
INTRODUCTION
Malignant pleural mesothelioma (MPM) is a biologically highly aggressive tumour arising from the pleura leading to a dismal prognosis [1–5].
Nowadays, cisplatin is the drug of choice for the treatment of MPM, and carboplatin seems to have comparable efficacy [1, 6–10]. Combined with antifolates they are considered as the most effective regimen for MPM [10–13]. However, patients only show response rates of approximately 40% with a progression-free survival (PFS) of 5.7 months [14].
In contrast to other solid tumours, mutations of the TP53 gene are extremely rare in MPM, so other mechanisms such as deletion of the locus or methylation contribute to inactivation of TP53 [15–17]. Overexpression of MDM2 in some tumour types can lead to a loss of TP53 regulatory function in cancer cells by its increased proteasomal degradation [18–23]. P14/ARF, the physiological inhibitor of MDM2 is recognized as a tumour suppressor and induces cell cycle arrest in a TP53-dependent and -independent manner [24–27]. Analysis of the signalling relationship between these genes indicates an additional role of RB1 in this signalling network [28–30]. In previous studies, we have demonstrated that MDM2 is strongly expressed in the nuclei in approximately 25% of MPM affecting only epithelioid or the epithelioid components of biphasic MPM [23, 31]. Moreover, patients with MDM2-positive MPM showed a significantly decreased overall survival (OS) and progression-free survival (PFS) compared to MDM2-negative MPM [23, 31]. This might be explained by a significantly decreased or completely abolished TP53 activity and/or stability mediated by an overexpression of MDM2 [18–22]. A promising approach to explain the differences in MDM2 expression is a differential expression of miRNAs [32], targeting some of these key enzymes and thereby reducing expression levels of those.
The recent study is designed to analyse the miRNA profile of pleural mesotheliomas with respect to immunohistochemical MDM2 expression (score 0 versus score ≥1) using the nCounter system (NanoString), a hybridization-based digital detection method that can be used to analyse mRNA, miRNA and DNA [31, 33–36].
RESULTS
136 miRNAs significantly differentially regulated between MDM2-positive and -negative samples were identified. 39 of them showed a p-value <0.01 and six of them (miR-106-5p, miR-15b-5p, miR24, miR-29a, miR-29c and miR-130a) a p-value <0.001.
Six miRNAs with p<0.01 were increased, whereas the remaining were decreased in MDM2-expression positive tumours.
All significant p-values are summarized in suppl. Table 1.
Furthermore, 17 miRNAs inhibiting TP53, 18 miRNAs binding MDM2 and eleven miRNAs suppressing CDKN2A were significantly decreased in MDM2-expressing mesotheliomas. Of note, five miRNAs (miR-29a, miR-29b, miR-29c, miR-125a, miR-125b) binding all three targets, five regulating both TP53 and CDKN2A (let-7a, let-7c, let-7d, let-7e, let-7g), and three binding both MDM2 and TP53 (miR-34a, miR-145, miR-185) were found to be significantly decreased, respectively. TP53 is predicted to be suppressed in MDM2-negative tumours by miRNAs with a prediction score (PS) of 11.67, RB1 with a PS of 8.02, CDKN2A with a PS 1.27 and MDM2 with a PS of 4.50 (Figure 1).
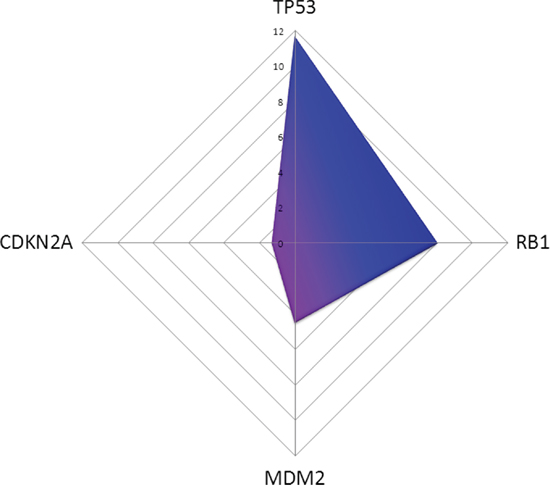
Figure 1: In silico prediction of miRNAs affecting TP53 pathway members in MDM2-negativ tumours compared to MDM2-positive ones is shown. TP53 is downregulated with a prediction score (PS) of 11.67, RB1 with a PS of 8.02, CDKN2A with a PS 1.27 and MDM2 with a PS of 4.50.
