
INTRODUCTION
Neuroblastoma (NB) is a highly aggressive pediatric malignancy and accounts for 13% of cancer morbidity in children with current cure rates less than 50% [1]. The MYCN oncogene is a primary driver of this malignancy and is also amplified in almost 50% of high-risk cases. Additional oncogenes including c-Src (cellular-Src) are known to increase NB invasiveness and proliferation in vitro [2, 3]. Here, we present in vitro, in vivo, and clinical data defining the major repressor of c-Src, PAG1 (Phosphoprotein Associated with Glycosphingolipid-enriched microdomains 1), as a novel tumor suppressor in high-risk NB.
PAG1 (Csk binding protein, Cbp) is a ubiquitous lipid-raft associated scaffold protein that inhibits Src Family Kinases (SFKs) including c-Src [4]. The activated transmembrane PAG1 binds and co-localizes both SFKs and Csk (C-terminal Src Kinase) to facilitate inhibition of SFKs by Csk. Thus PAG1 is essential for optimal Csk based repression of c-Src [5]. PAG1 can also inhibit c-Src independent of Csk by directly sequestering this kinase to lipid-rafts. PAG1 represses c-Src and prevents activation of multiple downstream effector proteins controlling cellular adhesion, proliferation, inflammation and differentiation [2, 6]. Importantly, negative feedback activation of PAG1 controls potent proliferative and oncogenic functions of c-Src in non-transformed cells [7, 8].
Elevated c-Src activity is found in multiple malignancies including NB that promotes proliferation, cell-cell adhesion, and anti-apoptotic functions [5, 9]. Small molecule tyrosine kinase inhibitors such as dasatanib and PP2 induce apoptosis and block proliferation of NB cells in culture [10, 11]. However, lack of specificity has made the dissection of the essential targets of these drugs difficult [12]. In addition, the endogenous mechanisms promoting aberrant c-Src activation in tumors remain poorly understood. Thus, our analysis of patient cohorts revealing a highly significant correlation of low PAG1 levels with poor survival, prompted us to further investigate the role of PAG1 in NB pathogenesis.
Herein, we demonstrate that PAG1 gain-of-function inhibits NB proliferation and colony formation in vitro, while xenografts with high PAG1 expression show limited tumorigenicity in vivo. These PAG1 effects correlate with inhibition of active c-Src (pY416) and downstream effector proteins AKT and ERK. Overall, our findings demonstrate potent tumor suppressor functions for PAG1 and suggest transcriptional repression of PAG1 is a major mechanism for c-Src hyperactivity in NB. These findings further suggest that reactivation of PAG1 may be a clinically significant novel therapeutic approach for NB.
RESULTS
PAG1 expression strongly correlates with overall survival of neuroblastoma patients
To evaluate how transcription of PAG1 correlates with NB outcomes, we analyzed multiple independent clinical cohorts of NB patients using the R2-data analysis platform. Kaplan-Meier analyses of datasets revealed that low PAG1 transcript levels strongly correlated with poor overall and event-free survival for the Versteeg cohort (n=88) (p<0.0001), for the Kocak dataset (n=476) (p<0.0001), and for the NCI-POB dataset (n=56) (p<0.0001) (Figure 1A, 1B, 1C). Other disease factors, such as relapse or tumor progression, and risk of death due to disease was also found to inversely correlate with PAG1 expression (p<0.001) (Figure 1D). In addition, higher stage, more aggressive tumors have significantly lower PAG1 expression suggesting that loss of PAG1 function may lead to de-differentiated invasive malignancy (Figure 1E). We observed a strong inverse correlation between PAG1 and MYCN expression levels (p<0.001) in all patient datasets analyzed (Supplementary Figure S1). MYCN non-amplified samples in clinical cohorts show worse overall survival with low PAG1 levels (p<0.001) while the MYCN amplified samples did not show significant correlation of PAG1 levels to overall survival (Supplementary Figure S2). This is due to aggressive disease progression in MYCN amplified tumors due in part to suppression of PAG1 (Supplementary Figure S1). These findings suggest that active suppression of PAG1 is an important factor influencing the biology and response to therapy of both MYCN amplified and non-amplified high-risk NB.
