
INTRODUCTION
Gastric cancer (GC) is one of the most common cancers and is the second leading cause of cancer-related death worldwide [1]. Disease development progresses stepwise from a normal stomach through inflammation and precancerous conditions to cancer, as described by Correa’s cascade [2]. Although environmental factors such as Helicobacter pylori infection are known risk factors for GC, genetic influences and interactions with environmental factors also play an essential role in its initiation [3]. Therefore, the screening and identification of genetic factors that are associated with risks of GC and its precancerous diseases would reveal the etiology and pathogenesis.
As the most common form of genetic variation, single nucleotide polymorphisms (SNP) have been widely investigated in relation to the risk of cancers. Genome-wide association studies (GWAS) have identified several SNPs that are significantly associated with high GC risk including: MUC1 rs2070803 G/A and PSCA rs2976392 A/G, associated with an increased risk of diffuse-type GC in a Japanese population (odds ratio (OR)=1.63, P =1.2 × 10−6; OR=1.62, P =1.1 × 10−9) [4]; PLCE1 rs2274223 A/G, associated with a high GC risk in a Chinese population (OR=1.31, P =8.4 × 10−9) [5]; and PRKAA1 rs13361707 T/C, which was associated with an increased risk of non-cardia GC (OR=1.41, P =7.6 × 10−29) [6]. A number of candidate gene association studies have also identified SNPs in genes encoding pepsinogen C and glutathione S-transferase pi 1, which appear to significantly alter individual susceptibility to GC [7, 8]. Although these studies have found several SNPs related to GC risk, most focused on scattered SNPs rather than integral gene-gene pathways or gene-environment interactions. Thus, the screening of additional key SNPs is still required to elucidate their role in different stages of gastric carcinogenesis.
Nucleotide excision repair (NER) is a versatile system that monitors and repairs DNA damage, including ultraviolet (UV)-induced cyclobutane pyrimidine dimers, DNA crosslinks, and bulky adducts [9]. NER stages include damage recognition, damage demarcation and unwinding, damage incision, and new strand ligation [10], all of which require corresponding functional proteins. Cellular DNA is constantly at risk from damage by endogenous and exogenous stimuli, and NER defects are likely to increase genome instability [11]. Polymorphisms of NER genes might change the NER ability by influencing the expression and function of key proteins, thereby altering individual susceptibility to GC and giving rise to gastric carcinogenesis[12, 13].
Polymorphisms of several key NER genes have previously been reported to alter the GC risk, including xeroderma pigmentosum, complementation group A (XPA) and xeroderma pigmentosum, complementation group C (XPC) in the damage recognition step, excision repair cross-complementation group 2 (ERCC2) in the damage unwinding step, and ERCC1, ERCC4, and ERCC5 in the damage incision step [13]. However, most of these studies investigated only a few SNPs of a single gene. For instance, Chen et al. reported three ERCC2 SNPs [14], while He et al. studied three ERCC5 SNPs [12]. No study has yet analyzed the role of SNPs from the entire NER pathway in gastric carcinogenesis. In the present study, therefore, we systematically analyzed 39 SNPs of eight key NER genes (ERCC1, ERCC2, ERCC3, ERCC4, ERCC5, XPA, XPC, and the damage-specific DNA binding protein 2 gene DDB2) in a total of 2686 northern Chinese subjects including 898 with GC, 851 with atrophic gastritis (AG), and 937 controls. The roles of these SNPs at different stages of gastric carcinogenesis as well as gene-gene and gene-environment interactions were investigated to determine whether they could be used to predict GC risk.
RESULTS
Characteristics of subjects
A total of 2686 subjects were selected for inclusion in the study, and their characteristics are shown in Supplementary Table S2. GC and AG groups had a significantly higher proportion of males (71.2 and 56.2%, respectively) compared with the control (CON) group (54.4%; P < 0.001). H. pylori infection rates (50.6 and 50.9%, respectively) were also significantly higher in GC and AG groups than the CON group (29.2%; P < 0.001). Based on Lauren’s histological classification of GC, 269 cases were intestinal-type (37.3%) and 453 cases were diffuse-type (62.7%).
NER pathway gene SNPs and disease risk
Our study focused on 39 SNPs in eight NER pathway genes (ERCC1, ERCC2, ERCC3, ERCC4, ERCC5, XPA, XPC, and DDB2). Six SNPs were not in accordance with Hardy-Weinberg equilibrium (HWE) so were not entered into the subsequent association study. Primary details of SNPs and allele frequencies are listed in Supplementary Table S1 and the associations of genotypes with disease risk by different genetic models are summarized in Supplementary Table S3. Subgroup analysis results for H. pylori infection positive/negative and intestinal/diffuse-type GC are shown in Supplementary Tables S4 and Table S5, respectively.
XPA SNPs rs10817938, rs2808668 and DDB2 SNP rs830083 were found to be significantly associated with the risk of AG, while XPC SNP rs2607775 and DDB2 SNPs rs2029298, rs326222, rs3781619, and rs830083 were significantly associated with GC risk (Table 1). After Bonferroni correction for multiple comparisons, DDB2 rs830083 and XPC rs2607775 remained significantly associated with increased GC risk: the DDB2 rs830083 GG genotype was significantly associated with an increased risk of GC compared with the wild-type CC genotype (OR=2.32, 95% confidence interval (CI)=1.75-3.08, P =6.62 × 10−9), and the XPC rs2607775 CG genotype conferred a 1.73-fold increased GC risk compared with the wild-type CC genotype (OR=1.73, 95%CI=1.28-2.32, P =3.04 × 10−4).
Table 1: NER SNPs that demonstrate significant association with disease risk
The combined analysis of the DDB2 rs830083 and XPC rs2607775 risk genotypes demonstrated an even higher GC risk, with individuals carrying both risk genotypes having a 3.05-fold increased risk of developing GC (Figure 1).
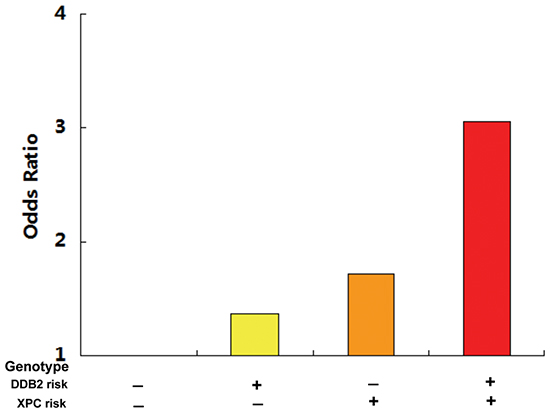
Figure 1: The combined detection of the DDB2 rs830083 and XPC rs2607775 risk genotypes demonstrated an even higher GC risk, with individuals carrying both risk genotypes having a 3.05-fold increased risk of developing GC.
