
INTRODUCTION
Intracranial aneurysms (IAs) account for about 80–85% of non-traumatic subarachnoid hemorrhages (SAH) [1–3]. The prevalence of IAs is about 1–5% (10 million to 12 million persons in the United States) and the incidence is 1 per 10,000 persons per year in the United States (approximately 27,000), with incidence highest in 30- to 60-year-olds [4, 5]. Women own more cases of IAs, by a ratio of 3 to 2 [4]. Little is known about the molecular pathogenesis of IAs. They are thought to result from the interplay of environmental and genetic factors [6]. Previous studies have indicated that hypertension, hypercholesterolemia, cigarette smoking and female gender are risk factors for IAs [7]. Also, there is increasing evidence to suggest that genetic factors play an important part in the pathogenesis of IAs [6, 8, 9].
Many studies have revealed that copy number variations (CNVs) contributed to part of the missing heritability for complex traits after the GWAS era [10–20]. Human WW domain-containing oxidoreductase (WWOX) is a tumor suppressor that has been reported to lose function due to genetic alterations carcinogenesis [21, 22]. It is found in all eukaryotes and play an important role in the regulation of a wide variety of cellular functions such as protein degradation, transcription, and RNA splicing [23–25]. The human WWOX gene (OMIM: 605131) is located on chromosome 16q23.3–24.1, a region spanning over the common fragile site 16D (FRA16D). Somatic and germline mutations of WWOX, including loss of heterozygosis (LOH), homozygous deletions and chromosomal translocations, has been reported to evolve in carcinogenesis and development in several types of cancers [22]. Very interestingly, we previously identified that deletion genotypes of CNV-67048 in WWOX gene predispose their carriers to gliomas, which are the most common primary tumors of the central nervous system (CNS) and representing more than 80% of all malignant brain tumors [21]. Basing on the evidence above, we hypothesized that the CNVs in WWOX might be also associated with risk of IAs by disturbing the function of WWOX. To test this hypothesis, we performed a two-stage, case-control study among Chinese people to evaluate whether WWOX CNV-67048, a representative CNV in the WWOX gene region, contributes to the risk of IAs.
RESULTS
Totally included in this study were 976 cases of IAs and 1200 matched healthy controls. As shown in Table 1, all the demographic characteristics of patients with IAs and controls are summarized. There were no statistically significant differences between groups with respect to age, gender, smoking status, drinking status, and body mass index (BMI) in both discovery stage and validation stage (all P value > 0.05). However, the hypertension rates of patients with IAs were significantly higher than those of healthy controls in both discovery stage (P < 0.001) and validation stage (P < 0.001).
Table 1: Characteristics of IAs patients and healthy controls used in this study
Category |
Discovery stage |
Validation stage |
||||
|---|---|---|---|---|---|---|
Cases |
Controls |
P Value |
Cases |
Controls |
P Value |
|
Age (yr) |
|
|
|
|
|
|
Mean ± SD |
54.0 ± 6.7 |
53.8 ± 7.1 |
0.655 |
53.2 ± 4.1 |
53.7 ± 6.3 |
0.108 |
Gerder |
|
|
|
|
|
|
Male |
167 (41.8%) |
249 (41.5%) |
0.937 |
228 (39.5%) |
233 (38.8%) |
0.792 |
Female |
233 (58.2%) |
351 (58.5%) |
|
348 (60.5%) |
367 (61.2%) |
|
Hypertension |
|
|
|
|
|
|
Yes |
240 (60.0%) |
127 (21.2%) |
P < 0.001 |
337 (58.5%) |
120 (20.0%) |
P < 0.001 |
No |
160 (40.0%) |
473 (78.8%) |
|
239 (41.5%) |
480 (80.0%) |
|
Ever smoker |
|
|
|
|
|
|
Yes |
74 (18.5%) |
86 (14.3%) |
0.078 |
108 (18.8%) |
88 (14.7%) |
0.060 |
No |
326 (81.5%) |
514 (85.7%) |
|
468 (81.2%) |
512 (85.3%) |
|
Ever drinker |
|
|
|
|
|
|
Yes |
89 (22.3%) |
121 (20.1%) |
0.428 |
138 (23.9%) |
123 (20.5%) |
0.154 |
No |
311 (77.7%) |
479 (79.9%) |
|
438 (76.1%) |
477 (79.5%) |
|
Body mass index (kg/m2) |
24.2 ± 3.6 |
23.9 ± 3.1 |
0.160 |
24.3 ± 4.1 |
23.9 ± 3.2 |
0.061 |
Table 2 presented the genotype distribution of WWOX CNV-67048 among both patients with IAs and healthy controls in the discovery stage. Consistent with results in DGV databases, we detected three kinds of genotypes for WWOX CNV-67048 (no deletion, one copy deletion, and two copy deletion) in all samples. When analyzing the data using a log-additive model and adjusted for age, gender, smoking status, and Hypertension, we found that there was a significantly higher risk of IAs for per copy deletion (OR = 1.43, 95% CI = 1.14–1.80; Ptrend = 1.79 × 10−3). Compared with the individuals having no deletion, those carrying 1 copy deletion harbored a 1.37-fold increased risk of lung cancer (OR = 1.37, 95% CI = 1.02–1.83), while those with 2 copy deletion had a much higher risk (OR = 1.88, 95% CI = 1.10–3.21).
