
INTRODUCTION
Gastric cancer (GC) is a leading cause of cancer-related death worldwide [1, 2], and about 90% of non-cardia GCs are the ultimate consequence of longstanding Helicobacter pylori (H. pylori) infection [3–7]. H. pylori is the main trigger of Correa’s multistep gastric carcinogenic cascade [8]. The stepwise changes occurring in the gastric mucosa have been well characterized, starting from longstanding inflammation, which may result in atrophic changes. Gastric mucosal atrophy is the “cancerization field” in which intra-epithelial neoplasia (IEN), and invasive cancer can develop [3, 9, 10].
In a minority of cases, the initiating cause of the mucosal inflammation is not environmental (H. pylori), but host-related (autoimmune gastritis [AIG]) [11, 12]. Autoimmune, corpus-restricted gastritis may result in mucosal atrophy too, and a higher risk of GC has been associated with AIG as well. The epidemiological and clinico-biological profiles of this “alternative” oncogenic pathway are less well defined than those of the H. pylori-related model [9, 13].
Despite recent extensive investigations on the molecular landscape of GC [14], the molecular grounds for the various steps in H. pylori-related carcinogenesis have yet to be fully elucidated, and no consistent and reliable biomarkers have become available for use in GC secondary prevention strategies [9, 15, 16].
Aberrant microRNA (miRNA) expression has consistently been reported in both H. pylori infection and H. pylori-related GC [17–19], but the miRNA dysregulation(s) associated with each of the phenotypic changes sequentially occurring in H. pylori-related carcinogenesis remain elusive. Among other miRNAs, the let-7 family members have reliably been found down-regulated in association with both H. pylori-associated gastritis [20, 21], and gastric cancer [22, 23]. Moreover, a significant down-regulation of let-7c (a let-7 family member) consistently occurs in the natural history of Barrett-adenocarcinoma (which features remarking similarities with H. pylori-related gastric carcinogenesis) [24].
This study focused on the involvement of let-7c over the whole series of non-neoplastic and neoplastic phenotypic changes occurring in H. pylori-infected gastric mucosa. Let-7c expression was profiled by qRT-PCR in a large series of formalin-fixed paraffin-embedded (FFPE) biopsy samples. The results obtained were then confirmed by miRNA in situ hybridization on surgical tissue specimens, and validated by exploring the publicly-available NCBI-GEO and TCGA databases.
RESULTS
Let-7c is down-regulated in Helicobacter pylori-related gastritis
In 80 biopsy samples (from 40 H. pylori-positive and 40 H. pylori-negative subjects), a significant let-7c down-regulation (qRT-PCR) was associated with H. pylori-positive non-atrophic gastritis, in both antral and oxyntic mucosa samples (p=0.011 and p=0.007, respectively; t-test; Figure 1A). For 10 patients enrolled as H. pylori-positive, paired biopsy samples were obtained after the bacterium’s eradication; in 8 of these 10 cases, let-7c expression was significantly higher after the eradication treatment (p=0.009; t-test; Figure 1B).
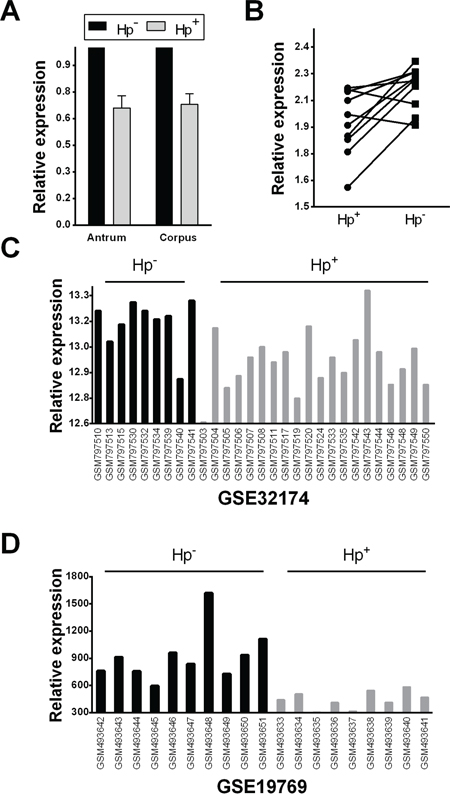
Figure 1: Let-7c is significantly down-regulated in Helicobacter pylori infection. A. Let-7c expression was analyzed by qRT-PCR in a series of 40 samples of normal gastric mucosa and 40 of H. pylori-related gastritis. The latter showed a significantly down-regulated let-7c expression in both gastric compartments (antrum and corpus; p<0.001; relative expression= fold changes compared to normal mucosa samples set as 1.0). B. Let-7c expression levels were significantly up-regulated after H. pylori eradication in 8/10 patients (p=0.009; relative expression= distribution of normalized data in the two classes). C–D. Two independent studies in the NCBI GEO database showed a significant down-regulation of let-7c in H. pylori-positive samples (GSE32174 [31], adjusted p=3.47E-02; and GSE19769 [20], adjusted p=4.3E-04; relative expression= distribution of normalized data obtained from the GEO database).