A summary of the miRNAs, the affected target, correlating p-values and correlation coefficients is given in Table 1.
Table 1: An overview of significances of miRNAs directly regulating TP53-pathway members in MDM2-expressing and non-expressing tumours is shown
Target |
miRNA Expression |
p-Value |
Correlation Coefficient |
|---|---|---|---|
TP53/CDKN2A/MDM2 |
hsa-miR-125a |
0.0078 |
-0.6030 |
TP53/CDKN2A/MDM2 |
hsa-miR-125b |
0.0019 |
-0.6316 |
TP53/CDKN2A/MDM2 |
hsa-miR-29a |
0.0000 |
-0.4055 |
TP53/CDKN2A/MDM2 |
hsa-miR-29b |
0.0344 |
-0.4055 |
TP53/CDKN2A/MDM2 |
hsa-miR-29c |
0.0000 |
-0.4055 |
TP53/CDKN2A |
hsa-let-7a |
0.0014 |
-0.5711 |
TP53/CDKN2A |
hsa-let-7c |
0.0117 |
-0.5711 |
TP53/CDKN2A |
hsa-let-7d |
0.0014 |
-0.5711 |
TP53/CDKN2A |
hsa-let-7e |
0.0142 |
-0.5711 |
TP53/CDKN2A |
hsa-let-7g |
0.0246 |
-0.5711 |
MDM2/TP53 |
hsa-miR-34a |
0.0006 |
|
MDM2/TP53 |
hsa-miR-145 |
0.0063 |
-0.3741 |
MDM2/TP53 |
hsa-miR-185 |
0.0470 |
-0.2059 |
CDKN2A |
hsa-miR-340 |
0.0470 |
-0.1303 |
MDM2 |
hsa-miR-140 |
0.0063 |
-0.1286 |
MDM2 |
hsa-miR-223 |
0.0206 |
-0.2969 |
MDM2 |
hsa-miR-23b |
0.0011 |
-0.1197 |
MDM2 |
hsa-miR-142 |
0.0246 |
|
MDM2 |
hsa-miR-191 |
0.0008 |
|
MDM2 |
hsa-miR-331 |
0.0403 |
|
MDM2 |
hsa-miR-605 |
0.0292 |
|
MDM2 |
hsa-miR-548d |
0.0142 |
|
MDM2 |
hsa-miR-374b |
0.0172 |
-0.1462 |
MDM2 |
hsa-miR-383 |
0.0292 |
-0.1209 |
TP53 |
hsa-miR-19b |
0.0031 |
-0.2945 |
TP53 |
hsa-miR-218 |
0.0050 |
-0.1678 |
TP53 |
hsa-miR-22 |
0.0078 |
-0.1955 |
TP53 |
hsa-miR-27b |
0.0019 |
-0.5795 |
Some pathways are stimulated in MDM2-positive tumours by decreased miRNA expression. In silico, the most influenced KEGG-pathways between MDM2-positive and negative tumours with respect to the expression levels are Ribosome (eight targets, PS 36.24), Wnt signaling pathway (122 targets, PS 29.45), metabolism of xenobiotics by cytochrome P450 (seven targets, PS 24.14), MAPK signaling pathway (184 targets, PS 23.49), focal adhesion (145 targets, PS 21.53), regulation of actin cytoskeleton (152 targets, PS 19.46), axon guidance (99 targets, PS 19.26), colorectal cancer (73 targets, PS 18.11), oxidative phosphorylation (35 targets, PS 16.49) and TGF-beta signaling pathway (74 targets, PS 15.87).
An overview of all differentially stimulated pathways can be found in suppl. Table 2. The results are visualized in Figure 2.