Table 2: WWOX gene deletion and risk of IAs in discovery stage
Genotypes |
No. of cases |
No. of controls |
OR (95% CI)a |
|---|---|---|---|
no deletion |
255 (63.8%) |
431 (71.8%) |
1.00 (reference) |
1 copy deletion |
115 (28.8%) |
142 (23.7%) |
1.37 (1.02–1.83) |
2 copy deletion |
30 (7.4%) |
27 (4.5%) |
1.88 (1.10–3.21) |
per copy deletion |
|
|
1.43 (1.14–1.80) |
Ptrend |
|
|
1.79 × 10−3 |
aAdjusted for age, gender, smoking status, and Hypertension.
Furthermore, the results were validated in an independent validation dataset (Table 3). A 1.29-fold increased risk of IAs were detected for per copy deletion (OR = 1.29, 95% CI = 1.05-1.58; P trend = 0.017). When pooled together, significantly higher risk of IAs was detected for both subjects carrying 1 copy deletion (OR = 1.24, 95% CI = 1.02–1.52) and subjects carrying 2 copy deletion (OR = 1.77, 95% CI = 1.24–2.53), compared with the individuals having no deletion. The effect size for per copy deletion was 1.35 (95% CI = 1.16–1.57; P trend = 1.18 × 10−4). Multiplicative interactions between CNV and demographic variables were evaluated using the likelihood ratio test when interaction terms were added to logistic regression models along with the main effect terms. However, we did not find any significant interaction terms on risk of IAs (data not shown).
Table 3: WWOX gene deletion and risk of IAs in validation stage and the pooled results
Genotypes |
No. of cases |
No. of controls |
OR (95% CI)a |
|---|---|---|---|
Validation stage |
|
|
|
no deletion |
384 (66.7%) |
429 (71.5%) |
1.00 (reference) |
1 copy deletion |
147 (25.5%) |
141 (23.5%) |
1.16 (0.89–1.52) |
2 copy deletion |
45 (7.8%) |
30 (5.0%) |
1.67 (1.04–2.70) |
per copy deletion |
|
|
1.29 (1.05–1.58) |
Ptrend |
|
|
0.017 |
Pooled results |
|
|
|
no deletion |
639 (65.5%) |
860 (71.7%) |
1.00 (reference) |
1 copy deletion |
262 (26.8%) |
283 (23.6%) |
1.24 (1.02–1.52) |
2 copy deletion |
75 (7.7%) |
57 (4.7%) |
1.77 (1.24–2.53) |
per copy deletion |
|
|
1.35 (1.16–1.57) |
P trend |
|
|
1.18 × 10−4 |
aAdjusted for age, gender, smoking status, and Hypertension.
To confirm the abnormal expression of WWOX in tissues of IA patients and influence of genotypes of CNV-67048, we evaluated the WWOX levels in 70 paired tissues of IA patients and corresponding normal tissues. As shown in Figure 1, the expression level of WWOX in IA tissues was significantly lower than that in corresponding normal tissues (P = 0.004). The deletion genotypes of CNV-67048 have lower WWOX mRNA levels in both tumor tissues (P = 0.001) and border tissues (P = 0.002). Then, to determine whether WWOX protein expression was altered in IAs, we also performed Western blot analysis of lysates from 10 paired IA tissues and adjacent normal tissues. As shown in Figure 2, densitometry measurements from Western blots revealed that all of the IA tissues had an average of 48% reduction of WWOX levels compared to adjacent normal tissues when normalized to actin.