The relationship between H. pylori infection and let-7c expression was further explored in the NCBI-GEO database and using the GEO2R microarray analysis tool. In 2 of the 3 available series, H. pylori-positive samples featured a significant let-7c down-regulation (GSE32174 [31], adjusted p=3.47E-02; and GSE19769 [20], adjusted p=4.3E-04; t-test) (Figure 1C–1D); in the third series, let-7c expression was still lower, though not significantly so, in H. pylori-positive than in normal mucosa (GSE54397 [32]; data not shown).
Let-7c is down-regulated during H. pylori-related gastric carcinogenesis
Let-7c expression was tested (qRT-PCR) in 115 FFPE samples representative of normal mucosa and each of the phenotypic lesions occurring in the oncogenic cascade (normal antral mucosa, 20 cases; IM, 35 cases; LG-IEN, 15 cases; HG-IEN, 15 cases; intestinal-type GC, 30 cases).
Overall, let-7c expression decreased significantly with the increasing severity of the lesions considered (p<0.001; ANOVA; Figure 2A). Compared with normal H. pylori-negative antral mucosa, let-7c down-regulation was significant in IM (p=0.041; t-test), LG-IEN (p=0.005; t-test), HG-IEN (p=0.002; t-test), and GC (p<0.001; t-test).
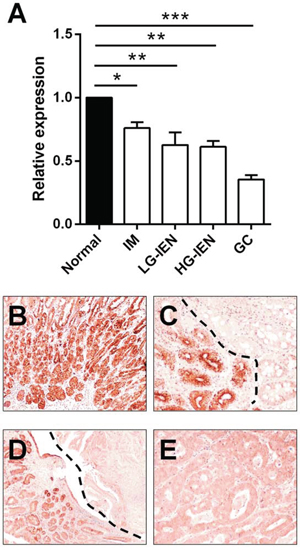
Figure 2: Let-7c is significantly down-regulated during Helicobacter pylori-related gastric carcinogenesis. A. Let-7c expression, analyzed by qRT-PCR in 115 FFPE biopsy samples, gradually decreased along the H. pylori-related gastric carcinogenic cascade (p<0.001; relative expression= fold changes compared to normal antral mucosa samples set as 1.0). Compared with normal antral mucosa, let-7c was significantly down-regulated in intestinal metaplasia (IM; p=0.041), low-grade intraepithelial neoplasia (LG-IEN; p=0.005), high-grade IEN (HG-IEN; p=0.002), and intestinal-type gastric cancer (GC; p<0.001). (*=p<0.05; **=p<0.01 ***=p<0.001) B–E. ISH consistently revealed a strong let-7c expression in normal antral epithelia B, whereas samples of IM (C, top right), IEN (D, top right), and GC E. showed faint or no staining. (Original magnifications 10x and 20x)
These results were validated by testing let-7c expression (in situ hybridization) on FFPE samples obtained from 5 gastrectomy specimens of intestinal-type GC (Figure 2B–2E). In the multiple tissue samples obtained from each of the surgical specimens considered, strong let-7c hybridization signals were consistently found in native antral mucosecreting epithelia (Figure 2B), while a significant let-7c down-regulation was associated with intestinal metaplasia (Figure 2C), IEN (Figure 2D), and overt adenocarcinoma (Figure 2E).
The above-mentioned results are basically consistent with the profile of let-7c (precursor and mature forms) in The Cancer Genome Atlas (TCGA). As emerged from the Atlas database, the levels of both precursor and mature forms of let-7c were significantly lower in cancer than in normal mucosa among 177 cases of intestinal-type GC, and 41 cases of normal gastric mucosa (both p<0.001; t-test; Figure 3A). Moreover, among the 15 TCGA cases of matched normal and GC samples, cancers featured let-7c down-regulation in its precursor (11/15) and/or mature form (13/15) (p=0.022 and p=0.029, respectively; paired t-test) (Figure 3B).