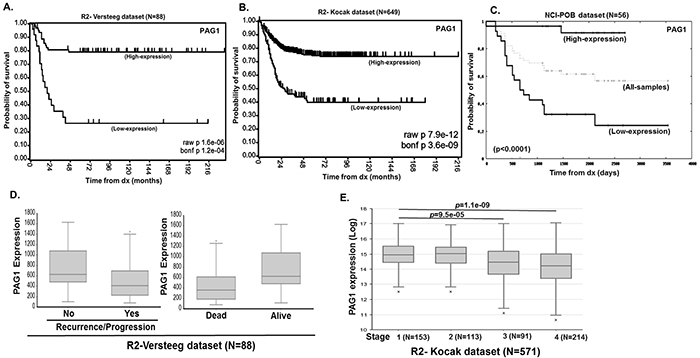
Figure 1: PAG1 is a prognostic factor in neuroblastoma. Kaplan-Meier curves shows the probability of overall survival based on PAG1 expression level A. 88 patients in the Versteeg dataset, B. 649 patients in the Kocak dataset and C. 56 patients in the NCI-POB dataset. D. R2-Versteeg dataset analysis showing correlation of PAG1 expression levels to neuroblastoma recurrence or progression and overall outcome. E. R2-Kocak dataset analysis showing correlation of PAG1 expression levels to neuroblastoma disease stages.
PAG1 inhibits oncogenic potential of neuroblastoma cell lines
To further investigate the effect of PAG1 in NB, we derived gain-of-function and loss-of-function NB cell lines from the MYCN amplified cell line NGP and non-amplified cell line SH-SY5Y. Validation of these cell lines for PAG1 mRNA expression by qPCR assay shows profound increase of PAG1 levels in PAG1 over-expression (PAG1-OEX) cell lines and significant decrease of PAG1 levels in PAG1 knockdown (PAG1-KD) cell lines (Figure 2A). In MTS proliferation assays, PAG1-OEX decreases proliferation in both NGP and SH-SY5Y lines compared to parental cell lines (p<0.001). Furthermore, PAG1-KD increases cell proliferation in NGP by 1.6 fold (p<0.01) and in SH-SY5Y by 2.6 fold (p<0.001) (Figure 2B). In soft-agar colony formation assays, PAG1-OEX markedly decreases anchorage-independent colony formation (p<0.0001) while PAG1-KD increases the colony formation in NGP by 1.2 fold (p=ns) and in SH-SY5Y by 3.4 fold (p<0.001) (Figure 2C).
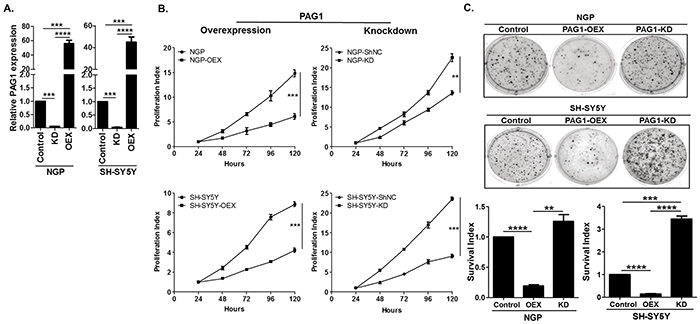
Figure 2: PAG1 is a tumor suppressor in neuroblastoma in vitro. A. Quantitative PCR (qPCR) analysis of PAG1 mRNA expression in PAG1 overexpression and knockdown cell lines in NGP and SH-SY5Y. Experiment was performed in triplicates and repeated thrice. Data represented as mean ± SD (T-test, ***p<0.001, ****p<0.0001). PAG1 mRNA expression in control cell lines NGP and SH-SY5Y was used to quantitate relative expression. B. Cellular proliferation assay of PAG1 gain and loss of function in NGP and SH-SY5Y NB cell lines. Cell proliferation was measured using MTS assay over 120 h. Proliferation index over the time was calculated by normalizing to the 24 h time point. Experiments were repeated three times with six replicates for each condition and represented as mean ± SD. (T-test, **p<0.01, ***p<0.001). C. Representative pictures of colony formation assay for PAG1 gain and loss of function in NB cell lines. Survival index shows the quantitation of relative inhibition of colony formation in different cell lines and is normalized to control or original cell line. Data represented as mean ± SD (T-test, **p<0.01, ***p<0.001, ****p<0.0001). OEX= PAG1-overexpression, KD= PAG1-knockdown, shNC= negative/mock control knockdown.