Stratified analysis for DDB2 rs830083 and XPC rs2607775
We next performed stratified analyses of DDB2 rs830083 and XPC rs2607775 by gender, presence of H. pylori infection, Lauren’s classification of GC, and smoking and alcohol consumption status of individuals. As shown in Table 2, the association of the DDB2 rs830083 GG genotype with GC risk was more obvious in subgroups of males (OR=2.61, P =4.26 × 10−7), non-smokers (OR=2.39, P =3.93 × 10−4), and non-drinkers (OR=3.10, P =3.41 × 10−6) than the controls. Individuals both positive and negative for H. pylori infection, and with either intestinal-type or diffuse-type all demonstrated an increased risk of GC if they also possessed the GG genotype. For XPC rs2607775, the CG genotype was significantly associated with an increased risk of GC in males (OR=2.00, P =4.03 × 10−4) and H. pylori-negative (OR=1.98, P =8.93 × 10−4) subgroups.
Table 2: Subgroup analysis of DDB2 rs830083 and XPC rs2607775 polymorphisms
CON→AG |
AG→GC |
CON→GC |
Non-cancer→GC |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
Group |
OR(95%CI) |
P |
PB-D test |
OR(95%CI) |
P |
PB-D test |
OR(95%CI) |
P |
PB-D test |
OR(95%CI) |
P |
PB-D test |
DDB2 rs830083 GG vs. CC |
||||||||||||
All |
1.48(1.11-1.97) |
0.008 |
1.63(1.25-2.13) |
3.29 × 10−4 |
2.32(1.75-3.08) |
6.62 × 10−9 |
1.94(1.54-2.44) |
2.18 × 10−8 |
||||
Gender |
0.960 |
0.605 |
0.646 |
0.589 |
||||||||
Male |
1.67(1.11-2.50) |
0.013 |
1.75(1.24-2.46) |
1.44 × 10−3 |
2.61(1.80-3.78) |
4.26 × 10−7 |
2.10(1.57-2.82) |
6.66 × 10−7 |
||||
Female |
1.35(0.89-2.04) |
0.157 |
1.45(0.95-2.23) |
0.086 |
1.97(1.26-3.08) |
0.003 |
1.68(1.15-2.47) |
0.008 |
||||
Hp infection |
0.673 |
0.190 |
0.418 |
0.432 |
||||||||
Positive |
1.37(0.88-2.11) |
0.162 |
1.91(1.34-2.72) |
3.53 × 10−4 |
2.70(1.71-4.26) |
1.89 × 10−5 |
2.14(1.54-2.97) |
5.75 × 10−6 |
||||
Negative |
1.59(1.08-2.32) |
0.018 |
1.32(0.88-1.98) |
0.181 |
2.09(1.45-3.02) |
7.49 × 10−5 |
1.75(1.26-2.43) |
8.01 × 10−4 |
||||
Lauren’s classification |
0.914 |
0.915 |
0.907 |
|||||||||
Intestinal |
1.48(1.11-1.97) |
0.008 |
1.82(1.23-2.69) |
0.003 |
2.75(1.82-4.17) |
1.79 × 10−6 |
2.18(1.51-3.14) |
2.91 × 10−5 |
||||
Diffuse |
1.48(1.11-1.97) |
0.008 |
1.70(1.24-2.34) |
1.02 × 10−3 |
2.40(1.71-3.36) |
3.45 × 10−7 |
2.04(1.52-2.72) |
1.6 × 10−6 |
||||
Smoking |
0.013 |
<0.001 |
0.271 |
0.011 |
||||||||
Smoker |
2.56(1.30-5.07) |
0.007 |
0.91(0.48-1.73) |
0.770 |
2.01(1.00-4.04) |
0.050 |
1.30(0.75-2.28) |
0.352 |
||||
Nonsmoker |
0.67(0.42-1.07) |
0.094 |
3.17(1.96-5.13) |
2.63 × 10−6 |
2.39(1.48-3.88) |
3.93 × 10−4 |
2.79(1.84-4.23) |
1.24 × 10−6 |
||||
Drinking |
0.861 |
0.094 |
0.111 |
0.061 |
||||||||
Drinker |
1.59(0.71-3.53) |
0.258 |
1.27(0.60-2.68) |
0.535 |
1.51(0.71-3.18) |
0.282 |
1.41(0.75-2.65) |
0.283 |
||||
Nondrinker |
0.94(0.60-1.45) |
0.762 |
2.99(1.88-4.76) |
4.10 × 10−6 |
3.10(1.92-5.00) |
3.41 × 10−6 |
2.97(1.98-4.48) |
1.77 × 10−7 |
||||
XPC rs2607775 CG vs. CC |
||||||||||||
All |
0.76(0.51-1.12) |
0.166 |
1.88(1.30-2.72) |
7.35 × 10−4 |
1.52(1.07-2.15) |
0.019 |
1.73(1.28-2.32) |
3.04 × 10−4 |
||||
Gender |
0.812 |
0.259 |
0.336 |
0.220 |
||||||||
Male |
0.79(0.44-1.43) |
0.436 |
2.19(1.33-3.61) |
0.002 |
1.84(1.15-2.92) |
0.010 |
2.00(1.36-2.94) |
4.03 × 10−4 |
||||
Female |
0.74(0.43-1.25) |
0.259 |
1.51(0.86-2.65) |
0.150 |
1.14(0.66-1.98) |
0.644 |
1.34(0.83-2.18) |
0.236 |
||||
Hp infection |
0.109 |
0.595 |
0.016 |
0.121 |
||||||||
Hp(+) |
0.58(0.33-1.00) |
0.051 |
1.79(1.08-2.96) |
0.025 |
0.99(0.58-1.70) |
0.975 |
1.44(0.93-2.22) |
0.105 |
||||
Hp(-) |
1.01(0.59-1.73) |
0.968 |
1.95(1.14-3.33) |
0.015 |
2.01(1.28-3.16) |
0.002 |
1.98(1.32-2.97) |
8.93 × 10−4 |
||||
Lauren’s classification |
0.747 |
0.733 |
0.714 |
|||||||||
Intestinal |
0.76(0.51-1.12) |
0.166 |
2.07(1.24-3.48) |
0.006 |
1.52(0.91-2.54) |
0.108 |
1.85(1.17-2.92) |
0.009 |
||||
Diffuse |
0.76(0.51-1.12) |
0.166 |
1.71(1.11-2.62) |
0.015 |
1.33(0.87-2.02) |
0.185 |
1.55(1.07-2.25) |
0.021 |
||||
Smoking |
0.876 |
0.099 |
0.041 |
0.027 |
||||||||
Smoker |
1.01(0.33-3.09) |
0.981 |
2.91(1.09-7.75) |
0.033 |
3.02(1.22-7.43) |
0.016 |
2.92(1.38-6.17) |
0.005 |
||||
Nonsmoker |
0.71(0.40-1.27) |
0.245 |
1.34(0.70-2.56) |
0.383 |
0.91(0.48-1.71) |
0.766 |
1.17(0.67-2.04) |
0.573 |
||||
Drinking |
0.420 |
0.247 |
0.665 |
0.397 |
||||||||
Drinker |
0.49(0.14-1.71) |
0.263 |
3.23(0.99-10.56) |
0.053 |
1.58(0.63-3.98) |
0.336 |
1.98(0.87-4.52) |
0.103 |
||||
Nondrinker |
0.83(0.47-1.47) |
0.529 |
1.51(0.81-2.82) |
0.194 |
1.25(0.68-2.33) |
0.473 |
1.46(0.85-2.49) |
0.172 |
||||
The association between each SNP and risk of AG and GC was estimated by calculating ORs and their 95% CIs by multivariate logistic regression with adjustments for gender, age, and H. pylori infection status.