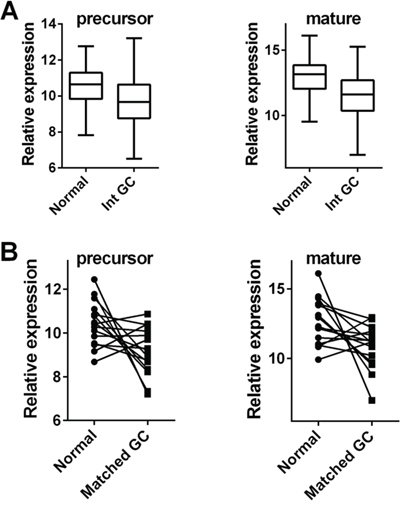
Figure 3: Let-7c is significantly down-regulated in intestinal-type gastric adenocarcinoma. Let-7c expression was explored in intestinal-type GCs and matched normal gastric mucosa series profiled in The Cancer Genome Atlas (TCGA) [14]. A. The levels of both precursor and mature forms of let-7c were significantly lower in GC (n=177) than in normal gastric mucosa (n=41) (both p<0.001; relative expression= distribution of normalized data obtained from the TCGA database). B. In 15 TCGA patients with matched normal/GC samples, both the precursor and the mature let-7c form were down-regulated in the neoplastic samples (p=0.022 and p=0.029, respectively; relative expression= distribution of normalized data obtained from the TCGA database).
In autoimmune gastritis, let-7c down-regulation is associated with intestinal type, but not pseudo-pyloric type metaplasia
As seen in the H. pylori-related gastritis model, corpus-restricted autoimmune gastritis may also result in the atrophic transformation of the native mucosa, often associated with metaplastic (pseudo-pyloric and intestinal) transformation of the resident specialized epithelia [25]. These lesions are currently considered at higher risk of cancer onset and frequently coexist with enterochromaffin-like (ECL) cell hyperplasia, both linear and micronodular (the precursor lesions of gastric carcinoids).
In this “autoimmune model”, let-7c status was tested to assess its potential involvement in the earliest phases of the morphogenesis involved in atrophic transformation unrelated to H. pylori.
Let-7c expression was assessed by qRT-PCR in 40 FFPE corpus biopsy samples (20 cases of corpus-restricted non-atrophic gastritis, and 20 of intestinal metaplasia) obtained from patients with AIG and no history of H. pylori infection. Compared with normal oxyntic mucosa samples, a significant let-7c down-regulation was consistently associated with both non-atrophic AIG and AIG-related IM (both p<0.001; t-test) (Figure 4A).
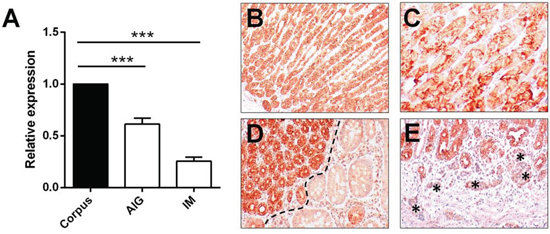
Figure 4: Let-7c is significantly down-regulated in autoimmune gastritis. A. Let-7c expression was measured (qRT-PCR) in 40 FFPE corpus biopsy samples obtained from patients with autoimmune gastritis (20 cases of corpus-restricted non-atrophic gastritis [AIG] and 20 of intestinal metaplasia [IM]). Both groups showed a significant down-regulation of the miRNA by comparison with normal oxyntic mucosa (***= p<0.001; relative expression= fold changes compared to normal oxynthic mucosa samples set as 1.0). ISH for let-7c consistently showed strong staining in oxyntic mucosa B, and a weaker expression in parietal cells by comparison with other oxyntic cell types C. Strong let-7c expression was also apparent in pseudo-pyloric metaplastic epithelia (on left of panel), whereas a significant down-regulation was seen in IM D. ECL hyperplasia (asterisks) showed faint let-7c staining E. (Original magnifications 10x and 20x)
Let-7c expression was further tested by in situ hybridization in a series of 15 FFPE samples representative of the histological spectrum of AIG (non-atrophic and atrophic variants). The biopsy samples included 5 cases of normal oxyntic mucosa, 5 of non-atrophic AIG, and 5 of oxyntic atrophic-metaplastic gastritis (including both intestinal and pseudo-pyloric metaplasia; Figure 4B–4E).
