
INTRODUCTION
Hepatocellular carcinoma (HCC) is the fifth most common cancer worldwide and the third leading cause of cancer-related deaths in China [1, 2]. Patient survival relies on early diagnosis, which remains challenging due to the lack of sensitive and specific tumor markers [3, 4]. HCC develops typically in the setting of cirrhosis associated with hepatitis B virus (HBV) infection in China [1, 4, 5]. HCC carcinogenesis involves multiple genetic changes including loss of tumor suppressor genes and activation of oncogenes, while HBV infection caused liver inflammation may induce mutation accumulation in hepatocytes [6]. The precise molecular mechanism of HBV-associated HCC remains unresolved.
Telomerase synthesizes telomeric TTAGGG repeats essential for chromosome stability and sustained cellular proliferation [7]. In normal somatic cells, length of telomeres shortens at each cycle of cell division, directing cells towards senescence and apoptosis. Telomerase is silent in most normal differentiated cells, while it is activated in up to 90% of human malignancies including HCC, leading to infinite proliferation potential [8–10]. The cancer-specific telomerase activation is primarily determined by telomerase reverse transcriptase (TERT) activity, which may be re-activated by epigenetic regulation, TERT amplification or TERT promoter mutations [8, 11, 12].
TERT promoter mutations have been found in many human cancers, including melanomas [12, 13], bladder carcinomas [14, 15], and thyroid carcinomas [16–18]. The mutations mainly occur in two hotspot sites located -124 (C228T) and -146 bp (C250T) upstream from the ATG start site, both of which create binding motifs for Ets/TCF transcription factors [19]. Either TERT promoter mutation is reported to elevate gene transcriptional activity by 2-4 fold [12].
TERT promoter mutations have recently been identified in 59% of HCC tissues from European patients with various etiologies [20]. TERT promoter mutations were also found in premalignant lesions within fibrous tissues, and thus are thought to be a new biomarker predictive of transformation of premalignant lesions into HCC [21]. Previous studies for TERT promoter mutations in HCC have been performed in in individuals infected with HCV from western countries and in Japan [11, 22–25]. However, little is known in patients with HCC secondary to HBV infection in Han Chinese, a population with the highest mortality rates. In the present study, we have determined promoter mutation status, TERT protein expression, and their clinical-pathological implications in HBV-associated HCC in 276 Chinese patients with comprehensive clinical, viral, and pathological data.
RESULTS
TERT promoter hotspot mutations in HCC
By direct sequencing of the TERT promoter region, we detected TERT hot spot mutations in 85 (31%) tumor tissues from 276 HCC cases (Table 1, Supplementary Figure S1). C228T was detected in 84 HCC tumors (30%) and C250T was detected in 1 HCC tumor (0.4%) (Figure 1). The two mutations were mutually exclusive, consistent with published observations [19]. In contrast, we found no TERT mutation in liver tissues from 20 control patients with cirrhosis or hepatolithiasis without HCC. TERT promoter mutations were significantly higher in HCC than in non-HCC patients (p = 0.003, Fisher Exact test).
Table 1: Characteristics of the HCC patients according to TERT promoter mutation status