Abbreviations: GC, gastric cancer; AG, atrophic gastritis; CON, control; Hp, H. pylori; B-D test, Breslow-Day test.
The association between each SNP and risk of AG and GC was estimated by calculating ORs and their 95% CIs by multivariate logistic regression with adjustments for gender, age, and H. pylori infection status.
Abbreviations: GC, gastric cancer; AG, atrophic gastritis; CON, control; Hp, H. pylori; B-D test, Breslow-Day test.
Haplotype analysis
Haplotypes and their frequencies for each gene were inferred by SHEsis online software based on observed genotypes (Table 3). ERCC5 CGTAG, XPA CTC, and DDB2 GTAC and GTAG haplotypes were observed to alter the AG risk, while ERCC5 CGCTG, TATAG, and TGTAG, XPA TCC, XPC GCAAG, and DDB2 GTAC and GTAG haplotypes conferred an altered risk to GC. After Bonferroni correction for multiple testing, the DDB2 GTAG haplotype remained significantly associated with disease risk at each step of the CON→AG→GC development (AG vs CON: OR=2.88, P =7.51 × 10−7; GC vs AG: OR=2.90, P =5.68 × 10−15; GC vs CON: OR=8.42, P =2.22 × 10−15); the DDB2 GTAC haplotype was associated with a reduced risk of GC compared with AG (OR=0.72, P =3.65 × 10−6), and a reduced risk of GC compared with CON (OR=0.63, P =8.31 × 10−12); while the XPC GCAAG haplotype conferred an increased risk of GC compared with AG (OR=1.88, P =6.98 × 10−4).
Table 3: Results of haplotype analysis
Gastric mucosa status |
CON→AG |
AG→GC |
CON→GC |
|||||||
|---|---|---|---|---|---|---|---|---|---|---|
Gene |
Haplotype |
GC(%) |
AG(%) |
CON(%) |
OR(95%CI) |
P |
OR(95%CI) |
P |
OR(95%CI) |
P |
ERCC1 |
CAAAGC |
692.27(39.0) |
688.12(41.1) |
711.71(38.6) |
1.11(0.97-1.28) |
0.131 |
0.92(0.80-1.05) |
0.207 |
1.01(0.88-1.16) |
0.900 |
CCAAGC |
54.02(3.0) |
43.19(2.6) |
40.47(2.2) |
/ |
/ |
1.19(0.79-1.78) |
0.408 |
1.39(0.92-2.10) |
0.115 |
|
CCAATA |
57.27(3.2) |
47.53(2.8) |
67.92(3.7) |
0.76(0.52-1.11) |
0.157 |
1.14(0.77-1.69) |
0.505 |
0.87(0.61-1.24) |
0.434 |
|
CCGCTA |
485.47(27.3) |
471.43(28.2) |
524.91(28.5) |
0.98(0.85-1.14) |
0.806 |
0.96(0.83-1.12) |
0.603 |
0.94(0.81-1.09) |
0.392 |
|
TCACGA |
394.30(22.2) |
338.82(20.2) |
394.59(21.4) |
0.93(0.79-1.09) |
0.374 |
1.13(0.96-1.33) |
0.149 |
1.04(0.89-1.22) |
0.607 |
|
ERCC2 |
CTGC |
96.50(5.4) |
95.68(5.8) |
78.99(4.3) |
1.34(0.99-1.82) |
0.060 |
0.95(0.71-1.27) |
0.708 |
1.27(0.94-1.72) |
0.128 |
CTGG |
750.54(42.3) |
700.52(42.1) |
816.64(44.9) |
0.89(0.77-1.01) |
0.076 |
1.01(0.89-1.16) |
0.838 |
0.90(0.79-1.03) |
0.111 |
|
CTTC |
710.61(40.1) |
664.07(40.0) |
724.72(39.8) |
1.00(0.87-1.14) |
0.962 |
1.01(0.88-1.16) |
0.869 |
1.01(0.88-1.15) |
0.904 |
|
CTTG |
74.34(4.2) |
78.71(4.7) |
66.65(3.7) |
1.30(0.93-1.82) |
0.121 |
0.88(0.64-1.22) |
0.455 |
1.15(0.82-1.61) |
0.418 |
|
TGGG |
95.46(5.4) |
86.44(5.2) |
83.44(4.6) |
1.14(0.83-1.55) |
0.419 |
1.04(0.77-1.41) |
0.790 |
1.18(0.88-1.60) |
0.275 |
|
ERCC3 |
AAC |
187.95(10.6) |
194.00(11.6) |
188.98(10.3) |
1.14(0.92-1.41) |
0.218 |
0.90(0.73-1.12) |
0.353 |
1.03(0.83-1.28) |
0.765 |
AGT |
532.84(30.0) |
500.97(30.0) |
572.98(31.2) |
0.94(0.81-1.09) |
0.409 |
1.01(0.87-1.16) |
0.941 |
0.95(0.82-1.09) |
0.446 |
|
GGC |
1018.86(57.3) |
946.97(56.6) |
1038.96(56.6) |
1.00(0.87-1.14) |
0.992 |
1.04(0.91-1.19) |
0.599 |
1.04(0.91-1.19) |
0.597 |
|
ERCC4 |
CCT |
362.77(20.6) |
357.90(21.5) |
382.91(21.1) |
1.03(0.87-1.21) |
0.757 |
0.95(0.81-1.12) |
0.535 |
0.97(0.83-1.15) |
0.749 |
TGC |
380.99(21.6) |
355.45(21.3) |
398.32(21.9) |
0.97(0.82-1.14) |
0.685 |
1.02(0.87-1.20) |
0.819 |
0.99(0.84-1.16) |
0.859 |
|
TGT |
1007.01(57.2) |
944.55(56.7) |
1028.68(56.6) |
1.01(0.88-1.15) |
0.935 |
1.02(0.89-1.17) |
0.748 |
1.03(0.90-1.17) |
0.681 |
|
ERCC5 |
CGCAG |