PAG1 reduces neuroblastoma tumorigenicity in vivo
To evaluate the effects of PAG1 loss and gain-of-function on NB tumorigenicity in vivo, we generated cohorts of xenografts in athymic immunodeficient nude mice via orthotopic injection into the renal capsule as previously described [18]. Results show that both NGP and SH-SY5Y PAG1-OEX cell line cohorts have significant (p<0.01) reduction in tumor weights in contrast to parental control cell line and PAG1-KD cell line xenograft cohorts (Figure 3A, 3B). We further verified the expression levels of PAG1 in harvested tumor tissues, and as expected, observed increased PAG1 expression in PAG1-OEX xenograft tumor tissues compared to parental control cell line tumors (Figure 3C). These in vivo studies further validate our in vitro observations that PAG1 is able to serve as a potent tumor suppressor in NB.
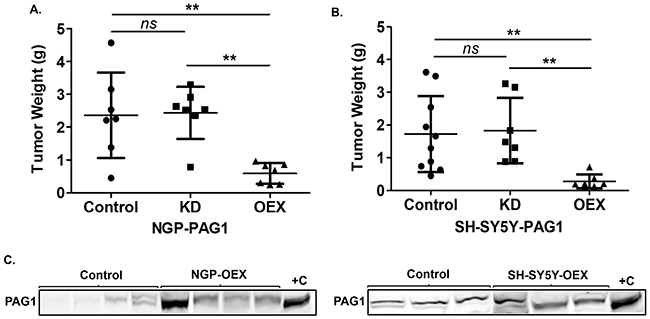
Figure 3: PAG1 is a tumor suppressor in neuroblastoma in vivo. A. Tumor xenografts of different PAG1 gain and loss of function cell lines in NGP and B. SH-SY5Y were developed and tumor weights were analyzed. Data represented for individual tumors in cohorts and as mean ± SD (Mann-Whitney test, ns= non significant, **p<0.01). C. Western immunobloting analysis of tumor samples shows overexpression of PAG1 in NGP-OEX and SH-SY5Y-OEX tumor xenografts in comparison to control NGP and SH-SY5Y xenografts respectively. Protein was isolated from flash frozen tumor tissues. +C represents positive control and is the protein isolated from respective OEX cell line. OEX= PAG1-overexpression, KD= PAG1-knockdown.
PAG1 inhibits c-SRC and downstream signaling pathways
In order to define the mechanism(s) of action for PAG1 as a tumor suppressor, we performed Western blot analysis for c-SRC and associated downstream signaling pathways. We observed that PAG1 over-expression in both NGP and SH-SY5Y cell lines significantly inhibits the phosphorylation and activation of the proto-oncogene c-SRC (phosphorylation at Tyr416). PAG1 over-expression also reduces phosphorylation of downstream RAS pathway effector proteins pERK, pAKT, and pSTAT3 (Figure 4A). In contrast, knockdown of PAG1 expression in both NGP and SH-SY5Y cell lines rescues these effects and induces phosphorylation and activation of c-SRC and downstream effector proteins (Figure 4A). An illustrated summary of the PAG1 tumor suppressor mechanism of action in NB is shown in Figure 4B. The transmembrane adaptor protein PAG1 recruits Csk and brings it in close proximity to active c-SRC. Csk then phosphorylates c-SRC at Tyr527 that leads to inhibition of c-Src kinase functions and consequently downstream effector protein signaling pathways [5].
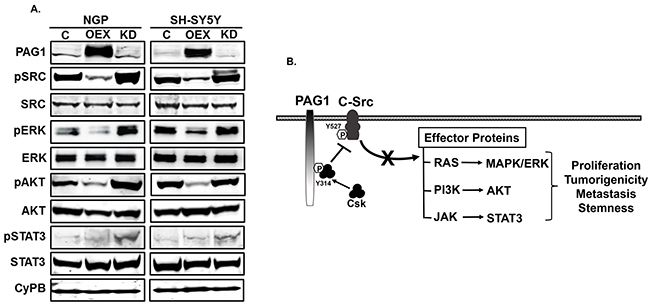
Figure 4: PAG1 inhibits c-Src and downstream effector proteins. A. Western blots of represented proteins in PAG1 overexpression (OEX), knockdown (KD) and control (C) NGP and SH-SY5Y cell lines. CyPB was used as a loading control for all blots, as it is not known and observed to be affected by PAG1 levels in NB cell lines. B. Mechanism of PAG1 action in neuroblastoma. PAG1 acts as an adaptor protein to recruit the Csk from cytoplasm at specific site Tyr-314 and bring it to close proximity with membrane bound c-SRC and other SFKs. Csk then directly inhibit c-Src and other SFks by phosphorylating the Tyr527 [5, 6]. c-Src is known to activate different cellular functions by controlling the effector proteins of different pathways (RAS, PI3K and JAK) either directly or indirectly [4]. Inhibition of c-Src by PAG1 leads to downstream inhibition of different cellular pathways that further translate in to reduction in overall proliferation and tumorigenicity, as confirmed by our in vitro and in vivo results in neuroblastoma.