Let-7c hybridization consistently disclosed a normal miRNA expression in normal oxyntic glands (Figure 4B–4C), and metaplastic pseudo-pyloric epithelia, while a significant down-regulation was apparent in intestinalized cells (Figure 4D). Any ECL hyperplasia showed faint let-7c staining (Figure 4E).
DISCUSSION
Lethal-7 (let-7) is a founding member of the miRNA family of genes that was first identified in Caenorhabditis elegans [26–28]. Several homologs of C. elegans let-7 have been recognized in humans, including hsa-let-7c [27]. Members of the human let-7 family share an identical seed sequence crucial to target recognition. They are distributed in 12 genomic loci known as breakpoint regions and fragile sites that are commonly affected in cancer [29, 30]. Several independent studies have indicated that let-7 members act as tumor suppressors on various tumor types by targeting KRAS and HMGA2 oncogenes [23, 31–35].
Like other malignancies, GC is consistently associated with let-7 down-regulation [23]. In a large series of GCs, Motoyama et al. first demonstrated that let-7 family members negatively regulated HMGA2 expression [23]. By profiling H. pylori-infected gastric mucosa using miRNA microarrays, Matsushima et al. demonstrated an important down-regulation (>2-fold) of 5 members of the let-7 family (let-7a, let-7b, let-7d, let-7e, let-7f) in H. pylori-positive mucosa [20]. In the 4 patients successfully treated to eradicate the bacterium, there was subsequently a significant increase in 14 miRNAs, including let-7a, let-7b, let-7d, and let-7e [20]. Similar results emerged from a comprehensive miRNA array analysis on gastric epithelial cells transfected with the H. pylori cagA gene [21]. In this in vitro model, Y. Hayashi et al. also found let-7 family down-regulation associated with an increased DNA-methyltransferase-DNMT3B activity, resulting in histone and DNA methylation of the let-7 gene promoter (21).
As for the present study, we first explored hsa-let-7c expression in the whole spectrum of phenotypic changes occurring in the H. pylori-related carcinogenic cascade. In a large series of representative gastric mucosa biopsy samples, let-7c was found significantly down-regulated in H. pylori-infected mucosa samples (from the antrum and corpus), and its expression was restored after H. pylori eradication.
We demonstrated that let-7c expression drops significantly (qRT-PCR) since the earliest phenotypic changes (Intestinal Metaplasia) occurring in H. pylori-associated carcinogenesis and its loss of expression parallels the increasing severity of the histological changes. The significant miRNA down-regulation featured already in IM samples (ISH) is also consistent with a significant down-regulation of let-7c in serum, as recently reported in a large series of patients with atrophic gastritis (n=222), and GC (n=214) [22]. In line with our results, these data further support the candidacy of let-7c as a promising noninvasive biomarker in GC [22]. Next step will be the evaluation of both tissue and circulating let-7c expression levels in a large series of atrophic gastritis patients stratified according to different gastritis stages [11] to assess let-7c prognostic impact as a noninvasive biomarker in the progression from initial gastritis phases to cancer-prone multifocal gastritis.
We also tested let-7c expression in a model of autoimmune gastritis. In this setting, both non-atrophic and IM-atrophic biopsy samples showed a significant miRNA down-regulation compared with normal oxyntic mucosa (both p<0.001). The significant down-regulation observed on ISH in IM (but not in pseudo-pyloric metaplasia) suggests a role for let-7c in driving the earliest changes occurring in the gastric carcinogenic cascade, while it does not support any elective oncogenic influence of pseudo-pylorized metaplastic cells [25, 36–38].
A recent study [39] pinpointed the significant role of HMGA2, a let-7c target, in the progression and prognosis of GC through the activation of the epithelial-mesenchymal transition process in GC cells. These data and the present results (i.e., let-7c expression decreased significantly with the increasing severity of the lesions considered) support an important role of let-7c in both the initiation and progression of the gastric neoplastic transformation.