209.08(12.0) |
205.77(12.5) |
213.66(11.8) |
1.06(0.86-1.30) |
0.580 |
0.96(0.78-1.18) |
0.721 |
1.02(0.83-1.25) |
0.845 |
CGCTG |
162.74(9.3) |
117.04(7.1) |
139.44(7.7) |
0.91(0.71-1.18) |
0.471 |
1.36(1.06-1.74) |
0.015 |
1.24(0.98-1.57) |
0.078 |
|
CGTAG |
136.67(7.8) |
112.95(6.8) |
161.47(8.9) |
0.75(0.58-0.96) |
0.022 |
1.17(0.90-1.51) |
0.246 |
0.87(0.69-1.10) |
0.252 |
|
TATAG |
614.64(35.3) |
650.29(39.4) |
688.84(38.0) |
1.05(0.92-1.21) |
0.471 |
0.84(0.73-0.97) |
0.018 |
0.89(0.77-1.02) |
0.090 |
|
TGTAA |
84.11(4.8) |
88.92(5.4) |
108.21(6.0) |
0.89(0.67-1.19) |
0.436 |
0.90(0.66-1.22) |
0.481 |
0.80(0.60-1.07) |
0.132 |
|
TGTAG |
502.97(28.9) |
453.82(27.5) |
467.50(25.8) |
1.08(0.93-1.26) |
0.298 |
1.08(0.93-1.25) |
0.323 |
1.17(1.01-1.36) |
0.039 |
|
XPA |
CTC |
370.15(20.8) |
391.46(23.3) |
369.03(19.9) |
1.22(1.04-1.44) |
0.014 |
0.87(0.74-1.02) |
0.086 |
1.06(0.90-1.25) |
0.460 |
TCC |
903.16(50.9) |
792.39(47.2) |
919.38(49.5) |
0.91(0.80-1.04) |
0.157 |
1.16(1.02-1.33) |
0.026 |
1.06(0.93-1.21) |
0.401 |
|
TTC |
334.90(18.9) |
327.15(19.5) |
374.68(20.2) |
0.96(0.81-1.13) |
0.593 |
0.96(0.81-1.14) |
0.663 |
0.92(0.78-1.09) |
0.322 |
|
TTT |
157.88(8.9) |
162.39(9.7) |
185.24(10.0) |
0.97(0.77-1.21) |
0.752 |
0.91(0.73-1.15) |
0.440 |
0.88(0.71-1.10) |
0.267 |
|
XPC |
CCAAC |
477.24(27.0) |
455.68(27.4) |
504.16(27.4) |
1.00(0.86-1.17) |
0.967 |
0.98(0.84-1.14) |
0.789 |
0.98(0.85-1.14) |
0.816 |
GCAAG |
86.07(4.9) |
44.15(2.7) |
64.99(3.5) |
0.75(0.51-1.10) |
0.139 |
1.88(1.30-2.72) |
6.98 × 10−4 |
1.40(1.01-1.95) |
0.044 |
|
GCCAC |
647.59(36.6) |
625.30(37.5) |
659.35(35.8) |
1.08(0.94-1.24) |
0.254 |
0.96(0.83-1.10) |
0.554 |
1.04(0.91-1.19) |
0.582 |
|
GTAAC |
526.97(29.8) |
509.63(30.6) |
584.35(31.7) |
0.95(0.83-1.10) |
0.505 |
0.96(0.83-1.11) |
0.592 |
0.92(0.79-1.06) |
0.220 |
|
DDB2 |
ACGG |
438.89(24.8) |
420.58(25.2) |
446.33(24.0) |
1.06(0.91-1.24) |
0.432 |
0.97(0.83-1.13) |
0.691 |
1.04(0.90-1.22) |
0.580 |
ATAC |
65.19(3.7) |
42.61(2.5) |
51.55(2.8) |
/ |
/ |
1.45(0.98-2.15) |
0.062 |
1.34(0.93-1.94) |
0.120 |
|
ATGG |
66.77(3.8) |
67.10(4.0) |
70.91(3.8) |
1.05(0.75-1.48) |
0.763 |
0.93(0.66-1.31) |
0.675 |
0.99(0.70-1.39) |
0.947 |
|
GCGG |
39.01(2.2) |
48.70(2.9) |
57.12(3.1) |
0.95(0.64-1.40) |
0.779 |
/ |
/ |
0.71(0.47-1.07) |
0.103 |
|
GTAC |
855.35(48.3) |
929.72(55.6) |
1108.04(59.6) |
0.84(0.73-0.96) |
0.013 |
0.72(0.63-0.83) |
3.65 × 10−6 |
0.63(0.55-0.72) |
8.31 × 10−12 |
|
GTAG |
206.44(11.7) |
72.37(4.3) |
28.73(1.5) |
2.88(1.86-4.47) |
7.51 × 10−7 |
2.90(2.20-3.82) |
5.68 × 10−15 |
8.42(5.67-12.51) |
2.22 × 10−15 |
|
GTGG |
86.99(4.9) |
77.40(4.6) |
84.57(4.5) |
1.02(0.74-1.40) |
0.912 |
1.06(0.77-1.44) |
0.739 |
1.09(0.80-1.47) |
0.604 |
For each comparison, genotypes other than the analyzed one were considered to be the reference, and all rare haplotypes with a frequency <0.03 were ignored.
Abbreviations: GC, gastric cancer; AG, atrophic gastritis; CON, control.
NER pathway SNP-environment and SNP-SNP interactions
The interactive effects of NER gene polymorphisms and the environment were next explored. XPA rs2808668 and the drinking of alcohol were found to have an interactive effect on AG risk (Pinteraction=0.009), while DDB2 rs326222, rs3781619, and rs830083 demonstrated interactive effects with smoking in the development of AG (Pinteraction=0.040, 0.005, and 0.007, respectively) (Table 4). XPC rs2607775 showed a significant interaction with smoking (Pinteraction=0.024) on GC risk: (CG+GG) genotype smokers had a 3.84-fold increased risk of developing GC (OR=3.84, 95%CI=1.71-8.64). No interaction was observed between SNPs and the drinking of alcohol nor between SNPs and H. pylori infection on GC risk.