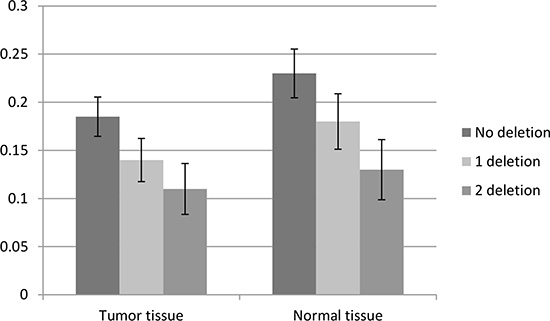
Figure 1: Association between the CNV-67048 and WWOX expression. The relative mRNA levels of WWOX in IA tissues compared with border normal tissues with different genotypes.
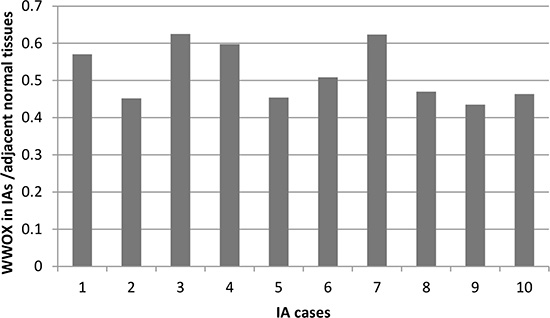
Figure 2: Densitometry measurements from Western blots of WWOX in paired IA tissues and adjacent normal tissues (WWOX in IAs tissues/adjacent normal tissues).
DISCUSSION
Fully exploration of CNVs and their role in carcinogenesis could possess a large number of potential clues for developing novel therapeutic agents for IAs. In the current two-stage, case–control study, we found that the CNV-67048 in WWOX was significantly associated with increased risk of IAs. The deletion genotypes of CNV-67048 were related with a higher rate of IAs and lower expression of WWOX than the no deletion genotype in IA tissues. To the best of our knowledge, this is the first study conducted to reveal a functional CNV in WWOX predisposing development of IAs. This provides evidence to implicate WWOX gene deletion as a novel susceptibility factor for IAs.
Rupture of IAs causes subarachnoid hemorrhage (SAH), a serious subtype of stroke, which leads to fatality in more than 50% of the cases and results in significant disability in 30% of the cases [26, 27]. Screening the susceptibility loci is an important work to effectively prevent the hazards of IAs. Although 9 genome-wide association studies (GWAS) have been conducted since 2008, only one locus (9p21.3) was consistently replicated across different studies, which means the majority of genetic factors for IAs needs further exploration [6]. Furthermore, the number of reports examining susceptibility loci for IA among Asian populations is particularly limited.
In 2000, Bednarek et al. [28] first identified that WWOX, a novel WW domain-containing protein mapping to human chromosome 16q23.3-24.1, as a region frequently affected in breast cancer. The ORF of WWOX is 1245 bp long, encoding a 414-amino acid protein with nine exons [29]. The distinct methylation patterns of WWOX could also influence the disease status of lung squamous cell carcinomas, invasive breast carcinomas, normal mammary tissues, and bladder transitional cell carcinomas [30]. Nowakowska et al. [31] found that WWOX expression were related with cell cycle and apoptosis regulation in neuroblastoma. In the current study, we showed that WWOX expression was lower in IA tissues compared with their adjacent normal tissues. Germline CNVs is a potential induction factor of somatic genetic alterations, which may be involved in the carcinogenesis process and development [32–35]. WWOX CNV-67048, which maps to chr16:76,929,120-76,942,453, has been linked to risk of lung cancer, gliomas, and COPD recently [21, 22, 36]. In our study, we first find that the WWOX gene deletion (CNV-67048) was associated with increased risk of IAs. As known, CNVs can directly influence gene expression and phenotypic variation, disrupt gene structure, alter gene dosage and indirectly regulate gene function through position effects. In this study, the deletion genotypes of CNV-67048 have lower WWOX mRNA levels in both tumor tissues and their borderline normal tissues, compared with the no deletion genotype. Findings of our study fit with current research about the suppressor gene function of WWOX.