Transcriptional repression of PAG1 in neuroblastoma
We sought to determine whether PAG1 is directly inhibited by MYCN since MYCN is a primary oncogenic driver of NB and more aggressive tumors typically have increased MYCN expression [19]. MYCN-ChIP-seq data analysis for the PAG1 promoter and 5′UTR demonstrates lack of a canonical CACGTG E-box [16]. However, sequence analysis of the PAG1 5′UTR reveals two non-canonical binding sites (CAGCTG and CATGTG) suggesting that PAG1 could be directly repressed by the MYCN oncogene (Figure 5A). We also found binding sites of several microRNAs, directly or indirectly activated by MYCN in the 3′UTR of PAG1 (Figure 5A and Supplementary Figure S3, S4), including sites for miR17a (a component of the miR-17-92a cluster) and multiple other microRNAs known to contribute to NB pathogenesis [16, 20, 21]. To validate that PAG1 is repressed by MYCN, we analyzed the PAG1 mRNA levels in two independent MYCN overexpression systems. Results shows significant reduction (<0.001) of PAG1 levels in stable MYCN overexpressing CHLA-255-MYCN cell line and inducible MYCN expressing MYCN3 cell line compared to control cell lines (Figure 5B). These findings and the lack of a canonical E-Box binding site suggest that MYCN indirectly represses PAG1 gene expression via its downstream microRNA targets or other mechanisms. We propose that MYCN represses PAG1 to promote c-Src activity and contributes to aggressive disease progression (Figure 5C).
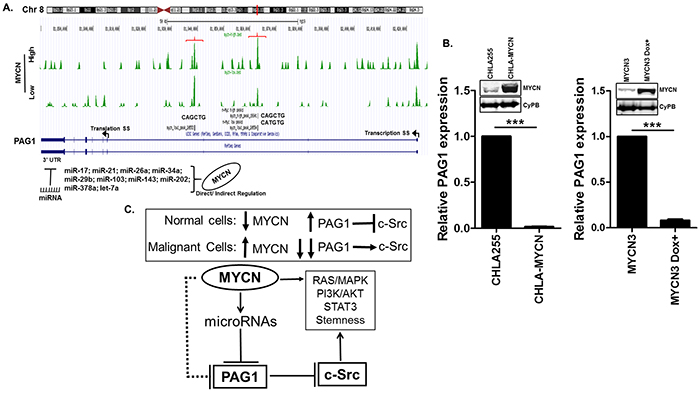
Figure 5: Transcriptional regulation of PAG1 in neuroblastoma. A. MYCN ChIP-seq database [16] analysis in MYCN high and low conditions at the PAG1 gene locus showing two binding sites with non-canonical e-boxes (brackets). Arrows represent the transcription and translation start sites on PAG1 gene. Analysis of PAG1 3′UTR shows binding sites of several miRNAs directly or indirectly activated by MYCN. B. Transcription profiling of PAG1 mRNA expression in two independent MYCN overexpression models in neuroblastoma. CHLA-MYCN cell line stably overexpress MYCN [17] and MYCN3 cell line express MYCN in response to doxycycline induction [16]. MYCN3 cell was either not treated or treated with doxycycline for 24 h. Western Immunoblot for MYCN was performed to validate the expression models and CyPB was used as loading control. PAG1 mRNA expression was analyzed by qPCR and represented as mean ± SD (T-test, ***p<0.001). Experiment was performed in triplicates and repeated thrice. PAG1 mRNA expression in control cell lines CHLA-255 and MYCN3 was used to quantitate relative expression. C. PAG1 repression contributes to neuroblastoma tumorigenesis. In normal non-malignant cells with normal PAG1 levels, c-Src activity is limited by PAG1/Csk induced negative phosphorylation. In malignant cells high levels of MYCN represses PAG1 that results in further c-Src activation. MYCN either directly or indirectly via miRNAs represses PAG1 expression. Inhibition of PAG1 leads to increased c-Src levels that further enhance activation of multiple oncogenic pathways including STAT3, PI3K and others.