Characteristics |
HCC, Total (N=276) |
TERT promoter mutation status |
OR |
95%CI |
p valuea |
|
|---|---|---|---|---|---|---|
Mutated |
Non-Mutated |
|||||
Number of tumor |
85 (30.8) |
191 (69.2) |
||||
Gender |
||||||
Female |
41 |
10(24.4) |
31(75.6) |
1.00 |
Reference |
|
Male |
235 |
75(31.9) |
160(68.1) |
1.45 |
0.68-3.12 |
0.34 |
Age median |
276 |
57.8±9.6 |
56.1±11.8 |
0.21b |
||
< 60 yr. |
158 |
41(25.9) |
117(74.1) |
1.00 |
Reference |
|
≥60 yr. |
118 |
44(36.8) |
74(63.2) |
1.70 |
1.01-2.84 |
0.04* |
HBV marker (HBsAg) |
||||||
Negative |
60 |
17(28.3) |
43(71.7) |
1.00 |
Reference |
|
Positive |
215 |
68(31.6) |
147(68.4) |
1.17 |
0.62-2.20 |
0.63 |
HBV marker (HBcAb or HBe Ab +) |
||||||
Negative |
16 |
7 (43.8) |
9 (56.2) |
1.0 |
||
Positive |
259 |
78 (30.1) |
181 (69.9) |
0.55 |
0.20-1.54 |
0.27 |
Family history |
||||||
No |
238 |
81(34.0) |
157(66.0) |
1.00 |
Reference |
|
Yes |
26 |
3(11.5) |
23(88.5) |
0.25 |
0.07-0.87 |
0.02* |
Tumor size |
||||||
<3cm |
78 |
23(29.5) |
55(70.5) |
1.00 |
Reference |
|
3-5cm |
82 |
29(35.4) |
53(64.6) |
1.31 |
0.67-2.54 |
0.43 |
>5cm |
53 |
13(24.5) |
40(75.5) |
0.77 |
0.35-1.72 |
0.53 |
Cirrhosis |
||||||
No |
52 |
22(42.3) |
30(57.7) |
1.00 |
Reference |
|
Yes |
195 |
62(31.8) |
133(68.2) |
0.64 |
0.34-1.19 |
0.16 |
Tumor Embolus |
||||||
No |
109 |
32(29.4) |
77(70.6) |
1.00 |
Reference |
|
Yes |
41 |
13(31.7) |
28(68.3) |
1.11 |
0.51-2.43 |
0.78 |
Tumor Capsule |
||||||
No |
20 |
6(30) |
14(70) |
1.00 |
Reference |
|
Yes |
163 |
43(26.4) |
120(73.6) |
0.84 |
0.30-2.31 |
0.73 |
Clinical stages |
||||||
I |
40 |
12(30) |
28(70) |
1.00 |
Reference |
|
II |
169 |
56(33.1) |
113(66.9) |
1.16 |
0.55-2.44 |
0.70 |
III |
40 |
9(22.5) |
31(77.5) |
0.68 |
0.25-1.85 |
0.45 |
IV |
17 |
4(23.5) |
13(76.5) |
0.72 |
0.19-2.66 |
0.86 |
Tumor differentiation |
||||||
I |
35 |
9(25.7) |
26(74.3) |
1.00 |
Reference |
|
II |
159 |
51(32.1) |
108(67.9) |
1.36 |
0.60-3.12 |
0.46 |
III |
67 |
21(31.3) |
46(68.7) |
1.31 |
0.53-3.30 |
0.55 |
ap values were from χ2 test, except for b, which was from t test. *: p < 0.05.
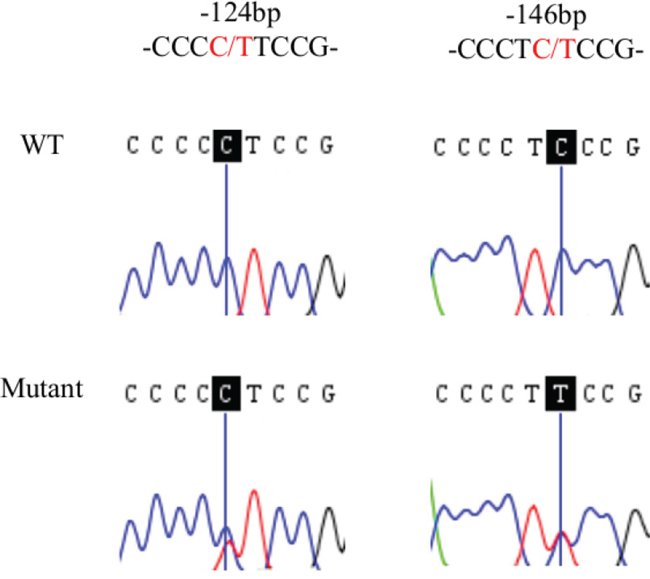
Figure 1: Sequence analysis of TERT promoter somatic mutations in HCC. Sanger sequencing of mutant or wildtype (WT) of the TERT promoter mutations at positions -124 and -146 bp (shadowed) from representative cases.
Clinical and pathological features of TERT promoter mutations in HCC Patients
Clinicopathological data, including demographic characteristics, laboratory results for HBV infection and liver function, and histopathology findings, were used for analysis. The relationship between TERT promoter mutation status (mutated/non-mutated) and clinicopathological features in HCC patients are presented in Table 1.
The presence of the TERT mutation was not associated with sex, HBV infection status, or the presence of HBV DNA, cirrhosis, tumor embolus, tumor capsule or tumor size (Table 1). TERT mutations were also not associated with HCC stages and tumor differentiation grades (Table 1) or with liver function tests (Supplementary Table S1). However, we found that HCC patients with a first-degree relative with HCC were less likely to carry TERT mutations (12%) compared to those without a family history (34%) (p = 0.02). HCC patients ≥60 yr were more likely to carry TERT mutations (37%) compared to those <60 yr (26%) (p = 0.04) (Table 1).