The pleotropic roles of WWOX was not only restricted to tumor suppression, but also expanded to neuropathy as well as metabolic diseases. Dayan et al. [37] reported that altering metabolism from glycolysis to oxidative phosphorylation causes stable increase in steady-state levels of transcripts of the WWOX gene. Additionally, exposure to hypoxic conditions could cause a down-regulation of WWOX mRNA. Due to WWOX often exhibits homozygous deletions and translocation breakpoints under multiple cellular stresses induced by extrinsic or intrinsic factors, such as hypoxia, UV, and DNA damage regents, evidence has shown that WWOX that contains a short-chain dehydrogenase/reductase (SDR) domain is involved in steroid metabolism and bone development, while reduced or lost expression of WWOX will lead to development of metabolic disease [38]. Aldaz et al. [39] also reviewed that WWOX acted at the crossroads of cancer, metabolic syndrome related traits and CNS pathologies. Functionally, the WW domain is not only a tumor suppressor, but also a participant in molecular interactions, signaling, and apoptosis in many diseases. Studies also revealed the potential mechanism by which WWOX/WOX1 may participate in the pathogenesis of AD with a focus on cell death signaling pathways in neurons [40].
Strengths of current study includes: (1) our large sample size and a two-stage, case– control study design; (2) functional analyses further supported the findings that the CNV has significant association with IA risk. However, some limitations should also be considered: (1) although the sample size was large enough for the main analyses, the statistical power for the interaction analyses was still limited; (2) the potential bias for case-control study; (3) the extrapolation of our results to other ethnic groups.
In conclusion, this study found that the deletion genotypes of CNV-67048 in WWOX was significantly associated with an increased risk of IAs among Chinese population, and reduced the expression of WWOX in vivo. The results suggest that the CNV-67048 in WWOX gene may be a new biomarker for susceptibility of IAs. Validations with larger population-based studies in different ethnic groups and further research into the function of WWOX deletion may be warranted.
MATERIALS AND METHODS
Subjects
Subjects included in this study were geographically homogenous Han Chinese derived from Chengdu Military General Hospital, Jinling Hospital of Nanjing University, and Fuzhou General Hospital of Nanjing Command before June 2015. Ruptured IAs were diagnosed according to the suggestive symptoms of SAH, subarachnoid blood on CT, or a proven aneurysm at angiography (angiogram, CT, or MRA/or MRI) or at surgery; un-ruptured IAs were ascertained using CT, MRA (or MRI), angiography, surgery, or autopsy. All suspected IAs were confirmed by digital subtraction angiography (DSA). Health controls were matched for area of residence, age and gender to eliminate the effect of population stratification by heterogeneity. A structured questionnaire was used to elicit detailed information on demographic factors. Then 5 ml blood samples were obtained from the subjects who participated in the study. Finally, included in this study were 400 subjects diagnosed with IAs, and 600 healthy controls in the discovery stage, as well as 576 IAs patients and 600 healthy controls in an independent validation stage. The study complied with the Declaration of Helsinki and was approved by the appropriate Institutional Review Board and Ethics Committee, and all participants provided written informed consent.
CNV selection and genotyping
The procedure of CNV selection and genotype has been described previously [21]. Briefly, through searching the DGV database, we found only two deletion of copy number CNVs (CNV-38092 and CNV-67048) to exist in Chinese populations with the minor allele frequency (MAF) > 0.05. As reported previously [21], 98% of individuals shared the deletion of CNV-38092 and CNV-67048, which means they could be used as the proxy of each other. In current study, we finally selected CNV-67048 as the representative CNV. Real-time qualitative PCR (qPCR) were used to conduct copy number analyses, and all CNV calls were conducted by two independent staff members.