Table 4: Interaction of NER pathway SNPs and environmental factors
CON→AG |
CON→GC |
|||||||
|---|---|---|---|---|---|---|---|---|
Genotype |
Nonsmoker |
Smoker |
Nondrinker |
Drinker |
Nonsmoker |
Smoker |
Nondrinker |
Drinker |
XPA rs2808668 |
||||||||
TT |
1(ref) |
0.94(0.58-1.55) |
1(ref) |
0.50(0.27-0.91) |
1(ref) |
1.63(0.96-2.75) |
1(ref) |
1.34(0.74-2.42) |
CC+CT |
0.94(0.68-1.30) |
0.81(0.56-1.16) |
0.81(0.60-1.09) |
0.83(0.57-1.20) |
1.10(0.76-1.60) |
1.66(1.12-2.47) |
1.01(0.70-1.44) |
1.99(1.32-3.00) |
Pinteraction=0.995 |
Pinteraction=0.009 |
Pinteraction=0.900 |
Pinteraction=0.221 |
|||||
XPC rs2607775 |
||||||||
CC |
1(ref) |
0.87(0.68-1.13) |
1(ref) |
0.91(0.68-1.20) |
1(ref) |
1.42(1.09-1.86) |
1(ref) |
1.78(1.33-2.38) |
CG+GG |
0.78(0.46-1.32) |
0.77(0.27-2.18) |
0.86(0.51-1.44) |
0.47(0.15-1.52) |
0.93(0.52-1.67) |
3.84(1.71-8.64) |
1.18(0.68-2.07) |
2.71(1.17-6.28) |
Pinteraction=0.544 |
Pinteraction=0.447 |
Pinteraction=0.024 |
Pinteraction=0.650 |
|||||
DDB2 rs326222 |
||||||||
CT+TT |
1(ref) |
0.84(0.65-1.10) |
1(ref) |
0.91(0.68-1.21) |
1(ref) |
1.50(1.15-1.95) |
1(ref) |
1.85(1.39-2.48) |
CC |
1.16(0.70-1.95) |
2.27(1.00-5.16) |
1.58(0.96-2.60) |
1.13(0.48-2.69) |
0.79(0.41-1.51) |
1.81(0.71-4.62) |
1.11(0.59-2.07) |
1.51(0.60-3.80) |
Pinteraction=0.040 |
Pinteraction=0.885 |
Pinteraction=0.551 |
Pinteraction=0.782 |
|||||
DDB2 rs3781619 |
||||||||
AG+AA |
1(ref) |
0.79(0.61-1.04) |
1(ref) |
0.87(0.64-1.17) |
1(ref) |
1.52(1.16-2.00) |
1(ref) |
1.87(1.39-2.51) |
GG |
0.93(0.62-1.41) |
1.67(0.94-2.98) |
1.16(0.79-1.71) |
1.22(0.62-2.38) |
0.76(0.46-1.25) |
1.19(0.59-2.39) |
0.89(0.55-1.45) |
1.25(0.58-2.70) |
Pinteraction=0.005 |
Pinteraction=0.287 |
Pinteraction=0.674 |
Pinteraction=0.396 |
|||||
DDB2 rs830083 |
||||||||
CG+CC |
1(ref) |
0.80(0.61-1.05) |
1(ref) |
0.88(0.65-1.19) |
1(ref) |
1.71(1.29-2.29) |
1(ref) |
2.07(1.51-2.84) |
GG |
0.84(0.56-1.25) |
1.31(0.76-2.25) |
1.04(0.72-1.51) |
0.95(0.51-1.76) |
2.22(1.51-3.27) |
2.72(1.58-4.67) |
2.57(1.76-3.75) |
3.11(1.75-5.50) |
Pinteraction=0.007 |
Pinteraction=0.372 |
Pinteraction=0.652 |
Pinteraction=0.167 |
|||||
SNP-environment interaction effects were assessed from the likelihood ratio test, comparing the fit of the logistic model that included the main effects of sex, age, environment risk factor and genotype with a fully parameterized model containing the multiplicative interaction terms of genotype and environment risk factor.
Abbreviations: GC, gastric cancer; AG, atrophic gastritis; CON, control.
Supplementary Table S6 shows the effects of SNP-SNP interactions. No significant interaction was found between SNPs except for that between XPA rs2808668 and DDB2 rs326222 in the development of GC from AG (Pinteraction=0.031).
DISCUSSION
Identifying biomarkers associated with high GC risk has long been a research goal to help improve early disease detection. Although several previous studies have suggested that NER gene polymorphisms could alter GC susceptibility, to the best of our knowledge this is the first large-scale investigation of the relationship between SNPs of the entire NER gene pathway and risks of developing GC and AG. We found that the DDB2 rs830083 GG genotype and XPC rs2607775 CG genotype were associated with increased GC risk, and haplotype analysis revealed that the DDB2 GTAG haplotype significantly increased GC risk. Significant interactions between XPA rs2808668 and the drinking of alcohol, as well as between DDB2 SNPs rs326222, rs3781619, and rs830083 and smoking, were identified in the pathogenesis of AG. Moreover, in GC development, XPC rs2607775 demonstrated a significant interaction with smoking. Thus, polymorphisms in DDB2 and XPC are associated with increased risks of developing GC and AG.
DDB2 encodes damage-specific DNA binding protein 2, which is responsible for damage recognition and the initiating NER. UV-induced DNA damage is recognized by a heterodimer formed from DDB1 and DDB2 [19]. Recent research has revealed that DDB2 is the downstream target of tumour suppressor genes p53 and BRCA1 [20, 21], and it is known to play an important role in regulating p53 function and controlling p53-mediated apoptosis [22, 23]. Yoon et al. found that DDB2-defective mice have a high risk of developing spontaneous tumours, which is indicative of a protective role for DDB2 in cancer development [24].
Human DDB2 is located on chromosome 11p12-p11, and the rs830083 C/G polymorphism is within its intronic region. Previously, rs830083 has been suggested to significantly increase the risk of lung cancer [25], but no study has yet investigated its association with GC. Here, carriers of the DDB2 rs830083 GG genotype demonstrated significantly increased risks of developing AG (OR=1.48) and GC (OR=2.32) compared with the wild-type CC genotype, suggesting that this polymorphism affects different stages of gastric carcinogenesis. Considering the critical role of DDB2 in the NER pathway, this polymorphism could alter the function of DDB2, thereby affecting individual susceptibility to GC. However, the molecular mechanism has yet to be investigated by future studies. We further performed subgroup analysis and showed that the association of the GG genotype and GC was more significant in males, non-smokers, and non-drinkers, suggesting that the DDB2 rs830083 polymorphism might have an improved predictive role in these subgroups. The consistent significant association seen in H. pylori-positive and -negative, as well as intestinal-type and diffuse-type GC indicated that the DDB2 rs830083 polymorphism has a stable predictive effect despite H. pylori infection and GC type. However, as the samples for subgroup analysis were relatively small, further large-scale studies are required to confirm these preliminary findings.
XPC also plays an essential role in the damage recognition step of NER. In mammals, the heterotrimeric XPC complex composed of XPC, RAD23, and centrin-2 recognizes DNA distortions and is indispensable for initiating global genome NER [26]. XPC, located on chromosome 3p25, consists of 16 exons and 15 introns and encodes a DNA-binding protein of 940 amino acids that preferentially binds damaged DNA [27]. The XPC rs2607775 polymorphism is located upstream of the XPC regulatory region and its effect on transcription is largely unknown. In the present study, carriers of the XPC rs2607775 CG genotype had a 1.73-fold increased risk of GC than subjects with the wild-type CC genotype. Subgroup analysis found that the XPC rs2607775 CG genotype still demonstrated a significantly increased GC risk in males and H. pylori-negative subgroups. Moreover, individuals with both DDB2 rs830083 and XPC rs2607775 risk genotypes had a 3.05-fold increased risk of GC (Fig.1), indicating that these two NER genes might serve as future joint predictive biomarkers for GC risk.