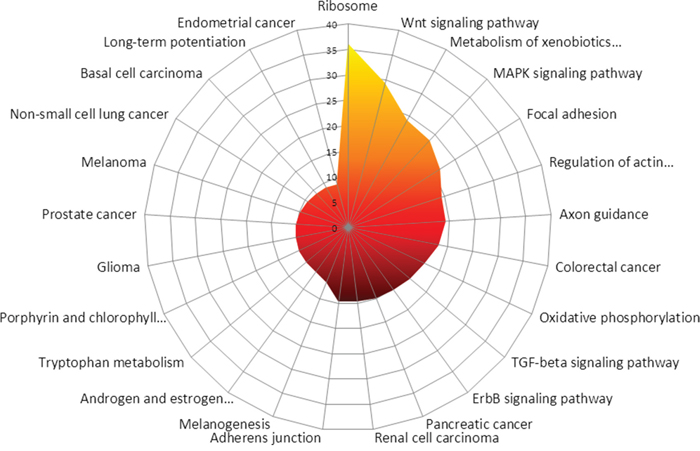
Figure 2: In silico prediction of the most affected KEGG-pathways between MDM2-positive and negative tumours is shown. Most likely affected are Ribosome, Wnt signaling pathway, Metabolism of xenobiotics by cytochrome P450, MAPK signaling pathway, focal adhesion, regulation of actin cytoskeleton, Axon guidance, Colorectal cancer, Oxidative phosphorylation as well as TGF-beta signaling pathway.
DISCUSSION
Recently, a number of miRNAs targeting both TP53 and MDM2 have been identified. In TP53 wild type plasmocytomas, TP53 can induce the expression of miR-192, miR-194 and miR-215, subsequently decreasing MDM2 expression. Hypermethylation of the promoter region of all three miRNAs impairs the MDM2 suppression, still resulting in a blunted TP53 function [38]. Additionally, miR-143, miR-45, miR-605, miR-34a and miR-29b are direct transcriptional targets of TP53 [44]. Together with the three former ones, they form a TP53-positive feedback loop by decreasing MDM2 and HDM4 expression levels [45, 46]. Nevertheless, 10 miRNAs suppressing MDM2 showing low overall expression levels (Figure 3) but still seem to be associated to TP53 activity and thereby are also significantly decreased in MDM2-positive tumours (Table 1).
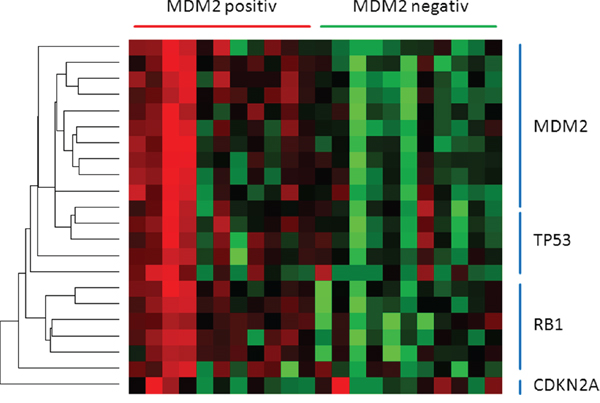
Figure 3: The heatmap presents miRNAs directing TP53 pathway members. miRNAs regulating either MDM2, TP53 or RB1 are downregulated in MDM2-positiv tumours. CDKN2A seems to be unaffected.
TP53 shows the strongest predicted suppression via miRNAs in MDM2-positive tumours with an overall PS of 11.67. Active TP53 stimulates the DROSHA complex, mediating the processing of tumour suppressor miRNAs [44, 47]. As already demonstrated, differences between epithelioid and sarcomatoid mesotheliomas regarding miR-34a expression were found [48]. These differences may be explained by the high incidence of loss of heterozygosity (LOH) in sarcomatoid mesotheliomas, whereas epitheloid subtypes more often show functional inhibition of TP53 via MDM2 [23, 31]. Furthermore, it has been reported that epigenetic silencing of miR-34 family members plays an important role in the pathogenesis of a majority of MPMs [49].
MDM2 suppresses wild type TP53 leading to a more or less impaired but still specific transcriptional activity [50]. This may shift to other promoters e.g. controlling oncogenic miR-128b. Of note, activation of TP53 leads to an genome-wide change of miRNA pattern, indicating that TP53 has a complex role in regulating miRNAs (Figure 4) [51, 52].