Correlation of TERT promoter mutations and α-fetoprotein (AFP) levels in HCC patients
TERT mutant carriers had lower serum AFP levels than the non-mutated group. The cutoff of AFP > 200 μg/l is standard in clinical practice in China to monitor HCC development in HBV infected patients [26, 27]. Mutation carriers (18.2%) were less likely to have AFP > 200 μg/l compared to the non-mutated group (18.2% versus 30.8%, respectively; p = 0.04, OR 0.5, 95% CI = 0.26-0.97, Table 2). Moreover, a sensitivity analysis of all HCC cases with abnormal AFP levels (using the conventional abnormal cutoff at AFP > 20μg/l) indicated TERT promoter mutations were significantly associated with lower AFP levels (p = 0.03; Figure 2A, Supplementary Figure S2, Table 2). Since HCC is more prevalent in males, we performed a stratified analysis for males and females; TERT promoter mutations were significantly associated with AFP levels and HBsAg positive males (p = 0.04), with adjustment for age.
Table 2: The AFP levels in HCC patients according to the TERT promoter mutation status
AFP Group |
Mutated (n = 77) |
Non-Mutated (n = 172) |
p-value |
OR |
95% CI |
|
|---|---|---|---|---|---|---|
① |
AFP ≤ 20 |
44 (57.1%) |
83 (48.3%) |
0.20a |
0.70 |
0.41-1.20 |
② |
20 < AFP ≤ 200 |
19 (24.7%) |
36 (20.9%) |
|||
③ |
AFP > 200 |
14 (18.2%) |
53 (30.8%) |
0.038b |
0.50 |
0.26-0.97 |
0.09c |
0.50 |
0.22-1.12 |
||||
AFP quantity mean±SD (> 20) |
870 ± 1638.7 |
1732.8 ± 2927.7 |
0.03* |
Note: a:①vs②+③; b:①+②vs③; c:②vs③, from χ2 test; * from t test.
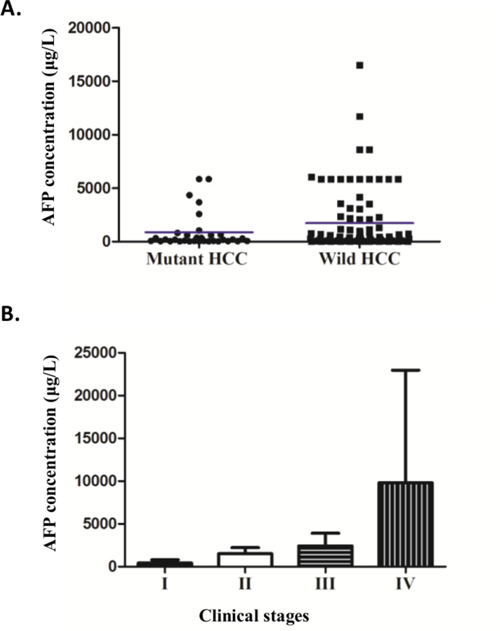
Figure 2: AFP levels in relationship with TERT promoter mutations and clinical stages. A. The scatter plot of the concentration of serum AFP (ug/L) in TERT mutant and wild-type groups (p = 0.045 excluded 2 outlier data points of more than 54000 in wild-type group; p = 0.03 including all data points as show in Supplementary Figure S2). Mean is shown as blue horizontal line. B. The relationship of AFP (ug/L) with clinical stages, ANOVA analysis (AFP>20, p=0.002).
We further observed a strong positive correlation of AFP levels and clinical stages (ANOVA AFP>20, p =0.002, Figure 2B), with AFP increasing with advancing stages. This result is consistent with previous findings in Chinese populations [28, 29]. Together, these data suggest that in those individuals with low AFP levels, detection of TERT promoter mutations may provide additional evidence for HCC diagnosis.
TERT protein expression measured by Immunohistochemistry (IHC)
We measured TERT protein expression by IHC in 28 HCCs (15 with mutation and 13 without mutation) and 8 non-HCC liver tissues including live cirrhosis tissues and normal liver tissues. In immunohistochemistry IHC, TERT staining was observed both in the nucleus and cytoplasm although nuclear staining was more intense (Figure 3). The intensity of TERT expression in the nucleus and cytoplasm did not differ by promoter mutation status. However, the intensity of TERT staining was higher in HCC tissues compared to non-tumor liver samples (p = 0.001).