Previous studies have identified relationships between some NER gene SNPs and risk of GC in different populations. For example, the ERCC1 rs2298881 and rs11615 polymorphisms were associated with a higher risk of GC in eastern Chinese individuals [28], but we only found the rs11615 polymorphism to be associated with an increased GC risk in H. pylori-positive northern Chinese subjects (Table S4). This might reflect the variation of H. pylori strains from different regions leading to differences in individual environmental exposure. Similarly, ERCC2 rs13181 was reported to alter individual susceptibility to GC [29], but we observed no significant association either in the total or subgroup analysis of our study. Previous studies have drawn inconsistent conclusions regarding the association between ERCC5 rs2296147 and GC risk [30, 31]. In the present study, we found ERCC5 rs2296147 to be associated with increased GC risk in H. pylori-positive and diffuse-type GC subgroups (Table S4 and Table S5). The ERCC5 rs2094258 AA genotype has been reported to be associated with decreased GC risk [31], which is consistent with our findings in H. pylori-negative and diffuse-type GC subgroups. Although ERCC5 rs751402 and rs873601 have been linked with altered GC risk [28, 30], we did not investigate these two polymorphisms further because they were not in accordance with HWE. It is worth noting that XPA rs2808668 showed a protective effect in “CON to AG”, but a risk effect in “AG to GC”. We then performed stratified analysis and found that the risk effect in “AG to GC” was only found in diffuse-type GC but not in intestinal-type GC (Supplementary Table S5). As described by Correa’s cascade, it is accepted that intestinal-type GC progresses stepwise from normal stomach, atrophic gastritis to carcinoma, while the pathogenesis of diffuse-type GC is still unclear[2]. Therefore, the findings of this study might indicate possible involvement of this polymorphism in the complex process of diffuse-type GC, and the underlying mechanisms still require further studies to elucidate.
Haplotype analysis covering SNPs of different regions would be effective at revealing the role of key NER factors and providing a pathway overview, but most previous studies only focused on a few SNPs rather than performing a combined analysis. In this study, we investigated the joint effect of multiple polymorphisms to determine the most important NER pathway factor associated with GC risk. The DDB2 rs2029298-rs326222-rs3781619-rs830083 GTAG haplotype was significantly associated with disease risk at each step of CON→AG→GC development, while the DDB2 GTAC haplotype was associated with a reduced risk of GC compared with CON. Moreover, the XPC rs1870134-rs2228000-rs2228001-rs2470352-rs2607775 GCAAG haplotype conferred an increased risk of GC compared with AG. These findings not only strongly suggest that DDB2 and XPC are GC susceptibility genes, but also highlight the potential of predicting GC risk by detecting key NER pathway haplotypes. Considering that both DDB2 and XPC are critical for the damage recognition stage of NER, we speculate that this might be the crucial step that determines whether GC develops.
Apart from genetic factors, gastric carcinogenesis is also influenced by environmental factors such as H. pylori infection, smoking, and the consumption of alcohol, of which H. pylori is the best known environmental pathogenic factor [32]. Chronic H. pylori infection, smoking, and alcohol consumption can induce persistent inflammation and/or the generation of reactive oxygen species, which leads to DNA damage [33]. Thus, polymorphisms of key NER genes responsible for damage repair may have synergistic effects with H. pylori infection in the development of GC. Previous research found that ERCC5 rs2296147 and rs2094258 interacted with H. pylori infection in the development of GC [31], and that XPC PAT(+/-) showed an interactive effect with smoking in GC [34]. However, no clear overview of the gene-environment interaction in the NER pathway has been provided. Our results suggested that XPA rs2808668 significantly interacted with alcohol consumption in the development of AG. Similarly, DDB2 rs326222, rs3781619, and rs830083 SNPs showed significant interactions with smoking in AG development. XPC rs2607775 had an interactive effect with smoking in GC development, with (CG+GG) genotype smokers having a 3.84-fold increased risk of GC. NER is a complex, multi-step process, and different NER gene polymorphisms might function jointly in gastric carcinogenesis. However, no significant interaction was found between SNPs, except for an interaction between XPA rs2808668 and DDB2 rs326222, in the development of GC from AG.
It is notable that polymorphisms demonstrating significant interactions with environmental factors are all located within genes responsible for the DNA damage recognition step of NER (XPA, XPC, and DDB2). This suggests that NER gene polymorphisms exert interactive effects with environmental factors during DNA damage recognition, thereby influencing an individual’s capacity for DNA repair and susceptibility to GC. This finding may not only provide direction for further investigations into the molecular interactions between NER gene polymorphisms, but may also suggest a potential target for the primary prevention of GC in the future. This could take the form of lifestyle interventions for carriers of high risk GC genotypes and the control of environmental factors. Thus, avoidance of a high salt diet, cessation of smoking and drinking alcohol, and eradication of H. pylori infection, which may all interact with a susceptible genotype to increase GC risk, could be an effective method for disease prevention. One limitation for this study is that the functions and mechanisms of the associated SNPs of this pathway were not analyzed, which need further functional studies to elucidate. Secondly, the gender and age of subjects between cases and controls were not matched although these factors have been adjusted in the multivariable analysis, which might influence the results of the disease association. In addition, because the information of smoking and drinking status was not available for all of the included subjects, we did not adjust the smoking and drinking status in multivariate logistic regression.
In summary, we found for the first time that the two polymorphisms DDB2 rs830083 and XPC rs2607775, affecting the damage recognition step of the NER pathway, were significantly associated with increased GC risk, and that the combined detection of these two polymorphisms demonstrated an even higher GC risk (OR=3.05). Future systematic SNP screening and GWAS studies focusing on the relevant NER pathway may identify more susceptible genes and their interactions with environmental factors. It is anticipated that this would not only provide an effective basis for the early warning of high GC risk individuals with the aim of individualized prevention, but also reveal important molecular mechanisms and novel therapies for GC.
MATERIALS AND METHODS
Study population
A total of 2686 subjects were selected for inclusion in the present study. All enrolled individuals were unrelated Han ethnic Chinese living in northern China, who were recruited from a health check program for GC screening or hospitals in Zhuanghe and Shenyang of Liaoning Province, China between 2002 and 2013. Patients with a history of other malignancies were excluded. The gastroscopy examination was performed by experienced endoscopists. Four biopsy specimens were obtained from the body, angulus, antrum, and site of the lesion. Each subject was assigned a global diagnosis based on the most severe lesion among the four biopsy specimens, and this was confirmed independently by two gastrointestinal pathologists. Histopathological findings were assessed according to the Consensus on Chronic Gastritis formulated at the National Symposium in combination with the updated Sydney System [15] and the World Health Organization criteria.
Three groups of subjects were retrospectively selected from the total participants based on their baseline diagnosis: (i) a control (CON) group of individuals with normal stomach and subjects with slight or moderate gastritis without atrophic or intestinal metaplasia lesions (n=937); (ii) an atrophic gastritis (AG) group with or without intestinal metaplasia (n=851); and (iii) a gastric cancer (GC) group (n=898). Subjects were excluded from the CON group if they had gastric erosion, peptic ulcer disease, gastric polyps, adenomas, or diseases related to or causing predisposition to cancer.