DISCUSSION
In summary, we demonstrate that PAG1 significantly alters NB tumor proliferation and xenograft formation. In addition, evaluation of NB patient cohorts demonstrates a significant inverse correlation of PAG1 expression with survival. Overexpression of PAG1 has also been shown to inhibit the growth of breast [22], lung [23], prostate [7] and colon cancer [24] primarily via repression of Src suggesting that repression of PAG1 may also be an important event in NB pathogenesis.
High stage NB displays elevated c-Src activity as well as increased downstream mTOR, PI3K/Akt and RAS/MAPK pathway activation [1, 19]. Earlier studies using tyrosine kinase inhibitors to block Src demonstrated sensitivity of NB cells to the non-specific Src inhibitors such as dasatanib [10], PP2 [11, 25] and other SFK inhibitors [3, 9, 26]. While these studies demonstrated in vitro proliferation arrest and apoptosis, the mechanisms for increased c-Src signaling in NB remained poorly defined. Here we demonstrate that increased expression of PAG1 decreases c-Src activation and inhibits tumor growth in vivo, and this correlates with reduced levels of pAKT and pERK.
PAG1 typically recruits Csk to co-localize and inhibit c-Src in lipid-rafts. Studies with Csk deficient cells demonstrated that PAG1 also inhibits Src through a secondary pathway by directly sequestering Src in lipid-rafts to prevent Src controlled transformation [24]. Additional PAG1 functions include Csk independent repression of RAS [27], binding with EBP5 [28], and regulating SOCS1–mediated ubiquitination and degradation of c-Src and other SFKs [29]. Thus accumulating evidence suggests that transcriptional regulation of PAG1 plays an important role regulating SFKs in cancer via multiple interactions. PAG1 is also reported to be controlled at epigenetic level in cancer cells, and treatment with a histone deacetylase (HDAC) inhibitor as well as by siRNA against HDAC1/2 restore PAG1 expression and activity [30]. We propose that antagomiRs or other approaches to block MYCN driven microRNAs that repress PAG1 transcription may present effective therapeutic strategies for high-risk NB.
There is increasing interest in developing specific inhibitors to c-Src and other non-receptor Src related tyrosine kinases due to the plethora of ascribed oncogenic functions [31]. NDRG1, a suppressor of metastasis is recently shown to inhibit c-Src and downstream signaling pathways in three tumor cell-types [32]. Metastasis represents a major cause of morbidity and mortality for NB and identification of upstream regulator of c-Src, PAG1 provides rationale for novel therapeutic strategies to inhibit this oncogene. The elucidation of the pathway(s) that lead to tumor suppressor PAG1 down-regulation may shed light on the mechanisms of NB tumor progression, and pave the way for developing new therapeutic interventions for children suffering with this deadly malignancy.
MATERIALS AND METHODS
Clinical patient cohorts
NCI-POB dataset include 56 primary patient tumors and their microarray gene expression profiling and clinical outcomes. This dataset is available from Oncogenomics platform (http://home.ccr.cancer.gov/oncology/oncogenomics). Versteeg dataset includes 88 and the Kocak dataset includes 649 [13] unique primary tumor profiles and are publically available from the gene expression databases at the R2: Genomic Analysis and Visualization Platform (http://hgserver1.amc.nl/cgi-bin/r2/main.cgi) [14].
Cell culture
Human neuroblastoma cell lines (NGP, SH-SY5Y, CHLA-255, MYCN3, CHLA-255-MYCN) were routinely maintained and cultured as described previously [15, 16]. CHLA-255-MYCN cell line was courtesy provide by Dr. Leonid Metelitsa, Baylor College of Medicine [17]. All cell lines were validated for CD56 expression and by genotyping within the past 12 months. All cell lines used in this article were routinely tested for Mycoplasma on a monthly basis.
Generating stable PAG1 overexpression and knockdown cell lines
Lentiviral vector plasmids EX-T0343-Lv105-B (Genecopeia, MD) and pGIPZ-PAG1-shRNA set (Thermo Fisher Scientific, MA) were used to transduce NB cell lines for PAG1 overexpression and knockdown as described previously [15]. Set of pGIPZ-PAG1-shRNA include four shRNA vectors and testing with transient transfection in NB cell lines shows that shRNA2 (mature antisense sequence: TCTCTTGTTAGTTTCACTG) efficiently knockdown PAG1, therefore we used this shRNA vector for further study. Non-silencing pGIPZ-control-shRNA vector was used to generate NGP-shNC and SH-SY5Y-shNC cell lines as a control.