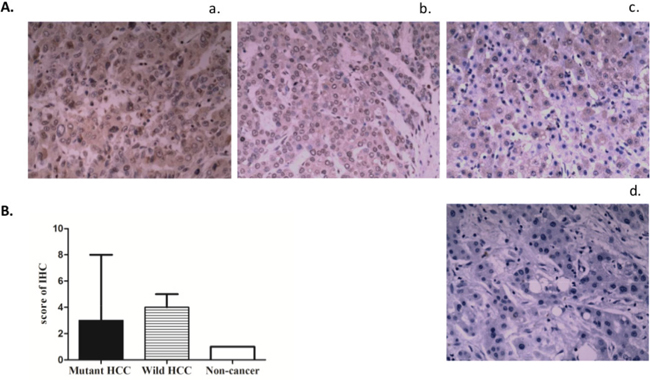
Figure 3: Expression of TERT protein in liver tissues stratified by TERT promoter mutation status. A. Immunostaining of TERT in representative cases. a: TERT promoter mutated live cancer; b: TERT promoter non-mutated live cancer; c: Non-cancer liver tissue (cirrhosis or hepatolith); d: negative control (without antibody to hTERT). Note the TERT staining in the nucleus. B. Bar graph of IHC scores representing TERT protein levels (Median with interquartile rage); mutant (n = 15) vs wild type (n = 13): p =0.94; HCC (n = 28) vs non-cancer (n= 8): p =0.0013 (U test).
TERT promoter hotspot mutations and TERT protein expression in the pre-neoplastic nodules
Dysplastic nodules may coexist with hepatocarcinoma tissues. We dissected preneoplastic lesions adjacent to the main HCC tissues from 7 cases (Supplementary Figure S3). In four of the 7 cases (57%), the C228T TERT mutation was found in both the paired preneoplastic lesions and the tumor tissue.
DISCUSSION
TERT promoter mutations are one of the most frequent somatic alterations in a variety of human cancers [19, 30]. In the present study, we screened for the presence of telomerase promoter mutations in liver tumor tissues from 276 HBV-associated HCC patients and investigated the clinical relevance of this gain-of-function somatic mutation. We found that 31% of HCC tissue harbored TERT promoter mutations and that the TERT promoter mutations were more often detected in those with low AFP level. HCC tissues also had higher TERT protein expression than either cirrhotic or normal liver tissues in a small sub-dataset. On the other hand, we found no correlation between TERT promoter mutation status and clinical stage, tumor differentiation grades or cirrhosis, consistent with other studies [21, 22, 25].
TERT mutations occurred at an early stage of tumorigenesis as they were observed in 30% of stage I HCCs. TERT promoter mutations were previously found in isolated cirrhotic premalignant nodules (6-19%) [20, 21]. Unexpectedly, TERT mutations were observed in our small subset of 4 out of 7 (57%) of preneoplastic lesions adjacent to HCC nodules with the TERT mutations. The mutant T allele had generally lower peaks in the sequence graph in the precursor compared to cancer tissues, possibly reflecting a mixture of tumor and non-tumor cells (Supplementary Figure S3). It may be indicative of in-situ tumorigenesis in preneoplastic lesions but is also possible that cancer cells invaded into the adjacent dysplastic lesions. This finding and its implication requires further investigation.
The frequency of TERT promoter mutational events varies markedly in different global geographical regions. In our cohort comprising Han Chinese patients with HBV-associated HCC, 31% carried TERT mutations. This rate is comparable with those of HBV infected patients from other studies in East Asians (20.7% - 38.8%) [20, 22, 25], but is much lower compared to other continental groups: Europeans (47-59%) [20, 23], Americans (44%) [11], and Africans (53%) [24]. These data indicate that TERT promoter mutations are less frequent among Asians, most of whom have HBV-associated HCC. HBV integration in the TERT locus inducing transcriptional activation may partially account for the lower rate of TERT promoter mutations in HBV-related HCC compared to HCV or alcohol- related HCC [25, 31].
Serum AFP is a key tumor marker of HCC. Current guidelines are to suspect HCC in cirrhosis patients whose AFP level ≥400 μg/L for at least one month or ≥200 μg/L for at least two months [26, 27]. Our study indicated that TERT promoter mutations are more frequent in tumors from patients with lower levels of AFP, in agreement with the Nault et al study [20]. In this study, 51% (127/249) of HCC patients had false-negative AFP (< 20 μg/L), among whom 34.7% (n = 44) carried TERT mutations. Detection of cancer-specific TERT mutations in patients with low serum AFP may thus complement the AFP assay which has insufficient sensitivity and specificity for early diagnosis of HCC [4, 26, 27].
Nuclear detection of TERT protein is a strong indication of telomerase activation and ongoing cell proliferation [32, 33]. We observed higher nuclear expression of TERT protein in HCC tumors compared to adjacent or control non-tumor liver tissues; however, we observed no differences in TERT expression between tumor tissues with or without the TERT promoter mutations, consistent with previous mRNA studies [20, 22]. High TERT protein expression or telomerase reactivation is likely a fundamental event in HCC.
In addition, we observed that TERT promoter mutations in HCC were associated with older patient age, consistent with other studies in HCC [20, 22, 25] and in in several other cancers [34–37]. The age effect may indicate a higher probability of TERT mutation with advancing age.