In conclusion, our findings in a large series of histologically well-profiled phenotypic lesions demonstrate that let-7c down-regulation plays a pathogenic part right from the earliest phenotypic changes taking place in the H. pylori-related carcinogenic process, and further supports the tumor suppressor activity of this miRNA family. The reversibility of let-7c down-regulation, as seen after H. pylori eradication, also testifies to a “functional plasticity” of this potential candidate for use as an adjunctive test on gastric mucosa status. We also demonstrated for the first time that let-7c is significantly down-regulated even in early cancer precursor lesions occurring in autoimmune gastric oncogenesis. Overall, these results provide a solid rationale for further exploring the diagnostic reliability of this miRNA family as a novel biomarker for use in GC secondary prevention strategies.
MATERIALS AND METHODS
Ethics statement
Investigation has been conducted in accordance with the ethical standards and according to the Declaration of Helsinki and according to national and international guidelines and has been approved by the authors’ institutional review board (#0012272/15).
Tissue samples
The cases considered in this study were retrospectively collected from the files of the Surgical Pathology and Cytopathology Unit of the Department of Medicine (DIMED - University of Padua). The Institute’s ethical regulations concerning research conducted on human tissues were followed.
A total of 195 endoscopic formalin-fixed paraffin-embedded (FFPE) biopsy samples were considered in the let-7c qRT-PCR analysis. The tissue samples were selected from 140 different biopsy sets obtained from 130 Caucasian patients (M/F: 1.5/1; mean age 63.2±8.4 years) who underwent gastro-esophageal endoscopy at the Gastroenterology Unit of the Department of Surgical Oncology and Gastroenterology (DiSCOG, University of Padua).
Biopsy samples were representative of each of the phenotypic lesions in the H. pylori-related gastric carcinogenic cascade [11, 40–43]. They included: i) 40 normal gastric mucosa samples (20 antral, 20 oxyntic) from 20 patients undergoing endoscopy for functional dyspepsia; ii) 40 H. pylori-positive non-atrophic gastritis samples (20 antral, 20 oxyntic) from 20 patients (whose H. pylori status was assessed histologically by means of Giemsa staining); iii) 20 antral mucosa samples obtained from 10 patients before and after H. pylori eradication therapy (median follow-up: 10 weeks; H. pylori eradication was always confirmed with the 13C-urea breath and/or stool antigen tests); iv) 35 antral mucosa samples with extensive intestinal metaplasia (IM; histochemically sub-typed as Type II-III IM by means of high iron diamine staining) obtained from patients with OLGA stage III and IV gastritis; v) 15 low-grade intra-epithelial neoplasia samples (LG-IEN; formerly called ‘low-grade dysplasia’); vi) 15 high-grade IEN samples (HG-IEN; formerly ‘high-grade dysplasia’); vii) 30 well- or moderately-differentiated intestinal-type GC samples (all G1/G2, and all pT1 or pT2).
A further 40 FFPE oxyntic mucosa samples were obtained from 40 Caucasian patients (M/F: 0.8/1; mean age 55.2±9.0 years) with a clinical history of autoimmune gastritis (AIG). In all cases, their autoimmune etiology was assessed histologically and confirmed by serologically testing anti-parietal cells and/or anti-intrinsic factor autoantibodies (Autozyme IFAb; Cambridge Life Sciences, Ely, UK) [13]. These biopsies included: i) 20 samples of non-atrophic corpus-restricted gastritis; ii) 20 oxyntic mucosa samples with extensive IM obtained from OLGA stage II and III autoimmune gastritis.
In situ hybridization (ISH) was performed on FFPE tissue samples obtained from 5 gastrectomy specimens for adenocarcinoma (all distal cancers associated with ongoing or previous H. pylori infection); and 10 biopsy samples of oxyntic mucosa obtained from AIG patients were also included. Where indicated, tissue samples were micro-dissected from the formalin-fixed paraffin-embedded tissues so that the tested tissue samples always included at least 70% of the target lesion (as further validated on serial H&E histology sections). All histological assessments were performed jointly by three pathologists (MF, DS, IC).
Reverse transcription and quantitative real-time PCR
Total RNA was extracted using the RecoverAll kit (Ambion, Austin, TX, USA). The NCodeTM miRNA qRT-PCR method (Invitrogen, Carlsbad, California, USA) was applied to detect and quantify mature hsa-let-7c according to the manufacturer’s instructions, using an ABI Prism 7900HT Sequence Detection System (Applied Biosystems, Foster City, CA) [24, 44]. Normalization was done with the small nuclear RNA U6B. All reactions were run in duplicate, including no-template controls. The fold difference for each sample was obtained using the ΔΔCT method.