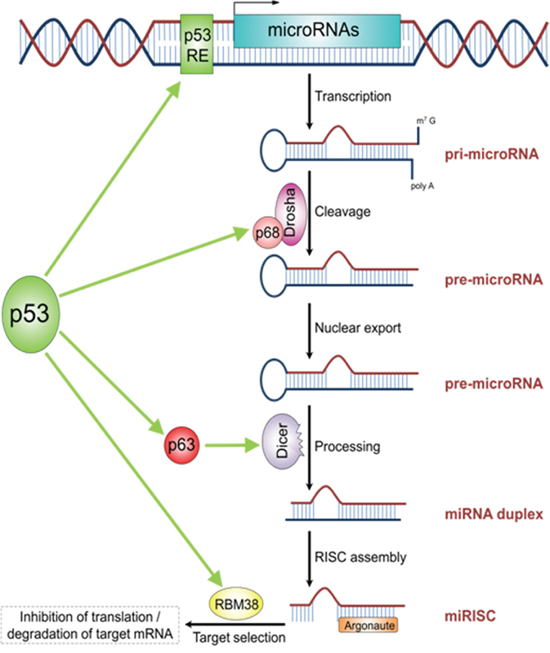
Figure 4: The schematic illustrations describes the role of TP53 in miRNA processing and regulation. TP53 acts as transcription factor for some miRNAs. Additionally, TP53 plays a role in miRNA processing by directly interacting with P68 (DROSHA-complex), P63 (DICER-complex) as well as RBM38. The figure is adapted from Rokavec et al. Clin Exp Gastroenterol, 2014 [44].
Due to the high number of targets measured in a relatively low number of cases, a classical p-value adjustment by FDR or Bonferroni correction was not suitable. To our opinion, this study should be more seen like a screening study to get a first insight in possible regulatory mechanisms but therefore is still of importance to smooth the way for larger validation cohorts.
An interesting finding was the strong difference in activation of the ribosomal pathway predicted in silico. RPS6 encoding for the ribosomal protein S6, but also other ribosomal components, rendered high PS (suppl. Table 2), suggesting it plays an important role in pleural tumour progression. Multiple studies reported RPS6 as a predictive marker in pleural mesotheliomas with platinum-based chemotherapy [53, 54], but these findings may be coincidental. More likely, the MDM2 driven knock-down of TP53 can predict the response to platinum-based regimes and the subsequent regulation of ribosomal components acts as an indirect parameter. RL26 can directly bind TP53 and therefore form a positive feedback loop with MDM2 [55, 56].
In summary, 136 miRNAs significantly differentially expressed in MDM2-positive and -negative tumours were identified. Furthermore, 17 miRNAs directly binding TP53, 18 miRNAs directly suppressing MDM2, and eleven miRNAs directly inhibiting CDKN2A are significantly decreased in MDM2-expressing mesotheliomas. TP53 is predicted to be strongly suppressed by miRNA depending on expression pattern, whereas the impact on MDM2 itself is just moderate and on CDKN2A is even weaker. The present data lead to the conclusion that MDM2 expression level noticeably impacts the overall miRNA expression and thereby cellular pathways (e.g. ribosomal translation) in MPM. This can lead to a better understanding of the biology of MPM and may open the chance for new therapeutic approaches and thereby augment patients’ outcome.
MATERIALS AND METHODS
Study design
For this exploratory miRNA study, twenty-four formalin-fixed paraffin-embedded (FFPE) tumour specimens were screened with respect to their MDM2 immunoexpression (twelve MDM2-positive and twelve MDM2-negative MPM) as described previously [31]. Tumor classification is based on the WHO classification of tumours guidelines (2004) [37], TNM-staging is based on the UICC classification of malignant tumours [38]. Both were confirmed by two experienced pathologists (JWO, KWS). The study included only patients with MPM, treated at the West German Cancer Centre or the West German Lung Centre between 2006 and 2009. Clinicopathological data including age, gender, histology and stages are summarized in Table 1. Surveillance for this study was stopped on August 31, 2014. The retrospective study was approved by the Ethics Committee of the Medical Faculty of the University Duisburg-Essen (identifier: 14-5775-BO). The investigation conforms to the principles outlined in the declaration of Helsinki.
RNA extraction and RNA integrity assessment
According to the manufacturer’s recommendations, three to five paraffin sections with a thickness of 4 μm per sample were deparaffinised with xylene prior to total RNA extraction including small RNAs using the miRNeasy FFPE kit (Qiagen, Venlo, Netherlands). RNA concentrations were measured using a Qubit 2.0 fluorometer with the appertaining RNA broad-range assay (Thermo Fisher Scientific, WA, USA). RNA integrity was assessed using a Fragment Analyzer (Advanced Analytical Inc., Ames, IA, USA).