We noted for the first time that TERT promoter mutations are rarely seen in the patients with HCC family history. It is possible that heritable germline genetic variation and non-TERT somatic gene mutations drive hepatocarcinogenesis in this subset. This relationship warrants further attention in future studies.
The small sample size used in the expression study limits the power to detect subtle differences among TERT genotype groups and we did not test for HBV integration into the TERT locus. HBV integration into the TERT locus was found in 50% of Japanese HBV-associated HCCs without TERT mutations [25]; HBV integration may activate TERT in those without promoter mutations [31]. Future studies are needed to clarify the role of HBV integration on the observed associations. Finally, the invasive nature of liver biopsies for detection of TERT mutation and TERT protein expression limits its clinical utility. The development of “liquid biopsies” should be explored in future studies.
In conclusion, TERT promoter mutations in HCC are common somatic mutations in Han Chinese infected with HBV, and are negatively correlated with family history and AFP serum level. TERT promoter mutations are not associated with TERT protein levels or HCC stages and grades. Detection of TERT promoter mutations in those with low levels of AFP may aid in the differential diagnosis of HCC with atypical presentation. Additional studies are required to determine if TERT promoter mutated-HCCs represent a subset of patients amenable to anti-TERT therapies.
MATERIALS AND METHODS
Patients and tumor tissue specimens
The study was conducted according to the principles in the Declaration of Helsinki and the national laws and regulations. The Ethics Committee of Wenzhou Medical University approved this retrospective study using archived pathological specimens and the de-identified health information (IRB00009844). The de-identified data were analyzed anonymously. An IRB exemption was obtained from the National Institutes of Health Office of Human Subjects Research (OHSRP Review #12836).
The tissue specimens were formalin-fixed and paraffin-embedded (FFPE) blocks that were archived at the Department of Pathology, from a consecutive series of 276 patients who underwent tumor resection at the First Affiliated Hospital of Wenzhou Medical University, Wenzhou, Zhejiang, China, during January 2010 and January 2014. Only specimens with pathologic diagnosis of hepatocellular carcinoma and of sufficient tissue specimens were used. The adjacent non-tumor preneoplastic lesions were separated on hematoxylin-eosin (HE) stained slides by microscopic-guided dissection, with aid of laser capture microdissection (ArcturusXT, Applied Biosystems) [38].
Tumor grading and staging were performed according to criteria of World Health Organization [39] and the Tumor Lymph Node Metastasis (TNM) classification of the international Union Against Cancer [40], which is based on tumor number, size, vascular invasion, lymphatic involvement and metastasis.
Of 276 HCC patients sequenced, 86% were male (n = 235) and 94% were HBV infected (based on HBV serological markers, among which 78% was HBsAg positive). Patients had a median age of 58 years (mean±SD, 56.5±11.2) ranging from 23 to 88 years old. Of 20 non-HCC controls, 16 had decompensated cirrhosis and 4 had hepatolithiasis. The study work flow is presented in Supplementary Figure S1.
AFP level in serum was measured using the Beckman Coulter Access alpha-fetoprotein (AFP) immunoassay kit on a Beckman Coulter UniCel DxI 800 instrument. The assay is a 2-site immunoenzymatic sandwich assay for the quantitative determination of AFP with a measuring range 0.50-51,000 ng/ml.
DNA extraction
Genomic DNA from FFPE tissues was isolated from tissue sections with the phenol extract method [41]. Briefly, paraffin-embedded sample were subjected to xylene treatment to dissolves the paraffin, and then rehydrated. Digestion with Lysis buffer containing dodecyl sulfate (SDS) and proteins K was carried out at 56°C overnight, followed by phenol-chloroform extraction, isopropanol precipitation and re-suspension in H2O. Amount and quality of DNA were measured by a NanoDrop 2000 (Thermo Fisher Scientific).
Sequencing of the TERT promoter region
The promoter region of TERT covering the two hotspot mutations -124C>T and -146C>T was amplified by polymerase chain reaction (PCR) as previously described [19]. The 163bp fragment of the TERT promoter region spanning the two hotspot mutations (-124, -146 sites) was amplified by KOD FXDNA polymerase (KFX-101, Toyobo Life Science) and performed with the following primer pairs: 5'-CAGCGCTGCCTGAAACTC-3'(sense) and 5'-GTCCTGCCCCTTCACCTT-3'(anti-sense). PCR products were visualized in 2% agarose gels and verified to have the expected size of 163 bp, which contained the site of C288T and C250T mutations. The direct sequencing was done on the 3730XL DNA Analyzer with BigDye Terminator V 3.1 (Applied Biosystems). Sequencing was carried out by BGI-Shanghai and Sunny Biotech Co., Ltd. (Shanghai, China). The detected mutations were validated in both strands.