Let-7c in situ hybridization (ISH)
Locked nucleic acid (LNA) probes with complementarity to let-7c were labeled with 5′-biotin and synthesized using Exiqon (Vedbaek, Denmark). Tissue sections were digested with ISH protease 1 (Ventana Medical Systems, Milan, Italy) and ISH was performed as previously described [45], with minor modifications. Positive (U6; Exiqon) and negative scrambled LNA probes were used as controls. Only cytoplasmic let-7c intensity was retained for scoring purposes.
Array database meta-analysis
The NCBI-GEO repository of published array data and the GEO2R microarray analysis tool were used (15 June 2015) to assess let-7c expression in H. pylori-related gastritis (using the keywords: microRNA, gastritis, Helicobacter pylori, Homo sapiens, gastric). The GEO2R algorithms were used for the statistical analysis on the differences in let-7c expression between the groups (normal gastric mucosa vs Hp-gastritis).
To test let-7c dysregulation in gastric cancer, let-7c expression was explored among intestinal-type gastric adenocarcinomas and matched normal gastric mucosa series profiled in The Cancer Genome Atlas (TCGA) initiative [14]. Normalized let-7c expression data were used in the analysis.
Statistical analysis
Differences between groups were tested by applying the (paired) t-test, and ordinary one-way ANOVA, as appropriate. P values <0.05 were considered significant. The statistical analysis was performed using STATA software (Stata Corporation, College Station, TX).
ACKNOWLEDGMENTS AND FUNDING
All authors took part in writing the manuscript and approved the final, submitted version. We would like to thank Dr. Roberta Salmaso for her technical assistance.
This work was partly supported by a grant from the Italian Association for Cancer Research (AIRC Regional grant 2008 N. 6421).
CONFLICTS OF INTEREST
The authors declare no conflicts of interest.
REFERENCES
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2015. CA Cancer J Clin. 2015; 65: 5-29.
2. Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015; 65: 87-108.
3. Rugge M, Fassan M, Graham DY. Clinical guidelines: Secondary prevention of gastric cancer. Nat Rev Gastroenterol Hepatol. 2012; 9: 128-129.
4. Graham DY. Helicobacter pylori update: gastric cancer, reliable therapy, and possible benefits. Gastroenterology. 2015; 148: 719-31.e3.
5. Plummer M, Franceschi S, Vignat J, Forman D, de Martel C. Global burden of gastric cancer attributable to Helicobacter pylori. Int J Cancer. 2015; 136: 487-490.
6. Uemura N, Okamoto S, Yamamoto S, Matsumura N, Yamaguchi S, Yamakido M, Taniyama K, Sasaki N, Schlemper RJ. Helicobacter pylori infection and the development of gastric cancer. NEJM. 2001; 345: 784-789.
7. Sugano K, Tack J, Kuipers EJ, Graham DY, El-Omar EM, Miura S, Haruma K, Asaka M, Uemura N, Malfertheiner P, faculty members of Kyoto Global Consensus Conference. Kyoto global consensus report on Helicobacter pylori gastritis. Gut. 2015; 64:1353-1367.
8. Correa P. Gastric cancer: overview. Gastroenterol Clin North Am. 2013; 42: 211-217.
9. Rugge M, Capelle LG, Cappellesso R, Nitti D, Kuipers EJ. Precancerous lesions in the stomach: from biology to clinical patient management. Best Pract Res Clin Gastroenterol. 2013; 27: 205-223.
10. Genta RM, Rugge M. Atrophic Gastritis in the Arctic. Clin Gastroenterol Hepatol. 2015; 10: 1601-1603.
11. Rugge M, Pennelli G, Pilozzi E, Fassan M, Ingravallo G, Russo VM, Di Mario F, Gruppo Italiano Patologi Apparato Digerente (GIPAD), Societa Italiana di Anatomia Patologica e Citopatologia Diagnostica/International Academy of Pathology, Italian division (SIAPEC/IAP). Gastritis: the histology report. Dig Liver Dis. 2011; 43 Suppl 4: S373-84.
12. Nordenstedt H, Graham DY, Kramer JR, Rugge M, Verstovsek G, Fitzgerald S, Alsarraj A, Shaib Y, Velez ME, Abraham N, Anand B, Cole R, El-Serag HB. Helicobacter pylori-negative gastritis: prevalence and risk factors. Am J Gastroenterol. 2013; 108: 65-71.