Each participant was interviewed face-to-face by trained interviewers using a standardized questionnaire. Written informed consent was obtained from each participant. Data including gender, age, history of illness, native origin, pathological diagnosis, and status of smoking, alcohol consumption, and H. pylori infection were coded in a specific databank. Individuals who smoked at least once a day for more than 1 year were defined as smokers, and the others were defined as non-smokers. Subjects who consumed alcohol at least once a week for more than 1 year were defined as drinkers, while the remainder were non-drinkers.
Fasting blood samples (5 ml) were collected from each patient for DNA extraction and measuring serum H. pylori immunoglobulin G levels. This study was approved by the human ethics review committee of China Medical University (Shenyang, China).
Candidate genes and SNP selection
We extracted genotype data from extended NER gene regions encompassing 5 kb of upstream and downstream flanking sequences from the HapMap Chinese Han Beijing population (Release 27, Phase I + II + III, http://www.HapMap.org). Haploview software (http://www.broadinstitute.org/mpg/haploview) was used to minimize the number of SNPs required to be genotyped, providing an important shortcut to carry out candidate gene association studies in a particular population. Tag SNPs were chosen based on pairwise linkage disequilibrium information to maximally capture (r2 > 0.8) common or rare variants (minor allele frequency [MAF] > 0.05) by Haploview 4.2. FastSNP Search was used to predict the potential SNP function (leading to amino acid substitutions, altering splicing or transcription factor-binding motifs, acting as intronic enhancers) [16, 17]. A total of 39 SNPs covering eight key NER pathway genes (ERCC1, ERCC2, ERCC3, ERCC4, ERCC5, XPA, XPC, and DDB2) were selected by integrating these two publicly available tools. The flow chart of the detailed SNP selecting strategy was summarized in Supplementary Figure 1.
Genotyping assay
Genomic DNA was isolated from blood samples using routine phenol-chloroform extraction and then diluted to working concentrations (50 ng/ μ l) for genotyping. Samples were placed randomly on the 384-well plates and blinded for disease status. The design of the assay and SNP genotyping were performed by Bio Miao Biological Technology (Beijing, China) using the Sequenom MassARRAY platform (Sequenom, San Diego, CA) according to the manufacturer’s instructions. The average genotyping rate was 99.3% and the results of all duplicated samples were 100% consistent.
H. pylori serology
H. pylori serology was performed to examine the status of H. pylori infection using an enzyme-linked immunosorbent assay. H. pylori immunoglobulin G concentrations of serum samples were detected by an ELISA kit (Biohit, Helsinki, Finland) according to the manufacturer’s instructions. A numerical reading exceeding 34 enzyme immune-units was considered to be H. pylori-positive.
Statistical analysis
HWE for each SNP was first evaluated among control subjects using either the chi-square (χ2) test or Fisher’s exact test. We excluded SNPs that deviated from HWE from subsequent association analysis. Age differences between groups were assessed using the analysis of variance test. The χ2 test was applied to evaluate differences in categorical variables including gender, H. pylori infection, smoking status, and consumption of alcohol. The association between each SNP and risk of AG and GC was estimated by calculating ORs and their 95% CIs by multivariate logistic regression with adjustments for gender, age, and H. pylori infection status. Recessive model, dominant model, co-dominant model and additive model were adopted. Stratified analysis by gender, Lauren’s classification, H. pylori infection, and smoking and alcohol consumption status was also conducted. To compare the SNP effect on the risks of AG and GC between different subgroups, the Breslow-Day test was used to assess the homogeneity of stratum-specific ORs across different subgroups. Statistical significance was set as P ≤ 0.10 for this test. To limit spurious findings, we used the Bonferroni correction for multiple comparisons considering significance thresholds for SNP association as P =1.5 × 10−3 (0.05/33 SNPs) and for haplotype association as P=6.25 × 10−3 (0.05/8 genes). This is a fairly stringent correction given that not all of the SNPs analyzed may be independent of each other because of linkage disequilibrium. SNP-environment interaction effects were assessed from the likelihood ratio test, comparing the fit of the logistic model that included the main effects of sex, age, environment risk factor and genotype with a fully parameterized model containing the multiplicative interaction terms of genotype and environment risk factor. Likelihood ratio test was also performed to assess the SNP-SNP interaction effects on the risk of GC by comparing the model only involving main effects of gender, age, genotype with the full model which also contained the SNP-SNP interaction term. The abovementioned statistical analyses for SNP association were performed using SPSS 16.0 software (SPSS, Chicago, IL). Haplotype association analyses were performed for all investigated genes by SHEsis online software (http://analysis.bio-x.cn/myAnalysis.php) [18] to explore the relation between haplotype and disease risk. For each comparison, genotypes other than the analyzed one were considered to be the reference, and all rare haplotypes with a frequency < 0.03 were ignored.
CONFLICT OF INTEREST
All of the authors declare that there is no conflict of interest.
FUNDING
This study is supported by grants from National Basic Research Program of China (973 Program Ref No.2010CB529304), the grants of Finance department of Liaoning province (Ref No. [2008]621) and the grants of the Science and Technology Project of Liaoning province (Ref No.2012225016).
REFERENCE
1. Siegel R, Ward E, Brawley O and Jemal A. Cancer statistics, 2011: the impact of eliminating socioeconomic and racial disparities on premature cancer deaths. CA Cancer J Clin. 2011; 61:212–236.
2. Correa P. Human gastric carcinogenesis: a multistep and multifactorial process--First American Cancer Society Award Lecture on Cancer Epidemiology and Prevention. Cancer Res. 1992; 52:6735–6740.
3. Zabaleta J. Multifactorial etiology of gastric cancer. Methods Mol Biol. 2012; 863:411–435.
4. Sakamoto H, Yoshimura K, Saeki N, Katai H, Shimoda T, Matsuno Y, Saito D, Sugimura H, Tanioka F, Kato S, Matsukura N, Matsuda N, Nakamura T, Hyodo I, Nishina T, Yasui W, et al. Genetic variation in PSCA is associated with susceptibility to diffuse-type gastric cancer. Nat Genet. 2008; 40:730–740.
5. Abnet CC, Freedman ND, Hu N, Wang Z, Yu K, Shu XO, Yuan JM, Zheng W, Dawsey SM, Dong LM, Lee MP, Ding T, Qiao YL, Gao YT, Koh WP, Xiang YB, et al. A shared susceptibility locus in PLCE1 at 10q23 for gastric adenocarcinoma and esophageal squamous cell carcinoma. Nat Genet. 2010; 42:764–767.
6. Shi Y, Hu Z, Wu C, Dai J, Li H, Dong J, Wang M, Miao X, Zhou Y, Lu F, Zhang H, Hu L, Jiang Y, Li Z, Chu M, Ma H, et al. A genome-wide association study identifies new susceptibility loci for non-cardia gastric cancer at 3q13.31 and 5p13.1. Nat Genet. 2011; 43:1215–1218.