Immunohistochemistry (IHC) for TERT protein expression
FFPE slides of 3 μm sections were used to detect the TERT protein expression by IHC. Human TERT (hTERT) reactivity was detected by use of a specific mouse monoclonal [2C4] antibody to hTERT (ab5181, Abcam) with immunoperoxidase staining. Briefly, deparaffinized and rehydrated sections were exposed to 3% H2O2 solution for 10 min, followed by antigen retrieval in the citrate buffer. Incubation with anti-hTERT at a 1:100 dilution was carried out at 4°C overnight, followed by incubation with horseradish peroxidase-conjugated polymer anti-mouse secondary antibody for 20 mins at 37°C. 3,3′-Diaminobenzidine (DAB) (ZSGB-Bio) was applied for visualization. IHC score was evaluated by experienced pathologists. HCC slides without treatment of anti-hTERT were served as negative controls and tonsil slides were served as positive controls. The degree of positive staining for TERT was defined on the intensity of the staining and the percentage of stained cells [42]. In particular, the intensity was defined as negative (0), low (1), medium (2) and high (3), meanwhile the percentage of stained cells were defined as none (0), 1–10% (1), 10–50% (2) 50-80% (3) and >80% (4). We evaluated semi-quantitatively using the product of intensity and percentage of stained as following scale: negative (0-1); moderate (2-6); strong (8-12).
Statistical analysis
We performed statistical analyses using SPSS software version 17.0 (SPSS Inc.). Student’s t test was used to test the differences of continuous variables between groups with and without the TERT promoter mutations. Fisher’s exact or χ2 test were used to examine associations between categorical variables. The results are expressed as a percentage or mean ± SD. A nonparametric analysis of Mann-Whitney U-test was used to test the immunostaining raw scores between the two groups because the analytical IHC scores were not normally distributed. All tests were two-tailed and results were considered statistically significant at p<0.05.
ACKNOWLEDGMENTS
The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.
CONFLICTS OF INTEREST
The authors disclose no conflicts of interest.
GRANT SUPPORT
This project has been supported by the Opening Project of Zhejiang Provincial Top Key Discipline of Clinical Medicine. This project has been funded in whole or in part with federal funds from the National Cancer Institute, National Institutes of health, under contract HHSN26120080001E. This Research was supported in part by the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research.
REFERENCES
1. El-Serag HB, Rudolph KL. Hepatocellular carcinoma: epidemiology and molecular carcinogenesis. Gastroenterology. 2007; 132:2557-2576.
2. Chen W, Zheng R, Baade PD, Zhang S, Zeng H, Bray F, Jemal A, Yu XQ, He J. Cancer statistics in China, 2015. CA Cancer J Clin. 2016; 66:115-132.
3. Singal AG, Pillai A, Tiro J. Early detection, curative treatment, and survival rates for hepatocellular carcinoma surveillance in patients with cirrhosis: a meta-analysis. PLoS Med. 2014; 11:e1001624.
4. Forner A, Llovet JM, Bruix J. Hepatocellular carcinoma. Lancet. 2012; 379:1245-1255.
5. Zhu Q, Li N, Zeng X, Han Q, Li F, Yang C, Lv Y, Zhou Z, Liu Z. Hepatocellular carcinoma in a large medical center of China over a 10-year period: evolving therapeutic option and improving survival. Oncotarget. 2015; 6:4440-4450. doi: 10.18632/oncotarget.2913.
6. Arzumanyan A, Reis HM, Feitelson MA. Pathogenic mechanisms in HBV- and HCV-associated hepatocellular carcinoma. Nat Rev Cancer. 2013; 13:123-135.
7. Mitchell M, Gillis A, Futahashi M, Fujiwara H, Skordalakes E. Structural basis for telomerase catalytic subunit TERT binding to RNA template and telomeric DNA. Nature structural & molecular biology. 2010; 17:513-518.
8. Pinyol R, Tovar V, Llovet JM. TERT promoter mutations: Gatekeeper and driver of hepatocellular carcinoma. J Hepatol. 2014; 61:685-687.
9. Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PdL, Coviello GM, Wright WE, Weinrich SL, Shay JW. Specific association of human telomerase activity with immortal cells and cancer. Science. 1994; 266:2011-2015.