NanoString CodeSet design and expression quantification
The commercially available human miRNA V2.1 code set containing probes and miRTags for the 800 most important miRNAs described in the context of cancerogenic events was chosen for miRNA expression analysis. Probe sets and miRTags for each target in the CodeSet were designed and synthesized by NanoString Technologies (Seattle, WA, USA). 200 ng total RNA of each sample was processed. For the sample preparation, the high-sensitivity program was chosen. The cartridge was read with maximum sensitivity (555 FOV).
NanoString data processing and statistical analysis
All statistical analyses were calculated with the R statistical programming environment (v2.15.2). NanoString data processing was done using the NanoStringNorm package [34]. Considering the counts obtained for positive control probe sets raw NanoString counts for each gene were subjected to a technical factorial normalization. Mean background plus 2x standard deviations were subtracted for background correction. Additionally, samples with less than mean background plus 2x standard deviations were interpreted as not expressed to overcome basal noise. After this procedure a biological normalization using the reference genes ACTB and GAPDH included in the CodeSet was performed. For in silico prediction of the functional impact of miRNAs we used DIANA-microT v4.0 [39], PicTar 4-way [40] as well as TargetScan5 [41]. KEGG-Pathway analysis was performed by the DIANA-mirPath tool for multiple miRNA analysis [42]. Analysis of already validated miRNA-target interactions was performed using the miRWalk database [43].
For statistical analysis of dichotomous factors such as gender and MDM2-positivity, the Wilcoxon Mann-Whitney rank sum test was applied. Associations between gene expression of tested genes and associations between gene expression and TNM-criteria were analysed by using the Spearman’s rank correlation test.
The level of statistical significance was defined as p≤0.05.
CONFLICTS OF INTEREST
All authors state that they have no conflicts of interest to declare.
ACKNOWLEDGMENTS
The skilful assistance of Dr. Dido Lenze is gratefully acknowledged.
REFERENCES
1. Goudar RK. Review of pemetrexed in combination with cisplatin for the treatment of malignant pleural mesothelioma. Therapeutics and clinical risk management. 2008; 4:205-211.
2. Liu Z and Klominek J. Regulation of matrix metalloprotease activity in malignant mesothelioma cell lines by growth factors. Thorax. 2003; 58:198-203.
3. Antman KH. Clinical presentation and natural history of benign and malignant mesothelioma. Seminars in oncology. 1981; 8:313-320.
4. Hazarika M, White RM, Jr., Booth BP, Wang YC, Ham DY, Liang CY, Rahman A, Gobburu JV, Li N, Sridhara R, Morse DE, Lostritto R, Garvey P, Johnson JR and Pazdur R. Pemetrexed in malignant pleural mesothelioma. Clinical cancer research. 2005; 11:982-992.
5. Weill H, Hughes JM and Churg AM. Changing trends in US mesothelioma incidence. Occupational and environmental medicine. 2004; 61:438-441.
6. Ramalingam SS and Belani CP. Recent advances in the treatment of malignant pleural mesothelioma. Journal of thoracic oncology. 2008; 3:1056-1064.
7. Vorobiof DA and Mafafo K. Malignant pleural mesothelioma: medical treatment update. Clinical lung cancer. 2009; 10:112-117.
8. Ceresoli GL, Castagneto B, Zucali PA, Favaretto A, Mencoboni M, Grossi F, Cortinovis D, Del Conte G, Ceribelli A, Bearz A, Salamina S, De Vincenzo F, Cappuzzo F, Marangolo M, Torri V and Santoro A. Pemetrexed plus carboplatin in elderly patients with malignant pleural mesothelioma: combined analysis of two phase II trials. British journal of cancer. 2008; 99:51-56.
9. Ceresoli GL, Zucali PA, Favaretto AG, Grossi F, Bidoli P, Del Conte G, Ceribelli A, Bearz A, Morenghi E, Cavina R, Marangolo M, Parra HJ and Santoro A. Phase II study of pemetrexed plus carboplatin in malignant pleural mesothelioma. Journal of clinical oncology. 2006; 24:1443-1448.
10. Kindler HL. Systemic treatments for mesothelioma: standard and novel. Current treatment options in oncology. 2008; 9:171-179.
11. Tomek S and Manegold C. Chemotherapy for malignant pleural mesothelioma: past results and recent developments. Lung cancer. 2004; 45:S103-119.