WWOX expression detection and western blot
The mRNA levels of WWOX were detected by SYBR-Green real-time PCR in IAs tissues and border of the IAs tissue of 70 cases. The primers were designed as previous [21, 22]: 5′-TGG GTT TAC TAC GCC AAT C-3′(forward) and 5′- GTC CGTTCT CAT CAG TTT CT -3′(reverse) to amplify 124 bp cDNA sequences from exon 2 to exon 3 of WWOX, while the b-actin was used as an internal reference gene. Western blot analyses were performed using protein extracts obtained from 10 paired IA tissues and adjacent normal tissues. 50 μg of total protein was separated by 12.5% SDS–PAGE and transferred onto PVDF membranes (Millipore, Billerica, MA, USA). Immunodetection was performed using Protein Detector™ (KPL, Gaithersburg, MD, USA) Western blotting reagents as described by the manufacturer. Actin was detected using monoclonal anti-actin antibody (1:1000) and HRP conjugated anti-mouse secondary antibody (1:5000). Quantitation of western blot autoradiographs was done using a Kodak digital science Image Station 440CF.
Statistical analyses
Cases and controls were compared with the Chi-square or T test for categorical or continuous variables, respectively. Unconditional logistic regression models were used to evaluated the associations between the CNV-67048 and risk of IAs using odds ratios (ORs) and 95% confidence intervals (CIs). ORs were estimated for 1 copy and 2 copy deletion genotypes compared with no deletion genotype. The OR (95% CI) was also estimated for per copy deletion based on a log-additive model and adjusted for age, gender, smoking status, and Hypertension. One-way ANOVA tests were used for analyzing the association between CNV-67048 genotypes and WWOX expression. All statistical analyses were conducted with SAS version 9.2 (SAS Institute Inc.). All statistical tests were 2-tailed, and P < 0.05 was interpreted as statistically significant.
ACKNOWLEDGMENTS AND FUNDING
The study was supported by the Twelfth five-year Plan for Medical Projects of Chengdu Military Command (grant No. C12045) and the Key Medical Project of Chengdu Military General Hospital (2013YG-A006).
CONFLICTS OF INTEREST
The authors declare that they have no conflicts of interest.
REFERENCES
1. Kassell NF, Torner JC, Jane JA, Haley EC, Jr., Adams HP. The International Cooperative Study on the Timing of Aneurysm Surgery. Part 2: Surgical results. Journal of neurosurgery. 1990; 73:37–47.
2. Sawyer DM, Amenta PS, Medel R, Dumont AS. Inflammatory mediators in vascular disease: identifying promising targets for intracranial aneurysm research. Mediators of inflammation. 2015; 2015:896283.
3. Tromp G, Weinsheimer S, Ronkainen A, Kuivaniemi H. Molecular basis and genetic predisposition to intracranial aneurysm. Ann Med. 2014; 46:597–606.
4. Brisman JL, Song JK, Newell DW. Cerebral aneurysms. N Engl J Med. 2006; 355:928–939.
5. Brown RD, Jr., Broderick JP. Unruptured intracranial aneurysms: epidemiology, natural history, management options, and familial screening. Lancet Neurol. 2014; 13:393–404.
6. Abrantes P, Santos MM, Sousa I, Xavier JM, Francisco V, Krug T, Sobral J, Matos M, Martins M, Jacinto A, Coiteiro D, Oliveira SA. Genetic Variants Underlying Risk of Intracranial Aneurysms: Insights from a GWAS in Portugal. PLoS One. 2015; 10:e0133422.
7. Low SK, Takahashi A, Cha PC, Zembutsu H, Kamatani N, Kubo M, Nakamura Y. Genome-wide association study for intracranial aneurysm in the Japanese population identifies three candidate susceptible loci and a functional genetic variant at EDNRA. Hum Mol Genet. 2012; 21:2102–2110.
8. Kremer PH, Koeleman BP, Pawlikowska L, Weinsheimer S, Bendjilali N, Sidney S, Zaroff JG, Rinkel GJ, van den Berg LH, Ruigrok YM, de Kort GA, Veldink JH, Kim H, et al. Evaluation of genetic risk loci for intracranial aneurysms in sporadic arteriovenous malformations of the brain. J Neurol Neurosurg Psychiatry. 2015; 86:524–529.
9. Chen J, Zhu Y, Jiang Y, Yu H, Sun K, Song W, Luan L, Lou K, Li Y, Jiang P, Pang Q, Hui R. A functional variant of the collagen type III alpha1 gene modify risk of sporadic intracranial aneurysms. Hum Genet. 2012; 131:1137–1143.