10. Kong F, Zheng C, Xu D. Telomerase as a “stemness” enzyme. Science China Life Sciences. 2014; 57:564-570.
11. Killela PJ, Reitman ZJ, Jiao Y, Bettegowda C, Agrawal N, Diaz LA, Jr., Friedman AH, Friedman H, Gallia GL, Giovanella BC, Grollman AP, He TC, He Y, Hruban RH, Jallo GI, Mandahl N, et al. TERT promoter mutations occur frequently in gliomas and a subset of tumors derived from cells with low rates of self-renewal. Proc Natl Acad Sci U S A. 2013; 110:6021-6026.
12. Huang FW, Hodis E, Xu MJ, Kryukov GV, Chin L, Garraway LA. Highly recurrent TERT promoter mutations in human melanoma. Science. 2013; 339:957-959.
13. Horn S, Figure lA, Rachakonda PS, Fischer C, Sucker A, Gast A, Kadel S, Moll I, Nagore E, Hemminki K, Schadendorf D, Kumar R. TERT promoter mutations in familial and sporadic melanoma. Science. 2013; 339:959-961.
14. Li C, Wu S, Wang H, Bi X, Yang Z, Du Y, He L, Cai Z, Wang J, Fan Z. The C228T mutation of TERT promoter frequently occurs in bladder cancer stem cells and contributes to tumorigenesis of bladder cancer. Oncotarget. 2015; 6:19542-19551. doi: 10.18632/oncotarget.4295.
15. Wang K, Liu T, Liu L, Liu J, Liu C, Wang C, Ge N, Ren H, Yan K, Hu S, Bjorkholm M, Fan Y, Xu D. TERT promoter mutations in renal cell carcinomas and upper tract urothelial carcinomas. Oncotarget. 2014; 5:1829-1836. doi: 10.18632/oncotarget.1829.
16. Xing M, Liu R, Liu X, Murugan AK, Zhu G, Zeiger MA, Pai S, Bishop J. BRAF V600E and TERT promoter mutations cooperatively identify the most aggressive papillary thyroid cancer with highest recurrence. J Clin Oncol. 2014; 32:2718-2726.
17. Wang N, Liu T, Sofiadis A, Juhlin CC, Zedenius J, Hoog A, Larsson C, Xu D. TERT promoter mutation as an early genetic event activating telomerase in follicular thyroid adenoma (FTA) and atypical FTA. Cancer. 2014; 120:2965-2979.
18. Liu X, Qu S, Liu R, Sheng C, Shi X, Zhu G, Murugan AK, Guan H, Yu H, Wang Y, Sun H, Shan Z, Teng W, Xing M. TERT Promoter Mutations and Their Association with BRAF V600E Mutation and Aggressive Clinicopathological Characteristics of Thyroid Cancer. J Clin Endocrinol Metab. 2014; 99:E1130-1136.
19. Heidenreich B, Rachakonda PS, Hemminki K, Kumar R. TERT promoter mutations in cancer development. Curr Opin Genet Dev. 2014; 24:30-37.
20. Nault JC, Mallet M, Pilati C, Calderaro J, Bioulac-Sage P, Laurent C, Laurent A, Cherqui D, Balabaud C, Zucman-Rossi J. High frequency of telomerase reverse-transcriptase promoter somatic mutations in hepatocellular carcinoma and preneoplastic lesions. Nat Commun. 2013; 4:2218.
21. Nault JC, Calderaro J, Di Tommaso L, Balabaud C, Zafrani ES, Bioulac-Sage P, Roncalli M, Zucman-Rossi J. Telomerase reverse transcriptase promoter mutation is an early somatic genetic alteration in the transformation of premalignant nodules in hepatocellular carcinoma on cirrhosis. Hepatology. 2014; 60:1983-1992.
22. Chen YL, Jeng YM, Chang CN, Lee HJ, Hsu HC, Lai PL, Yuan RH. TERT promoter mutation in resectable hepatocellular carcinomas: a strong association with hepatitis C infection and absence of hepatitis B infection. Int J Surg. 2014; 12:659-665.
23. Quaas A, Oldopp T, Tharun L, Klingenfeld C, Krech T, Sauter G, Grob TJ. Frequency of TERT promoter mutations in primary tumors of the liver. Virchows Archiv. 2014; 465:673-677.
24. Cevik D, Yildiz G, Ozturk M. Common telomerase reverse transcriptase promoter mutations in hepatocellular carcinomas from different geographical locations. World J Gastroenterol. 2015; 21:311-317.
25. Kawai-Kitahata F, Asahina Y, Tanaka S, Kakinuma S, Murakawa M, Nitta S, Watanabe T, Otani S, Taniguchi M, Goto F, Nagata H, Kaneko S, Tasaka-Fujita M, Nishimura-Sakurai Y, Azuma S, Itsui Y, et al. Comprehensive analyses of mutations and hepatitis B virus integration in hepatocellular carcinoma with clinicopathological features. J Gastroenterol. 2015. PMID:26553052. [Epub ahead of print].