12. Mairinger F, Vollbrecht C, Halbwedl I, Hatz M, Stacher E, Gully C, Quehenberger F, Stephan-Falkenau S, Kollmeier J, Roth A, Mairinger T and Popper H. Reduced folate carrier and folylpolyglutamate synthetase, but not thymidylate synthase predict survival in pemetrexed-treated patients suffering from malignant pleural mesothelioma. Journal of thoracic oncology. 2013; 8:644-653.
13. Mairinger F, Vollbrecht C, Mairinger T and Popper H. The issue of studies evaluating biomarkers which predict outcome after pemetrexed-based chemotherapy in malignant pleural mesothelioma. Journal of thoracic oncology. 2013; 8:e80-82.
14. Vogelzang NJ, Rusthoven JJ, Symanowski J, Denham C, Kaukel E, Ruffie P, Gatzemeier U, Boyer M, Emri S, Manegold C, Niyikiza C and Paoletti P. Phase III study of pemetrexed in combination with cisplatin versus cisplatin alone in patients with malignant pleural mesothelioma. Journal of clinical oncology. 2003; 21:2636-2644.
15. Papp T, Schipper H, Pemsel H, Bastrop R, Muller KM, Wiethege T, Weiss DG, Dopp E, Schiffmann D and Rahman Q. Mutational analysis of N-ras, p53, p16INK4a, p14ARF and CDK4 genes in primary human malignant mesotheliomas. International journal of oncology. 2001; 18:425-433.
16. Papp T, Schipper H, Pemsel H, Unverricht M, Muller KM, Wiethege T, Schiffmann D and Rahman Q. Mutational analysis of the PTEN/MMAC1 tumour suppressor gene in primary human malignant mesotheliomas. Oncology reports. 2001; 8:1375-1379.
17. Toyooka S, Kishimoto T and Date H. Advances in the molecular biology of malignant mesothelioma. Acta medica Okayama. 2008; 62:1-7.
18. Jones SN, Roe AE, Donehower LA and Bradley A. Rescue of embryonic lethality in Mdm2-deficient mice by absence of p53. Nature. 1995; 378:206-208.
19. Marine JC, Francoz S, Maetens M, Wahl G, Toledo F and Lozano G. Keeping p53 in check: essential and synergistic functions of Mdm2 and Mdm4. Cell death and differentiation. 2006; 13:927-934.
20. Montes de Oca Luna R, Wagner DS and Lozano G. Rescue of early embryonic lethality in mdm2-deficient mice by deletion of p53. Nature. 1995; 378:203-206.
21. Parant J, Chavez-Reyes A, Little NA, Yan W, Reinke V, Jochemsen AG and Lozano G. Rescue of embryonic lethality in Mdm4-null mice by loss of Trp53 suggests a nonoverlapping pathway with MDM2 to regulate p53. Nature genetics. 2001; 29:92-95.
22. Ringshausen I, O'Shea CC, Finch AJ, Swigart LB and Evan GI. Mdm2 is critically and continuously required to suppress lethal p53 activity in vivo. Cancer cell. 2006; 10:501-514.
23. Mairinger FD, Walter RF, Ting S, Vollbrecht C, Kollmeier J, Griff S, Hager T, Mairinger T, Christoph DC, Theegarten D, Kurt Werner S and Wohlschlaeger J. Mdm2 protein expression is strongly associated with survival in malignant pleural mesothelioma. Future oncology. 2014; 10:995-1005.
24. Kanellou P, Zaravinos A, Zioga M and Spandidos DA. Deregulation of the tumour suppressor genes p14(ARF), p15(INK4b), p16(INK4a) and p53 in basal cell carcinoma. The British journal of dermatology. 2009; 160:1215-1221.
25. Chen D, Kon N, Li M, Zhang W, Qin J and Gu W. ARF-BP1/Mule is a critical mediator of the ARF tumor suppressor. Cell. 2005; 121:1071-1083.
26. Miao L, Song Z, Jin L, Zhu YM, Wen LP and Wu M. ARF antagonizes the ability of Miz-1 to inhibit p53-mediated transactivation. Oncogene. 2010; 29:711-722.
27. Huang Y, Tyler T, Saadatmandi N, Lee C, Borgstrom P and Gjerset RA. Enhanced tumor suppression by a p14ARF/p53 bicistronic adenovirus through increased p53 protein translation and stability. Cancer research. 2003; 63:3646-3653.