Cell proliferation and soft agar colony formation assay
Cell proliferation assay was performed using CellTiter 96® Cell Proliferation Assay (G4000, Promega) according to manufacturer’s instructions and as described previously [15]. Proliferation index was calculated by normalizing each reading to the 24 h time point. Colony formation soft agar assays were performed using standard conditions as described previously [15]. All assays were performed in triplicates and repeated twice with proper controls.
PAG1 quantitative PCR assay
PAG1 mRNA expression in different cell lines were measured using quantitative real-time RT-PCR assay. Extraction of RNA, cDNA synthesis and qPCR was performed as described previously [18]. Assays were performed in triplicates for each sample using Power SYBR Green PCR Master Mix (Applied Biosystems) and normalized to GAPDH. Primers used were, PAG1-F 5′ATCACCCTGTGGGGAAGTCT3′ and PAG1-R 5′TCCTTGTCTGAAGGCACGTT3′, GAPDH-F 5′GGTCGTATTGGGCGCCTGGTC3′ and GAPDH-R 5′GCCAGCATCGCCCCACTTGA3′. Dissociation curves were analyzed for primer pairs as a means to ensure the quality of amplicon and to monitor primer dimers.
Mice and xenograft assay
Four to six week-old female inbred athymic immunodeficient Nude mice (Nu/Nu) (Taconic Biosciences, NY) were used for all xenograft studies. Mice were implanted using a previously described orthotopic xenograft model of NB [15]. Briefly, 1×106 NB cells were surgically implanted in the sub-renal capsule of mice, and tumor growth was monitored bi-weekly by bioluminescent imaging (IVIS Lumina XR System, Caliper Life Sciences, MA). After 28 days of implantation all mice were euthanized and tumors were resected and weighed. All animal experiments are approved by the nstitutional animal care and use committee.
Western blot
Total and phospho- proteins were extracted by lysing cells in RIPA buffer supplemented with protease inhibitor cocktail (Complete mini EDTA free, Roche) and phosphatase inhibitor cocktail (PhosSTOP, Roche). Extracted proteins were analyzed by Western blotting as described previously [15]. Primary antibodies for PAG1 (MAB5285, R&D systems), MYCN (OP13L, EMD Millipore), pSRC (6943), SRC (2102), pERK (4370), ERK (4695), pAKT (4060), AKT (9272), pSTAT3 (9145), STAT3 (4904) were purchased from Cell Signaling Technology, MA, and loading control CyPB antibody (sc20361) from Santa Cruz Biotech, TX.
Statistical analysis
Data values for in vivo experiments are expressed as mean ± SEM and compared using Mann-Whitney (two-tailed nonparametric analysis) test. Student’s t-test (two-tailed or one-tailed distribution with unequal variance) was applied to compare the results shown for in vitro experiments unless otherwise stated. Assays were performed in triplicate and repeated. Correlation between PAG1 and MYCN expression in human NB patients was examined with Pearson’s correlation. Survival analyses were performed using Kaplan-Meier method and two-sided log-rank tests. A p-value less than 0.05 was considered significant.
ACKNOWLEDGMENTS
The Authors would like to express our gratitude to the private foundations funding this work. In addition we thank Dr. Javed Khan for providing access to the Oncogenomics dataset platform and to Dr. Leonid Metelitsa for providing CHLA-MYCN cell line. Authors would also like to thank Dr. Meenakshi Mehrotra for assisting with Ingenuity pathway analysis. This work is dedicated to all the children and parents struggling with neuroblastoma who inspire us to search for a cure.
CONFLICTs OF INTEREST
The authors disclose no potential conflicts of interest.
Authors’ Contributions
S.A. designed and performed experiments, developed methodology, acquired and analyzed data, wrote the paper. R.G. provided materials, analyzed data. Z.C. performed experiments, acquired data. AL performed experiments. P.G. provided materials, analyzed data. E.K. performed experiments, analyzed data, wrote the paper. J.M.S. designed experiments, analyzed data and wrote the paper.
GRANT SUPPORT
This work was supported by grants from American Cancer Society, Alex’s Lemonade Stand Foundation, Gilson-Longenbaugh Foundation, and the Children’s Neuroblastoma Research Foundation to Jason M. Shohet.