7. He C, Tu H, Sun L, Xu Q, Li P, Gong Y, Dong N and Yuan Y. Helicobacter pylori-related host gene polymorphisms associated with susceptibility of gastric carcinogenesis: a two-stage case-control study in Chinese. Carcinogenesis. 2013; 34:1450–1457.
8. Zhang Y, Sun LP, Xing CZ, Xu Q, He CY, Li P, Gong YH, Liu YP and Yuan Y. Interaction between GSTP1 Val allele and H. pylori infection, smoking and alcohol consumption and risk of gastric cancer among the Chinese population. PLoS One. 2012; 7:e47178.
9. de Laat WL, Jaspers NG and Hoeijmakers JH. Molecular mechanism of nucleotide excision repair. Genes Dev. 1999; 13:768–785.
10. Nouspikel T. DNA repair in mammalian cells : Nucleotide excision repair: variations on versatility. Cell Mol Life Sci. 2009; 66:994–1009.
11. Hoeijmakers JH. Genome maintenance mechanisms for preventing cancer. Nature. 2001; 411:366–374.
12. He J, Qiu LX, Wang MY, Hua RX, Zhang RX, Yu HP, Wang YN, Sun MH, Zhou XY, Yang YJ, Wang JC, Jin L, Wei QY and Li J. Polymorphisms in the XPG gene and risk of gastric cancer in Chinese populations. Hum Genet. 2012; 131:1235–1244.
13. Liu J, He C, Xing C and Yuan Y. Nucleotide excision repair related gene polymorphisms and genetic susceptibility, chemotherapeutic sensitivity and prognosis of gastric cancer. Mutat Res Fundam Mol Mech Mutagen. 2014; 765C:11–21.
14. Chen Z, Zhang C, Xu C, Li K, Hou R, Li D and Cheng X. Effects of selected genetic polymorphisms in xeroderma pigmentosum complementary group D on gastric cancer. Mol Biol Rep. 2011; 38:1507–1513.
15. Dixon MF, Genta RM, Yardley JH and Correa P. Classification and grading of gastritis. The updated Sydney System. International Workshop on the Histopathology of Gastritis, Houston 1994. Am J Surg Pathol. 1996; 20:1161–1181.
16. Tabor HK, Risch NJ and Myers RM. Candidate-gene approaches for studying complex genetic traits: practical considerations. Nat Rev Genet. 2002; 3:391–397.
17. Yuan HY, Chiou JJ, Tseng WH, Liu CH, Liu CK, Lin YJ, Wang HH, Yao A, Chen YT and Hsu CN. FASTSNP: an always up-to-date and extendable service for SNP function analysis and prioritization. Nucleic Acids Res. 2006; 34(Web Server issue):W635–641.
18. Shi YY and He L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005; 15:97–98.
19. Payne A and Chu G. Xeroderma pigmentosum group E binding factor recognizes a broad spectrum of DNA damage. Mutat Res. 1994; 310:89–102.
20. Hwang BJ, Ford JM, Hanawalt PC and Chu G. Expression of the p48 xeroderma pigmentosum gene is p53-dependent and is involved in global genomic repair. Proc Natl Acad Sci U S A. 1999; 96:424–428.
21. Hartman AR and Ford JM. BRCA1 induces DNA damage recognition factors and enhances nucleotide excision repair. Nat Genet. 2002; 32:180–184.
22. Itoh T, Cado D, Kamide R and Linn S. DDB2 gene disruption leads to skin tumors and resistance to apoptosis after exposure to ultraviolet light but not a chemical carcinogen. Proc Natl Acad Sci U S A. 2004; 101:2052–2057.
23. Itoh T, O'Shea C and Linn S. Impaired regulation of tumor suppressor p53 caused by mutations in the xeroderma pigmentosum DDB2 gene: mutual regulatory interactions between p48(DDB2) and p53. Mol Cell Biol. 2003; 23:7540–7553.
24. Yoon T, Chakrabortty A, Franks R, Valli T, Kiyokawa H and Raychaudhuri P. Tumor-prone phenotype of the DDB2-deficient mice. Oncogene. 2005; 24:469–478.
25. Hu Z, Shao M, Yuan J, Xu L, Wang F, Wang Y, Yuan W, Qian J, Ma H, Wang Y, Liu H, Chen W, Yang L, Jin G, Huo X, Chen F, et al. Polymorphisms in DNA damage binding protein 2 (DDB2) and susceptibility of primary lung cancer in the Chinese: a case-control study. Carcinogenesis. 2006; 27:1475–1480.
26. Araki M, Masutani C, Takemura M, Uchida A, Sugasawa K, Kondoh J, Ohkuma Y and Hanaoka F. Centrosome protein centrin 2/caltractin 1 is part of the xeroderma pigmentosum group C complex that initiates global genome nucleotide excision repair. J Biol Chem. 2001; 276:18665–18672.
27. Sugasawa K. XPC: its product and biological roles. Adv Exp Med Biol. 2008; 637:47–56.
28. He J, Xu Y, Qiu LX, Li J, Zhou XY, Sun MH, Wang JC, Yang YJ, Jin L, Wei QY and Wang Y. Polymorphisms in ERCC1 and XPF genes and risk of gastric cancer in an eastern Chinese population. PLoS One. 2012; 7:e49308.
29. Long XD, Ma Y, Huang YZ, Yi Y, Liang QX, Ma AM, Zeng LP and Fu GH. Genetic polymorphisms in DNA repair genes XPC, XPD, and XRCC4, and susceptibility to Helicobacter pylori infection-related gastric antrum adenocarcinoma in Guangxi population, China. Mol Carcinog. 2010; 49:611–618.
30. Duan Z, He C, Gong Y, Li P, Xu Q, Sun LP, Wang Z, Xing C and Yuan Y. Promoter polymorphisms in DNA repair gene ERCC5 and susceptibility to gastric cancer in Chinese. Gene. 2012; 511:274–279.
31. Yang WG, Zhang SF, Chen JW, Li L, Wang WP and Zhang XF. SNPs of excision repair cross complementing group 5 and gastric cancer risk in Chinese populations. Asian Pac J Cancer Prev. 2012; 13:6269–6272.
32. Wroblewski LE and Peek RM, Jr. Helicobacter pylori in gastric carcinogenesis: mechanisms. Gastroenterol Clin North Am. 2013; 42:285–298.
33. Obst B, Wagner S, Sewing KF and Beil W. Helicobacter pylori causes DNA damage in gastric epithelial cells. Carcinogenesis. 2000; 21:1111–1115.
34. Palli D, Polidoro S, D'Errico M, Saieva C, Guarrera S, Calcagnile AS, Sera F, Allione A, Gemma S, Zanna I, Filomena A, Testai E, Caini S, Moretti R, Gomez-Miguel MJ, Nesi G, et al. Polymorphic DNA repair and metabolic genes: a multigenic study on gastric cancer. Mutagenesis. 2010; 25:569–575.