13. Rugge M, Fassan M, Pizzi M, Zorzetto V, Maddalo G, Realdon S, De Bernard M, Betterle C, Cappellesso R, Pennelli G, de Boni M, Farinati F. Autoimmune gastritis: histology phenotype and OLGA staging. Aliment Pharmacol Ther. 2012; 35: 1460-1466.
14. Cancer Genome Atlas Research Network. Comprehensive molecular characterization of gastric adenocarcinoma. Nature. 2014; 513: 202-209.
15. Fassan M, Baffa R, Kiss A. Advanced precancerous lesions within the GI tract: the molecular background. Best Pract Res Clin Gastroenterol. 2013; 27: 159-169.
16. Bornschein J, Leja M, Kupcinskas J, Link A, Weaver J, Rugge M, Malfertheiner P. Molecular diagnostics in gastric cancer. Front Biosci (Landmark edition). 2014; 19: 312-338.
17. D'Angelo E, Vicentini C, Agostini M, Kiss A, Baffa R, Scarpa A, Fassan M. MicroRNAs as tools and effectors for patient treatment in gastrointestinal carcinogenesis. Curr Drug Targets. 2015; 16: 383-392.
18. Fassan M, Croce CM, Rugge M. miRNAs in precancerous lesions of the gastrointestinal tract. World J Gastroenterol. 2011; 17: 5231-5239.
19. Petrocca F, Visone R, Onelli MR, Shah MH, Nicoloso MS, de Martino I, Iliopoulos D, Pilozzi E, Liu CG, Negrini M, Cavazzini L, Volinia S, Alder H, et al. E2F1-regulated microRNAs impair TGFbeta-dependent cell-cycle arrest and apoptosis in gastric cancer. Cancer Cell. 2008; 13: 272-286.
20. Matsushima K, Isomoto H, Inoue N, Nakayama T, Hayashi T, Nakayama M, Nakao K, Hirayama T, Kohno S. MicroRNA signatures in Helicobacter pylori-infected gastric mucosa. Int J Cancer. 2011; 128: 361-370.
21. Hayashi Y, Tsujii M, Wang J, Kondo J, Akasaka T, Jin Y, Li W, Nakamura T, Nishida T, Iijima H, Tsuji S, Kawano S, Hayashi N, et al. CagA mediates epigenetic regulation to attenuate let-7 expression in Helicobacter pylori-related carcinogenesis. Gut. 2013; 62: 1536-1546.
22. Liu WJ, Xu Q, Sun LP, Dong QG, He CY, Yuan Y. Expression of serum let-7c, let-7i, and let-7f microRNA with its target gene, pepsinogen C, in gastric cancer and precancerous disease. Tumour Biol. 2015; 36: 3337-3343.
23. Motoyama K, Inoue H, Nakamura Y, Uetake H, Sugihara K, Mori M. Clinical significance of high mobility group A2 in human gastric cancer and its relationship to let-7 microRNA family. Clin Cancer Res. 2008; 14: 2334-2340.
24. Fassan M, Volinia S, Palatini J, Pizzi M, Baffa R, De Bernard M, Battaglia G, Parente P, Croce CM, Zaninotto G, Ancona E, Rugge M. MicroRNA expression profiling in human Barrett's carcinogenesis. Int J Cancer. 2011; 129: 1661-1670.
25. Neumann WL, Coss E, Rugge M, Genta RM. Autoimmune atrophic gastritis--pathogenesis, pathology and management. Nat Rev Gastroenterol Hepatol. 2013; 10: 529-541.
26. Reinhart BJ, Slack FJ, Basson M, Pasquinelli AE, Bettinger JC, Rougvie AE, Horvitz HR, Ruvkun G. The 21-nucleotide let-7 RNA regulates developmental timing in Caenorhabditis elegans. Nature. 2000; 403: 901-906.
27. Di Leva G, Garofalo M, Croce CM. MicroRNAs in cancer. Annu Rev Pathol. 2014; 9: 287-314.
28. Fassan M, Baffa R, Kiss A, Zaninotto G, Rugge M. MicroRNA dysregulation in esophageal neoplasia: the biological rationale for novel therapeutic options. Curr Pharm Des. 2013; 19: 1236-1241.
29. Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M, Croce CM. Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci U S A. 2004; 101: 2999-3004.
30. Roush S, Slack FJ. The let-7 family of microRNAs. Trends Cell Biol. 2008; 18: 505-516.
31. Takamizawa J, Konishi H, Yanagisawa K, Tomida S, Osada H, Endoh H, Harano T, Yatabe Y, Nagino M, Nimura Y, Mitsudomi T, Takahashi T. Reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res. 2004; 64: 3753-3756.
32. Johnson SM, Grosshans H, Shingara J, Byrom M, Jarvis R, Cheng A, Labourier E, Reinert KL, Brown D, Slack FJ. RAS is regulated by the let-7 microRNA family. Cell. 2005; 120: 635-647.
33. Yanaihara N, Caplen N, Bowman E, Seike M, Kumamoto K, Yi M, Stephens RM, Okamoto A, Yokota J, Tanaka T, Calin GA, Liu CG, Croce CM, et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell. 2006; 9: 189-198.
34. Kumar MS, Armenteros-Monterroso E, East P, Chakravorty P, Matthews N, Winslow MM, Downward J. HMGA2 functions as a competing endogenous RNA to promote lung cancer progression. Nature. 2014; 505: 212-217.
35. Zhu X, Wu L, Yao J, Jiang H, Wang Q, Yang Z, Wu F. MicroRNA let-7c Inhibits Cell Proliferation and Induces Cell Cycle Arrest by Targeting CDC25A in Human Hepatocellular Carcinoma. PloS one. 2015; 10: e0124266.
36. Nomura S, Baxter T, Yamaguchi H, Leys C, Vartapetian AB, Fox JG, Lee JR, Wang TC, Goldenring JR. Spasmolytic polypeptide expressing metaplasia to preneoplasia in H. felis-infected mice. Gastroenterology. 2004; 127: 582-594.
37. Weis VG, Sousa JF, LaFleur BJ, Nam KT, Weis JA, Finke PE, Ameen NA, Fox JG, Goldenring JR. Heterogeneity in mouse spasmolytic polypeptide-expressing metaplasia lineages identifies markers of metaplastic progression. Gut. 2013; 62: 1270-1279.
38. Petersen CP, Weis VG, Nam KT, Sousa JF, Fingleton B, Goldenring JR. Macrophages promote progression of spasmolytic polypeptide-expressing metaplasia after acute loss of parietal cells. Gastroenterology. 2014; 146: 1727-38.e8.
39. Lee J, Ha S, Jung CK, Lee HH. High-mobility-group A2 overexpression provokes a poor prognosis of gastric cancer through the epithelial-mesenchymal transition. Int J Oncol. 2015; 46: 2431-2438.
40. Rugge M, Capelle LG, Fassan M. Individual risk stratification of gastric cancer: evolving concepts and their impact on clinical practice. Best Pract Res Clin Gastroenterol. 2014; 28: 1043-1053.
41. Rugge M, Correa P, Dixon MF, Fiocca R, Hattori T, Lechago J, Leandro G, Price AB, Sipponen P, Solcia E, Watanabe H, Genta RM. Gastric mucosal atrophy: interobserver consistency using new criteria for classification and grading. Aliment Pharm Ther. 2002; 16: 1249-1259.
42. Rugge M, Correa P, Dixon MF, Hattori T, Leandro G, Lewin K, Riddell RH, Sipponen P, Watanabe H. Gastric dysplasia: the Padova international classification. Am J Surg Pathol. 2000; 24: 167-176.
43. Rugge M, Genta RM. Staging and grading of chronic gastritis. Hum Pathol. 2005; 36: 228-233.
44. Fassan M, Pizzi M, Realdon S, Balistreri M, Guzzardo V, Zagonel V, Castoro C, Mastracci L, Farinati F, Nitti D, Zaninotto G, Rugge M. The HER2-miR125a5p/miR125b loop in gastric and esophageal carcinogenesis. Hum Pathol. 2013; 44: 1804-1810.
45. Perilli L, Vicentini C, Agostini M, Pizzini S, Pizzi M, D'Angelo E, Bortoluzzi S, Mandruzzato S, Mammano E, Rugge M, Nitti D, Scarpa A, Fassan M, et al. Circulating miR-182 is a biomarker of colorectal adenocarcinoma progression. Oncotarget. 2014; 5: 6611-6619. doi: 10.18632/oncotarget.2245.