REFERENCES
1. Louis CU and Shohet JM. Neuroblastoma: molecular pathogenesis and therapy. Annual review of medicine. 2015; 66:49-63.
2. Liu ST, Pham H, Pandol SJ and Ptasznik A. Src as the link between inflammation and cancer. Frontiers in physiology. 2013; 4:416.
3. Finlay D and Vuori K. Novel noncatalytic role for caspase-8 in promoting SRC-mediated adhesion and Erk signaling in neuroblastoma cells. Cancer research. 2007; 67:11704-11711.
4. Saitou T, Kajiwara K, Oneyama C, Suzuki T and Okada M. Roles of raft-anchored adaptor Cbp/PAG1 in spatial regulation of c-Src kinase. PloS one. 2014; 9:e93470.
5. Okada M. Regulation of the SRC family kinases by Csk. International journal of biological sciences. 2012; 8:1385-1397.
6. Parsons SJ and Parsons JT. Src family kinases, key regulators of signal transduction. Oncogene. 2004; 23:7906-7909.
7. Svec A. Phosphoprotein associated with glycosphingolipid-enriched microdomains/Csk-binding protein: a protein that matters. Pathology, research and practice. 2008; 204:785-792.
8. Oneyama C and Okada M. MicroRNAs as the fine-tuners of Src oncogenic signalling. Journal of biochemistry. 2015; 157:431-438.
9. Radi M, Brullo C, Crespan E, Tintori C, Musumeci F, Biava M, Schenone S, Dreassi E, Zamperini C, Maga G, Pagano D, Angelucci A, Bologna M, et al. Identification of potent c-Src inhibitors strongly affecting the proliferation of human neuroblastoma cells. Bioorganic & medicinal chemistry letters. 2011; 21:5928-5933.
10. Vitali R, Mancini C, Cesi V, Tanno B, Piscitelli M, Mancuso M, Sesti F, Pasquali E, Calabretta B, Dominici C and Raschella G. Activity of tyrosine kinase inhibitor Dasatinib in neuroblastoma cells in vitro and in orthotopic mouse model. International journal of cancer. 2009; 125:2547-2555.
11. Hishiki T, Saito T, Sato Y, Mitsunaga T, Terui E, Matsuura G, Saito E, Shibata R, Mise N, Yokoyama Y and Yoshida H. Src kinase family inhibitor PP2 induces aggregation and detachment of neuroblastoma cells and inhibits cell growth in a PI3 kinase/Akt pathway-independent manner. Pediatric surgery international. 2011; 27:225-230.
12. Reynolds AB, Kanner SB, Bouton AH, Schaller MD, Weed SA, Flynn DC and Parsons JT. SRChing for the substrates of Src. Oncogene. 2014; 33:4537-4547.
13. Kocak H, Ackermann S, Hero B, Kahlert Y, Oberthuer A, Juraeva D, Roels F, Theissen J, Westermann F, Deubzer H, Ehemann V, Brors B, Odenthal M, et al. Hox-C9 activates the intrinsic pathway of apoptosis and is associated with spontaneous regression in neuroblastoma. Cell death & disease. 2013; 4:e586.
14. R2: Genomics Analysis and Visualization Platform. http://r2.amc.nl.
15. Agarwal S, Lakoma A, Chen Z, Hicks J, Metelitsa LS, Kim ES and Shohet JM. G-CSF Promotes Neuroblastoma Tumorigenicity and Metastasis via STAT3-Dependent Cancer Stem Cell Activation. Cancer research. 2015; 75:2566-2579.
16. Shohet JM, Ghosh R, Coarfa C, Ludwig A, Benham AL, Chen Z, Patterson DM, Barbieri E, Mestdagh P, Sikorski DN, Milosavljevic A, Kim ES and Gunaratne PH. A genome-wide search for promoters that respond to increased MYCN reveals both new oncogenic and tumor suppressor microRNAs associated with aggressive neuroblastoma. Cancer research. 2011; 71:3841-3851.
17. Song L, Ara T, Wu HW, Woo CW, Reynolds CP, Seeger RC, DeClerck YA, Thiele CJ, Sposto R and Metelitsa LS. Oncogene MYCN regulates localization of NKT cells to the site of disease in neuroblastoma. The Journal of clinical investigation. 2007; 117:2702-2712.
