarhoven HW, Westenberg AH, de Wilt JH, Blanken-Peeters CF. Inflammatory breast cancer: an overview. Crit Rev Oncol Hematol. 2015; 93:116–126.
4. Monneur A, Bertucci F, Viens P, Goncalves A. [Systemic treatments of inflammatory breast cancer: an overview]. Bull Cancer. 2014; 101:1080–1088.
5. Wachtel-Galor S, Benzie IFF. Herbal Medicine: An Introduction to Its History, Usage, Regulation, Current Trends, and Research Needs. In: Herbal Medicine: Biomolecular and Clinical Aspects. Edited by Benzie IFF, Wachtel-Galor S, 2nd edn. Boca Raton (FL). 2011.
6. Vidyasagar MS, Ramanujam AS, Fernandes DJ, Koteshwar Rao K, Jadhav GK, Hospet CS, Seetharamaiah T, Vidyasagar S, Subramanyam K. Vincristine (Vinca-alkaloid) as a sclerosing agent for malignant pleural effusions. Acta Oncol. 1999; 38:1017–1020.
7. Rajput S, Mandal M. Antitumor promoting potential of selected phytochemicals derived from spices: a review. Eur J Cancer Prev. 2012; 21:205–215.
8. Eichhorst ST, Muerkoster S, Weigand MA, Krammer PH. The chemotherapeutic drug 5-fluorouracil induces apoptosis in mouse thymocytes in vivo via activation of the CD95(APO-1/Fas) system. Cancer Res. 2001; 61:243–248.
9. Karunagaran D, Rashmi R, Kumar TR. Induction of apoptosis by curcumin and its implications for cancer therapy. Curr Cancer Drug Targets. 2005; 5:117–129.
10. Desoize B, Sen S. [Apoptosis or programmed cell death: concepts, mechanisms and contribution in oncology]. Bull Cancer. 1992; 79:413–425.
11. Reed JC. Regulation of apoptosis by bcl-2 family proteins and its role in cancer and chemoresistance. Curr Opin Oncol. 1995; 7:541–546.
12. Sun SY, Hail N Jr, Lotan R. Apoptosis as a novel target for cancer chemoprevention. J Natl Cancer Inst. 2004; 96:662–672.
13. Xu DB, Ye WW, Han Y, Deng ZX, Hong K. Natural products from mangrove actinomycetes. Mar Drugs. 2014; 12:2590–2613.
14. Fardin KM, Young MC. Antifungal potential of Avicennia schaueriana Stapf & Leech. (Acanthaceae) against Cladosporium and Colletotrichum species. Lett Appl Microbiol. 2015; 61:50–7.
15. Bhimba BV, Agnel Defora Franco DA, Mathew JM, Jose GM, Joel EL, Thangaraj M. Anticancer and antimicrobial activity of mangrove derived fungi Hypocrea lixii VB1. Chin J Nat Med. 2012; 10:77–80.
16. Sharaf M, El-Ansari MA, Saleh NA. New flavonoids from Avicennia marina. Fitoterapia. 2000; 71:274–277.
17. Ragionieri L, Fratini S, Vannini M, Schubart CD. Phylogenetic and morphometric differentiation reveal geographic radiation and pseudo-cryptic speciation in a mangrove crab from the Indo-West Pacific. Mol Phylogenet Evol. 2009; 52:825–834.
18. Williamson G, Carughi A. Polyphenol content and health benefits of raisins. Nutr Res. 2010; 30:511–519.
19. Sudha S, Masilamani SM. Characterization of cytotoxic compound from marine sediment derived actinomycete Streptomyces avidinii strain SU4. Asian Pac J Trop Biomed. 2012; 2:770–773.
20. Momtazi-Borojeni AA, Behbahani M, Sadeghi-Aliabadi H. Antiproliferative activity and apoptosis induction of crude extract and fractions of avicennia marina. Iran J Basic Med Sci. 2013; 16:1203–1208.
21. Han L, Huang X, Dahse HM, Moellmann U, Fu H, Grabley S, Sattler I, Lin W. Unusual naphthoquinone derivatives from the twigs of Avicennia marina. J Nat Prod. 2007; 70:923–927.
22. Ainsworth EA, Gillespie KM. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat Protoc. 2007; 2:875–877.
23. Ghasemzadeh A, Jaafar HZ, Rahmat A. Antioxidant activities, total phenolics and flavonoids content in two varieties of Malaysia young ginger (Zingiber officinale Roscoe). Molecules. 2010; 15:4324–4333.
24. Huang C, Lee SY, Lin CL, Tu TH, Chen LH, Chen YJ, Huang HC. Co-treatment with quercetin and 1, 2, 3, 4, 6-penta-O-galloyl-beta-D-glucose causes cell cycle arrest and apoptosis in human breast cancer MDA-MB-231 and AU565 cells. J Agric Food Chem. 2013; 61:6430–6445.