26. Sturgeon CM, Duffy MJ, Hofmann BR, Lamerz R, Fritsche HA, Gaarenstroom K, Bonfrer J, Ecke TH, Grossman HB, Hayes P, Hoffmann RT, Lerner SP, Lohe F, Louhimo J, Sawczuk I, Taketa K, et al. National Academy of Clinical Biochemistry Laboratory Medicine Practice Guidelines for use of tumor markers in liver, bladder, cervical, and gastric cancers. Clin Chem. 2010; 56:e1-48.
27. Qin S, Primary Liver Cancer D and Treatment Expert Panel of the Chinese Ministry of H. Guidelines on the diagnosis and treatment of primary liver cancer (2011 edition). Chin Clin Oncol. 2012; 1:10.
28. Peng SY, Chen WJ, Lai PL, Jeng YM, Sheu JC, Hsu HC. High alpha-fetoprotein level correlates with high stage, early recurrence and poor prognosis of hepatocellular carcinoma: significance of hepatitis virus infection, age, p53 and beta-catenin mutations. Int J Cancer. 2004; 112:44-50.
29. Yao M, Zhao J, Lu F. Alpha-fetoprotein still is a valuable diagnostic and prognosis predicting biomarker in hepatitis B virus infection-related hepatocellular carcinoma. Oncotarget. 2016; 7: 3702-3708. doi: 10.18632/oncotarget.6913.
30. Vinagre J, Pinto V, Celestino R, Reis M, Populo H, Boaventura P, Melo M, Catarino T, Lima J, Lopes JM, Maximo V, Sobrinho-Simoes M, Soares P. Telomerase promoter mutations in cancer: an emerging molecular biomarker? Virchows Arch. 2014; 465:119-133.
31. Sung WK, Zheng H, Li S, Chen R, Liu X, Li Y, Lee NP, Lee WH, Ariyaratne PN, Tennakoon C, Mulawadi FH, Wong KF, Liu AM, Poon RT, Fan ST, Chan KL, et al. Genome-wide survey of recurrent HBV integration in hepatocellular carcinoma. Nat Genet. 2012; 44:765-769.
32. Chen Y, Kong Q. Nuclear translocation of telomerase reverse transcriptase: a critical process in chemical induced hepatocellular carcinogenesis. Neoplasma. 2010; 57:222-227.
33. Zavlaris M, Angelopoulou K, Vlemmas I, Papaioannou N. Telomerase reverse transcriptase (TERT) expression in canine mammary tissues: a specific marker for malignancy? Anticancer Res. 2009; 29:319-325.
34. Huang HN, Chiang YC, Cheng WF, Chen CA, Lin MC, Kuo KT. Molecular alterations in endometrial and ovarian clear cell carcinomas: clinical impacts of telomerase reverse transcriptase promoter mutation. Mod Pathol. 2015; 28:303-311.
35. Vinagre J, Almeida A, Pópulo H, Batista R, Lyra J, Pinto V, Coelho R, Celestino R, Prazeres H, Lima L. Frequency of TERT promoter mutations in human cancers. Nature communications. 2013; 4.
36. Liu T, Wang N, Cao J, Sofiadis A, Dinets A, Zedenius J, Larsson C, Xu D. The age-and shorter telomere-dependent TERT promoter mutation in follicular thyroid cell-derived carcinomas. Oncogene. 2014; 33:4978-4984.
37. Simon M, Hosen I, Gousias K, Rachakonda S, Heidenreich B, Gessi M, Schramm J, Hemminki K, Waha A, Kumar R. TERT promoter mutations: a novel independent prognostic factor in primary glioblastomas. Neuro Oncol. 2015; 17:45-52.
38. Jin C, Chen J, Meng Q, Carreira V, Tam NN, Geh E, Karyala S, Ho SM, Zhou X, Medvedovic M, Xia Y. Deciphering gene expression program of MAP3K1 in mouse eyelid morphogenesis. Dev Biol. 2013; 374:96-107.
39. Kleihues P, Sobin LH. World Health Organization classification of tumors. Cancer. 2000; 88:2887-2887.
40. Sobin LH, Fleming ID. TNM classification of malignant tumors, fifth edition (1997). Cancer. 1997; 80:1803-1804.
41. Pikor LA, Enfield KS, Cameron H, Lam WL. DNA extraction from paraffin embedded material for genetic and epigenetic analyses. J Vis Exp. 2011;:e2763.
42. Brustmann H. Immunohistochemical detection of human telomerase reverse transcriptase (hTERT) and c-kit in serous ovarian carcinoma: a clinicopathologic study. Gynecologic oncology. 2005; 98:396-402.