28. Martin JW, Chilton-MacNeill S, Koti M, van Wijnen AJ, Squire JA and Zielenska M. Digital expression profiling identifies RUNX2, CDC5L, MDM2, RECQL4, and CDK4 as potential predictive biomarkers for neo-adjuvant chemotherapy response in paediatric osteosarcoma. PloS one. 2014; 9:e95843.
29. Xu XL, Fang Y, Lee TC, Forrest D, Gregory-Evans C, Almeida D, Liu A, Jhanwar SC, Abramson DH and Cobrinik D. Retinoblastoma has properties of a cone precursor tumor and depends upon cone-specific MDM2 signaling. Cell. 2009; 137:1018-1031.
30. Xu XL, Singh HP, Wang L, Qi DL, Poulos BK, Abramson DH, Jhanwar SC and Cobrinik D. Rb suppresses human cone-precursor-derived retinoblastoma tumours. Nature. 2014; 514:385-388.
31. Walter RF, Werner R, Ting S, Vollbrecht C, Theegarten D, Christoph DC, Kurt Werner S, Wohlschlaeger J and Mairinger FD. Identification of deregulation of apoptosis and cell cycle in neuroendocrine tumors of the lung via NanoString nCounter expression analysis. Oncotarget. 2015; 6:24690-24698. doi: 10.18632/oncotarget.3992.
32. Mairinger FD, Ting S, Werner R, Walter RF, Hager T, Vollbrecht C, Christoph D, Worm K, Mairinger T, Sheu-Grabellus SY, Theegarten D, Schmid KW and Wohlschlaeger J. Different micro-RNA expression profiles distinguish subtypes of neuroendocrine tumors of the lung: results of a profiling study. Modern pathology: an official journal of the United States and Canadian Academy of Pathology, Inc. 2014; 27:1632-1640.
33. Geiss GK, Bumgarner RE, Birditt B, Dahl T, Dowidar N, Dunaway DL, Fell HP, Ferree S, George RD, Grogan T, James JJ, Maysuria M, Mitton JD, Oliveri P, Osborn JL, Peng T, et al. Direct multiplexed measurement of gene expression with color-coded probe pairs. Nature biotechnology. 2008; 26:317-325.
34. Waggott D, Chu K, Yin S, Wouters BG, Liu FF and Boutros PC. NanoStringNorm: an extensible R package for the pre-processing of NanoString mRNA and miRNA data. Bioinformatics. 2012; 28:1546-1548.
35. Walter RF, Mairinger FD, Werner R, Ting S, Vollbrecht C, Theegarten D, Christoph DC, Zarogoulidis K, Schmid KW, Zarogoulidis P and Wohlschlaeger J. SOX4, SOX11 and PAX6 mRNA expression was identified as a (prognostic) marker for the aggressiveness of neuroendocrine tumors of the lung by using next-generation expression analysis (NanoString). Future oncology. 2015; 11:1027-1036.
36. Mairinger FD, Walter RF, Werner R, Christoph DC, Ting S, Vollbrecht C, Zarogoulidis K, Huang H, Li Q, Schmid KW, Wohlschlaeger J and Zarogoulidis P. Activation of angiogenesis differs strongly between pulmonary carcinoids and neuroendocrine carinomas and is crucial for carcinoid tumourgenesis. Journal of Cancer. 2014; 5:465-471.
37. Travis WD, World Health Organization., International Agency for Research on Cancer., International Association for the Study of Lung Cancer. and International Academy of Pathology. (2004). Pathology and genetics of tumours of the lung, pleura, thymus and heart. (Lyon Oxford: IARC Press Oxford University Press (distributor)).
38. Sobin LH, Gospodarowicz MK, Wittekind C and International Union against Cancer. (2010). TNM classification of malignant tumours. (Chichester, West Sussex, UK ; Hoboken, NJ: Wiley-Blackwell).
39. Maragkakis M, Vergoulis T, Alexiou P, Reczko M, Plomaritou K, Gousis M, Kourtis K, Koziris N, Dalamagas T and Hatzigeorgiou AG. DIANA-microT Web server upgrade supports Fly and Worm miRNA target prediction and bibliographic miRNA to disease association. Nucleic acids research. 2011; 39:W145-148.