18. Hsu DM, Agarwal S, Benham A, Coarfa C, Trahan DN, Chen Z, Stowers PN, Courtney AN, Lakoma A, Barbieri E, Metelitsa LS, Gunaratne P, Kim ES. et al. G-CSF receptor positive neuroblastoma subpopulations are enriched in chemotherapy-resistant or relapsed tumors and are highly tumorigenic. Cancer research. 2013; 73:4134-4146.
19. Brodeur GM and Bagatell R. Mechanisms of neuroblastoma regression. Nature reviews Clinical oncology. 2014; 11:704-713.
20. Li Y, Choi PS, Casey SC, Dill DL and Felsher DW. MYC through miR-17-92 suppresses specific target genes to maintain survival, autonomous proliferation, and a neoplastic state. Cancer cell. 2014; 26:262-272.
21. Buechner J and Einvik C. N-myc and noncoding RNAs in neuroblastoma. Molecular cancer research. 2012; 10:1243-1253.
22. Jiang LQ, Feng X, Zhou W, Knyazev PG, Ullrich A and Chen Z. Csk-binding protein (Cbp) negatively regulates epidermal growth factor-induced cell transformation by controlling Src activation. Oncogene. 2006; 25:5495-5506.
23. Kanou T, Oneyama C, Kawahara K, Okimura A, Ohta M, Ikeda N, Shintani Y, Okumura M and Okada M. The transmembrane adaptor Cbp/PAG1 controls the malignant potential of human non-small cell lung cancers that have c-src upregulation. Molecular cancer research. 2011; 9:103-114.
24. Oneyama C, Hikita T, Enya K, Dobenecker MW, Saito K, Nada S, Tarakhovsky A and Okada M. The lipid raft-anchored adaptor protein Cbp controls the oncogenic potential of c-Src. Molecular cell. 2008; 30:426-436.
25. Beierle EA, Ma X, Trujillo A, Kurenova EV, Cance WG and Golubovskaya VM. Inhibition of focal adhesion kinase and src increases detachment and apoptosis in human neuroblastoma cell lines. Molecular carcinogenesis. 2010; 49:224-234.
26. Navarra M, Celano M, Maiuolo J, Schenone S, Botta M, Angelucci A, Bramanti P and Russo D. Antiproliferative and pro-apoptotic effects afforded by novel Src-kinase inhibitors in human neuroblastoma cells. BMC cancer. 2010; 10:602.
27. Smida M, Posevitz-Fejfar A, Horejsi V, Schraven B and Lindquist JA. A novel negative regulatory function of the phosphoprotein associated with glycosphingolipid-enriched microdomains: blocking Ras activation. Blood. 2007; 110:596-615.
28. Itoh K, Sakakibara M, Yamasaki S, Takeuchi A, Arase H, Miyazaki M, Nakajima N, Okada M and Saito T. Cutting edge: negative regulation of immune synapse formation by anchoring lipid raft to cytoskeleton through Cbp-EBP50-ERM assembly. Journal of immunology. 2002; 168:541-544.
29. Whiting RJ, Payne CJ, Satiaputra J, Kucera N, Qiu TW, Irtegun S, Gunn NJ, Slavova-Azmanova NS, Mulhern TD and Ingley E. Targeting Lyn tyrosine kinase through protein fusions encompassing motifs of Cbp (Csk-binding protein) and the SOCS box of SOCS1. The Biochemical journal. 2012; 442:611-620.
30. Suzuki K, Oneyama C, Kimura H, Tajima S and Okada M. Down-regulation of the tumor suppressor C-terminal Src kinase (Csk)-binding protein (Cbp)/PAG1 is mediated by epigenetic histone modifications via the mitogen-activated protein kinase (MAPK)/phosphatidylinositol 3-kinase (PI3K) pathway. The Journal of biological chemistry. 2011; 286:15698-15706.
31. Roskoski R, Jr. Src protein-tyrosine kinase structure, mechanism, and small molecule inhibitors. Pharmacological research. 2015; 94:9-25.
32. Liu W, Yue F, Zheng M, Merlot A, Bae DH, Huang M, Lane D, Jansson P, Lui GY, Richardson V, Sahni S, Kalinowski D, Kovacevic Z and Richardson DR. The proto-oncogene c-Src and its downstream signaling pathways are inhibited by the metastasis suppressor, NDRG1. Oncotarget. 2015; 6:8851-8874. doi: 10.18632/oncotarget.3316