10. Deregowska A, Skoneczny M, Adamczyk J, Kwiatkowska A, Rawska E, Skoneczna A, Lewinska A, Wnuk M. Genome-wide array-CGH analysis reveals YRF1 gene copy number variation that modulates genetic stability in distillery yeasts. Oncotarget. 2015.
11. Hugen N, Simmer F, Mekenkamp LJ, Koopman M, van den Broek E, de Wilt JH, Punt CJ, Ylstra B, Meijer GA, Nagtegaal ID. Reduced rate of copy number aberrations in mucinous colorectal carcinoma. Oncotarget. 2015; 6:25715–25725.
12. Li Z, Yu D, Gan M, Shan Q, Yin X, Tang S, Zhang S, Shi Y, Zhu Y, Lai M, Zhang D. A genome-wide assessment of rare copy number variants in colorectal cancer. Oncotarget. 2015; 6:26411–26423.
13. Lu TX, Fan L, Wang L, Wu JZ, Miao KR, Liang JH, Gong QX, Wang Z, Young KH, Xu W, Zhang ZH, Li JY. MYC or BCL2 copy number aberration is a strong predictor of outcome in patients with diffuse large B-cell lymphoma. Oncotarget. 2015; 6:18374–18388.
14. Czubak K, Lewandowska MA, Klonowska K, Roszkowski K, Kowalewski J, Figlerowicz M, Kozlowski P. High copy number variation of cancer-related microRNA genes and frequent amplification of DICER1 and DROSHA in lung cancer. Oncotarget. 2015; 6:23399–23416.
15. Peng CH, Liao CT, Ng KP, Tai AS, Peng SC, Yeh JP, Chen SJ, Tsao KC, Yen TC, Hsieh WP. Somatic copy number alterations detected by ultra-deep targeted sequencing predict prognosis in oral cavity squamous cell carcinoma. Oncotarget. 2015; 6:19891–19906.
16. Casadevall D, Gimeno J, Clave S, Taus A, Pijuan L, Arumi M, Lorenzo M, Menendez S, Canadas I, Albanell J, Serrano S, Espinet B, Salido M, et al. MET expression and copy number heterogeneity in nonsquamous non-small cell lung cancer (nsNSCLC). Oncotarget. 2015; 6:16215–16226.
17. Jin DH, Lee J, Kim KM, Kim S, Kim DH, Park J. Overexpression of MAPK15 in gastric cancer is associated with copy number gain and contributes to the stability of c-Jun. Oncotarget. 2015; 6:20190–20203.
18. Kubler K, Heinenberg S, Rudlowski C, Keyver-Paik MD, Abramian A, Merkelbach-Bruse S, Buttner R, Kuhn W, Schildhaus HU. c-myc copy number gain is a powerful prognosticator of disease outcome in cervical dysplasia. Oncotarget. 2015; 6:825–835.
19. Liu B, Morrison CD, Johnson CS, Trump DL, Qin M, Conroy JC, Wang J, Liu S. Computational methods for detecting copy number variations in cancer genome using next generation sequencing: principles and challenges. Oncotarget. 2013; 4:1868–1881.
20. Gualtieri A, Andreola F, Sciamanna I, Sinibaldi-Vallebona P, Serafino A, Spadafora C. Increased expression and copy number amplification of LINE-1 and SINE B1 retrotransposable elements in murine mammary carcinoma progression. Oncotarget. 2013; 4:1882–1893.
21. Yu K, Fan J, Ding X, Li C, Wang J, Xiang Y, Wang QS. Association study of a functional copy number variation in the WWOX gene with risk of gliomas among Chinese people. Int J Cancer. 2014; 135:1687–1691.
22. Yang L, Liu B, Huang B, Deng J, Li H, Yu B, Qiu F, Cheng M, Wang H, Yang R, Yang X, Zhou Y, Lu J. A functional copy number variation in the WWOX gene is associated with lung cancer risk in Chinese. Hum Mol Genet. 2013; 22:1886–1894.
23. Baryla I, Styczen-Binkowska E, Bednarek AK. Alteration of WWOX in human cancer: a clinical view. Exp Biol Med (Maywood). 2015; 240:305–314.