40. Chen K and Rajewsky N. Natural selection on human microRNA binding sites inferred from SNP data. Nature genetics. 2006; 38:1452-1456.
41. Agarwal V, Bell GW, Nam JW and Bartel DP. Predicting effective microRNA target sites in mammalian mRNAs. eLife. 2015; 4.
42. Vlachos IS, Zagganas K, Paraskevopoulou MD, Georgakilas G, Karagkouni D, Vergoulis T, Dalamagas T and Hatzigeorgiou AG. DIANA-miRPath v3.0: deciphering microRNA function with experimental support. Nucleic acids research. 2015; 43:W460-466.
43. Dweep H, Gretz N and Sticht C. miRWalk database for miRNA-target interactions. Methods Mol Biol. 2014; 1182:289-305.
44. Rokavec M, Li H, Jiang L and Hermeking H. The p53/microRNA connection in gastrointestinal cancer. Clinical and experimental gastroenterology. 2014; 7:395-413.
45. Mandke P, Wyatt N, Fraser J, Bates B, Berberich SJ and Markey MP. MicroRNA-34a modulates MDM4 expression via a target site in the open reading frame. PloS one. 2012; 7:e42034.
46. Okada N, Lin CP, Ribeiro MC, Biton A, Lai G, He X, Bu P, Vogel H, Jablons DM, Keller AC, Wilkinson JE, He B, Speed TP and He L. A positive feedback between p53 and miR-34 miRNAs mediates tumor suppression. Genes & development. 2014; 28:438-450.
47. Brachova P, Mueting SR, Devor EJ and Leslie KK. Oncomorphic 53 Mutations in Gynecologic Cancers Lose the Normal Protein:Protein Interactions with the microRNA Microprocessing Complex. Journal of cancer therapy. 2014; 5:506-516.
48. Ghawanmeh T, Thunberg U, Castro J, Murray F and Laytragoon-Lewin N. miR-34a expression, cell cycle arrest and cell death of malignant mesothelioma cells upon treatment with radiation, docetaxel or combination treatment. Oncology. 2011; 81:330-335.
49. Kubo T, Toyooka S, Tsukuda K, Sakaguchi M, Fukazawa T, Soh J, Asano H, Ueno T, Muraoka T, Yamamoto H, Nasu Y, Kishimoto T, Pass HI, Matsui H, Huh NH and Miyoshi S. Epigenetic silencing of microRNA-34b/c plays an important role in the pathogenesis of malignant pleural mesothelioma. Clinical cancer research. 2011; 17:4965-4974.
50. Donzelli S, Strano S and Blandino G. microRNAs: short non-coding bullets of gain of function mutant p53 proteins. Oncoscience. 2014; 1:427-433. doi: 10.18632/oncoscience.52.
51. Hattori H, Janky R, Nietfeld W, Aerts S, Madan Babu M and Venkitaraman AR. p53 shapes genome-wide and cell type-specific changes in microRNA expression during the human DNA damage response. Cell Cycle. 2014; 13:2572-2586.
52. Wang N, Xu ZW and Wang KH. Systematical analysis of cutaneous squamous cell carcinoma network of microRNAs, transcription factors, and target and host genes. Asian Pacific journal of cancer prevention. 2014; 15:10355-10361.
53. Bitanihirwe BK, Meerang M, Friess M, Soltermann A, Frischknecht L, Thies S, Felley-Bosco E, Tsao MS, Allo G, de Perrot M, Seifert B, Moch H, Stahel R, Weder W and Opitz I. PI3K/mTOR signaling in mesothelioma patients treated with induction chemotherapy followed by extrapleural pneumonectomy. Journal of thoracic oncology. 2014; 9:239-247.
54. Cedres S, Montero MA, Martinez P, Martinez A, Rodriguez-Freixinos V, Torrejon D, Gabaldon A, Salcedo M, Ramon YCS and Felip E. Exploratory analysis of activation of PTEN-PI3K pathway and downstream proteins in malignant pleural mesothelioma (MPM). Lung cancer. 2012; 77:192-198.
55. Solozobova V and Blattner C. Regulation of p53 in embryonic stem cells. Experimental cell research. 2010; 316:2434-2446.
56. Takagi M, Absalon MJ, McLure KG and Kastan MB. Regulation of p53 translation and induction after DNA damage by ribosomal protein L26 and nucleolin. Cell. 2005; 123:49-63.