24. Gao G, Smith DI. WWOX, large common fragile site genes, and cancer. Exp Biol Med (Maywood). 2015; 240:285–295.
25. Ramos D, Aldaz CM. WWOX, a chromosomal fragile site gene and its role in cancer. Adv Exp Med Biol. 2006; 587:14–159.
26. Clark CE, 3rd, Jupiter DC, Thomas JS, Papaconstantinou HT. Rectal prolapse in the elderly: trends in surgical management and outcomes from the American College of Surgeons National Surgical Quality Improvement Program database. J Am Coll Surg. 2012; 215:709–714.
27. Inagawa T. Trends in surgical and management outcomes in patients with aneurysmal subarachnoid hemorrhage in Izumo city, Japan, between 1980–1989 and 1990–1998. Cerebrovasc Dis. 2005; 19:39–48.
28. Bednarek AK, Laflin KJ, Daniel RL, Liao Q, Hawkins KA, Aldaz CM. WWOX, a novel WW domain-containing protein mapping to human chromosome 16q23.3-24.1, a region frequently affected in breast cancer. Cancer Res. 2000; 60:2140–2145.
29. Ludes-Meyers JH, Bednarek AK, Popescu NC, Bedford M, Aldaz CM. WWOX, the common chromosomal fragile site, FRA16D, cancer gene. Cytogenet Genome Res. 2003; 100:101–110.
30. Iliopoulos D, Guler G, Han SY, Johnston D, Druck T, McCorkell KA, Palazzo J, McCue PA, Baffa R, Huebner K. Fragile genes as biomarkers: epigenetic control of WWOX and FHIT in lung, breast and bladder cancer. Oncogene. 2005; 24:1625–1633.
31. Nowakowska M, Pluciennik E, Wujcicka WI, Sitkiewicz A, Kazanowska B, Zielinska E, Bednarek AK. The correlation analysis of WWOX expression and cancer related genes in neuroblastoma- a real time RT-PCR study. Acta Biochim Pol. 2014; 61:91–97.
32. Nik-Zainal S, Wedge DC, Alexandrov LB, Petljak M, Butler AP, Bolli N, Davies HR, Knappskog S, Martin S, Papaemmanuil E, Ramakrishna M, Shlien A, Simonic I, et al. Association of a germline copy number polymorphism of APOBEC3A and APOBEC3B with burden of putative APOBEC-dependent mutations in breast cancer. Nat Genet. 2014; 46:487–491.
33. Krepischi AC, Pearson PL, Rosenberg C. Germline copy number variations and cancer predisposition. Future Oncol. 2012; 8:441–450.
34. Kuiper RP, Ligtenberg MJ, Hoogerbrugge N, Geurts van Kessel A. Germline copy number variation and cancer risk. Curr Opin Genet Dev. 2010; 20:282–289.
35. Sasso EH, Johnson T, Kipps TJ. Expression of the immunoglobulin VH gene 51p1 is proportional to its germline gene copy number. J Clin Invest. 1996; 97:2074–2080.
36. Yang L, Qiu F, Fang W, Zhang L, Xie C, Lu X, Huang D, Guo Y, Pan M, Zhang H, Zhou Y, Lu J. The Functional Copy Number Variation-67048 in WWOX Contributes to Increased Risk of COPD in Southern and Eastern Chinese. Copd. 2015:1–8.
37. Dayan S, O’Keefe LV, Choo A, Richards RI. Common chromosomal fragile site FRA16D tumor suppressor WWOX gene expression and metabolic reprograming in cells. Genes Chromosomes Cancer. 2013; 52:823–831.
38. Li J, Liu J, Ren Y, Yang J, Liu P. Common Chromosomal Fragile Site Gene WWOX in Metabolic Disorders and Tumors. International journal of biological sciences. 2014; 10:142–148.
39. Aldaz CM, Ferguson BW, Abba MC. WWOX at the crossroads of cancer, metabolic syndrome related traits and CNS pathologies. Biochim Biophys Acta. 2014; 1846:188–200.
40. Teng CC, Yang YT, Chen YC, Kuo YM, Sze CI. Role of WWOX/WOX1 in Alzheimer’s disease pathology and in cell death signaling. Front Biosci (Elite Ed). 2012; 4:1951–1965.